UNIVERSIDAD TÉCNICA PARTICULAR DE LOJA
La Universidad Católica de Loja
TITULACIÓN DE BIOQUÍMICO FARMACÉUTICO
Estudio de la expresión del gen
SERK
en respuesta al
ataque de hongos patógenos
Colletotrichum acutatum
y
Botrytis cinerea
en
Cattleya maxima
Lindl
Trabajo de fin de Titulación
AUTOR
Janeth Paola Rivera Requelme
DIRECTORA
Augusta Yadira Cueva Agila, Ph.D.
Loja- Ecuador
II CERTIFICACIÓN
Ph.D.
Augusta Cueva Agila
DIRECTORA DEL TRABAJO DE FIN DE TITULACIÓN
CERTIFICA:
Que el presente trabajo, denominado: “Estudio de la expresión del gen SERK en
respuesta al ataque de hongos patógenos Colletotrichum acutatum y Botrytis cinerea
en Cattleya maxima Lindl” realizado por el profesional en formación: Srta. Janeth
Paola Rivera Requelme, cumple con los requisitos establecidos en las normas generales para la Graduación en la Universidad Técnica Particular de Loja, tanto en el aspecto de forma como de contenido, por lo cual me permito autorizar su presentación para fines pertinentes.
Loja, Marzo 2013
f) _______________________
CI: 110385221-4
Augusta Cueva Agila, Ph.D
III
CESIÓN DE DERECHOS
“Yo Janeth Paola Rivera Requelme declaro ser autora del presente trabajo y eximo expresamente a la Universidad Técnica Particular de Loja y a sus representantes
legales de posibles reclamos o acciones legales”.
Adicionalmente declaro conocer y aceptar la disposición del Art. 67 del Estatuto Orgánico de la Universidad Técnica Particular de Loja que en su parte pertinente
textualmente dice: “Forman parte del patrimonio de la Universidad la propiedad
intelectual de investigaciones, trabajos científicos o técnicos y tesis de grado que se realicen a través, o con el apoyo financiero, académico, o institucional (operativo) de la
Universidad”.
f) ________________________
CI: 1104708027
Janeth Paola Rivera Requelme.
IV AUTORÍA
Las ideas, conceptos procedimientos y resultados vertidos en el desarrollo del presente trabajo de investigación son de absoluta responsabilidad de su autora
V DEDICATORIA
A Dios, por haberme brindado la salud y fortaleza necesaria para
culminar con éxito mis objetivos planteados.
A mis padres: Horacio y Patricia, por su apoyo incondicional durante
todo el transcurso de mi vida, por sus sabios consejos y enseñanzas, y
además por ser ejemplo de superación y perseverancia.
A mis hermanos Horacio, Giovanna y Cristian, por estar ahí cuando
más los necesito, por compartir conmigo la alegría de mis triunfos y por
todo el amor que siempre me brindan.
A mis sobrinos Dome y Luchito, quienes con su sonrisa y ocurrencias
despiertan en mí una inmensa alegría.
VI AGRADECIMIENTOS
Expreso mi gratitud:
A la Universidad Técnica Particular de Loja que por medio del Departamento de Ciencias Naturales, Departamento de Ciencias Agropecuarias y Alimentos, y la Titulación de Bioquímica y Farmacia me ha dado la oportunidad de preparar la presente investigación.
A mi directora de tesis Augusta Cueva Ph. D.,por brindarme su tiempo y por ser la fuente para cada pregunta que surgía durante la investigación.
A mi Co – Tutora Ing. Jaqueline Rojas por su ayuda y disposición en la elaboración de mi proyecto de fin de carrera.
A mis Padres y hermanos por su apoyo y confianza depositada en mí durante mi carrera universitaria, sobre todo a las palabras sabias que me han iluminado en el periodo de estudio.
VII
ÍNDICE DE CONTENIDOS
CONTENIDO
PÁG
Portada I
Certificación II
Contrato de Cesión de Derechos de Tesis III
Dedicatoria VI
Autoría V
Agradecimiento VI
Índice de Contenidos VII
Índice de Figuras IX
Índice de Tablas IX
Resumen X
Artículo científico XI
1. PRESENTACION DEL FIN, PROPÓSITO Y COMPONENTES
DEL PROYECTO
1
1.1Fin del proyecto. 2
1.2 Propósito del proyecto. 2
1.3 Componentes del Proyecto. 2
II. INTRODUCCIÓN
3
2.1 Generalidades Orquídeas. 4
2.1.1 Cattleya maxima Lindl. 4
2.1. 2 Cultivo in vitro de orquídeas. 5
2.2 Hongos. 5
2.3 Enfermedades producidas por hongos. 6
2.3.1 Colletotrichum acutatum. 6
2.3.1.1 Características morfológicas. 7
2.3.1.2 Taxonomía. 7
2.3.2 Botrytis cinerea. 7
2.3.2.1 Características morfológicas. 8
2.3.1.2 Taxonomía. 8
2.4 Receptor de Embriogénesis Somática tipo quinasa (SERK). 8
2.5 PCR en tiempo Real. 10
III. MATERIALES Y METODOLOGÍA
12VIII
3.2 Aislamiento de Colletotrichum acutatum y Botrytis cinerea de
material vegetal enfermo.
13
3.2.1 Obtención de cultivos monospóricos. 14
3.3 Identificación morfológica. 14
3.4 Identificación molecular. 14
3.4.1 Extracción de ADN. 14
3.4.2 Amplificación de ADN mediante PCR. 15
3.4.3 Cloning y purificación de productos. 16
3.4.4 Digestión enzimática. 16
3.4.5 Secuenciación. 17
3.4.5.1 Análisis de secuenciación. 17
3.5 Inoculación de hongosen plantas in vitro de Cattleya maxima Lindl. 18
3.6 Extracción de ARN, síntesis de cDNA y PCR de Tubulina. 19
3.6.1 Extracción de ARN y síntesis de cDNA. 19
3.6.2 PCR de Tubulina. 19
3.7 PCR de tiempo Real. 20
3.8 Análisis de datos. 21
IV. RESULTADOS Y DISCUSIÓN
224.1 Identificación morfológica Colletotrichum acutatum y Botrytis cinerea. 23
4.2 Identificación molecular. 25
4.2.1 Extracción ADN. 25
4.2.2 Amplificación mediante PCR. 26
4.2.3 Secuenciación y análisis. 27
4.3 Inoculación de patógenos en plantas in vitro de Cattleya maxima
Lindl.
28
4.4 Extracción de ARN y síntesis de cDNA. 30
4.5 Expresión del gen SERK en respuesta al ataque de hongos
patógenos.
31
V. CONCLUSIONES
34VI. ANEXOS
361. Protocolo de extracción de ADN ( DNeasy Plant Mini Kit de Qiagen). 37
2. Protocolo para Cloning (Invitrogen). 38
3. Protocolo para purificación S.N.A.P.™ MidiPrep Kit (Invitrogen). 39
4. Protocolo para extracción de ARN (Plant Rneasy Mini de Quiagen). 40
IX
ÍNDICE DE FIGURAS
FIGURAS
PAG.
1. Planta de Cattleya maxima Lindl. 5
2. Sintomas de C. acutatum y B. cinerea sobre tejido vegetal. 13
3. Estructura morfológica de Colletotrichum acutatum. 23
4. Estructura morfológica de Botrytis cinerea. 24
5. ADN de Colletotrichum acutatum y Botrytis cinerea. 26
6. Productos amplificados con cebadores universales para Colletotrichum
acutatum y Botrytis cinerea, y cebadores específicos para B. pseudocinerea.
27
7. Vista general de Cattleya maxima inoculada con Colletotrichum
acutatum.
29
8. Vista general de Cattleya maxima inoculada con Botrytis cinerea. 29
9. Vista general de Cattleya maxima Testigo. 30
10. ARN de tejido infectado con Colletotrichum acutatum , Botrytis cinerea y tejido sano.
30
11.Expresion del gen CmSERK en respuesta al ataque de hongos
patógenos.
32
ÍNDICE DE TABLAS
TABLAS
PAG.
1. Preparación de solución para amplificar ADN mediante PCR. 16
2. Preparación de solución para digestión enzimática. 17
3. Tratamientos para inoculación de hongos en Cattleya maxima Lindl. 18
4. Preparación de solución para PCR de Tubulina. 20
5. Preparación de solución para PCR de tiempo Real. 20
6.Similitud de secuencias de hongos obtenidos mediante la búsqueda en
el GenBank (Herramienta Blastn).
X RESUMEN
El género Cattleya agrupa orquídeas originarias de zonas tropicales del Sur y de
Centro América, una de las especies de importancia ornamental es Cattleya maxima
Lindl. En este estudio se aisló Colletotrichum acutatum y Botrytis cinerea y se
determinó su patogenicidad mediante inoculación en plantas in vitro de Cattleya
maxima Lindl. Las pruebas de patogenicidad resultaron positivas a los siete y dos días
de la inoculación para Colletotrichum acutatum y Botrytis cinerea respectivamente.
Mediante la técnica de PCR en tiempo real se reveló que el gen CmSERK es más
expresado en el tejido infectado por el ataque de hongos. Estos resultados sugieren que el gen CmSERK juega un importante papel en la activación de respuestas
relacionadas con la defensa.
Palabras clave: Cattleya maxima Lindl, Colletotrichum acutatum, Botrytis cinerea,
XI
Study of
SERK
gene expression in response to the attack of
pathogen fungi
Colletotrichum acutatum
and
Botrytis cinerea
in
Cattleya maxima
Lindl
Janeth Rivera
a, 1*, Jaqueline Rojas
b, 1, Augusta Cueva
c, 1a
Titulación de Bioquímica y Farmacia
b
Departamento de Ciencias Agropecuarias y Alimentos
c
Departamento de Ciencias Naturales
1
Universidad Técnica Particular de Loja, San Cayetano Alto s/n C.P. 11 01 608, Loja – Ecuador
ABSTRACT
The genus Cattleya groups orchids originate in tropical zones of South and Central America, one species of ornamental importance is Cattleya maxima Lindl. In this study Colletotrichum acutatum and Botrytis cinerea were isolated and their pathogenicity was determinedby in vitro
inoculation of Cattleya maxima. Pathogenicity tests resulted positive for infection with C. acutatum after seven days of inoculation while as for B. cinerea the symptoms of infection appeared after two days. Quantitative PCR revealed that CmSERK gene is more expressed in tissue under fungal attack. These results suggest that the CmSERK gene plays an important role in the activation of defense-related responses.
Keywords:Cattleya maxima, Colletotrichum acutatum, Botrytis cinerea, Quantitative PCR,
SERK gene
*
XII
1. Introduction
Orchidaceae family gathers great quantity of species from the Plant Kingdom. It has been estimated that there 25,000 species grouped in about 900 genera, besides 70,000 registered artificial hybrids [1, 2]. The genus Cattleya orchids groups together originating in tropical zones of South and Central America, comprises something more than 60 species commonly epiphytes and a small part of lithophytes, with pseudobulbs from few centimeters up to one meter, that carry at its end one or two sheets without petiole [3], the roots have a spongy membrane which protects them from drying out and also serve to absorb water and minerals [4]. Cattleya maxima is an epiphytic orchid which is distributed in Colombia, Ecuador and the northern Peru. In Ecuador is more abundant in the provinces of El Oro, Guayas, Loja, Santa Elena, Manabí and Los Rios [5].
In Orchids the development of fungi pathogens as Botrytis cinerea and Colletotrichum acutatum
is favored by the presence of weak tissues and injury in plants [6]. C. acutatum (teleomorph
Glomerella acutata) is a pathogen which may be latent within the host tissues for a short or long period of time. Once the mold has sufficiently developed inside the plant, dark fruiting bodies are produced, causing the typical symptoms. Mycelium in culture is usually white, pale-grey or pale-orange. Conidia are, fusiform, thin-walled, aseptate, hyaline, clavate to circular and light to dark-brown [7, 8, 9].
Botrytis cinerea (teleomorph Botryotinia fuckeliana) also known as gray mold, is a pathogen that penetrates through wounds and dying foliage, the incidence of the disease increases when there are prolonged periods of moisture and low temperatures (15-20 °C). The characteristic symptoms of the disease are grayish mildew on fruits, in leaf can be observed necrosis around the point of entry being able advance to the rest of the leaf if conditions are favorable for the fungus [10].
The effectiveness of a pathogen fungus of producing disease depends in large part on the conditions in which it develops and inoculates. Conditions of growth must be controlled considering the culture medium, its volume, aeration, light, temperature. The strains of the pathogens need multiply to inoculate hosts, because they only infect through their spores [11]. Plants have developed multi defense strategies against infections that allow recognition specific pathogens; this early recognition is one of the keys that activate the defense response effective [12].
Many receptor like - kinases (RLKs) play an important role in cell metabolism leading to growth and defense response for activate a large number of gene expression. The Somatic Embryogenesis Receptor Kinase (SERK) gene is claimed to have an important role. SERK gene was first isolated from carrot embryogenic cells, hailed as a molecular marker for somatic embryogenesis [13]. During the last few years, it has been associated the function of this gene mediating the signal transduction leading to the defense and disease resistance [14, 15]. Recent studies have revealed that deletion of the SERK gene expression increased the susceptibility to pathogenic microbial attack on cocoa [16].
Through molecular studies has been shown that the resistance genes encode components of the host immune system, which confer the ability to recognize and respond to pathogens [17]. Recently it has been used to characterize the SERK gene in two species of orchids distributed in the South of the country: Cyrtochilum loxense (Cueva et al., 2012) and Cattleya maxima
XIII
maxima Lindl plants, study of SERK gene expression by real-time quantitative PCR in response to the attack of fungal pathogens Botrytis cinerea and Colletotrichum acutatum, order to know the functionality of the gene in other plant processes.
2. Material and methods
2.1 Biological material
Fungus samples were collected from tree tomato (Solanum betaceum) and strawberry (Fragaria vesca) with symptoms of Colletotrichum acutatum and Botrytis cinerea respectively. The fruit surface was disinfected with alcohol 70% and the vegetable tissue with disease symptoms was sown directly in Petri plates containing PDA medium. The Petri plates were incubated in the dark in an inverted position at 25oC during ten days. With the purpose for establishing a more dependable cultivation, monosporic isolates were performed to ensure the authenticity and purity of both fungi.
2.2 Morphological Identification
The macroscopic characteristics as shape and color of the pathogenic cultures were reviewed after ten days of incubation. The culture purity was checked through microscopic visualization as well as the shape of conidias according to Barnett and Hunter [19].
2.3 Molecular Identification
DNA was extracted from macerated mycelium, was used the extraction kit DNeasy Plant Mini Kit from Qiagen ®, by following the manufacturer's protocol. The quality of DNA extracted was determined by electrophoresis in 1% agarose gel and by spectrophotometry in Nanodrop 2000.
PCR amplification was performed with universal primers. For C. acutatum and Botrytis cinerea
the ITS1primer (5'-TCC GTA GGT GAA CCT GCG G-3 ') was used, together with the conserved ITS4 primer (5’-TCC TCC GCT TAT TGA TAT GC -3’) [20]. MS547 specific primers (5'-AAG GAG GAC GT TGGA AGG AT-3') / (5'-AAG TCC AGA ATC TCG ATG TAT TTG T-3 ') [21] were used for B. pseudocinerea, a kind of Botrytis with morphological characteristics similar to the one studied in our research. MS547 (encoding the ATP-dependent RNA helicase DBP7; reference gene in the B. cinerea B0510 genome: BC1G_03202.1)
The amplification reaction was performed for a final volume of 20 μl, containing 2 μl of fungic DNA, 0.4 μl each primer, 0.8 μl of BSA, 10μl Phusion polymerase mix and 6.4 μl sterile
deionized distilled water to complete the final volume. Amplification was carried out under the following conditions for C. acutatum: one cycle at 98oC for 30 seconds; 30 cycles at 98oC for 10 seconds, 60oC for 20 seconds and 72oC for 30 seconds; one final cycle at 72oC for 7 minutes, and a final cooling temperature of 4 ° C. The conditions for B. cinerea are one cycle at 95oC for 5 minutes; 40 cycles at 95oC for 30 seconds, 62oC for 30 seconds and 72oC for 90 seconds; one final cycle at 72oC for 5 minutes, and a final cooling temperature of 4 ° C. The amplification products were verified in 1% agar gel through electrophoresis at 128V, 300mA for 20 minutes, the products were observed under ultraviolet light.
2.3.1 Cloning and DNA purification
XIV
2.3.2 Sequencing and Analysis
DNA purified was sequenced using Applied Biosystems 3500 Genetic Analyzer; universal primers M13-Fw (5’TGT AAA ACG ACG GCC AGT3’) and M13-Rv (5’TGC CAG GAA
ACA GCT ATG AC3’) were used. The sequences obtained were analyzed using the website
VecScreen NCBI (National Center for Biotechnology Information), SMS web (Sequence Manipulation Suite) tool Reverse complement (http://www.bioinformatics.org/sms2/rev_comp.html). The identified sequences were compared with GenBank database in NBCI (http://blast.ncbi.nlm.nih.gov/Blast.cgi) using the BLASTn web interface.
2.4 In vitro inoculation Cattleya maxima Lindl plants with pathogenic fungi
Healthy plants of Cattleya maxima in vitro 8-month-old maintained on Murashige & Skoog (MS) medium were inoculated with a fungal suspension of 1x106 spores/ml of C. acutatum and B. cinerea [22]. The spore inoculation was performed in the adaxial part of two leaves (apex). Inoculation of distilled water was used as negative control. The plants were placed in a growth chamber with proper incubation conditions for the development of fungi: temperature 19 ° C, humidity 18%, under photoperiod of 16h light and 8 hours darkness.
To determine the presence of symptoms of the disease on the apexes was seen in the external appearance: appearance of spots time, necrosis or invasion of the fungus to the whole plant.
2.5 RNA extraction, cDNA synthesis and PCR Tubulin
Four samples per plant (apexes), previously stored at - 80° C were selected (2 healthy and 2 infected) for RNA extraction. RNA was extracted using the kit Planta RNeasy Mini (Qiagen) following the manufacturer's instructions. Each RNA sample was analyzed by electrophoresis in 1% agarose gel.
The cDNA synthesis was performed by preparing a mix solution with 10µl of 2x RT Buffer, 1µl of 20x RT Enzyme (Applied Biosystem) and 9µl of RNA, obtaining a final volume of 20µl.
To verify the synthesis of cDNA it used as reference gene Tubulin using the following primers: Tubulin_Onc_Hou_Fw: 5' GGA TTA GGC TCT CTG CTG TTG G 3' and Tubulin_Onc_Hou_Rv:5' GTG TGG ATA AGA CGC TGT TGT ATG 3' [23]. For e Tubulin
PCR we performed a mix with: 10.05μl of sterile deionized distilled water, 4μl of Buffer, 1.6μl of MgCl2,1μl of Fw Tubulin primer, 1μl of Rv Tubulin primer, 0.3μl of dNTPs, 0.05µl of GoTAQ and 2µl of cDNA, obtaining a final volume of 20μl per sample, this volume was
collected in microcentrifuge tubes for PCR and then was placed in the Applied Biosystems thermocycler, taking into account the following conditions: one cycle at 95oC for 5 minutes; 34 cycles at 95oC for 30 seconds, 58oC for 40 seconds and 72oC for 60 seconds; one final cycle at 72oC for 5 minutes, and a final cooling temperature of 4 ° C. Each sample was analyzed by electrophoresis in 1% agarose gel.
2.6 Real – Time quantitative PCR
Real time PCR analysis was performed using the Fast SYBR® Green Master Mix (Applied Bisystems) following the manufacturer's protocol. For each sample was performed a mix with: 10µl of Master Mix, 1 µl of each primer, 6 µl of H2Odde and 2 µl of cDNA, obtaining a final volume of 20µl. The gene expression was performed in the Applied Biosystems 7500 Fast Team Real-Time PCR System.
To normalize the expression of SERK gene we used as reference gene Tubulin [23], and SERK
XV
SERK1_Cm_RT_Rv: 5' AAT GTT CTT GTG GCT TAC GAC G 3' [24]. The analyses of the obtained data were using LingReg program.
4. Results and Discussion
4.1 Morphological and Molecular identification
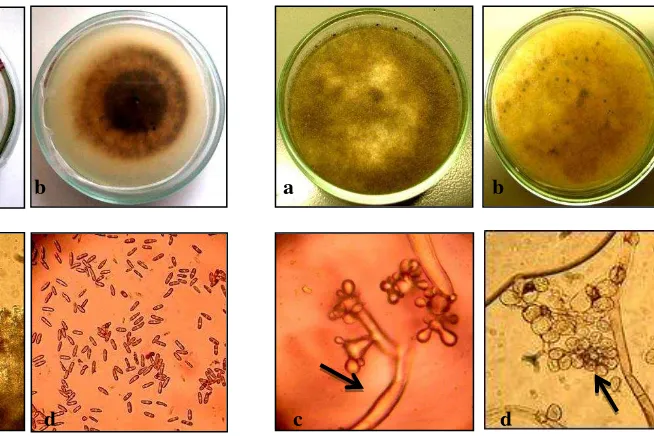
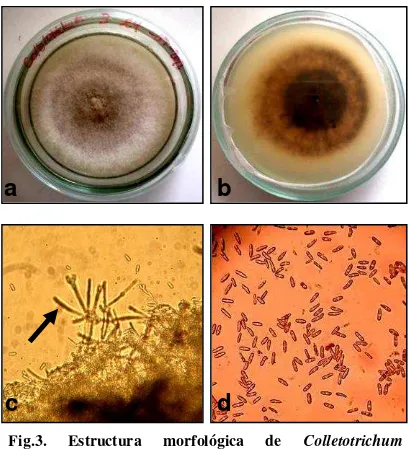
The C. acutatum isolates submitted a greyish - white mycelium, cottony [25, 26] and compact with conididiales masses dispersed color orange / salmon, the conidia were cylindrical, elongated, with pointed ends, these characteristics are similar to those reported for
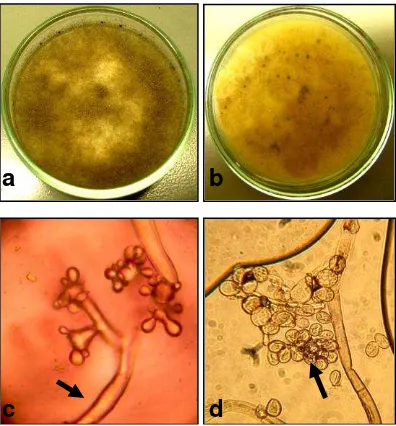
Colletotrichum acutatum by Walker et al [27] (Fig.1). In contrast B. cinerea presented a gray mycelium, long branched conidiophores with conidia oval and globose (Fig.2), corresponding to
Botrytis cinerea morphology, similar to that observed by Barnett and Hunter [18] and Domsch et al. [28]
a b a b
c d c d
The morphology of conidia and macroscopic review are not satisfactory elements to distinguish species [29, 30], that's why we also performed molecular characterization of fungi using molecular techniques such as PCR and sequencing, useful to classify or differentiate these species.
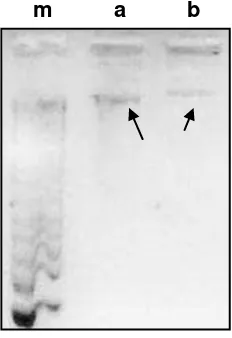
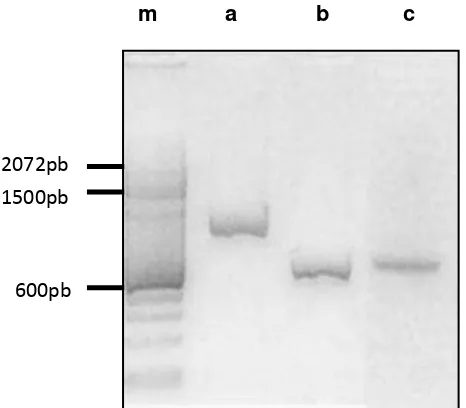
The quality of DNA extracted was determined by electrophoresis and spectrophotometry in Nanodrop 2000, giving the following concentrations: C. acutatum 170 ng/ μl y B. cinerea 118 ng/ μl, amount sufficient for performing PCR, since this technique allows amplification mold DNA from amounts as low as 0.1ng [31]
Amplification of the ITS region of ribosomal DNA (rDNA) with the primers ITS1/ITS4, amplified a fragment of ~ 600pb, for C. acutatum and Botrytis cinerea. The entire ITS region measured from 600 to 800 bp and can be amplified easily with universal primers [32]. Ms547 primers amplified a product of ~ 900pb.
4.2 Sequence and analysis
[image:15.595.178.505.258.476.2]According to the analysis in the web interface Blast, the sequences obtained from the isolates of the pathogens turned out to be Colletotrichum acutatum (theleomorph Glomerella acutata) and
[image:15.595.93.314.259.476.2]Fig. 2. Morphologic structure of B. cinerea. a. Mycelium obverse b. Mycelium reverse c. Conidiophores d. Conidia
XVI
[image:16.595.321.538.603.729.2]Botrytis cinerea (theleomorph Botryotinia fuckeliana); with an identity percentage between 98-100% in GenBank database (Table 1).
Table 1. Similarity of fungal sequences obtained by searching the GenkBank (Blastn tool).
Code Access Code Description Identities Max identity
(Genk Bank)
Colletotrichum spp JN715841.1 Glomerella acutata 583/584 99% ITS1/ITS4
Botrytis spp HM849615.1 Botryotinia fuckeliana 539/539 100% ITS1/ITS4
Botrytis spp XM_001558120.1 Botryotinia fuckeliana 919/938 98% MS547
The E-value was equal to zero in all analyzed sequences.
Morphological characteristics of tomato isolates indicated that the causal agent could be C. acutatum but the morphological characteristics of conidia were not enough. However, the nucleotide sequence amplified with ITS1/ITS4 primers confirmed a 99% identity of the agent that was isolated from tomato to C. acutatum theleomorph state of Glomerella cingulata.
The nucleotide sequence amplified with ITS1/ITS4 primers and MS547 primers for Botrytis cinerea was identified in the GenBank database with a maximum of 100% and 97% identity respectively, asserting that the agent isolated from strawberry tissue was Botrytis cinerea but not Botrytis pseudocinerea. These results demonstrate that the rDNA analysis is a reliable method for taxonomic species identification [33, 34].
4.3 In vitro inoculation of Cattleya maxima Lindl plants with pathogenic fungi
Pathogenicity tests were positive after inoculation, for seven days in the case of Colletotrichum acutatum and two days in the case of Botrytis cinerea with manifestation of symptoms characteristic of pathogenic fungi. Plants infected with C. acutatum showed dark-brown lesions, sunken and raised rims (Fig. 3). Plants inoculated with Botrytis cinerea showed necrotic lesions around the site of infection (Fig.4).
a b a b
Fig. 3. a. General view Cattleya maxima inoculated with C. acutatum b. apex with disease symptoms
[image:16.595.84.300.603.729.2]XVII
Control plants showed no symptoms during these time periods (Fig.5)
4.4 SERK gene expression in response to attack by pathogen fungi in Cattleya maxima Lindl.
Resistant plants have the ability to recognize a pathogen invasion because they are molecularly equipped with an alert signaling system [35]. Several components are involved in this signaling event. The first is a unique receptor protein that is located either at the outer limits of the plant cell or within the cytosol. Other components include proteins that are responsible to transduce the signal to the nucleus where the induced expression of defense genes is activated [36]. Included within this group the SERK gene. Several lines of evidence have demonstrated that
SERKs, as a group of LRR-RLKs, play important roles in the recognition of pathogens that activate effective defense responses.
In this study, we evaluated the CmSERK gene expression facing pathogenic fungi attack in Cattleya maxima plants. The Real – Time Quantitative PCR analysis revealed that the fungal infection, is related to gene expression CmSERK. In plants, there are numerous receptor-like kinases (RLKs), which are involved in perceiving the external pathogenic signals and transferring the signals inside plant cells to activate a large number of gene expressions [37].
The data presented in Fig. 6 indicated that tissues infected with Colletotrichum acutatum and
[image:17.595.214.371.118.254.2]Botrytis cinerea led to increase CmSERK gene expression, as compared with healthy tissue. These results are presented in a manner similar to the results reported by Hu et al. [14] which found that OsSERK1, a newly identified rice SERK gene, was induced by pathogen infection and by defense signaling molecules such as salicylic acid, jasmonic acid, and abscisic acid and that constitutive overexpression of OsSERK1 led to an increase in host resistance to the blast fungus, these transgenic rice plants showed an increase resistance to Magnapothe grisea.
Fig. 5. Cattleya maxima control
[image:17.595.182.427.604.734.2]XVIII
Moreover lettuce plants exhibited LsSERK gene silencing, showed reduced ability and became more susceptible to Sclerotinia attack. Thus, the elimination of LsSERK via RNA silencing corroborates the hypothesis that SERK is involved in plant defense, since these plants had to resist fungus attack [38].
The SERK genes belong to a small family of five plant receptor kinases that are involved in at least five different signaling pathways. [39]. One member of this family, BRASSINOSTEROID INSENSITIVE1 (BRI1)-ASSOCIATED KINASE1 (BAK1), also known as SERK3, is the coreceptor of the brassinolide (BR)-perceiving receptor BRI1, a function that is BR dependent and partially redundant with SERK1. BAK1 (SERK3) alone controls plant innate immunity, is also the coreceptor of the flagellin receptor FLS2, and, together with SERK4, can mediate cell death control. [40, 41].
In conclusion, SERK gene in addition to being involved in developmental pathways such as somatic embryogenesis also plays an important role against pathogen infection, in our study it was determined that CmSERK gene expression was stronger in infected tissue by fungal suspension corroborating that is involved in signaling pathways of the plant, when attacked by pathogens.
Acknowledgements
The author wants to thank the financial support and the facilities given by Universidad Técnica Particular de Loja through the Departamento de Ciencias Naturales and Departamento de Ciencias Agropecuarias y Alimentos
References
[1] Pupulin, F. Conservación de orquídeas: Una guía para autoridades administrativas. San José, Costa Rica, Editorial SEIDIN. 2005; Pp. 37.
[2] Rivera, G. Orquídeas: Generalidades y cultivo. Editorial Fundación UNA, Heredia, Costa Rica, 1998; Pp.266.
[3] Kuan, C y González, L . Introduccion al cultivo y manejo de las orquideas. Instituto Nacional de Aprendizaje. San José, Costa Rica. 1993; Pp 1 – 16, 47 -53, 75 -76.
[4] Warner, J . Descripción botánica del hibrido Cattleya Kinneri x Cattleya maxima. Jardin Botanico Lankester. Cartago, Costa Rica; 2000.
[5] Dodson, C., Luerand, C y Gustavo L. Orchids of Ecuador. Soluciones Graficas D Yg. Quito – Ecuador; 2004.
[6] Wit, P. How plants recognize pathogens and defend themselves. Cell Mol Life Sci. 2007;64 : 2726 – 2732.
[7] Mordue, J.E.M. Colletotrichum acutatum. CMI Descriptions of Pathogenic Fungi and Bacteria No.
630. CAB International, Wallingford, UK, 1979.
[8] Baxter, A.P.; Westhuizen, G.C.A. van der; Eicker, A. Morphology and taxonomy of South African
isolates of Colletotrichum. South African Journal of Botany 2, 1983; 259-289
[9] Gunnell, P.S.; Gubler, W.D.Taxonomy and morphology of Colletotrichum species pathogenic to
strawberry. Mycologia 84, 1992;157-165.
[10] Latorre, BA; Agostín, E; San Martín, R; Vázquez, GS . Effectiveness of conidia of Trichoderma harzianum produced by liquid fermentation against Botrytis bunch rot of table grape in Chile. Crop Protection. 1997; 16: 209- 214.
[11] Freeman, S., Katan, T and Shabi, E. Characterization of Colletotrichum species responsible for anthracnose diseases of various fruits. Plant Disease. 1998; 82: 596 – 605
[12] Martin GB, Bogdanove AJ, Sessa G. Understanding the functions of plant disease resistance proteins. Annu Rev Plant Biol. 2003; 54:23–61
[13] Schmidt ED, Guzzo F, Toonen MA, de Vries SC. A leucine-rich repeat containing receptor-like kinase marks somatic plant cells competent to form embryos. Development. 1997; 124:2049–2062
[14] Hu H, Xiong L, Yang Y. Rice SERK1 gene positively regulates somatic embryogenesis of cultured
XIX
[15] Song D, Li G, Song F, Zheng Z. Molecular characterization and expression analysis of OsBISERK1, a gene encoding a leucinerich repeat receptor-like kinase, during disease resistance responses in rice. MolBiol Rep 35. 2008; (2):275–283
[16] Santos MO, Romano E, Vieira LS, Baldoni AB, Aragão FJL. Suppression of SERK gene expression
affects fungus tolerance and somatic embryogenesis in transgenic lettuce. Plant Biol. 2008; doi:10.1111/j.1438-8677.2008.00103.x.
[17] Chisholm S, Coaker G, Day B, and Staskawicz B. Host–microbe interactions: Shaping the evolution of the Plant Immune Response. Cell, Volume 124. Issue 4, 2006; 803–81.
[18] Cueva A, Concia L, Cella R. Molecular characterization of a Cyrtochilum loxense Somatic
Embryogenesis Receptor-like Kinase (SERK) gene expressed during somatic embryogenesis. Plant Cell Rep, Volume 31, Issue 6, 2012; Pp. 1129 – 1139.
[19] Barnett, H y Hunter, B. Illustrated Genera of Imperfect Fungi. Cuarta edicion. The American Psychopathological Society. 1998; Pp. 188-189.
[20] Cullings KW. Molecular phylogeny of the Monotropoideae (Ericaceae) with a note on the placement of the Pyroloideae. J. Biol1994; Evol 7: 501-516.
[21] Walker, A.-S., Gautier, A., Confais, J., Martinho, D., Viaud, M., Le Pêcheur, P., Dupont, J., and Fournier, E. Botrytis pseudocinerea, a new cryptic species causing gray mold in French vineyards in
sympatry with Botrytis cinerea. 2011; Phytopathology 101:1433-1445.
[22] Oliviera R, Moral J, Bouhmidi K, Trapero A. Caracterización Morfológica y cultural de aislados de Colletotrichum spp. causantes de la Antracnosis del Olivo. Bol. San. Veg. Plagas, 2005; 31: 531-548 [23] Hou C y Yang C . Functional Analysis of FT and TFL1 Orthologs from Orchid (Oncidium Gower Ramsey) that Regulate the Vegetative to Reproductive Transition. Plant and Cell Physiology, Vol. 50, No. 8., pp. 1544-1557. 2009 ;doi:10.1093/pcp/pcp099
[24] Cueva Agila A.. Somatic Embryogenesis in Ecuador Orchids. Dottorato di Ricerca in Scienze Genetiche e Biomolecolari. Universita degli Studi di Pavia. 2010.
[25] Sutton, B. The genus Glomerella and its anamorph Colletotrichum. In: Bailey JA, Jeger MJ (Eds.)
Colletotrichum: Biology, Pathology and Control. Wallingford UK. CAB International. 1992; pp.1-26.
[26] Rodriguez, A. Caracterización molecular de poblaciones de Colletotrichum spp. Asociadas a Coffea
arabica en Colombia y su aplicación en el diagnóstico del CBD. Bogotá. Pontificia Universidad Javeriana.; 2007.
[27] Walker J., Nikandrow A y Millar, G.D . Species of Colletotrichum on Xanthium (Asteraceae) with
comments on some taxonomic and nomenclatural problem in the genus Colletotrichum. Micological Research 95, 1991; 1175 -1193
[28] Domsch, K., Gams, W and Anderson, T. Compedium Of soil Fungi. IHW –Verlang Eching. Second edition USA. 2007.
[29] Freeman, S., Katan, T. y Shabi, E. Characterization of Colletotrichum species responsible for Anthracnose diseases of various fruits. En: Plant Disease. 1998; 82(6): 596 - 604.
[30] Bailey, J. y Jeger, M . Colletotrichum biology, Pathology and control. Red Wood Press London, 1992; England p 388.
[31] Lee, S y Taylor, J. Isolation of DNA from Fungal mucelia and singles spores. Rn: Innis, M., Gelfand, D., Sninsky, J., White, T. PCR Protocols. A guide to metohds and applications. Academic Press. San Diego. USA. 1990; 482p.
[32] Ochoa D y Suárez, J.P. Caracterización molecular de 60 hongos que forman parte del cepario micológico del C.B.C.M. de la Universidad Técnica Particular de Loja. Tesis Bioquímico Farmacéutico. Universidad Técnica Particular de Loja. Escuela de Bioquímica y Farmacia. Loja, Ecuador. 2008.
[33] Sreenivasaprasad S , Sharada K, Brown A, y Mills P. Sequence-based detection of Colletotrichum
acutatum on strawberry. Plant Pathology. 1996; 45: 650-655
[34] Freeman S , Minz D., Jurkevitch E , Maymon M y Shabi E . Molecular analyses of Colletotrichum species from almond and other fruits. Phytopathology. 2000; 90:608-614.
[35] Sessa, G. and Martin, G. Protein Kinases in the Plant Defense Response. Advances in Botanical Research, 2000; 32:379-404.
[36] Vanoosthuyse, V., Tichtinsky, G., Dumas, C., Gaude, T. and Cock, J.M. Interaction of Calmodulin, a sorting nexin and kinase-associated protein phosphatase with the Brassica oleracea S locus receptor kinase. Plant Physiology, 2003; 133:919-929.
[37] Becraft PW. Receptor kinases in plant development. Trends Plant. 1998;Sci 3:384–388
[38] Santos MO, Romano E, Vieira LS, Baldoni AB, Aragão FJL. Suppression of SERK gene expression
affects fungus tolerance and somatic embryogenesis in transgenic lettuce. Plant Biol 2. 2009; 11:83–89. [39] Schmidt E, Guzzo F, Toonen M, de Vries S. A leucine-rich receptor-like kinase marks somatic plant
XX
[40] Hecht V, Velle-Calzada JP, Hartog MV, Schmidt EDL, Boutilier K, Grossniklaus U, de Vries SC . The Arabidopsis SOMATIC EMBRYOGENESIS RECEPTOR KINASE 1 gene is expressed in developing ovules and embryos and enhances embryogenic competence in culture. Plant Physiol, 2001; 127: 803–816
- 1 -
I.
Presentación de Fin,
- 2 -
I. Presentación de fin, propósito y componentes del Proyecto.
1.1 Fin del proyecto
Contribuir al conocimiento de las funciones del gen SERK en orquídeas.
1.2 Propósito del proyecto
Determinar la expresión del gen SERK de Cattleya maxima Lindl en respuesta al
ataque de hongos patógenos, Colletotrichum acutatum y Botrytis cinerea
1.3 Componentes del proyecto
Inoculación de hongos patógenos Colletotrichum acutatum y Botrytis cinerea en Cattleya maxima Lindl.
Patrón de expresión del gen SERK de Cattleya maxima Lindl, determinado por el
análisis qRT- PCR.
- 3 -
- 4 - II. INTRODUCCIÓN
2.1 ORQUÍDEAS
La familia Orchidaceae agrupa gran cantidad de especies del reino vegetal. Se estima
que existen 25.000 especies (1), agrupadas en alrededor de 900 géneros, además de
70.000 híbridos artificiales inscritos (2). Las orquídeas son plantas herbáceas,
perennes, cuyo tamaño oscila de 3,0 mm hasta varios metros de altura; presentan diversos hábitos de crecimiento debido a su gran variabilidad morfológica, crecen sobre los árboles (epífitas), rocas (litófitas) y en el suelo (terrestres), pero la mayor parte de la familia es epífita (3). Esta familia se encuentra prácticamente en todos los
continentes; sin embargo, su distribución no es uniforme, sino que se concentra en las regiones tropicales del planeta. Su distribución en el Ecuador va desde el trópico hasta el páramo, el país se considera como uno de los más diversos del planeta en cuanto a especies de esta familia (4).
2.1.1 Cattleya maxima Lindl
El nombre del género Cattleya se dio en honor al botánico y horticultor inglés
William Cattley, el cual cultivó las hojas hasta obtener floración, siendo la primera Cattleya en descubrirse (5). El género Cattleya agrupa orquídeas
originarias de zonas tropicales del Sur y Centro América, comprende algo más de 60 especies comúnmente epífitas y en pequeña parte litófitas, con pseudobulbos desde pocos centímetros hasta un metro, que llevan en su extremo una o dos hojas sin pecíolo (6), las raíces presentan una membrana
esponjosa que las protege de la desecación y sirven también para absorber agua y minerales (7).
Cattleya maxima es una orquídea epífita que se distribuye
en Colombia, Ecuador y en el norte del Perú. En Ecuador es más abundante su distribución geográfica en las provincias de El Oro, Guayas, Loja, Santa Elena,
1
Pupulin, F (2005). Conservación de orquídeas: Una guía para autoridades administrativas. San José, Costa Rica, Editorial SEIDIN. Pp. 37
2
Rivera, G (1998). Orquídeas: Generalidades y cultivo. Heredia, Costa Rica, Editorial Fundación UNA.Pp.266
3
Ortiz-Arias, B (2002). Identificación de tres especies de poty virus en orquídeas nativas (en línea). Tesis de Bach. de Ing. Biotecnológica presentada en la Escuela de Biología. Cartago, CR, TEC. 69 p .
4 Dodson, C. H., Escobar R (2003). Orquídeas Nativas del Ecuador. Oncidium-Restrepiopsis. Editorial
Colina, Medellín. Volumen IV.
5
Horich, C (1980). Las Cattleyas en Costa Rica. En: Orquídeas: su cultivo en Costa Rica. Ed. por la Asociación Costarricense de Orquideología. San José, Costa Rica, Impresora Delta, S.A. Pp. 9-11.
6
Kuan, C y González, L (1993). Introduccion al cultivo y manejo de las orquideas. San José, Costa Rica. Instituto Nacional de Aprendizaje. Pp 1 – 16, 47 -53, 75 -76.
7
- 5 -
Manabí y Los Ríos (8).Con el objetivo de incrementar las tasas de multiplicación
de orquídeas en menor tiempo, en la actualidad se utiliza la técnica de cultivo
in vitro.
Figura 1. Planta de Cattleya maxima Lindl Fuente:(9)
Orquidario Virtual
2.1.2 Cultivo in vitro de orquídeas
Las técnicas de cultivo in vitro son una importante herramienta para la
germinación de orquídeas.
Las semillas de las orquídeas no pueden germinar en un sustrato común o un invernadero debido a que no poseen endosperma que provea nutrientes al embrión para su germinación, por ello en la naturaleza se asocian a hongos micorrízicos que le proporcionan nutrientes. En condiciones in vitro el medio
de cultivo con sales minerales, vitaminas, sacarosa y reguladores de crecimiento, posibilita la germinación (10,11)
2.2 Hongos
Los hongos son probablemente los organismos más abundantes del planeta y forman parte de ciclos vitales. Se estima que existen aproximadamente 1.5 millones de especies en todo el planeta, de éstos son conocidos únicamente 80000, los hongos pueden ser microscópicos o macroscópicos, eucarióticos,
8 Dodson, C., Luerand, C y Gustavo L (2004). Orchids of Ecuador. Soluciones Graficas D Yg. Quito –
Ecuador.
9 Orquidario Virtual (http://www.orquidariovirtual.com/cattleya-maxima/) 10 Arditti, J y R. Ernst (1993). Micropropagation of Orchids. Canada. 11
[image:25.595.193.461.159.341.2]- 6 -
ramificados, a menudo filamentosos, carecen de clorofila y tienen una pared celular que contiene quinina, celulosa o ambos componentes (12).
2.3 Enfermedades producidas por hongos
En las orquídeas el desarrollo de hongos patógenos como Colletotrichum
acutatumy Botrytis cinerea es favorecido por tejidos débiles y presencia de heridas
en las plantas. Los hongos son uno de los organismos que mayores problemas fitosanitarios causan en este tipo de cultivo (13). Los síntomas más frecuentes que
presentan las orquídeas atacadas por hongos son manchas marrones en hojas y bulbos, también pueden afectar a las raíces causando la pudrición de las mismas
(14)
La efectividad de un hongo patógeno de producir enfermedad depende en gran parte de las condiciones en que se desarrolla e inocula. Se deben controlar las condiciones de crecimiento tomando en cuenta el medio de cultivo, su volumen, aireación, luz, temperatura. Las cepas de los patógenos necesitan multiplicarse para inocular a los hospederos, ya que éstas solo infectan por medio de sus esporas (15)
2.3.1 Colletotrichum acutatum
Es causante de la enfermedad conocida como antracnosis, produce graves lesiones en plantas y frutos en su estado de madurez, es un patógeno que puede estar latente dentro de los tejidos del huésped durante un corto o largo periodo de tiempo. Una vez que el micelio se ha desarrollado suficientemente dentro de la planta, se producen cuerpos fructíferos, causando los síntomas típicos (16,17,18). Inicialmente las hojas afectadas presentan manchas foliares
ovaladas, circulares o de forma irregular, color café oscuro, negras o grisáceas. Después de haberse depositado en la superficie de la planta, los conidios y
12
Piepenbring, M (2006). Inventoring the fungi of Panama. Springer Science. 16: 73 - 84
13
Bustos, T (2006). Ecuador Patria de Orquídeas. Universidad Técnica Particular de Loja. Loja –Ecuador
14 Rollke, F (2006). Orquideas. Hispana Europea. Barcelona – España 15
Hoog, G., Guarro, J., Gene, J and Figueroas, M (2000). Atlas of Clinical Fungi. Central albulreau Voor Schimmelcultures Utrecht. The Netherlands. Second edition.
16
Mordue, J.E.M (1979). Colletotrichum acutatum. CMI Descriptions of Pathogenic Fungi and Bacteria No. 630. CAB International, Wallingford, UK.
17
Baxter, A.P.; Westhuizen, G.C.A. van der; Eicker, A (1983). Morphology and taxonomy of South African isolates of Colletotrichum. South African Journal of Botany 2, 259-289.
18
- 7 -
ascosporas dan lugar a la germinación, proceso fundamental para la infección
(19).
2.3.1.1 Caracteristicas morfológicas
Las colonias en cultivo son generalmente de color blanco, gris pálido o anaranjado pálido, a veces produciendo pigmentos de color rosado-púrpura. Las células conidiógenas son cilíndricas, fusiformes, de paredes delgadas, aseptadas e hialinas, con apresorios claviforme a circular y claro a oscuro-marrón (17, 18,19,20)
2.3.1.2 Taxonomía
Colletotrichum acutatum pertenece al reino Fungi; Hongos Mitospóricos
(División Eumycota, Subdivisión Deuteromycotina, Clase Coelomycetes); género Glomerella; especie Glomerella acutata (fase teleomórfica, sexual o perfecta) o Colletotrichum acutatum (fase anamórfica, asexual o imperfecta) (21)
2.3.2 Botrytis cinerea
Responsable de la enfermedad conocida como podredumbre gris, es un patógeno que se implanta una vez que se ha producido un daño sobre el tejido, las conidias son liberadas desde los conidióforos hasta llegar al hospedero para comenzar el proceso de infección. El hongo sobrevive en el suelo (esclerocios) y en plantas muertas donde crece como micelio, la incidencia de la enfermedad aumenta cuando hay periodos prolongados de humedad y temperaturas bajas de 15 a 20°C (22) .Los síntomas de esta enfermedad son
variables dependiendo de la planta y del órgano que se vea afectado, pero en general, se producen podredumbres blandas, y se puede observar un característico moho de color grisáceo, especialmente en frutos; en hojas se puede observar una necrosis alrededor del punto de entrada pudiendo avanzar al resto de la hoja si las condiciones son favorables para el hongo (23)
19 Agrios, G (2000). Fitopatología. Ed. Limusa, S.A. México, D.F. Segunda edición 838p. 20
Sutton, B.C (1980). The Coelomycetes. Fungi Imperfecti with pycnidia, acervuli and stromata, 696 pp. Commonwealth Mycological Institute, Kew, UK.
21 Barrau, C, de los Santos, B. y Romero, F (2001). First report of Colletotrichum acutatum in blueberry
plants in Spain. Plant Disease 85: 1285
22Latorre, BA; Agostín, E; San Martín, R; Vázquez, GS (1997). Effectiveness of conidia of Trichoderma
harzianum produced by liquid fermentation against Botrytis bunch rot of table grape in Chile. Crop Protection 16: 209- 214.
23
- 8 - 2.3.2.1 Caracteristicas morfológicas
Botrytis cinerea, posee conidios hialinos con forma semejante a un huevo,
formado en ramas de conidióforos sobre la superficie. La organización de las esporas en forma de racimos da el nombre a éste género, en griego Botrytis
significa grupo de uvas. Forma esclerocios lisos de color negro en forma de barra o hemisferio debajo de la cutícula o la epidermis del huésped y se aferran firmemente a esta. Las especies de Botrytis causan comúnmente moho gris. En
hojas las lesiones se presentan con sintomas característicos de necrosis, pueden extenderse a través de ésta hasta debilitarlo y ocasionar quebramiento
(24)
2.3.2.2 Taxonomía
Botrytis cinerea pertenece al reino Fungi, filo Ascomycota, subfilo
Pezizomycotina, clase Leotiomycetes, orden Helotiales, familia Sclerotiniaceae,
género Botryotinia y especie Botryotinia fuckeliana (fase teleomórfica, sexual o
perfecta) o Botrytis cinerea (fase anamórfica, asexual o imperfecta)(25)
2.4 Receptor de Embriogénesis Somática tipo quinasa (SERK)
Muchos receptores quinasas (RLKs) juegan un importante papel en el metabolismo celular que conduce al crecimiento y respuestas de defensa para activar un gran
número de expresión de genes. El gen SERK (Somatic Embryogenesis Receptor like
Kinase) fue aislado por primera vez a partir de células embriogénicas de zanahoria, resultando ser un marcador molecular para la embriogénesis somática ( 26), durante los
últimos años se ha asociado la función de este gen mediando la transducción de señales que conducen a la defensa y resistencia a enfermedades (27).Se han
identificado cinco miembros de la familia SERK en Arabidopsis (SERK1, SERK2, SERK3, SERK4, SERK5) (28) .Estudios recientes han indicado que la supresión de la
24
Holz, G., Coertze, S. & Willianson, B (2004). The ecology of Botrytis on plant surfaces. Y. Elad et al. (eds.), Botrtytis: Biology, Pathology and Control. Kluwer Academic Publishers. Netherlands. Pp, 9-27.
25 Fernandez, F (2006). Aplicación de la proteómica a la caracterización de los mecanismos de
patogenicidad de Botrytis cinerea. Utilización y evaluación de nuevos fungicidas. Tesis Doctoral, Departamento de Bioquímica y Biología Molecular, Facultad de Ciencias del Mar y Ambientales. Universidad de Cádiz, España.
26 Schmidt ED, Guzzo F, Toonen MA, de Vries SC (1997). A leucine-rich repeat containing receptor-like
kinase marks somatic plant cells competent to form embryos. Development. 1997; 124:2049–2062
27 Hu H, Xiong L, Yang Y (2005) Rice SERK1 gene positively regulates somatic embryogenesis of cultured
cell and host defense response against fungal infection. Planta 222:107–1
28 Shiu S, Bleecker A (2001). Receptor-like kinases from Arabidopsis form omonophilitic gene family
- 9 -
expresión del gen SERK aumenta la susceptibilidad al ataque de agentes patógenos
en cacao (29).
En arroz la inoculación de agentes patógenos y moléculas de defensa de señalización como el ácido salicílico y el ácido abcísico, inducen sobre la expresión de OsSERK1
llevando a un aumento en la resistencia del huésped y por ende la supresión del hongo (23)
Una serie de genes SERK han sido identificados hasta el momento en
monocotiledóneas y dicotiledóneas, incluyendo 5 genes en Arabidopsis, AtSERK1-5 (30), 3 en
Zea mays ZmSERK1–3 (31), 2 en Oriza sativa OsSERK1-2 (32), y seis en Medicago truncatulaMtSERK1- 6(33).
A través de estudios moleculares se ha demostrado que los genes de resistencia del huésped codifican componentes del sistema inmune que confieren la capacidad para reconocer y responder a patógenos, mientras que dichos patógenos han desarrollado proteínas efectoras que pueden suprimir respuestas inmunes del huésped. Además, los efectores pueden por sí mismos actuar como elicitores y pueden ser deshabilitados por el huésped (34)
En la cara externa de la célula huésped, ésta conserva elicitores microbianos llamados patrones moleculares asociados a patógenos (PAMP), que son reconocidos por proteínas llamadas receptores de reconocimiento de patrones (PRRs) (35). La
estimulación de PRRs conduce a la inmunidad activada por PAMP (PTI). A pesar de que diversas PAMP activan PTI a través de receptores diferentes, las respuestas intracelulares resultantes se superponen ampliamente. En plantas, el PRR mejor estudiado es el FLS2 (flagellin sensitive 2), y es el responsable del reconocimiento de la flagelina en Arabidopsis thaliana, compuesto por un ectodominio de repeticiones
ricas en Leu, un dominio transmembrana y otro unido a proteínas quinasas mediante
29 Santos MO, Romano E, Vieira LS, Baldoni AB, Aragão FJL (2008) Suppression of
SERK gene expression affects fungus tolerance and somatic embryogenesis in transgenic lettuce. Plant Biol. doi:10.1111/j.1438-8677.2008.00103.x
30 Hecht V, Vielle-Calzada JP, Hartog MV, Schmidt EDL, Boutilier K, Grossniklaus U, de Vries SC (2001).
The Arabidopsis Somatic Embryogenesis Receptor Kinase 1 Gene is Expressed in Developing Ovules and Embryos and Enhances Embryogenic . Competence in Culture. Plant Physiol 127:803–816
31
Baudino S, Hansen S, Brettschneider R, Hecht VFG, Dresselhaus T, Lorz H, Dumas C, Rogowsky PM (2001). Molecular characterization of novel maize LRR receptor-like kinases, which belong to the SERK gene family. Planta 213:1–10
32
Ito Y, Takaya K, Kurata N (2005). Expression of SERK family receptor-like protein kinase genes in rice. Biochim Biophys Acta 1730 :253–258
33
Nolan KE, Irwanto RR, Rose RJ (2011). Auxin up-regulates MtSERK1 Expression in Both Medicago truncatula Root-Forming and Embryogenic Cultures. Plant Physiol 133:218–230
34
Chisholm S, Coaker G, Day B, and Staskawicz B (2006) Host–microbe interactions: Shaping the evolution of the Plant Immune Response. Cell, Volume 124, Issue 4, 803–814
35
- 10 -
una unión Ser-Thr (36) El reconocimiento de flg22 por su receptor FLS2 en
A. thaliana,
causa formación de callos, una inhibición fuerte del crecimiento celular, y la expresión de los genes de defensa (37). En los últimos años se ha identificado el gen
SERK3
asociado a BAK1 (brassinosteroid - insesitive 1), como un factor implicado en la PTI. La interacción SERK3/BAK1 se requiere para las respuestas tempranas a PAMPs y
para incorporar rápidamente el complejo flg22 inducido. Por ello, se especula que en respuestas a PAMPs, SERK3/BAK1 actúa de forma semejante a su papel en la
señalización del BR (brasinoesteroide) asociado al receptor BRI1. La implicación de
SERK3/BAK1 en PTI proporciona evidencia de concordancias entre la inmunidad de
la planta y la señalización del BR(39) . Por lo tanto,
SERK3/BAK1 parece integrar
diversos eventos de percepción en las respuestas PAMP, dando lugar a la inmunidad frente a una amplia gama de microrganismos invasores(40).
Recientemente se ha realizado la caracterización del gen SERK de dos especies de
orquídeas distribuidas en el Sur del país: Cyrtochilum loxense (38) y Cattleya maxima
(Cueva, datos sin publicar). Sin embargo la expresión del gen SERK ha sido estudiada
únicamente durante el proceso de embriogénesis somática. Estudios sobre el papel de este gen en respuesta a patógenos no se han realizado en ninguna especie de orquídea hasta el momento. Los objetivos planteados para la presente investigación fueron dirigidos a: (1) Aislar hongos patógenos en medio de cultivo PDA (Potato Dextrose Agar), (2) identificación morfológica y molecular, (3) inoculación de hongos en plantas in vitro de Cattleya maxima Lindl, (4) estudiar la expresión del gen SERK
mediante PCR de tiempo real en respuesta al ataque de hongos patógenos
Colletotrichum acutatum y Botrytis cinerea, con el fin de conocer la funcionalidad del
gen en otros procesos vegetales.
2.5 PCR en Tiempo Real
También llamada PCR cuantitativa en tiempo real, está basada en la reacción en cadena de polimerasa (PCR) y se usa para amplificar y al mismo tiempo cuantificar
36
Heese A, Hann DR, Giménez-Ibanez S, Jones AME, He K, Li J, Schroeder JI, Peck SC y Rathjen JP (2007) The receptor-like kinase SERK3/BAK1 is a central regulator of innate immunity in plants. Plant Biology 104: 12217-12222
37
Ryan CA, Huffaker A and Yamaguchi Y (2007) New insights into innate immunity in Arabidopsis. Cellular Microorganismology 9: 1902-1908
38
- 11 -
moléculas de ADN o ADN complementario (cDNA) específicas y así acceder a datos fiables y precisos sobre la expresión genética de las células en estudio (39).
En la PCR de tiempo real, los procesos de amplificación y detección se producen de manera simultánea en el mismo vial cerrado, sin necesidad de ninguna acción posterior. Además, mediante detección por fluorescencia se puede medir durante la amplificación la cantidad de ADN sintetizado en cada momento, ya que la emisión de fluorescencia producida en la reacción es proporcional a la cantidad de ADN formado. Esto permite conocer y registrar en todo momento la cinética de la reacción de amplificación.
Los termocicladores para llevar a cabo la PCR en tiempo real incorporan un lector de fluorescencia y están diseñados para poder medir, en cualquier momento, la fluorescencia emitida en cada uno de los viales donde se realice la amplificación. Los sistemas de detección por fluorescencia empleados en la PCR de tiempo real pueden ser agentes intercalantes, que son fluorocromos que aumentan notablemente la emisión de fluorescencia cuando se unen a ADN de doble hélice. El más empleado en PCR de tiempo real es el SYBR Green I (40).
39
Heid,C.A., Stevens,J., Livak,K.J. & Williams,P.M (1996). Real time quantitative PCR. Genome Research 6: 986-994
40 Higuchi R, Fokler C, Dollinger G, Watson R. Kinetic PCR analysis: Real-time monitoring of DNA
- 12 -
- 13 - III. MATERIALES Y MÉTODOS
3.1 Material biológico
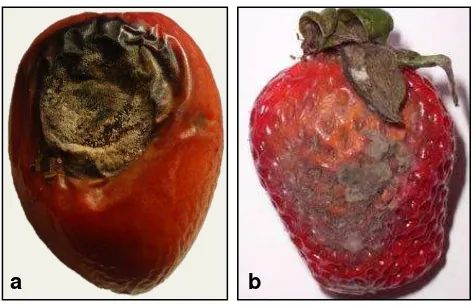
Para este estudio se obtuvieron muestras de tomate de árbol (Solanum betaceum) y fresa (Fragaria vesca) con síntomas de Colletotrichum acutatum y Botrytis cinerea
respectivamente (Figura. 2), éstas fueron recogidas en el barrio Zalapa ubicado al Norte de la ciudad de Loja.
Una vez recolectadas las muestras, fueron llevadas al laboratorio del Departamento de Ciencias Agropecuarias y Alimentos de la Universidad Técnica Particular de Loja para su aislamiento y posterior identificación.
a b
3.2 Aislamiento de Colletotrichum acutatum y Botrytis cinerea de material
vegetal enfermo.
Se preparó inicialmente una solución de 39 g/l de medio agar papa dextrosa (PDA), medio utilizado para el desarrollo de hongos, y se esterilizó a una temperatura de 121°C y 80 mmHg de presión por 20 minutos.
Dentro de la cámara de flujo, el medio fue dispensado colocando aproximadamente 10 ml en placas Petri estériles (9cm diámetro), se dejó reposar en la cámara hasta que solidifique el medio.
Se desinfectó la superficie de la fruta con alcohol al 70%, luego se realizó un raspado superficial con un asa de transferencia punta redonda esteril sobre el tejido vegetal con síntomas de la enfermedad, el cual fue sembrado directamente en placas Petri que contenían medio PDA, éstas fueron etiquetadas, selladas con parafilm e
[image:33.595.197.435.310.464.2]- 14 -
incubadas en la oscuridad en una posición invertida a 25°C por un lapso de diez días.
Con la finalidad de establecer un cultivo más confiable se realizaron aislamientos monospóricos que garantizan la autenticidad y pureza de los hongos.
3.2.1 Obtención de cultivos monospóricos
Se disolvió una alícuota de micelio con abundante esporulación en un tubo eppendorf con 2ml de agua destilada estéril, se agitó ligeramente por 40 segundos, obteniéndose una suspensión de esporas.
En placas Petri con medio PDA se dispensó la suspensión, cada aislamiento fue incubado a una temperatura de 25°C durante 24h.
Al día siguiente se procedió a seleccionar un conidio individual germinado y se transfirió a una nueva placa Petri con medio PDA (41).
3.3 Identificación morfológica
Las características macroscópicas como la forma y el color de los cultivos fueron revisadas después de diez días de incubación. La pureza del cultivo se comprobó mediante visualización microscópica, utilizando el microscopio óptico Zeiss (modelo Axiostar plus) con los objetivos 10X y 40X, se determinó la forma de las conidias según lo propuesto por Barnett y Hunter (42).
Los patógenos se replicaron en siete placas Petri cada uno, con la finalidad de conservar más cepas, éstas fueron refrigeradas a 4 °C.
3.4 Identificación molecular
3.4.1 Extracción ADN
De cada cultivo monospórico se realizó la extracción de ADN utilizando una porción micelial evitando llevar demasiado agar. Se empleó el kit de extracción DNeasy Plant Mini Kit de Qiagen ®, siguiendo el protocolo del fabricante
(Anexo1).
41
Falconi, C (1998). Fitopatología Práctica, Facultad de Ciencias Agropecuarias, Escuela Politécnica del Ejército. Primera Edición. Sangolqui, Ecuador.
42
- 15 - 3.4.2 Amplificación de ADN mediante PCR
Este proceso se llevó a cabo después de haber confirmado la existencia de ADN de buena calidad (mediante electroforesis y cuantificación en el Nanodrop 2000). En la tabla 1 se indica la preparación de la mezcla para la PCR.
Tabla. 1. Preparación de soluciones para amplificar ADN mediante PCR
REACTIVO MIX 1
Phusion polimerasa 10 μl
H2Odde 6,4 μl
Cebador Fw 0,4 μl
Cebador Rw 0,4 μl
BSA 0,8 μl
ADN 2 μl
TOTAL 18 μl
Se utilizaron primers universales para Colletotrichum acutatum y Botrytis
cinerea , se amplificó un segmento de la región intergénica con el cebador
universal ITS1 Fw (5'-TCC GTA GGT GAA CCT GCG G-3 '), el cual fue
combinado con el cebador ITS4 Rv (5’-TCC TCC GCT TAT TGA TAT GC -3’)
(43), además se utilizaron cebadores específicos para
Botrytis pseuocinerea, el
gen MS547 (5'-AAG GAG GAC GT TGGA AGG AT-3') y (5'-AAG TCC AGA
ATC TCG ATG TAT TTG T-3 ') (44), una especie de
Botrytis con características
morfológicas similares a la estudiada en nuestra investigación. MS547 codifica la ATP-dependiente del RNA helicasa DBP7 (referencia de genes en el genoma de B. cinerea B0510: BC1G_03202.1)
Las muestras se llevaron al termociclador Applied Biosystem programado de la siguiente manera para Colletotrichum acutatum: Desnaturalización inicial
98°C por 30s, seguida de 30 ciclos, cada ciclo consta de un paso de desnaturalización a 98°C por 10 s, anillamiento de primers 60°C por 20 s,
43
White, T. J., Bruns, T., Lee, S. & J. Taylor (1990). Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. Pp. 315-322. En M. Innis, D. Gelfand, J. Snisky & T. White (eds). PCR protocols: a guide to methods and applications. San Diego: Academic Press.
44 Walker, A.-S., Gautier, A., Confais, J., Martinho, D., Viaud, M., Le Pêcheur, P., Dupont, J., and Fournier,
- 16 -
extensión a 72°C por 30 s, una extensión final a 72°C por 7min y con una temperatura final de enfriamiento de 4°C.
En el caso de Botrytis cinerea se programó de la siguiente manera
Desnaturalización inicial 95°C por 5min, seguida de 40 ciclos, cada ciclo consta de un paso de desnaturalización a 95°C por 30 s, anillamiento de primers 62°C por 30 s, extensión a 72°C por 90 s, una extensión final a 72°C por 5min y con una temperatura final de enfriamiento de 4°C.
Los productos de PCR se verificaron por medio de electroforesis (128 V, 300 mA, 20 min) en gel de agarosa al 1%, se colocó en el primer pocillo del gel 1,5
μl de marcador molecular de 100pb y en el resto de pocillos se cargo 2 μl de cada producto de PCR combinado con 1.5 μl de azul de bromofenol y se realizó la observación en el transiluminador UV.
3.4.3 Cloning y Purificación de productos
Para realizar el cloning de los fragmentos de ADN obtenidos por PCR, se utilizó cepas bacterianas E.coli TOP10, medio solido LB (Bacto - Agar) con Kanamicina en cantidad 100mg/ μl, vector Zero Blunt TOPO PCR y el kit de
Cloning (Invitrogen) siguiendo el protocolo modificado por el grupo de micorrizas del departamento de ciencias naturales (Anexo 2).
La transformación de E. coli se realizó mediante shock térmico, utilizando 2 μl
de la mix de cloning, sometiéndose a 42°C durante 1 min. Posteriormente, las colonias sembradas y crecidas en la placa Petri se dejaron crecer en un medio de cultivo líquido LB con antibiótico durante 12 horas, para la posterior purificación , para lo cual se utilizó S.N.A.P.™ MidiPrep Kit (Invitrogen) siguiendo el protocolo del fabricante (Anexo 3).
3.4.4 Digestión enzimática
Se realizó un corte en la cadena del ADN utilizando la enzima de restricción EcoR1, (Tabla 2) incubando a 37°C durante 3 horas, luego se realizó una electroforesis en gel de agarosa al 1% para la observación del producto
.
- 17 -
Tabla. 2. Preparación de soluciones para digestión enzimática
REACTIVO MIX 1
ADN plasmídico 2,5 μl
Enzima EcoR1 0,5 μl
Buffer 0,5 μl
H2Odde 1,5 μl
TOTAL 5 μl
3.4.5 Secuenciación
Con el fin de confirmar la identidad de las especies encontradas, se recurrió al uso de técnicas de secuenciación las cuales se llevaron a cabo en un volumen total de 16 μl, cada tubo conteniendo: 1 μl Big dye® Terminador del ciclo de la secuenciación, 1 μl Cebador de Secuenciación, 2 μl de ADN plasmídico, 2 μl
de 5x Buffer y 10 μl de Agua destilada desionizada estéril. Las reacciones de PCR se realizaron en un termociclador 3500 Genetic Analyzer Applied Biosystem. El termociclador fue programado para realizar: desnaturalización de 96oC durante 1 minuto, 50 ciclos de 96oC por 10 segundos, la temperatura
apropiada de anillado 60oC por 4 minutos.
La purificación de la secuenciación luego de la amplificación fue realizada usando el kit BigDye XTerminator, para lo cual se utilizó: 20μl de muestra de solución SAM y 5μl del ADN amplificado, luego se aplica vortex por 30 minutos. La muestra se centrifugo por 2 minutos y 10μl del sobrenadante se usó para correr la secuencia de análisis.
Se utilizó primers universales M13-Fw (5’TGT AAA ACG ACG GCC AGT3’) y
M13-Rv ( 5’TGC CAG GAA ACA GCT ATG AC3’), para la secuencia de
nucleótidos clonados anteriormente.
3.4.5.1 Análisis de secuenciación