1 CARACTERIZACIÓN E IDENTIFICACIÓN MOLECULAR DE HONGOS DE SUELO AISLADOS DE LOS
PÁRAMOS DE GUASCA Y CRUZ VERDE, CUNDINAMARCA - COLOMBIA
NATALIA GARZON GRAJALES
PONTIFICIA UNIVERSIDAD JAVERIANA FACULTAD DE CIENCIAS
PROGRAMA PREGADO DEPARTAMENTO QUIMICA (GIFUJ)
2 CARACTERIZACIÓN E IDENTIFICACIÓN MOLECULAR DE HONGOS DE SUELO AISLADOS DE LOS
PÁRAMOS DE GUASCA Y CRUZ VERDE, CUNDINAMARCA – COLOMBIA
NATALIA GARZON GRAJALES
TRABAJO DE GRADO
Presentado como requisito parcial para optar por el título
MICROBIOLOGA INDUSTRIAL
Grupo de investigación en fitoquímica Universidad Javeriana (GIFUJ) – Instituto de Errores Innatos del Metabolismo (IEIM)
Director: Jorge Robles Camargo PhD
3 BOGOTA, DC.
CARACTERIZACIÓN E IDENTIFICACIÓN MOLECULAR DE HONGOS DE SUELO AISLADOS DE LOS PÁRAMOS DE GUASCA Y CRUZ VERDE, CUNDINAMARCA – COLOMBIA
NATALIA GARZON GRAJALES
APROBADO
___________________________ ___________________________
Ingrid Schuler PhD Janeth Arias M.SC., M.Ed Bióloga Bacterióloga
4 NOTA DE ACEPTACION
_________________________________
_________________________________
_________________________________
_________________________________
_______________________________
FIRMA DIRECTOR DE TESIS
______________________________
5 NOTA DE ADVERTENCIA
6 ABREVIATURAS
β-tubulina Beta- tubulina
g/L Gramo/litro
GOX Glucoxidasa
°C Grados Celcius
g Gramos
mL Mililitro
mm milímetro
mM mili Molar
μm Micrómetro
μL Microlitro
rpm Revoluciones por minuto
ng nanogramos
nm Nanómetros
NaCl Cloruro de sodio
min Minutos
pb Pares de bases
PCR Reacción en cadena de la
Polimerasa
PDA Papa Dextrosa Agar
%p/v % peso/volumen
psi Libras de presión
rRNA ARN ribosomal
7 Contenido
1. INTRODUCCIÓN……….……….…… 12
2. JUSTIFICACIÓN Y PLANTEAMIENTO DEL PROBLEMA……….………13
3. MARCO TEORICO……….………14
4. OBJETIVO 4.1. GENERAL………..…18
4.2. ESPECIFICO………..………18
5. METODOLOGÍA 5.1. Obtención de la cepa……….……….19
5.2. Elaboración de Cultivos Monospóricos………19
5.3. Elaboración de Micro- cultivos……….………20
5.4. Descripción Macroscópica y Microscópica……….………….……….……20
5.5. Extracción de ADN genómico……….………..20
6. RESULTADOS Y DISCUSIÓN 6.1. Cultivos Monospóricos……….……22
6.2. Extracción de ADN genómico y cuantificación……….…….25
6.3. Amplificación por PCR……….….26
6.4. Árboles Filogenéticos………30
6.5. Potencial Industrial……….33
7. CONCLUSIONES Y RECOMENDACIONES………35
8 INDICE DE TABLAS
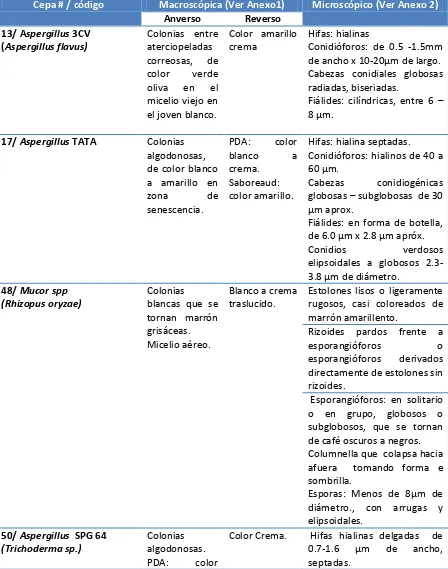
TABLA 1 Descripción macroscópica y microscópica de los cultivos monospóricos en Agar PDA y Saboreaud.
TABLA 2 Contenido ADN obtenido en la extracción en ng/μL con la absorbancia.
9 INDICE DE FIGURAS
Fig.1 Amplificación de las regiones ITS y del gen β-tubulina. ,(a) Bt2a-Bt2b- (b) ITS1-ITS4; (1)Aspergillus terreus, (2)Aspergillus oryzae, (3)Aspergillus CV7, (4)Aspergillus CV29, (5)Aspergillus CV3, (6)Penicillium sp.(61) , (10)Mucor circinelloides, (11)Aspergillus niger con bandas entre 300-550 pb.
Fig.2 Amplificación cepas AspergillusSPG 64 y Rhizopus oryzaecon β-tubulina 2a/b.
Fig.3 Árbol Filogenético de las cepas deducido de un análisis de Neighbour- Joining de una secuencia parcial del gen β-tubulina. Las cepas estudiadas en el presente trabajo están señaladas con el punto rojo.
10 ANEXOS
ANEXO 1. MACROSCOPÍA DE CADA HONGO EN PDA, SABOREAUD, EXTRACTO DE MALTA, CZAPECK O AVENA.
ANEXO 2. MICROSCOPIA DE LAS CEPAS.
ANEXO 3. MEDIOS DE CULTIVO
11 RESUMEN
12 1. INTRODUCCIÓN
Por años, los hongos han sido estudiados por su variada aplicación a nivel industrial para la obtención de productos alimenticios, cosméticos o farmacéuticos; o para evitar sus efectos perjudiciales y de esta forma contrarrestarlos (1).
Los hongos filamentosos son microorganismos implicados en procesos biotecnológicos debido a su alto potencial en el desarrollo de productos industriales de origen natural, ya que estos producen gran cantidad de compuestos que pueden ser aislados y utilizados en diferentes campos a nivel industrial tales como ambiental, alimenticio, farmacéutico, entre otros(2,3).
Para la aplicación y el conocimiento del potencial de estos hongos filamentosos a nivel industrial es necesaria la completa caracterización de las cepas con las que se desea trabajar, tanto a nivel microscópico y macroscópico, como a nivel molecular, para así determinar el potencial del hongo y de los compuestos que produce (2,3).
Debido al uso de estos compuestos y su alto potencial biotecnológico, es necesaria la búsqueda de mejoras en los procesos de aislamiento y producción de estos mismos por medio de diferentes técnicas, para las cuales se hace necesaria la identificación de dichos microorganismos, recurriendo a métodos moleculares con la finalidad de identificar genes potenciales cuya función esté relacionada con la producción de metabolitos secundarios con potencial industrial, permitiendo el mejoramiento de los procesos industriales en los que el microorganismo se encuentra implicado (2, 3, 4,5).
Hongos filamentosos pertenecientes a los géneros como Aspergillus sp. y Mucor sp. son conocidos en diferentes campos a nivel industrial. Por ejemplo en el campo farmacéutico Mucor circinelloides es empleado en la producción de ácido kojico, que se utiliza como principio activo de cremas desmanchadoras del rostro, en el área industrial se encuentran aplicaciones en la producción de biodiesel, isoprenoides, vitaminas, etc. (5, 6, 7,8).
13 2. JUSTIFICACIÓN Y PLANTEAMIENTO DEL PROBLEMA.
El grupo de Investigación en Fitoquímica Universidad Javeriana (GIFUJ) ha realizado la identificación y la caracterización microscópica y macroscópica de hongos pertenecientes a los géneros como Aspergillus sp. y Mucor sp. cuyos resultados fueron obtenidos por medio del uso de claves taxonómicas, encontrándose que de 33 a 86 cepas pertenecen a estos géneros.
Para asegurar el género y especie de un hongo es necesario el uso de varios métodos para su identificación. Debido a la gran variedad fenotípica dentro de un mismo género el empleo de métodos moleculares, sumado a la clasificación morfológica, representa una importante alternativa para la adecuada clasificación del hongo. Otros métodos importantes consideran la composición química de la membrana celular; la cual ejerce una importante función en su clasificación taxonómica. Por ello es necesario combinar los métodos para asegurar la identidad de los hongos y adicionalmente emplear y garantizar la conservación de estos sin ningún tipo de cambios en su morfología o fisiología, lo cual es de gran importancia en el hallazgo de potenciales biotecnológicos que puedan constituir un avance importante en campos de diversas industrias. (9)
14 3. MARCO TEORICO
Estudio de Hongos en Colombia
Colombia es un país conocido por su biodiversidad, Yepes et al. 2005 afirma que el 40% de estudios nacionales enfocan sus trabajos en plantas, 33% en vertebrados, 21% en invertebrados y el 14 % en microorganismos (14). A pesar que los estudios con microorganismos representan el porcentaje más pequeño, es el que hacen énfasis los investigadores que tienen como objetivo principal el enfoque biotecnológico. Para ello los ecosistemas constituyen un papel fundamental en el descubrimiento de nuevas cepas y sus potenciales. En Colombia uno de los ecosistemas de mayor potencial son los páramos, en el que la mayor influencia es dada por condiciones ambientales y principalmente el suelo ya que este es rico en materia orgánica, y donde se dan gran cantidad de interacciones biológicas, en la que están presentes bacterias, hongos, actinomicetos y protozoarios (10).
Estudio de Hongos por el grupo de investigación de fitoquímica Universidad Javeriana (GIFUJ) En el laboratorio de Química Microbiológica del departamento de Química de la PUJ se han llevado a cabo una serie de investigaciones para el aislamiento, caracterización y estudio de metabolitos secundarios de microorganismos aislados de suelos del páramo de Guasca, Cundinamarca. Trabajos de grado de pregrado y maestría, permitieron la caracterización morfológica de 147 cepas (10, 11, 12,13). En estos estudios se logró la caracterización a nivel microscópico y macroscópico determinando por medio de claves taxonómicas los géneros de las cepas estudiadas, obteniendo características microscópicas como esporangios globosos, demateaceos, rectos o curvos de 50 -70 μm de diámetro, columnellas lisas, semiesféricas, hifas hialinas y esporas ovoides, lisas, incoloras individualmente que corresponden a el género Mucor sp. Para algunas de estas cepas se caracterizó su actividad antimicrobiana, encontrándose que los extractos polares poseen actividad inhibitoria sobre como Bacillus subtilis (10, 11, 12,13). A pesar de los importantes logros obtenidos en los últimos años con los microorganismos de este banco, aun es necesaria la caracterización de estos microorganismos desde el punto de vista molecular, lo que serviría como un punto importante de partida para la realización de futuros proyectos que permitan mejorar la producción de metabolitos secundarios con intereses industriales.
Hongos Filamentosos y su Importancia industrial
15 Métodos de Identificación Molecular
Existen diferentes herramientas para la detección, diferenciación e identificación de hongos, entre las que se encuentran métodos fenotípicos, bioquímicos, inmunológicos y moleculares. A pesar de que el método de caracterización fenotípica es el estándar para la identificación de la mayoría de los microorganismos, los métodos moleculares también son una herramienta indispensable en la mayoría de estudios que surgieron como consecuencia de los tediosos procesos de identificación bioquímica y fenotípica (16). Sistemas comerciales como kits basados en el uso de la enzima taq polimerasa se desarrollaron inicialmente para ser usados en aislamientos humanos, de comida o bebidas con el fin de identificar cepas desconocidas; y de allí las pruebas moleculares empezaron a formar parte complementaria o alternativa a las pruebas fenotípicas, determinando una mayor especificidad y sensibilidad de los ensayos (15).
Se conoce que el trabajo de identificación a nivel molecular de hongos filamentosos, constituye una de las tareas más complejas debido a la composición de estos, haciendo de la extracción de ADN uno de los procesos más importantes de los métodos moleculares, en los que se ve involucrada directamente la composición de la membrana celular, constituida por un 10% en proteínas y glicoproteínas; seguida de un 8% de lípidos y iones inorgánicos, tales como calcio, fosforo y magnesio, micro elementos fundamentales en la clasificación taxonómica. (9)
Dentro de estos ensayos moleculares el método de la reacción en cadena de la polimerasa (PCR) es una de las primeras técnicas de la generación molecular cuyo propósito es la ampliación de un fragmento de ADN de la muestra que se somete a estudio. Existen varias modalidades de esta técnica como polimorfismos de longitud de fragmentos de restricción PCR-RFLP, (PCRtr) PCR en tiempo real, RAPD (Randomly Amplified Polymorphic DNA), PMM (Polymorphic Microsatellite Markers) y MLST (Multilocus Sequencing Typing), por ejemplo en el futuro la búsqueda del diseño de un microarray que permita la identificación de patógenos fúngicos presentes en muestras clínica, muestra que los métodos moleculares tienen particular interés en avanzar tecnológicamente para monitorear los microorganismos en un ecosistema natural (16,17).
Secuencias Diana
Con fines de identificación molecular se suele amplificar regiones diana multicopia y sitios altamente conservados ya que hace fácil la amplificación en todos los hongos (18). Las regiones diana multicopia utilizadas generalmente son los genes ribosomales. El ribosoma de células eucariotas tiene dos subunidades de ARN ribosomal (ARNr): 40S y 60S, que están codificados por tres genes: el gen 5.8S, la subunidad pequeña (18S) y por último la subunidad grande (28S), los cuales están separados por regiones espaciadores de transcripción interna (ITS). La subunidad 18S una de las más utilizada en comparaciones taxonómicas de base filogenética (24), así como la región ITS, debido a que estas últimas son intrones variables, ya que acumula mutaciones neutrales a través del tiempo, permitiendo distinguir entre organismos genéticamente relacionados a nivel de especie y subespecie. (19, 20, 21, 22, 23, 24)
16 eucariotas. A pesar que esta proteína es altamente conservada entre eucariotas, la comparación de regiones intrónicas permite diferencias entre individuos de una misma especie. (25)
La secuencia nucleotídica del gen de β-tubulina ha formado parte de análisis filogenéticos donde los polimorfismos en el intrón 3 han sido los más utilizados en estudios. (26) Las regiones ITS no siempre proporcionan una buena resolución para especies filogenéticamente próximas, el análisis de una segunda región, como el gen de β-tubulina, puede ser necesario para asegurar una identificación adecuada. (26,27) Así lo lograron determinar Gonzalez et al. (2007) cuando realizaron un análisis de diversidad genética de una población de Colletotrichum spp. a partir de la comparación en la distribución de subunidades e intrones en secuencias de ARNr (ITS) y β -tubulina, basados en estudios de O’Donnel et al. en los que reportó que hay 3.5 veces más información filogenética en los genes de la β-tubulina que en los genes de ARNr. (28)
Secuenciación y Filogenia
Una vez son obtenidos las secuencias de las regiones amplificadas se procede a hacer una comparación de estas secuencias con las reportadas en las bases internacionales, como GenBank. Esta comparación permite la identificación de las especies fúngicas, incluso puede hasta tipificar cepas de una misma especie. (29, 30,31)
17 4. OBJETIVO
4.1 GENERAL
Caracterizar desde el punto de vista molecular cepas nativas de hongos aislados del páramo de Guasca Colombia, registrados en la colección de microorganismos del laboratorio de química microbiológica del departamento de Química de la PUJ
4.2 ESPECÍFICOS
1. Identificar morfológicamente las cepas nativas de Aspergillus sp y Mucor sp.
2. Identificar molecularmente las especie de las cepas nativas de Aspergillus sp. y Mucor sp. 3. Realizar una revisión de literatura relacionada con la producción de metabolitos secundarios
18 5. METODOLOGÍA
Obtención de las cepas
Las cepas a utilizar fueron previamente aisladas de muestras de suelo tomadas de los páramos de Guasca y Cruz Verde (Cundinamarca) por el (GIFUJ). Los microorganismos fueron identificados por medio de claves taxonómicas, a las cuales se le realizó un cultivo monospórico.
A partir de este banco de microorganismos se seleccionaron los siguiente hongos: 56 Aspergillus CV7, 82 Aspergillus CV29, 81 Aspergillus A-26,98 Aspergillus oryzae, 17 Aspergillus TATA, 13 Aspergillus 3CV, 50 Aspergillus SPG64, 96 Aspergillus niger, 61Mucor circinelloides pf (papel filtro), 61 Mucor circinelloides, 48 Mucor spp. Los microorganismos fueron reactivados a partir de los bancos conservados en suelo, papel filtro y glicerol. En el primer caso, se tomó con una pinza la mayor cantidad de gránulos posibles, los que posteriormente se sembraron en Agar Papa Dextrosa (PDA), e incubando a 25°C durante 7 días. Por su parte, para la reactivación a partir del banco de glicerol, las muestras se descongelaron a temperatura ambiente y luego se resuspendieron en caldo nutritivo, para posteriormente ser incubadas a 25° C durante 7 días. (10, 11, 12,13)
Elaboración de cultivos monospóricos.
La obtención de un cultivo axénico se obtiene gracias a la realización de un cultivo monospórico. Para este fin, se inocularon tubos con medio PDA inclinado, con los microorganismos a ser evaluados, los cuales fueron incubados a 25°C durante 5 días. Pasado el periodo de incubación, se adicionaron 10 mL de solución Tween 80 al 0.1% hasta obtener el desprendimiento de los conidios. El recuento de las esporas se realizó en cámara de Newbauer, para el cual se mezclaron 0.5mL de la suspensión de conidios en Tween 80 al 0.1% con 4.5 mL de agua peptonada al 0.1% en la que se hizo diluciones de 10-1 a 10-7 para obtener una concentración de conidios entre 50 y 100 conidios. Posteriormente se sembró por agotamiento en cajas de Petri con medio de cultivo PDA, las cuales fueron incubadas a 25° C hasta observar el crecimiento de una espora, aproximadamente entre el segundo y tercer día de incubación. Con ayuda de un estereoscopio se realizó la selección de un conidio que posteriormente se transfirió a otra caja de Petri con medio de cultivo PDA, la cual se incubó a 25°C durante 5 días. (10)
La verificación de la pureza se realizó mediante tinción con azul de lactofenol y tinción de Gram. (19)
Elaboración de Microcultivos
19 Descripción macroscópica
Para la descripción macroscópica de los hongos para cada una de las cepas se realizó repiques a partir de los cultivos monospóricos por punción central en los medios de cultivo PDA, Extracto de Malta, Extracto de Malta, Czapeck o Avena según requerimientos de cada hongo. Los microorganismos fueron incubados por 5 días a 25°C, se describió el anverso y reverso de cada cepa según textura, color, diámetro de la colonia y la presencia de pigmentos difusibles al medio. Descripción microscópica
Para la descripción microscópica de los hongos se hizo la observación de los microcultivos con azul de lactofenol, en Microscopio Motic Type 102M en objetivo 40X y 100X siguiendo las claves taxonómicas de Sansom et al y Bernnet et al; determinando el género y la especie presuntiva de cada uno de los hongos estudiados. (33,34)
Extracción de ADN genómico de las cepas
Para la extracción de ADN genómico se inoculó en Caldo Nutritivo con disco de agar cada uno de los hongos, incubando a temperatura ambiente de 5- 8 días en agitación a 120 rpm.
La extracción del ADN genómico se realizó empleando el protocolo descrito por John Weiland (http://www.fgsc.net/fgn44/weiland.html), con modificación. Un gramo de micelio se pulverizó con nitrógeno líquido, el cual se mezcló con 400 µL de buffer de lisis (100mM Tris HCl pH 8,0, 20mM EDTA disódico, 0,5M NaCl, 1% SDS), la emulsión obtenida se transfirió a un tubo de micro centrífuga con 1 mL de fenol/cloroformo (1:1). La mezcla se homogenizó por inversión, se transfirió a tubos de micro centrífuga y se centrifugó a 14000g por 5 min a temperatura ambiente. La fase acuosa se transfirió a tubos nuevos y se adicionaron 0,6 volúmenes de isopropanol, incubando la mezcla por 10 minutos a temperatura ambiente. El precipitado se obtuvo por centrifugación a 14000g por 15 minutos a 4°C. El sobrenadante se descartó y el precipitado de (ADN) se lavó con etanol al 70%, se secó durante 10 min a temperatura ambiente y se reconstituyó en 200 μl en buffer Tris-EDTA-. (2)
La cuantificación del ADN se realizó espectrofotométricamente empleando un Nanodrop 2000(Thermo Fisher Scientific. La presencia y calidad del ADN se analizó mediante electroforesis en gel de agarosa al 1% que fue revelada. (2)
Los cebadores empleados para la identificación molecular de los hongos fueron ITS1
(5’TCCGTAGGTGAACCTGCGG-3’) e ITS4 (5’TCCTCCGCTTATTGATATGC-3’), los cuales
amplifican un fragmento de las ITS, y
ᵦ
ta2a (5’GGTAACCAAATCGGTGCTGCTTTC-3’)20 Para verificar el funcionamiento de cebadores y el tamaño de fragmentos, se tomaron 5 µL de ADN genómico (aprox 1μg) se mezclaron con 2.5 μL de buffer 10X PCR (Corpogen), 0.5 μL dNTPs (10mM), 1.5 μL MgCl (25mM) (Corpogen), 0,25 μL Taq polimerasa (5U/μL, Corpogen), 1 μL de cebadores (10mM) y agua desionizada para un volumen final de 25 μL. La reacción se realizó bajo las siguientes condiciones: 35 ciclos de 1 min a 95 0C, 1 min a 59 0C y 45 seg a 72 0C. Los productos de PCR se evidenciaron en gel de agarosa al 2% y tinción con bromuro de etidio. Una vez confirmado la funcionalidad de los cebadores y la obtención de fragmentos de ADN de los tamaños esperados, se realizó la reacción de amplificación de los fragmentos que se enviaron a secuenciación. Para este fin, se utilizó el estuche Expand Long Template PCR System (Expand Long Template PCR System), siguiendo las recomendaciones del fabricante y bajo las siguientes condiciones de reacción: 40 ciclos de 1 min a 95 0C, 1 min a 58 0C y 30 seg a 680C. Los fragmentos se purificaron a partir de gel de agarosa al 2% empleando el estuche Wizard PCR preps DNA purification system, siguiendo las recomendaciones del fabricante. Los fragmentos purificados se secuenciaron por el Instituto de Genética de la Universidad Nacional de Colombia (20).
Las secuencias obtenidas se evaluaron manualmente y se compararon con la base de datos GenBank empleando la herramienta BLAST para establecer la especie de los microorganismos sometidos al estudio.
Los arboles filogenéticos se construyeron con el programa MEGA5.2 en un análisis de Neighbour-Joining con un test bootstrap de 500 repeticiones.
Revisión Bibliográfica
Para la revisión bibliográfica se emplearon las bases de datos Scopus, springerlink, en artículos relacionados con el tema entre (2006 a 2013) Las palabras claves de búsqueda fueron: Aspergillus sp. Mucor sp. Biotecnología, ADN, genómica, metabolitos secundarios, genética, micología industrial.
6. RESULTADOS Y DISCUSIÓN
1. Cultivos Monospóricos
21 Tabla1. Descripción macroscópica y microscópica de los hongos aislados en Agar PDA y Saboreaud.
Cepa # / código Macroscópica (Ver Anexo1) Microscópico (Ver Anexo 2)
Anverso Reverso
13/ Aspergillus 3CV (Aspergillus flavus)
Colonias entre aterciopeladas correosas, de color verde oliva en el micelio viejo en el joven blanco.
Color amarillo crema
Hifas: hialinas
Conidióforos: de 0.5 -1.5mm de ancho x 10-20μm de largo. Cabezas conidiales globosas radiadas, biseriadas.
Fiálides: cilíndricas, entre 6 – 8 μm.
17/ Aspergillus TATA Colonias
algodonosas, de color blanco a amarillo en zona de senescencia. PDA: color blanco a crema. Saboreaud: color amarillo.
Hifas: hialina septadas. Conidióforos: hialinos de 40 a 60 μm.
Cabezas conidiogénicas globosas – subglobosas de 30 μm aprox.
Fiálides: en forma de botella, de 6.0 μm x 2.8 μm apróx.
Conidios verdosos elipsoidales a globosos
2.3-3.8 μm de diámetro. 48/ Mucor spp
(Rhizopus oryzae)
Colonias
blancas que se tornan marrón grisáceas. Micelio aéreo.
Blanco a crema traslucido.
Estolones lisos o ligeramente rugosos, casi coloreados de marrón amarillento.
Rizoides pardos frente a esporangióforos o esporangióforos derivados directamente de estolones sin rizoides.
Esporangióforos: en solitario o en grupo, globosos o subglobosos, que se tornan de café oscuros a negros. Columnella que colapsa hacia afuera tomando forma e sombrilla.
Esporas: Menos de 8μm de diámetro., con arrugas y elipsoidales.
50/ Aspergillus SPG 64 (Trichoderma sp.)
Colonias algodonosas.
PDA: color
22 verde oscuro
zona senescencia, blanco micelio joven. Saboreaud: Color amarillo zona senescencia a blanco.
Conidios elipsoidales de 4.0 – 4.9 μm.
56/ Aspergillus CV7 Penicillium spp. Colonias pulverulentas, verdes con bordes blancos y Pigmentos difusibles al medio (PDM) amarillo.
Color Amarillo. Hifas: hialinas delgadas. Conidióforos hialinos de 53.4 μm de largo apróx x 1.5 μm de ancho.
Métula cuatriverticialiado de 8.2μm de largo apróx.
Fiálides: cilíndricas, 6.5μm – 2.5μm apróx.
Conidios globosos: de 3.0-4.0 μm apróx.
61/ Mucor circinelloides Colonias con presencia de micelio aéreo, grisáceas a verdes.
Color blanco a crema.
Hifas: Cenocíticas, demateaceas.
Esporangios: altos y cortos, llenos con gotas en partes jóvenes. Blanquecinos a amarillentos en inicio y en madurez pardos.
Columnella ovoide elipsoidal en esporangios largos, y globosa en cortos.
61/Penicillium sp. (Penicillium brevicompactum) Colonias pulverulentas, ceribriformes, color verde militar a blanco. Colonias diámetro de 2 a 3 cm.
Color amarillo o crema.
Hifas: hialinas sin septos. Métula: de 5.5μm largo. Fiálides: de 9.3μm x 3.0μm
Conidios: globosos a
subglobosos, verdosos de 2.9μm de diámetro.
81/ Aspergillus A-26 (Aspergillus terreus.)
Colonias entre amarillo y marrón
aterciopeladas, con PDM de color amarillo. Saboreaud: cerebriforme.
Color amarillo crema.
Hifas: hialinas de μm de grosor.
Conidióforos: hialinos, cortos, verdes.
Cabezas conidiales: Globosas biseriadas.
23 82/Aspergillus CV29
(Aspergillus fumigatus)
Colonias aterciopeladas al inicio del crecimiento, color blanco a verde oscuro.
Color: Amarillo crema.
Hifas: hialinas, micelio cenocítico.
Cabezas conidiales radiales a globosas, biseriadas.
Conidióforos con vesículas ampliamente claviformes. 96/Aspergillus niger
(Aspergillus tubingensis)
Colonias correosas de
color negro
Hifas: hialinas a marrones, sin septos. Conidióforos hialinos, erguidos. De más de 37.6 μm Métula cilíndricas de 6.5μm apróx de largo.
Fiálides: cilíndricas de 6.1μm x 2.9μm de ancho.
Conidios: Globosos oscuros de 2.8 a 3.0 μm.
98/ Aspergillus oryzae (Aspergillus oryzae)
Color flocosa raramente aterciopelada, de color verde oliva a blanco.
Color crema amarilloso.
Hifas: hialinas
Conidióforos: hialinos, largos. Cabezas conidiales globosas a
radiadas. A menudo
biseriadas.
Vesículas: subglobosas de 67.8 μm aprox.
Fiálides cilíndricas, en forma de botella de 8.6μm x 2.7μm. Conidios: hialinos de 3.2 μm apróx.
Adicionalmente se realizaron siembras en medio de cultivo Czapeck, Extracto de Malta y Avena (para mucorales) para una descripción morfológica más detallada (Ver Anexo 1), colonias de aterciopeladas y pulverulentas a algodonosas de colores verdosos a amarillo para algunas y otras blancas y algodonosas en cepas codificadas como Aspergillus sp, en caso de los mucorales con micelio aero de color grisáceo a marrón. Se encontró que las cepas codificadas bajo el género de Aspergillus sp. pertenecen a géneros como Penicilliun sp y Trichoderma sp; en el caso de mucorales se encontró que la cepa 48 Mucor spp. es Rhizopus sp. Los dos medios para la realización del cultivo monospórico fueron PDA y Saboreaud por ser medios comunes que dan los nutrientes mínimos para el crecimiento de cualquier microorganismo por ello se suplementaron con cloranfenicol para evitar el crecimiento de bacterias. Haber incubado un cultivo a partir de una sola espora garantiza la pureza de los cultivos, manteniendo un mismo genotipo y fenotipo manifestados en una morfología única. (34) Al observar los hongos microscópicamente después de un periodo de 5 días se hizo posible la observación de todas las estructuras características de cada hongo como hifas, conidióforos, esporangióforos, columnellas, métulas, fiálides etc.
24 Investigación de Fitoquímica de la Universidad Javeriana (GIFUJ)(10, 11, 13, 14); mientras que las cepas Mucor spp, Aspergillus niger, Aspergillus SPG 64, Aspergillus CV7 y Mucor circinelloides (papel filtro) mostraron resultados diferentes a las cepas que se encuentran documentadas en la colección de GIFUJ, las cepas con género y especie presuntiva identificadas son: Rhizopus oryzae, Aspergillus niger, Trichoderma sp, Penicillium sp y Penicillium brevicompactum. Todo esto basado en las claves taxonómicas de Samson et al. (2011) y Barnett et al. (1999) con lo que se puede afirmar que no es sencillo solo por este método identificar hasta nivel de especie un hongo. (33,34)
2.
Extracción de ADN genómico y cuantificación
1.
Electroforesis
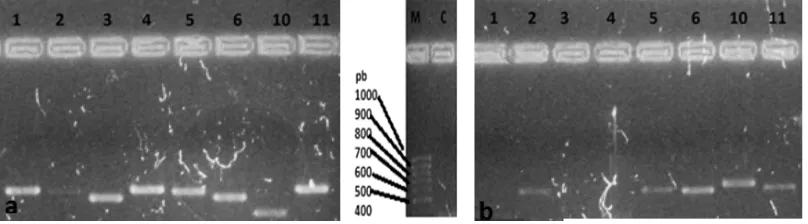
Para el análisis molecular las cepas estudiadas fueron enumeradas de la siguiente forma:
1. Aspergillus terreus 2. Aspergillus oryzae 3. Aspergillus CV7 4. Aspergillus CV29 5. Aspergillus CV3 6. Mucor circinelloides pf 7. Aspergillus SPG 64 8. Aspergillus TATA 9. Mucor spp.
10. Mucor circinelloides 11. Aspergillus niger
[image:24.612.223.391.584.720.2]Para todos los hongos se obtuvo ADN genómico con la pureza adecuada y concentración suficiente para realizar el análisis molecular de las cepas (Tabla 2 y Anexo 4)
Tabla 2. Contenido obtenido en la extracción de ADN en ng/μL vs. absorbancia.
Cepa
ng/μL 260/280nm 1 1002.87 2.012 390.56 1.97
3 451.80 2.06
4 416.34 1.85
5 242.43 2.02
6 526.56 2.12
7 3023.15 2.13
25
9 296.02 2.14
10 1275.46 2.16 11 372.34 1.77
2.
Amplificación por PCR
En el ensayo de PCR realizado para comprobar el funcionamiento de los cebadores, solo funcionaron los ITS1-ITS4 y Bt2a-Bt2b. La Figura 1 muestra los resultados de amplificación con los cebadores ITS1-ITS4 y Bt2a-Bt2b, con los cebadores ITS y β-tubulina para las cepas Aspergillus terreus, Aspergillus oryzae, Aspergillus CV7, Aspergillus CV29, Aspergillus CV3, Penicillium sp., Mucor circinelloides, y Aspergillus niger, en los que se observaron bandas entre 300 y 600 pb, mientras que la Figura 3 muestra el resultado de amplificación para las cepas Aspergillus SPG 64 y Mucor spp con los cebadores de β-tubulina.
Fig.1 Amplificación de las regiones ITS y del gen β-tubulina. (a) Bt2a-Bt2b- (b) ITS1-ITS4; (1)Aspergillus terreus, (2)Aspergillus oryzae, (3)Aspergillus CV7,(4)Aspergillus CV29,(5)Aspergillus CV3,(6)Penicillium sp.(61) ,(10)Mucor circinelloides,(11)Aspergillus niger con bandas entre 300-550 pb. C: control, M: marcador.
[image:25.612.122.526.407.519.2]26 Fig.3 Amplificación cepas AspergillusSPG 64 y Mucor spp.con los cebadores para β-tubulina
2a/b.
En total ocho cepas fueron amplificadas con los cebadores para las regiones ITS y β-tubulina: Aspergillus terreus, Aspergillus oryzae, Aspergillus CV7, Aspergillus CV29, Aspergillus CV3, Mucor circinelloides pf, Mucor circinelloides y Aspergillus niger. Estudios realizados por Jung, et al (2009) mostraron que para especies de Aspergillus sp. amplificadas con β-tubulina se obtuvieron fragmentos de 402 a 510 pb, en donde Aspergillus oryzae/flavus presentaron un tamaño de 497 pb que es similar el de la cepa de este estudio, para la cual se obtuvo un fragmento de aproximadamente 500 pb. Adicionalmente Aspergillus tubingensis en el estudio de Jung et al (2009)presentó fragmento de 506pb siendo menor de la cepa codificada como Aspergillus niger que dio 580pb, en donde el tamaño si pudo variar indicando que no son la misma cepa pero si la misma especie. En cuanto al tamaño de las banda obtenidas para la región ITS se observó un rango de 400 a 630 pb aproximadamente, que concuerda con la cepa de Mucor circinelloides descrita por los mismo autores, con un tamaño de 599 pb siendo de menor tamaño, que la obtenida en el presente estudio Mucor circinelloides. (3)
Solo las cepas de Aspergillus SPG 64 y Mucor spp.Amplificaron con β-tubulina obteniéndose para Aspergillus SPG 64 una banda de 400 pb y para Mucor spp. una banda de 390 pb valores que se encuentran dentro del rango del tamaño de los fragmentos que se obtienen con este gen de la β -tubulina.
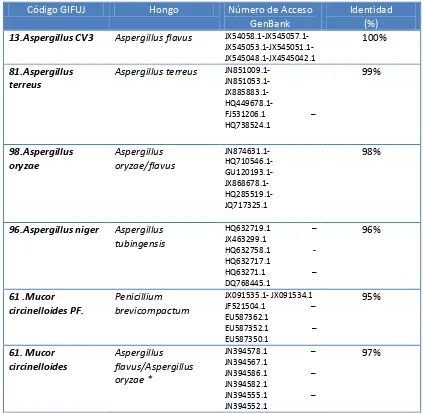
27 identificación de los hongos. Del total de muestras procesadas para secuenciación, sólo se obtuvo un resultado positivo para las seis cepas presentadas en la Tabla3.
Tabla 3. Identificaciones moleculares de hongos de la colección de microorganismos de GIFUJ.
Código GIFUJ Hongo Número de Acceso
GenBank
Identidad (%) 13.Aspergillus CV3 Aspergillus flavus
JX54058.1-JX545057.1- JX545053.1-JX545051.1-JX545048.1-JX4545042.1
100%
81.Aspergillus terreus
Aspergillus terreus JN851009.1- JN851053.1- JX885883.1- HQ449678.1-
FJ531206.1 – HQ738524.1 99% 98.Aspergillus oryzae Aspergillus oryzae/flavus JN874631.1- HQ710546.1- GU120193.1- JX868678.1- HQ285519.1- JQ717325.1 98%
96.Aspergillus niger Aspergillus tubingensis
HQ632719.1 – JX463299.1
HQ632758.1 - HQ632717.1
HQ63271.1 –
DQ768445.1 96% 61 .Mucor circinelloides PF. Penicillium brevicompactum JX091535.1- JX091534.1 JF521504.1 – EU587362.1
EU587352.1 – EU587350.1 95% 61. Mucor circinelloides Aspergillus flavus/Aspergillus oryzae *
JN394578.1 – JN394567.1
JN394586.1 – JN394582.1
JN394555.1 – JN394552.1
97%
28 secuencias provenientes de hongos de otros géneros. La primera mostró una identidad del 96% con Aspergillus tubingensis cepa IHEM21971 beta-tubulin gene (GenBank No. HQ632719.1), mientras que Mucor circinelloides tomada de papel filtro resultó idéntica a Penicillium brevicompactum cepa CV2475 beta-tubulin gene (GenBank No. JX091535.1), y Mucor circinelloides tomada de glicerol al 10% tuvo identidad del 98% con Aspergillus flavus aislamiento CA35 beta-tubulin gene (GenBank No. JN394578.1). Mientras en los dos primeros casos los resultados concuerdan con la morfología obtenida de la reconstitución de la cepa (ver Anexo1). Para el segundo aislamiento de la cepa de Mucor circinelloides no se observa una concordancia entre los resultados de identificación morfológica y molecular, por lo que se recomienda confirmar estos resultados mediante la re secuenciación del fragmento y el uso de una segunda pareja de cebadores.
En cuanto a las identificaciones de Aspergillus oryzae y Aspergillus flavus, es notorio que si solo se hubiese hecho una identificación molecular no habría sido posible realizar la adecuada identificación de los hongos, debido a que estos hongos pertenecen a la sección ‘Flavi’ y donde Aspergillus oryzae desciende de Aspergillus flavus. Sin embargo Thomas Cleveland et al. (2006) y
Changet al. (2009) en sus estudios hacen notable la semejanza morfológica entre estas especies, es allí donde se hizo importante para este estudio la combinación de estos métodos para distinguir morfológicamente Aspergillus flavus de Aspergillus oryzae.(36) En estudios realizados por Reis et al (2012), la identificación polifásica de la sección "Flavi"en Aspergillus la muestran ahora como la mejor alternativa para hacer de la identificación de hongos más confiable, puesto que allí no solo se determina por vía molecular y morfológica sino que incluye la producción de astrolitos y resistencia fúngica al calor. En el caso de Mucor circinelloides es evidente que por sus características estructurales encontradas en microscopía y macroscopía, que la parte morfológica diverge de la parte molecular, en cuyo caso la identificación molecular ayudaría a establecer un mejor perfil para su identificación. (37, 38, 39,40)
Lo mismo ocurre en cuanto a la sección "nigri" común para Aspergillus niger y Aspergillus tubingensis, Silva et al. (2011) afirmó que estas especies son difícilmente diferenciales por métodos morfológicos, lo que explicaría la razón de que con el uso de las claves taxonómicas en este estudio se identificará como Aspergillus niger. Samson et al. (2011)afirma que en ocasiones se puede visualizar la formación de esclerocios de color blanco a rosa pero no siempre ocurre así, por lo cual la identificación molecular tiene importancia para confirmar la parte morfológica. (33, 38,41)
3.
Arboles filogenéticos
30 Fig.4 Árbol filogenético de las cepas deducidas de un análisis Neigbour-Joining de una secuencia génica
parcial de ITS. Las cepas estudiadas en el presente trabajo están señaladas con el punto rojo.
31 Para un completo análisis de las secuencias obtenidas es importante la construcción de los árboles filogenéticos, con ellos es posible la observación de la relación evolutiva entre las cepas estudiadas y otras anteriormente confirmadas por estudios de alta confiabilidad. (40)
En la Figura 4 sería posible inferir que las cepas de Aspergillus flavus GIFPUJ13 y Mucor circinelloides GIFPUJ61 son muy cercanas; lo que sugiere una incongruencia entre los resultados obtenidos morfológicamente con los obtenidos molecularmente, pues Mucor circinelloides debería encontrarse dentro del grupo de los Zygomicetes y no Aspergillus sp. Estos resultados muestran la importancia de realizar la identificación de los hongos por más de un método, y para el caso de la identificación molecular usar más de una pareja de cebadores. Schuster et al. (2002) en su revisión relacionada con Aspergillus niger y otras especies de Aspergillus mostraron una relación indistinguible cuando se usó la amplificación con ITS, debido a la dificultad para la construcción de árboles filogenéticos por ser una región altamente conservada, gen común dentro de todos los géneros pertenecientes al reino Hongos.(3,43,44)
Para la identificación molecular es recomendado el uso de varios cebadores entre los cuales se puedan comparar resultados, analizando los microorganismos identificados, como en la identificación de Aspergillus flavus que usando los dos genes muestra mejores resultados con la región amplificada con β-tubulina, ubicado dentro del árbol filogenético en el clado de Aspergillus flavus y Aspergillus oryzae que comparten la sección "flavi" (Figura 3), mientras que en el árbol realizado con la amplificación de regiones ITS queda ubicado fuera de este clado. Las regiones ITS son conocidas por su uso universal y por ello se utilizan en estudios donde hay diversidad de géneros de hongos (45). Con base en la afirmación anterior, se recomienda la identificación de las cepas que amplificaron con un solo par de cebadores resecuenciadas por otro par de cebadores, es decir si Aspergillus terreus se logró identificar con ITS se recomienda resecuenciarlo con el gen de β-tubulina. (45, 46,47)
32 4.
Potencial Industrial
Tabla 4. Genes involucrados en la producción de los metabolitos tanto primarios con potenciales implicaciones industriales.
Código cepa Microorganismos
Relacionados
Genes Producto Aplicación Industrial
(13) Aspergillus flavus Aspergillus oryzae, Aspergillus fumigatus 1. aflC(pksA)Pol icetido sintasa 2. sidD(sid2) 3. gliP(A.fumig atus con A.flavus)
1. policétidos (ácidos grasos)
2. Peptido sintasas no ribosomales
3. Biosíntesis de nano partículas de oro
1,2,3. Productos naturales, antibióticos, antifungicos,
antiparasíticos etc. (48) 4. Química verde,
polímeros ecológicos, nanotecnología.(49)
(81) Aspergillus terreus
Saccharomyces cerevisiae
1. Gen suc 1/Gen sir 1
2. CAD gene
1. Invertasas 2. Ácido itacónico
1. Industrial de
alimentos confites, edulcorantes.(50) 2. Polímeros
biodegradables
(98) Aspergillus oryzae
Aspergillus flavus Rhizopus oryzae
1. Gen suc 1/Gen sir
2. ProRol
1.Invertasas 2.Proteínas hererologas
1.Industrial de alimentos confites,
edulcorantes.(50) 2. Biotecnología
Molecular.(51)
(96) Aspergillus tubingensis
Colonización micorrizica Aspergillus niger Rhizopus oryzae
1. Aun no
encontrado 1. xKiA gene
(D-xilulosa quinasa, exón 1-2 2. xyrA gene
D-xilosa reductasa
pol gene
poliproteina completa
1. Residuos de Bauxite 2. Ácido láctico
1.Biorremediación
2. Acidulante y
conservante: Alimentos. Industrias químicas solubilizador y agente controlador de pH. En pinturas y resinas: solvente biodegradable. Plásticos : precursor del ácido poliláctico (PLA),polímero
biodegradable.(52,53)
(61) Penicillium brevicompactu m
Penicillium oxalicum
1. Aun no
encontrado
2. B-xilosidasa 3. lovastatina
1. Bioetanol(54) 2.Papel y pulpa 3.Farmacéutica(55) (61) Mucor
circinelloides Rhizopus oryzae Aspergillus niger 1. CaRA, carS,carA 2. Gpd1p-gox 3. CarG 1.Betacarotenos, Vitamina A
2. Glucoxidasa (GOX) 3.Carotenos
33 La (Tabla 4) muestra los resultados de la revisión de bibliografía sobre el potencial de las cepas estudiadas en el presente trabajo. Algunos de los hongos identificados en este estudio no solo muestran que tienen potencial industrial sino que son importantes desde el punto de vista sanitario al ser patógenos directos e indirectos causantes de enfermedades en humanos por la producción de micotoxinas (aflatoxinas) (36,50). En este caso, la biotecnología juega un rol importante en la búsqueda de la transformación de estos hongos de patógenos a hongos con potencial industrial como es el caso de Aspergillus fumigatus, Aspergillus flavus, Aspergillus oryzae, Aspergillus tubingensis y Penicillium brevicompactun. (48, 52, 58,59) En el caso de Aspergillus fumigatus, Aspergillus oryzae y Aspergillus flavus Cleveland et al. (2006) hacen una revisión de como la genómica logra transformar las rutas metabólicas que producen las micotoxinas en rutas que induzcan la producción de metabolitos secundarios con aplicación farmacéutica (Tabla 4). (50)
Se encontró que hay genes en común para un mismo género como es Aspergillus sp. que puede producir en más de una de sus especies invertasas de distinto tipo y con diferente rendimiento en la industria de alimentos. (52) Dentro de los seis hongos identificados es importante destacar el estudio de Veanna et al. (2011) que incluye las cepas de este estudio pertenecientes al género Aspergillus sp y las combina con técnicas de biología molecular para buscar genes que produzcan a nivel biotecnológico nuevas sustancias y con las que se puedan mejorar procesos biotecnológicos, sugiriendo la incursión de los hongos en nuevas industrias que podrían se amigables al ambiente haciendo de la identificación molecular de hongos un cambio en la perspectiva biotecnológica y clínica. Como por ejemplo Aspergillus terreus que interviene en la producción de ácido itacónico, en cuyo caso se obtienen mayores rendimientos que los observados por procesos de síntesis química. (62,63)
El avance en la identificación de nuevas cepas también implica el descubrimiento de nuevos productos y el mejoramiento de bioprocesos, como es el caso de Penicillium brevicompactum y Aspergillus tubingensis en las que se ha descubierto la producción de compuestos como biocombustibles, industria en la que se ha implicado ampliamente el uso de mucorales como Mucor circinelloides.
Para el caso de Aspergillus tubingensis y Penicillium brevicompactumRezende et a. (2013) con el objetivo de encontrar el potencial enzimático y toxigénico de hongos aislados de vainas de café encontraron que P. brevicompactum tiene la capacidad de degradar pectina, haciéndolo aplicable en el área de la biorremediación. (54, 64, 65,66)
34 7. CONCLUSIONES
1. Se realizó la identificación morfológica de las once cepas de la colección de microorganismos del GIFUJ, encontrando que la identificación molecular es importante debido a que algunas cepas presentan dificultad diferencial a nivel morfológico y esta es complementaria para la obtención de resultados más versátiles en la identificación de hongos filamentosos en género y especie.
2. La Identificación Molecular de las cepas permitió confirmar la clasificación para Aspergillus A-26, Aspergillus 3CV, Aspergillus oryzae, mientras que para Mucor circinelloides (papel filtro) se encontró que se trataba de Penicillium brevicompactum, Aspergillus niger se trataba de Aspergillus tubingensis.
3. Se identificó el potencial biotecnológico de estos hongos encontrándose que Aspergillus terreus es uno de los mejores hongos en la producción de ácido itacónico. Que tanto Aspergillus oryzae como Aspergillus flavus y Penicillium brevicompactum son patógenos que se han convertido en hongos con potencial en biorremediación, en la producción de sustancias para alimentos, en la producción de etanol entre otros.
4. La identificación molecular constituye una importante herramienta para el avance de proceso biotecnológicos mejorando su rendimiento, o encontrando métodos de producción más ecológicos.
8. RECOMENDACIONES
1. Se recomienda que las cepas que fueron identificadas por un solo gen (ITS o β-tubulina) se realice de nuevo la secuenciación con el otro par de cebadores para comprobar la especie de cada hongo.
2. Se recomiendo la re-secuenciación de los hongos que no se lograron identificar molecularmente, y de los que no se logró hacer el análisis filogenético por no tener un tamaño adecuado.
3. Se recomienda la estandarización de los métodos moleculares para la identificación de las cepas aisladas de los páramos de Guasca y Cruz Verde – Cundinamarca – Colombia hechos por el grupo de investigación de fitoquímica Universidad Javeriana (GIFPUJ).
35 9. BIBLIOGRAFIA
1. Nicolas Chemidlin Pre´vost-Boure´ 1, Richard Christen2, Samuel Dequiedt3, Christophe Mougel1,3,Me´ lanie Lelie`vre3, Claudy Jolivet4, Hamid Reza Shahbazkia5, Laure Guillou6, Dominique Arrouays4,Lionel Ranjard1,3. Validation and Application of a PCR Primer Set to Quantify Fungal Communities in the Soil Environment by Real-Time Quantitative PCR.2011. PLoS ONE.vol.6(9)
2. Muhammad Mukhtarb, Zahida Parveenc, David A. Logana. Isolation of RNA from the filamentous fungus Mucor circinelloides. Journal of Microbiological Methods 1998; 33: 115–118.
3. Kim, Ji Yeun, Soo-Hwan Yeo, Sung Yeol Baek, and Hye Sun Choi*. Molecular and Morphological Identification of Fungal Species Isolated from Bealmijang Meju. Journal Microbiology Biotechnology. (2011), 21(12): 1270–1279
4. Abolfazl Lotfi2, Mohammad Ali Tajick Ghanbary1*, Gholam Ali Ranjbar2 and Ahmad Asgharzadeh3. Screening of some Zygomycetes for cellulase activity.African Journal of Biotechnology 2010; Vol. 9(27), pp. 4211-4216
5. Rohlfs, M. Churchill, S. Fungal secondary metabolites as modulators of interactions with insects and other arthropods. Fungal Genetics and Biology.2011. 23-34.
6. Mouna Sahnouna, Samir Bejara, ∗, Aida Sayaria, Mohamed Ali Triki b, Mouna Kriaaa, Radhouane Kammouna. Production, purification and characterization of two -amylase isoforms from a newly isolated Aspergillus Oryzae strain S2. Process Biochemistry 2012; 47 ( 18–25)
7. Johannes Wöstemeyer, Anke Grünler, Christine Schimek and Derstin Voigt. Genetic Regulation of Carotenoid Biosynthesis in Fungi. Applied Mycology and Biotechnology.2005 5: 257-271.
8. Yasunobu Terabayashi, Motoaki Sano, Noriko Yamame, Junichiro Marui, Koichi Tamano, Junichi Sagara, Mitsuko Dohmoto, Ken Oda, Eiji Ohshima, Kuniharu Tachibana, Yoshitaka Higa, Shinichi Ohashi, Hideaki Koike, Masayuki Machida. Identification and characterization of genes responsible for biosynthesis of Kojic acid, an industrially important compound from Aspergillus oryzae. Fungal Genetics and Biology. 2010; 47: 953-961.
9. Lurá, María C., Benitez, Joel D; Jáuregui, Soledad; Gonzáles, Ana M. Evaluación de diferentes técnicas de extracción del ADN de hongos filamentosos. Revista FABICIB 2003; 7: 37-44.
10. Jorge Robles, Andrea García, Alejandra Ocampo. Obtención de metabolitos secundarios obtenidos en fermentación líquida de una cepa nativa aislada del páramo de Guasca, Cundinamarca de Mucor circinelloides y evaluación de su actividad antimicrobiana. Tesis pregrado. Facultad de Ciencias. Pontificia Universidad Javeriana, Bogotá, 2011, 12-14 pág.
11. Nancy P. Huérfano, Identificación, caracterización morfológica y conservación de hongos filamentosos aislados previamente de los páramos de Cruz verde y Guasca. Tesis pregrado. Facultad de Ciencias. Pontificia Universidad Javeriana, Bogotá, 2010, 2-9.
36 Colombia. Tesis de pregrado. Facultad de Ciencias. Pontificia Universidad Javeriana, Bogotá, 2011,140 pág.
13. Paula Juliana Rodríguez Pardo. Conformación de un banco de hongos filamentosos aislados previamente de los páramos de Guasca y Cruz verde. Tesis de pregrado. Facultad de Ciencias. Pontificia Universidad Javeriana, Bogotá, 91 pág.
14. Chaparro, A. Aislamiento e identificación de metabolitos producidos por la cepa nativa SPG 321 de Mucor circinelloides y evaluación de su actividad antimicrobiana. Tesis posgrado. Facultad de Ciencias. Pontificia Universidad javeriana, Bogotá 2010, 116 pág.
15. Vargas A, Villamizar D. Estudio preliminar de la producción, extracción y purificación de T2 toxina por Fusarium sporotrichioides NRRL 3299 en dos medios sintéticos. Tesis pregrado. Pontificia Universidad Javeriana. Facultad de Ciencias. Bogotá D.C. 172 pág.
16. L. Settani, A. Corsetti. The use of multiplex PCR to detect and differentiate food- and beverage-associated microorganisms: A review. Journal of Microbiological Methods.2007; 69:1-22.
17. Antoine Hermet, Delphine Méheust, Jéróme Mounier, Georges Barbier, Jean-Luc Jany. Molecular systematics in the genus Mucor with special regards to species encountered in cheese. Fungal Biology.2012; 116: 692-705.
18. Bruns T.D. & Shefferson R.P. Evolutionary studies of ectomycorrhizal fungi: recent advances and future directions. Canadian Journal of Botany. (2004).82: 1122-1132.
19. García, A. Realizar protocolos de producción y uso de microorganismos benéficos. Se inicia la fase i del banco de microorganismos utilizando claves taxonómicas macroscópicas y microscópicas. Alcaldía Mayor de Bogotá Jardín Botánico José Celestino Mutis.
20. Instituto De Genética - Universidad Nacional De Colombia. http://www.ssigmol.unal.edu.co/support.php. Consultado el 28 de Octubre de 2012
21. Gonzales R.Andrés. F. Caracterización Molecular de Poblaciones de Colletotrichum spp. Asociadas a Coffe arabica en Colombia y su aplicación en el diagnóstico del CBD. Trabajo de Grado. Pontificia Universidad Javeriana, Bogotá, 2007,126 pág.
22. Bulat, S.A., Beck, M.L: Alekhina, I.A., Jensen, D.F., Knudsen, I.M.B. and Stephensen, P. 2000. Identification of a Universaly Primed-PCR-Derived Sequence-Characterized Amplified Region Marker for an Antagonistic Strain of Clonostachys rosea and development of a Strain-Specific PCR Detection Assay. Applied and Environmental Microbiology.P.4758-4763
23. Cruz, L.P.2003.Uso de la diversidad genética de Beauveria bassiana para el diseño de alternativas de control de la Broca del café. Microbióloga Industrial. Pontificia Universidad Javeriana. Facultad de Ciencias Básicas. Carrera de Microbiología Industrial. Bogotá.Colombia.71p.
24. Vilgalys Lab. Duke University. http://biology.duke.edu/fungi/mycolab/primers.htm. Conserved primers sequences for PCR amplification and sequencing from nuclear ribosomal RNA. Consultado Mayo 9 2013
37 26. Amrani, L. &Corio-Costet M.F. A single nucleotide polymorphism in the β-tubulin gene di stinguishing two genotypes of Erysiphe necátor expressing differente symtoms on grapevine. Plant Pathology 2006 55: 3-15.
27. Guarro, J., Gené, J., Cano, F.J (2011) Caracterización fenotípica y molecular de hongos filamentosos oportunistas: Scedosporium, Acremonium, Phialemonium, Lecythophora Y Paecilomyces. Tesis de Doctorado. Departamento de ciencias médicas básicas. Universidad Rovira I Virgili.Reus-España.
28. Gil-Lamaignere C, Roilides E, Hacker J, Muller FMC. Molecular typing for fungi a critical review of the possibilities and limitations of currently and future methods. Clin Microbiol Infect 2003; 9: 172-185. 29. O’Donnell K, Cigelnik E, Nirenberg H. Molecular systematics and phylogeography of the Giberella
fujikuroi species complex. Mycologia. (1998). 90(3): 465-493
30. Webster J, Weber RWS. Introduction to fungi. 3rd ed. Cambridge University press,New York.
31. Harrison CJ, Langdale JA. A step by step guide to phylogeny reconstruction. Plant Journal.2006.45: 561– 572.
32. Salemi M, Vandamme AM. The phylogenetic Handbook. A practical approach to DNA and protein phylogeny. Cambridge University Press, United Kingdom.
33. Samson, R.A, Houbraken, J., Thrane U., Frisvad J.C, & Andersen B. Food and Indoor Fungi. Séptima Edición CBS.The Netherlands.(2011)
34. Barnett H, Hunter B. Illustrated genera of imperfect fungi. Curate Edición. The American Phytopathological Society. Pilot Knob Rodal, St Paul, Minnesota, USA.1999, 216 p.
35. Gilman J. A manual of soil fungi. The iowa state university press, Ames, Iowa, U.S.A. 1950. 572p.
36. Chang,P-K.,Ehrlich C.K.,T.Hua.S.S. Cladal relatedness among Aspergillus oryzae isolates and Aspergillus flavus S and L morphotype isolates. International Journal of Food Microbiology (2006) 108: 172-177.
37. Polizzoto R.cAndersen B., Martini M., Grisan S., Assante G., Musetti R. A polyphasic approach for the characterization of endophytic Alternaria strains isolated from grapevines. Journal of Microbiological Methods. (2012) 88: 162-171.
38. Silva.M.D. Batista.R.L., Rezende.F.E, Fungaro.P.M. Sartori D., Alves E. Identification of fungi of the genus Aspergillus section Nigri using polyphasic taxonomy.Brazilian Journal of Microbiology (2011) 42: 761-773.
39. Reis A.T., Oliveira T.D., Baquião A.C., Gonçalves S.S., Zorzete P., Corrệa B. Mycobiota and mycotoxins in Brazil nut samples from different states of the Brazilian Amazon region. International Journal of Food Microbiology (2012) 159: 61-68.
38 41. Culebras, M.P., Sempere C.A., Hervás S.M., Elizaquivel P., Aznar R., Ramón D. Molecular characterization of the black Aspergillus isolates responsibles for ochratoxin A contamination in grapes and wine in relation to taxonomy of Aspergillus section Nigri. International Journal of Food Microbiology.(2009). 132:33-41.
42. Gibbons G.John, Rokas A. The function and evolution of the Aspergillus genome. Trends in Microbiology, (2013) 21: 1-17.
43. Perrone G., Stea G.,Epifani F., Varga J., Frisvad C.J., Samson A. R. Aspergillus niger contains the cryptic phylogenetic species A. awamori. Fungal Biology (2011)115: 1138-1150.
44. Dyer S.P., O’Gorman M.C., A fungal sexual revolution: Aspergillus and Penicillium show the way. Current Opinion in Microbiology (2011) 14: 649-654.
45. Ho.Y.S. Chen F., Genetic characterization of Mortierella alpina by sequencing the 18S-28S ribosomal gene internal transcribed spacer region. Letters in Applied Microbiology (2008) 47: 250-255.
46. Fedora, N.D. Khaldi N., Joardar S.V., Amedeo P., Anderson J.M., et al. Genomic islands in the pathogenic filamentous fungus Aspergillus fumigatus. PLoS Genetics. (2008) 4:e1000046.
47. Andersen M.R., Salazar P.M., Schaap J.P., et al. Comparative genomics of citric-acid-producing Aspergillus niger ATTC 1015 versus enzyme-producing CBS 513.88. Genome Research. (2011) 21 :885-897.
48. Scazzocchio C. Aspergillus genomes: secret sex and the secrets of sex. Trends in Genetics. 22: 521-525.
49. Lubertozzi D. Keasling D.J. Developing Aspergillus as a host for heterologous expression. Biotechnology Advances .(2009)27:53-75
50. Cleveland E.T.,Yu J., Fedorova N., Bhatnagar D., Payne A.G.et al. Potential of Aspergillus flavus genomics
for applications in biotechnology. Trends in Biotechnology. (2009)27:3,151-157.
51. Rodrigues G.A.,Ping Y.L., Marcato D.P., Alves L.O., Silva P.C.M.et al. Biogenic antimicrobial silver
nanoparticles produced by fungi. Applied Microbiology biotechnology. (2013)97:775-782.
52. Veana F., Aguilar N.C., Salvado-Viader J.M., Herrera R.R. Invertasa del Género Aspergillus y su impacto Biotecnológico. Biotecnología. (2011). 15: 1, 1-29.
53. Hama S., Tamalampudi S., Shindo N., Numata T., Tamaji I., et al. Role of N-terminal 28-amino-acid región of Rhizopus oryzae lipase in directing proteins to secretory pathway of Aspergillus oryzae. Applied Microbiology biotechnology. (2008) 79: 1009-1018.
54. Babu G.A., Reddy S.M., Aspergillus tubingensis Improves the Growth and Native Mycorrhizal Colonization of Bermudagrass in Bauxite Residue. Biorremediation Journal. (2011)15(3): 157-164.
39 56. Lee J., Jang Y., Lee H., Lee S., Gyu H.K., Jae J.K. Screening for xylanase and β-xylosidase production from wood-inhabiting Penicillium strains for potential use in biotechnological applications. Environmental Science and Ecological Engineering. (2011).66(2):267-271.
57. Zhao J.Z., Pan Z.Y., Liu J.Q., Li H.X., Exposure assesment of lovastatin in Pu-erh tea. International Journal of Food Microbiology. (2013).164(1): 26-31.
58. Wöstemeyer J., Grünler A., Schimek C., Vougt K. Genetic Regulation of carotenoid biosynthesis in Fungi.Applied Mycology and Biotechnology. (2005).5: 257-274.
59. Csemetics A.,Nagy G.,Iturriaga A.E.,Szekeres A.,Eslava P.A.,et al. Expression of three isoprenoid biosynthesis genes and their effects on the carotenoid production of the zygomycete Mucor circinelloides. Fungal Genetics and Biology. (2011) 48:696-703.
60. Trivedi S.,Divecha J.,Shah A. Optimization of inulinase production by a newly isolated Aspergillus tubingensis CR16 using low cost substrates. Carbohydrate Polymers. (2012). 90: 483-490.
61. Simon X., Philippe D., Feasibility of Generating Peaks of Bioaerosols for Laboratory Experiments. Aerosol and Air Qualitiy Research. (2013).13:877-886.
62. Kuenz A., Gallenmüller. Willke T., Vorlop D.K. Microbial production of itaconic acid: developing a stable platform for high product concentrations. Applied Microbiology Biotechnology.(2012) 96: 1209-1216.
63. Klement T., Büchs J., Itaconic acid- A biotechnological process in change. Bioresource Technology. (2013). 135: 422-431.
64. Rezende, E.F., Couto, F.A., Borges, J.G., da Silva, D.M., Batista, L.R. Enzymatic and toxigenic potential of fungi isolated from coffee beans. Coffe Science. (2013). 8(1): 69-77.
65. Gouda N., Raoult D., Drancourt M. Plant and Fungal Diversity in Gut Microbiota as Revealed by Molecular and Culture Investigations. PLOS ONE. (2013). 8(3): e59474.
66. Yamada O., Takara R., Hamada R., Hayashi. Tsukahara M., Mikami S. Molecular biological researches of Kuro-koji molds, their classification and safety. (2011). 112(3):233-237.
40 ANEXO1. Macroscópia de cada Hongo en PDA, Saboreaud, Extracto de Malta, Czapeck o Avena.
CEPA(COD) PDA Saboreaud Extracto de Malta Czapeck/ Avena
Aspergillus terreus (81)
Aspergillus flavus (13)
Aspergillus fumigatus
41 Aspergillus
niger (96)
Aspergillus oryzae (98)
42 Aspergillus
TATA
Penicillium brevicompac -tum (61)
43 Rhizopus
oryzae (48)
Mucor circinelloide
44 ANEXO 2. MICROSCOPIA DE CADA CEPA. Fotografías microscópicas con estructuras claves
para la identificación morfológica de cada cepa.
Cepa/Código GIFUJ Microscopia
Aspergillus (A-26)terreus (81)
Aspergillus (CV3) flavus
(13)
Aspergillus oryzae (98)
Aspergillus (CV29)
fumigatus (82)
Aspergillus niger (96)
45 Aspergillus SPG 64 (50)
Aspergillus TATA (17)
Penicillium
brevicompactum (61)
Penicillium spp. (CV7)(56)
Mucor circinelloides
(61)
46 Rhizopus oryzae
(Mucor spp.) (48)
ANEXO 3. MEDIOS DE CULTIVO
MEDIO PAPA DEXTROSA + CLORANFENICOL (39g /L)
Extracto de papa ……… 4,0 g Glucosa ………. .20,0 g Agar………..15,0 g Cloranfenicol………0.4g
pH
final:5,6 +- 0,2
Esterilizar a 121°C a 15 psi/15min
MEDIO SABOREAUD (65g/L)
Glucosa Saboureaud, Agar
D (+)-Glucosa...40,0g Mezcla de Peptonas...10,0g Agar...15,0g Cloranfenicol………0.4g pH final: 5,6 ±0,2
Esterilizar a 121°C a 15 psi/15min MEDIO EXTRACTO DE MALTA (48g/L)
Extracto de Malta...12,75g
Dextrina...2,75g
Glicerina...2,35g
Peptona de Gelatina...0,78g
Agar...15,0g
pH final: 4,6 ±0,2
Esterilizar a 121°C a 15 psi/15min
MEDIO AVENA
Avena……….………..60 g. Agar………12 g
Extracto de levadura………... 2 g. Esterilizar a 121°C a 15 psi/15min
MEDIO CZAPECK
Sacarosa………30 g Nitrato sódico………..…3.0 g Cloruro potásico……….0.5 g Sulfato de hierro (III)………...0.01 g Hidrógeno fosfato di potásico…...1.0 g Agar-agar………...13 g
CALDO NUTRITIVO
Extracto de Carne...3,0g Peptona de Gelatina...5,0g pH final: 6,8 ±0,2
2 ANEXO 4. ELECTROFORESIS PUREZA DE ADN
3