1
UNIVERSIDAD TÉCNICA PARTICULAR DE LOJA
La Universidad Católica De Loja
ÁREA BIOLÓGICA
TITULACIÓN DE BIÓLOGO
Anatomía foliar de tres especies de Bromeliaceae como componentes de la
dieta del oso andino (Tremarctos ornatus) en el Parque Nacional Podocarpus.
TRABAJO DE FIN DE TITULACIÓN
AUTORA:
Vallejo Ochoa, Natalia Salomé
DIRECTOR:
Moreira Palacios, Máximo Oswaldo, Blgo
LOJA - ECUADOR
II
APROBACIÓN DEL DIRECTOR DEL TRABAJO DE FIN DE TITULACIÓN
Biólogo.
Máximo Oswaldo Moreira Palacios.
DOCENTE DE LA TITULACIÓN
De mi consideración:
El presente trabajo de fin de titulación: Anatomía foliar de tres especies de Bromeliaceae como componentes de la dieta del oso andino (Tremarctos ornatus) en el Parque
Nacional Podocarpus realizado por Vallejo Ochoa Natalia Salomé ha sido orientado y revisado durante su ejecución, por cuanto se aprueba la presentacióndel mismo.
Loja, octubre de 2014
III
DECLARACIÓN DE AUTORÍA Y CESIÓN DE DERECHOS
Yo Natalia Salomé Vallejo Ochoa, declaro ser la autora del presente trabajo de fin de titulación:
Anatomía foliar de tres especies de Bromeliaceae como componentes de la dieta del oso andino (Tremarctos ornatus) en el Parque Nacional Podocarpus de la titulación de
Biología siendo el Biólogo. Máximo Moreira Palacios director del presente trabajo; y eximo expresamente a la Universidad Técnica Particular de Loja y a sus representantes legales de posibles reclamos o acciones legales. Adicionalmente declaro conocer y aceptar la disposición del Art. 67 del Estatuto Orgánico de la Universidad Técnica Particular de Loja que en su parte pertinente textualmente dice: “Forman parte del patrimonio de la Universidad la propiedad intelectual de investigaciones, trabajos científicos o técnicos y tesis de grado que se realicen a través, o con el apoyo financiero, académico o institucional (operativo) de la Universidad”
f.……….
IV
DEDICATORIA
La presente investigación va dedicado a mis padres María Teresa y Ángel Isidro (+), ellos han sido un pilar fundamental en este caminar, mi guía, mi cimiento, mis raíces en todos estos años de vida; su apoyo incondicional, sus sabios consejos y enseñanzas me han dado fuerzas para llegar culminar esta etapa y continuar las restantes aventuras que me faltan por vivir, hoy mi Ángel me cuida desde el cielo iluminando mi camino y mi alma, Papá te extraño y te amo, gracias.
A mis hermanas; Paola gracias por tu amor, tu protección, tus consejos que han sido fortalezas para continuar; a mi hermana Gabriela que aunque a la distancia siempre me ha brindado su apoyo y amor, a mi hermano Nikolay por estar apoyándome siempre, por tu paciencia, por estar conmigo en cada momento importante de mi vida, han sido, es y serán mi ejemplo a seguir.
A mis abuelitos Dorita y Olmedito (+) por consentirme y darme su cariño, sus pláticas me han brindado mucho amor, las bendiciones de cada noche inundaron mi alma de paz.
A mi pequeña inspiración Mateo Nikolas por ser mi alegría, ver cómo va cursando sus primeros años de vida me inundan de alegría y me dan endereza para ser cada vez una mejor tía.
V
AGRADECIMEINTO
Expreso un cordial agradecimiento a la Universidad Técnica Particular de Loja; por haberme permitido ser parte de su alumnado y culminar con éxito mi carrera profesional.
A cada uno de mis profesores de la UTPL por brindarme sus enseñanzas y transmitirme sus conocimientos.
Un agradecimiento especial al Biólogo Máximo Moreira, por su acertada dirección de la tesis y constate guía, aspectos que ayudaron a culminar el proyecto de tesis..
A la Dra. Lorena Guevara; gracias por brindarme su tiempo, su espacio, y sus conocimientos.
Agradezco al Dr. Zhofre Aguirre catedrático de la Universidad Nacional de Loja, y de igual manera al Ing. Bolívar Merino por su asesoría en la identificación de especies, y a Luis Marcelo por su apoyo y colaboración en el trabajo de campo.
A mi maestro Carlos Ortega por el apoyo brindado para poder culminar con éxito este proyecto.
A todas y cada una de las personas que me brindaron su ayuda reciban un agradecimiento personal de mi parte.
1
INDICE DE CONTENIDOS
CERTIFICACIÓN ... II DECLARACIÓN DE AUTORÍA Y CESIÓN DE DERECHOS ... III DEDICATORIA ... IV AGRADECIMEINTO ... V
RESUMEN EJECUTIVO ... 2
ABSTRACT ... 3
INTRODUCCIÓN ... 4
CAPÍTULO 1. MARCO TEÓRICO ... 6
1.1 La familia Bromeliaceae ... 7
1.2 Genero Puya ... 8
1.3 Algunos estudios anatómicos en Bromeliaceae ... 9
1.4 Utilidad de las técnicas histológicas o microhistológicas ... 10
1.5 El Parque Nacional Podocarpus (PNP) y su especie bandera: el oso de anteojos ... 11
1.6. OBJETIVOS ... 12
CAPÍTULO 2. MATERIALES Y MÉTODOS ... 13
2.1 Selección de especies ... 14
2.2 Sitios de colección de especímenes ... 15
2.3 Colección de muestras y especímenes (muestreo) ... 15
2.4. Evaluación de las estructuras morfológicas de las hojas de bromelias. ... 16
2.5 Metodología para diseñar una herramienta para la identificación de bromelias en estudios de dieta de herbívoros ... 18
CAPÍTULO 3. RESULTADOS ... 19
3.1. Caracterización anatómica de Puya eryngioides ... 19
3.2 Caracterización anatómica de Puya maculata ... 26
3.3 Caracterización anatómica de Puya nítida ... 33
CAPÍTULO 4 DISCUSIÓN ... 44
4.1. Aspectos metodológicos para estudios morfoanatómicos ... 45
4.2. Implicaciones de los estudios morfológicos ... 45
CONCLUSIONES ... 49
RECOMENDACIONES ... 50
2
RESUMEN EJECUTIVO
El oso andinoTremarctos ornatus, es una especie clave para la conservación de ecosistemas
de altoandinos; por ello, se estudió las estructuras anatómicas de Puya eringioides, Puya
maculata y Puya nítida. Se colectaron 10 hojas por especie en el Parque Nacional
Podocarpus; luego fueron fijadas en FAA, y se realizaron cortes a mano alzada, macerado y raspado de la epidermis. Se encontró que la epidermis foliar de las tres especies están compuestas por células rectangulares, casi rectas de paredes anticlinales sinuosas, y engrosadas; los estomas están ubicados al mismo nivel que las células epidérmicas; las escamas en las tres especies se distribuyen en hileras longitudinales, pero en P. eryngiodes
y P. nítida están formadas por escudos y alas; el parénquima acuífero está muy desarrollado
y la forma de sus células es variable; el clorénquima se ubica en la zona inferior contiguo a la hipodermis de la superficie abaxial, alternando con parénquima esponjoso en P. eringiodes y
en cámaras ocupadas por células aerenquimáticas irregulares en P. maculata. Los haces
vasculares presentan vainas fibrosas conectados entre sí.
3
ABSTRACT
The Andean bear Tremarctos ornatus, is a key species for conservation of paramos and
andean forests ecosystem. Therefore, we studied the anatomic structure of the Puya
eryngioides, Puya maculata and Puya nitida, which constitute the main diet of the Andean bear
in the Podocarpus National Park (PNP). Ten leaves of each species of paramos of PNP were collected and after in the laboratory each leaf was divided into middle and basal part; and were fixed in FAA (formaldehyde 5ml, 5ml acetic acid and ethyl alcohol 70% 90ml), and cutting, assembly and analysis was done; the technique used was cut freehand, marinated and hand scraping the epidermis. The leaf epidermis of the three species are composed of rectangular cells, nearly straight anticlinal walls sinuous, thickened. The scales (trichomes) in the three species is distributed in longitudinal rows and consist of shields and wings in P. eryngiodes
and P. nitida; its presence is related to the absorption of water and nutrients, and reducing
perspiration normal basic qualities of the vegetable species develops.
4
INTRODUCCIÓN
En la presente investigación se estudió las características morfo-anatómicas de tres especies de bromeliáceas pertenecientes al género Puya (Puya eryngioides, Puya maculata y Puya
nítida), las cuales están distribuidas en el ecosistema de páramo del Parque Nacional
Podocarpus; estas bromelias son parte fundamental de la dieta del oso de anteojos
(Tremarctos ornatus), considerada una especie clave para la conservación de los páramos y
los bosques nublados del Ecuador.
Esta investigación está dividido en cinco capítulos: marco teórico, materiales y métodos, resultados, discusión, y conclusiones. En el marco teórico se presenta una descripción botánica de la familia y del género Puya, se analiza las principales estructuras anatómicas del
género; y se argumenta la importancia de este género en la dieta de una de las especies símbolo de los ecosistemas alto andinos como lo es el oso de anteojos. En el segundo capítulo se habla de los materiales y métodos utilizados para realizar la investigación, y se describe el procedimiento metodológico tanto para la colecta del material en campo (hojas de las tres especies de Puya) como del procedimiento en laboratorio que se utilizó para el análisis anatómico de las estructuras celulares de cada una de las especies estudiadas. El capítulo tres sintetiza los principales resultados obtenidos en este estudio, donde se presentan las características del sistema epidérmico, el sistema fundamental y el sistema vascular de cada una de las especies. El capítulo cuatro se hace una discusión de estos resultados. Finalmente, se presentan las principales conclusiones a las cuales se ha llegado en este estudio, así como se plantean algunas recomendaciones relacionadas con aspectos técnicos, metodológicos y de gestión de la información que deberían considerarse.
A más de ampliar el conocimiento anatómico de las tres especies del género Puya antes
mencionadas; también se generó los insumos necesarios que permitieron construir una clave
de identificación anatómica para cada una de las tres especies de bromelias; clave que constituye una herramienta metodológica que podrá ser usada por actores interesados en el conocimiento y manejo de procesos de especies claves y su relación con los ecosistemas que los rodea; como es el caso del manejo del oso de anteojos en los ecosistemas de páramo del Parque Nacional Podocarpus.
5
incipientes en el Ecuador y sobre todo en la región sur del Ecuador; por ello, el uso de técnicas anatómicas que apoyen a la identificación de especies mediante características anatómicas fue uno de los aspectos que direccionó ésta investigación; pues la funciones están relacionadas con las estructuras celulares y su conocimiento ayuda a mejorar la capacidad entendimiento de los usos actuales.
Esta investigación tuvo como finalidad el estudio de las estructuras anatómicas de tres especies de Bromeliaceae del género Puya que constituyen parte de la dieta principal del oso
de anteojos en el Parque Nacional Podocarpus. Para lo cual se realizó la caracterización anatómica de las tres especies de Bromeliaceae del género Puya; y se complementó con el diseño de una herramienta (clave anatómica) cuya utilidad será la identificación de las tres bromelias del género Puya en estudios de dieta de herbívoros.
La metodología que fue utilizada en esta investigación inició con una revisión analítica de trabajos y publicaciones científicos relacionados con estudios de anatomía foliar de varios grupos de plantas y autores, de ellos se citan: Tomlinson (1969), Bezing (2000), Aoyama y Sajo (2003), Proença y Sajo (2007), Horres et al. 2007, Derwidueé y González (2010); y a
6
7
1.1 La familia Bromeliaceae
La familia Bromeliaceae constituye una familia muy diversa (Aguilera et al., 2011) y la más
grande de especies endémicas en el Neotrópico. Aproximadamente existen 3.140 especies distribuidas en 56 géneros (Luther, 2006; 2008). Se subdividen en tres subfamilias: Bromelioideae, Pitcairnioideae y Tillandsioideae (Tomlinson, 1969; Stevens, 2001). Sin embargo en los últimos años esta clasificación ha tenido algunos cambios.
Para la subfamilia Pitcairnioideae Smith (1974) en su monografía “Flora Neotrópica” agrupa a
13 géneros: Puya (168 especies); Encholirium (14 especies); Fosterella (13 especies);
Connellia (4 especies); Cottendorfia (24 especies); Deuterocohnia (7 especies); Abromeitiella
(2 especies); Pitcairnia (262 especies); Brocchinia (18 especies); Ayensua (1 especie); Navia
(74 especies); Dyckia (106 especies); y Hechtia (45 especies). En total se registras 738
especies. La subfamilia Pitcairnioideae reúne a todos los géneros con especies usualmente provistas de hojas con espinas en sus márgenes sin embargo se presentan excepciones en algunos especies (Manzanares, 2005).
Las bromelias son plantas usualmente de hojas arrosetadas, con hábitos, terrestres, saxícolas o epífitas, éstas últimas bien diversas y abundantes en selvas húmedas, es decir comprenden diversos tipos de vegetación y rangos de altitud, desde el nivel del mar hasta los 4000 msnm (Smith y Downs, 1974; 1979; Benzing, 2000). Por esta razón son importantes desde el punto de vista ecológico debido al hábito epífito de muchas de sus especies, así como a las diversas adaptaciones que presentan para vivir en hábitats xéricos (Benzing 2000); como también son de primordial importancia para el funcionamiento de ciertos ecosistemas, ya que contribuyen notablemente a la riqueza de especies, ocupan un lugar importante dentro del ciclo de nutrientes y establecen un gran número de interacciones con otros organismos (Strong, 1977; Castellaro et al. 2007 ).
También incluye especies consideradas como plantas suculentas ya que sus hojas presentan estructuras carnosas con caracteres xerofíticos que son capaces de retener agua en sus tejidos (Cruz et al., 2010) poseen parénquima almacenador de agua, tejidos de sostén y
8
1.2 Genero Puya
El género Puya fue establecido por Giovanni Ignazio Molina (1737-1829), originalmente
identificado como Juan Ignacio Molina, sacerdote jesuita, chileno, misionero, botánico y colector de plantas. Es el segundo género más numeroso de la subfamilia Pitcairnioideae, con 199 especies y 11 variedades (Luther, 2002); y está distribuido a lo largo de la cordillera de los Andes, desde el sur Venezolano hasta el norte de Chile y Argentina. En Ecuador se han registrado un total de 34 especies, una variedad y 2 híbridos naturales entre las especies registradas se consideran que 27 son endémicas (Manzanares, 2005).
Son plantas terrestres o saxícolas que forman una densa roseta donde no se almacena el agua de lluvia, es decir sin fitotelma; los asientos de las hojas suelen ser carnosos y constituyen parte del tallo, son gruesos y almacenan los hidratos de carbono necesarios para el momento de floración y fructificación; las láminas de las hojas son triangulares, coriáceas y están provistas de fuertes espinas en sus márgenes que les sirven para protegerse de los animales herbívoros, y no presentan angostamiento en su base. Las flores son grandes, conspicuas y actinomorfas; los pétalos se enroscan después de la antesis; a excepción de la
Puya ferruginea que puede tener dos callos verticales cerca de la base, ningún pétalo presenta
apéndices; presentan un ovario siempre súpero, y las semillas están provistas de un conspicuo alerón a su alrededor (Manzanares, 2005).
Existen dos tipos de Puya, las especies que mueren después de la floración (llamadas
monocárpicas) y las que mantienen un continuo desarrollo después de la floración, llamadas policárpicas); en las ultimas se observa un tallo ramificado mientras que en las monocárpicas el tallo es simple (Manzanares, 2005).
Según Manzanares (2005), se pueden encontrar dos subgéneros de Puya el Puyopsis y el
Puya; el primero, abarca todas las especies de Puya con las ramas completamente fértiles
9
1.3 Algunos estudios anatómicos en Bromeliaceae
La familia Bromeliaceae ha sido objeto de investigaciones a nivel anatómico por diversos autores, a continuación se presentan algunas de los más importantes estudios:
Tomlinson (1969), caracterizó cada una de las tres subfamilias de Bromeliaceae, basándose en los tricomas, estomas y mesófilos.
Benzing (1980; 1990) y Robinson (1969), publicaron los trabajos más comprensivos de anatomía de tallo y hojas de algunas bromelias.
Pita y Menezes (2002), reportaron por primera vez, velamen en raíces de especies terrestres de algunos géneros de Pitcairnioideae.
Brighigna et al. (1984), reportaron la anatomía foliar tipo Kranz asociada a un
metabolismo C4 en especies de Tillandsia.
Tomlinson (1969) y Aoyama y Sajo (2003), manifiestan que las hojas presentan caracteres xerofíticos, frecuentemente son suculentas, poseen parénquima almacenador de agua, tejidos de sostén y tricomas capaces de absorber agua y nutrientes.
Horres et al. (2007) realizaron el primer análisis filogenético considerando caracteres
anatómicos en Bromeloideae, indican que la anatomía puede dar información para discernir grupos a nivel de subgéneros como en Aechmea subgénero Lamprococcus
(Beer) Benth y en Aechmea subgénero Pothuava Baker, aunque hay caracteres que
han evolucionado independientemente, pero agregan que la anatomía es una fuente de información inexplorada y subutilizada.
10
1.4 Utilidad de las técnicas histológicas o microhistológicas
La técnica histológica se refiere a una serie de pasos ordenados para la obtención de muestras de tejidos que conservan todos sus detalles estructurales para poder estudiar las características morfológicas con la ayuda de un microscopio (Sandoval, 2005). Existen varias técnicas que se utiliza para el procesamiento de los tejidos, éstas dependen de las características que se desea observar.
Las técnicas histológicas se las ha aplicado en múltiples disciplinas como en anatomía, fisiología, histoquímica, etnobotánica, ecofisiología, etc. En cada una de ellas, han sido usados para: (i) estudiar las características morfológicas, moleculares, el funcionamiento de órganos, tejidos y una posible relación de sus características morfológicas y su funcionamiento; (ii) también para estudiar los efectos que causa la salinidad y el estrés hídrico en los cambios anatómicos y morfológicos; (iii) conocer aspectos básicos de la biología reproductiva y obtener información más precisa sobre la reproducción de las especies mediante estas técnicas; (iv) así como también para estudiar la composición botánica de la dieta de herbívoros (Sandoval 2005 ; D’ Ambrogio 1970).
Además el conocimiento de la histología es esencial para poder comprender los procesos bioquímicos y fisiológicos y para formarse una idea sobres las alteraciones estructurales que dan lugar a cambios funcionales y bioquímicos.
Las técnicas microhistológicas han sido ampliamente utilizadas para estudiar la dieta de herbívoros silvestres y domésticos (Sparks y Malechek, 1968; Klee et al., 2000; Holechek et al., 2001; Castellaro et al., 2007). Es por ello, que estas técnicas pueden establecer patrones
para identificar, determinar, comparar y cuantificar estructuras particulares de cada una de las especies vegetales (Ortega et al., 1993; Castellaro et al., 2007). Esta técnica consiste en la
obtención de tejidos epidérmicos vegetales desde una o más estructuras morfológicas de las principales especies que participan en el medio en que los animales se desarrollan (Borgnia, 2009); además proporcionan datos relevantes para el manejo de los sistemas ecológicos y de las relaciones entre sus componentes (Catán et al., 2003).
11
establecen que es una de las especies más usadas en la alimentación del oso de anteojos y cabe mencionar que no se cuenta con una clave de identificación anatómica de las mismas en el Parque Nacional Podocarpus.
1.5 El Parque Nacional Podocarpus (PNP) y su especie bandera: el oso de anteojos
El Parque Nacional Podocarpus (PNP) es conocido por su gran biodiversidad y alto grado de endemismo, debido principalmente a su ubicación en la depresión de Huancabamba y al amplio rango altitudinal que va desde los 800 hasta los 3800 msnm (Lozano et al., 2005). En
el PNP existen diferentes zonas de vida las cuales son caracterizadas por sus plantas y animales. En sus páramos una de las familias presentes en gran medida son las Bromeliaceae, las cuales constituyen lo más importante de la dieta del oso de andino
(Tremarctos ornatus).
La alimentación del oso de anteojos se basa principalmente en una dieta de fibra que la obtiene de algunas plantas como las bromelias terrestres y epífitas entre otras (Suárez, 1988; Cuesta et al., 2001; Castellanos et al., 2005; Achig, 2009). A pesar de las espinas en las
puyas, las plantas jóvenes son un bocado exquisito para los osos de anteojos (Manzanares, 2005). Estudios recientes de las especies de la familia de Bromeliaceae establecen que es una de las especies más usadas por el oso de anteojos recalcando en abundancia la Puya
maculata y la Puya eryngiodes como también se ha reportado el consumo de Puya nítida.
Siendo las Bromelias uno de los principales alimentos del oso andino, su estudio y conocimiento de aspectos claves puede resultar clave para el entendimiento de los hábitos alimenticios del oso andino
Los hábitos alimentarios proporcionan información crucial de la historia de vida y patrones conductuales de especies de fauna silvestre (Rodríguez et al., 2003). Por ejemplo se ha
encontrado que la dieta del oso andino, es básicamente omnívora, y que contiene en gran proporción lípidos y grasas que obtiene de bromelias terrestres y poaceas (Goldstein, 2008), mientras que el azúcar la obtiene de frutos ricos en azúcares (Peyton, 1999).
12
osos viven mayormente en el bosque nublado donde obtienen alimentación y protección; sin embargo cuando el alimento es escaso, este se desplaza al páramo en su búsqueda (Cuesta
et al., 2001).
1.6. Objetivos
El estudio de la anatomía foliar de tres especies de Bromeliaceae como componentes de la dieta del oso andino en el Parque Nacional Podocarpus se realizó a través del siguiente objetivo general:
- Describir las estructuras anatómicas de tres especies de Bromeliaceae del género
Puya que constituyen parte de la dieta principal del oso de anteojos en el Parque
Nacional Podocarpus.
Los objetivos específicos que se plantearon para la presente investigación fueron:
1. Caracterizar anatómicamente las tres especies de Bromeliaceae del género Puya
2. Diseñar una herramienta que pueda ser usada para la identificación de las tres bromelias del género Puya en estudios de dieta de herbívoros.
Las hipótesis planteadas y respondidas con este estudio fueron:
1. Cada una de las especies del género Puya estudiadas se identifican en base de sus estructuras anatómicas
2. Las tres especies de puya pueden ser identificadas siguiendo claves anatómicas
13
14
El estudio de la anatomía foliar de tres especies de Bromeliaceae como componentes de la dieta del oso andino, se realizó en dos fases de trabajo. La fase de campo que consistió en la selección de sitios con presencia de las especies seleccionadas y visita a estos ecosistemas para la colección de materia vegetal; y la fase de laboratorio, donde se realizó la caracterización anatómica de cada especie de Puya. El procedimiento y los materiales
utilizados en estas dos fases se detallan a continuación:
2.1 Selección de especies
En esta investigación se utilizó tres especies del genero Puya (Bromeliaceae), mismas que
fueron seleccionadas sobre la base que constituyen una de los principales alimentos del oso andino (Castellanos 2003, Ontaneda y Armijos, 2012). Las especies seleccionadas fueron:
Puya eryngioides, Puya maculata y Puya nítida; a continuación se presentan algunas
características ecológicas de cada una de las tres especies de Puya.
Puya eryngioides.- es una planta de 30-20 cm de largo con la inflorescencia policárpica, tallo
ramificado, presenta un follaje denso, subcoriáceo, rosulado y numeroso. La lámina de las hojas tienen forma triangular 15-20 cm de largo, 1-1,5cm de ancho márgenes serrados con espinas 2-3mm de largo, ápice largo acuminado y punzante. La base de la hoja es orbicular 2cm de largo, 3cm de ancho. Esta es una especie endémica del Ecuador, y se distribuye en páramos húmedos de 2500 - 3500 msnm desde las provincias de Loja, Azuay, Zamora Chinchipe. Las principales amenazas son incendios, fragmentación de los bosques y actividades antropogénicas. En 1997 fue clasificada como especie vulnerable y se encuentra en la categoría de preocupación menor (Manzanares y Pitman, 2003).
Puya maculata.- es una planta de 100 cm de largo con la inflorescencia monocárpica tallo no
ramificado, el follaje es denso, numeroso, punzante. La lamina de la hoja es triangular de 25-40 cm de largo, márgenes fuertemente serrados, espinas 10 mm de largo, negras y la base orbicular 4.5cm de largo, 4cm de ancho, y maculada marrón castaño, ápice largo atenuado y punzante. Es endémica de Ecuador, y se encuentra ampliamente distribuida en Azuay, Loja, Morona-Santiago y Sucumbíos en altitudes entre los 3000 y 3500 msnm. Las principales amenaza son incendios forestales y destrucción del hábitat. En 1997 la especie fue clasificada en peligro de extinción, debido a amplios estudios de distribución y en el 2005 fue catalogado en peligro crítico (Manzanares, 2005).
Puya nítida.-es una planta de 150-200 cm de largo con la inflorescencia policárpica, rosulada,
15
numeroso, punzante. La lámina de la hoja es triangular 40-60 cm de largo, 3.5-5cm de ancho, arqueada, márgenes serrados, espinas 6mm de largo, marrón castaño, ápice atenuado y punzante La base es suborbicular aproximadamente 9 cm de largo, 7 cm de ancho. Prefiere los ambientes nublados y húmedos, o zonas donde se aglomeran las nubes bajas (Manzanares, 2005). Habita en los páramos aproximadamente a una altitud de 2850 - 3270 msnm
2.2 Sitios de colección de especímenes
Sobre la base de información de los estudios realizados por Ontaneda y Armijos (2012), y el proyecto de investigación de la dieta del oso liderado por Cisneros 2014, y del Herbario Reinaldo Espinosa de la UNL, se seleccionaron sitios con presencia de las tres especies. Estos sitios corresponden al sector el Tiro y Cajanuma, los dos dentro del complejo de ecosistemas de páramo del Parque Nacional Podocarpus1 (ver Figura 1). El rango altitudinal
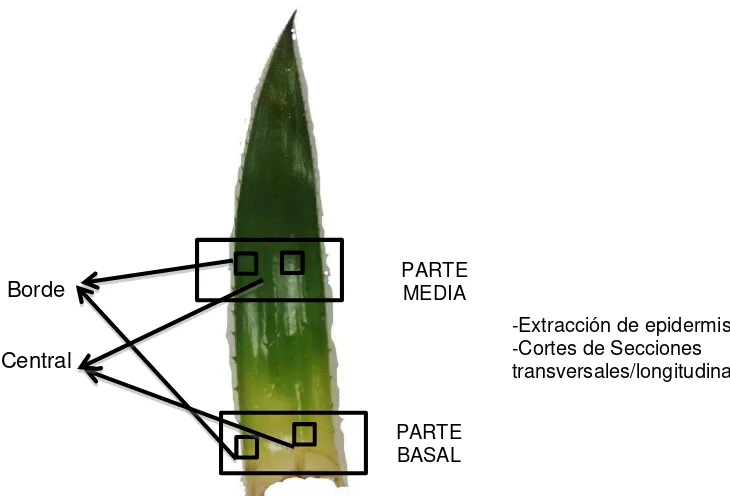
[image:20.595.90.396.382.643.2]de colección está entre los 2800- 3200 msnm
Figura 1. Sitios de colección de hojas de las tres especies de Puya, dentro del ecosistema de paramo del Parque Nacional Podocarpus.
2.3 Colección de muestras y especímenes (muestreo)
1Los páramos son ecosistema compuestos de vegetación herbácea y arbustiva y albergan un alto endemismo,
16
La colección de muestras (10 hojas de diferentes individuos de cada especie), se realizó en los dos sectores antes señalados y que se pueden ver en la figura 1. En el sector el Tiro se colectaron muestras de Puya eringioides y Puya nítida; mientras que en el sector de Cajanuma
se colectó Puya maculata.
Se colectaron 10 hojas de distintos individuos (una hoja por planta), por cada una de las tres especies, bajo el criterio que las plantas estén en excelentes condiciones fitosanitarias, y sin presencia de indicios de plagas y enfermedades; las cuales fueron transportadas al laboratorio de Fisiología Vegetal de la UTPL
Posteriormente, de cada hoja se cortó la parte media y basal y fueron fijadas por 7 días en FAA (Formaldehido 5ml, ácido acético 5ml y alcohol etílico 70% 90ml) (Figura 2), para conservar la estructura morfológica. Tanto de la parte media y basal, se prepararon muestras de las partes adaxial y abaxial de la hoja; y además se colectaron muestras de la parte media y el extremo (borde) de las secciones antes indicadas (ver Figura 2)
[image:21.595.157.522.404.652.2]
Figura 2. Secciones muestreadas de la hoja de Puya nítida. Se tomaron muestras de la parte media y basal de cada hoja colectada, además se diferenció la parte central y el borde de la hoja. La figura muestra la parte adaxial, sin embargo considerar que se muestreó también la parte abaxial.
2.4. Evaluación de las estructuras morfológicas de las hojas de bromelias.
-Extracción de epidermis -Cortes de Secciones transversales/longitudinales
Borde
Central
PARTE MEDIA
17
En primer lugar se procedió a la obtención de láminas semipermanentes, a través de cortes paradérmicos transversales y longitudinales.
Para los cortes transversales se utilizó la técnica a mano alzada desarrollado por Johansen (1940), a continuación se presenta el resumen del procedimiento que se utilizó para obtener las láminas semipermanentes:
- Se seleccionó la región media (es decir la parte más ancha de la lámina) y la región basal de cada hoja.
Selección de pequeñas porciones de material vegetal, aproximadamente (1cm x 2cm) de la parte media y del borde de las regiones antes mencionadas.
Se prosiguió a cortar con la ayuda de una hojilla de afeitar.
Se colocaron cada lámina en hipoclorito de sodio al 5%, por unos tres minutos aproximadamente, hasta ver que el tejido cambia de color.
Posteriormente se procedió a lavar las muestras con agua destilada
A continuación se tiñeron con azul de toluidina (al 0,01%) por aproximadamente dos minutos, o hasta observar que se tiña la muestra..
Finalmente se montaron en porta objetos utilizando glicerina-agua (1:1) y las preparaciones se sellaron con esmalte transparente para uñas (Salas et al. 2001).
Para los cortes paradérmicos (obtención de la epidermis adaxial y abaxial), se realizó a través de macerado con hipoclorito de sodio al 5.5%, a través del siguiente procedimiento:
Se colocó las porciones de material vegetal aproximadamente (1cm x 2cm) de la parte media (adaxial y abaxial) en hipoclorito de sodio al 5.5% con agua (1:1) aproximadamente por 48 horas dependiendo de la especie.
Luego se lavaron las muestras con agua destilada.
Posteriormente, se procedió a separar la epidermis con la ayuda de una aguja histológica.
Se tiñeron con azul de toluidina 0.01% por aproximadamente 1 minuto hasta ver que el tejido se tiña.
Las muestras se montaron en porta objetos con glicerina-agua (1:1) y se sellaron con esmalte transparente para uñas, siguiendo las recomendaciones de Salas et al.
(2001).
18
Una vez obtenidas las láminas de cada sección (20 por especie); se procedió a observar la epidermis, cortes transversales y longitudinales tanto adaxiales y abaxiales. Para ello se utilizó un microscopio óptico Labomed Lx 400 con aumentos de 10x, 20x y 40x.
Finalmente, se procedió a realizar fotografía de cada estructura observada, en el microscopio axiostar plus con aumentos de 10x, 40x y 100x con cámara digital adaptada al microscopio marca Canon EOS Rebelt t2i con un lente macro Canon 60mm.
2.5 Metodología para diseñar una herramienta para la identificación de bromelias en estudios de dieta de herbívoros.
Con la información generada en el primer objetivo, se construyó una clave de identificación para las tres especies de Puya estudiadas. Para ello, se sistematizaron los resultados
obtenidos en los diferentes análisis anatómicos foliares, los cuales fueron agrupados siguiendo patrones estructurales y procedimientos estandarizados para la construcción de claves dicotómicas que permitan diferenciar cada especie. Los caracteres de diagnóstico se detallan en las tablas 1, 2 y 3. Las cuales se las puede observar en la sección de resultados.
Para el establecimiento de la clave dicotómica se consideraron las siguientes variables: - En la epidermis: tipo de tricoma, ubicación de los estomas (adaxial/abaxial), nivel de
estomas en la epidermis, y cámara subestomática
- En el sistema fundamental: tipo de parénquima, y grado de lignificación de las paredes. - En el sistema vascular: prolongación de la vaina, y presencia de vaina
19
20
3.1. Caracterización anatómica de Puya eryngioides
Tejido epidérmico
La parte basal de las hojas de P. eryngioides presentan células de epidermis ovaladas casi
planas con lumen reducido, aparentemente incluye cuerpos de sílice casi esféricos (Figura 3) la hipodermis no se distingue claramente de las células del parénquima acuífero en la cara adaxial (Figura 3A), mientras que en la superficie abaxial la hipodermis está formada por una capa de células de forma redondeada con paredes ligeramente engrosadas anticlinalmente y también ligeramente esclerificadas (Figura 3B); y el contorno de la epidermis superior o adaxial es ligeramente ondulado, mientras que el contorno de la superficie inferior o abaxial es lisa.
Figura 3. Sección transversal de la epidermis de P. eryngiodes. (A) epidermis adaxial con células ovaladas con lumen reducido e hipodermis no diferenciada con el parénquima acuífero; (B) epidermis abaxial con células ovaladas e hipodermis formada por una capa de células ligeramente esclerificadas. Barra = 100um
La vista paradérmica de la parte media de la lámina foliar revela que las epidermis adaxial y abaxial, están formadas por células rectangulares de paredes engrosadas, sinuosas en mayor o menor grado, dependiendo de su posición sobre las nervaduras o zonas intercostales (Figura 4A-D).
[image:25.595.90.561.320.501.2]21 Figura 4. Vista paradérmica de la lámina foliar de la parte media de la epidermis adaxial y abaxial de P. eryngiodes. (A) células de la epidermis adaxial constituidas por células rectangulares con paredes engrosadas; (B) células de la epidermis abaxial formada por células rectangulares con paredes engrosadas, sinuosas y mostrando el patrón de distribución de estomas; (C) detalle de la superficie adaxial; (D) detalle de superficie abaxial. Barra 100um
Además, los cortes transversales revelan que ambas epidermis de la lámina foliar son uniestratificadas con células ovalada a poligonales, con pared tangencial externa y pared radial relativamente delgadas, sin embargo la pared tangencial interna es muy engrosada, formando un lumen muy reducido (Figura 5A y 5B). La hipodermis presenta un número variable de capas que van de dos a tres capas de células, con engrosamiento desuniforme, tienen una pared externa más delgada en comparación con la pared interna y presentan células esclerificadas en la cara abaxial (Figura 5B), mientras que en la cara adaxial la hipodermis presenta una sola capa con células ligeramente esclerificadas, aparentemente las células son más engrosadas en la cara abaxial (Figura 5A). Tanto la epidermis adaxial como la abaxial de la parte media son lisas y no presentan ondulaciones. Y se encuentra revestida por una cutícula engrosada.
B A
22 Figura 5. Vista frontal de la lámina foliar de la parte media de la epidermis e hipodermis adaxial y abaxial de P. eryngioides. (A) epidermis de la superficie adaxial con pared tangencial interna muy engrosada, seguida por una capa de hipodermis ligeramente esclerificadas; (B) epidermis e hipodermis abaxial mostrando engrosamiento desuniforme que delimitan un lumen muy reducido con células esclerificadas. Barra = 100um.
Las hojas de P. eryngiodes son hipoestomáticas, es decir sus estomas se encuentran
únicamente en la cara abaxial, ubicándose en las zonas costales e intercostales de la lámina (Figura 4B). Los estomas están constituidos por cuatro células subsidiarias, dos laterales y dos polares, por lo que se clasifican como tetracíticos (Figura 6A).
Con los cortes transversales en la cara abaxial se observó la presencia de cámara subestomática, y además se observó que las células oclusivas presentan engrosamiento de pared y están ubicadas al mismo nivel que las células epidérmicas.
Figura 6. Vista paradérmica de la parte media de la lámina foliar de P. eryngiodes. (A) detalle del estoma tetracítico formado por cuatro células subsidiarias; csl= célula subsidiaria lateral, cst = célula subsidiaria terminal; (B) estoma al mismo nivel de las células epidérmicas, detallando la cámara subestomática; cámara subestomática=cs. Barra= 50um (Figura A); 100um (Figura B).
B A
csl
cst
A B
[image:27.595.94.537.517.688.2]23
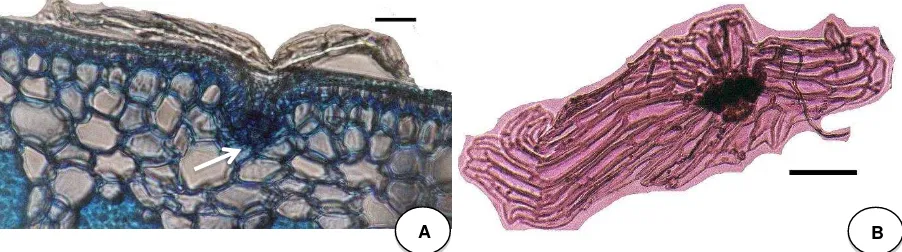
[image:28.595.94.545.242.368.2]En la superficie superior de la lámina foliar se presentaron tricomas tipo escamas peltadas, distribuidas en surcos longitudinales y constituidos por un número muy variable de células (Figura 6B). Están compuestos por tres partes: un disco central, un pie o base y un ala, Derwidueé y González (2010). Las células periféricas son alargadas. La base del tricoma está formado por paredes engrosadas, es hundida en depresiones epidérmicas y multicelular compuesta por células esclerenquimáticas tipo esclereidas (Figura 7A-B).
Figura 7. Vista frontal de la lámina foliar de hoja de P. eryngioides, (A) detalle de la base de la escama peltada compuesto por más de dos células; (B) detalle de la escama pelta (tricoma) con sus alas alargadas. Barra = 100um.
Sistema fundamental
El mesófilo es el conjunto de tejidos ubicados entre las dos epidermis. Desde la parte adaxial hacia adentro, se encuentra un parénquima acuífero de tamaño variable, seguido de parénquima en empalizada y clorénquima.
En la parte media de la lámina, el parénquima acuífero está compuesto por células redondeadas con paredes celulares muy tenues, son translucidas y de diversos tamaños (Figura 8C). A continuación se encuentra el parénquima en empalizada, el cual está formado por células alargadas con un número variado de capas que pueden ser de tres a cuatro células, y finalmente se encuentra el parénquima esponjoso con pequeños espacios intracelulares, el cual está formado por cinco a ocho capas de células y que presenta mayor cantidad de cloroplastos (Figura 8A).
En la parte basal de la lámina, se encuentran los haces vasculares en su zona media, y alrededor de estos se encuentra el parénquima acuífero que presenta diferencias de tamaño ya que el de la superficie adaxial las células son grandes y ovaladas con las paredes anticlinales y periclinales ligeramente engrosadas, mientras que en la superficie abaxial las células son pequeñas y redondeadas, ligeramente esclereficadas (Figura 8D). El aerénquima
24
[image:29.595.90.548.181.630.2]presenta canales longitudinales de aireación situados entre los haces vasculares, estos canales están atravesados por células braciformes irregulares los cuales presentan una ligera prolongación hacia la superficie abaxial (Figura 8B y 8D).
Figura 8. Vista frontal de la lámina foliar de la parte media y basal de P. eryngiodes. (A) parte media, parénquima en empalizada formado por células alargadas y parénquima esponjoso con pequeños espacios intracelulares; (B) parte basal, aerénquima con canales longitudinales situados entre los haces vasculares; (C) parte media de la lámina foliar mostrando su parénquima acuífero redondeadas con paredes celulares muy tenues; (D) detalle del aerénquima de la parte basal; (E) parte basal de la lámina foliar mostrando el parénquima acuífero formado por células de diferentes tamaños. Barra 100um
pa
es
A
B
C
D
25
Sistema vascular
En la parte media de la lámina, los haces vasculares son colaterales cerrados de diferentes tamaños pudiendo encontrarse grandes y alargados, pequeños y alargados; y pequeños y cortos, los cuales se encuentran alternados (Figura 9A y Figura 9B). Se encuentran rodeados por fibras en forma de vaina o casquetes fibrosos. La vaina parenquimática está compuesta por células redondeadas de diversos tamaños, a veces es interrumpida por el esclerénquima del haz y se observa que están asociadas a los haces vasculares. Tanto los haces grandes como los pequeños tienen conexión con la epidermis en la cara abaxial, por ello se denominan transcurrentes. Los haces de mayor tamaño se prolongan hacia ambas caras de la epidermis, hacia la cara adaxial colindando con el parénquima acuífero y hacia la cara abaxial contiguo con la hipodermis, mostrando claramente extensiones de las vainas de los haces. En los bordes de la hoja se observó que en ocasiones los haces se conectan entre sí, es decir que existen haces con anastomosis (Figura 9C). Además se observó conexiones entre los haces de mayor a menor longitud a través de las comisuras (Figura 9B).
En la parte basal, la haces vasculares están rodeados por fibras, presenta diferentes tamaños, alternándose los más grandes con los más pequeños, los de mayor tamaño están rodeados por células esclerenquimáticas mientras que en los de menor tamaño se extienden hacia la cara abaxial (Figura 8B,D).
A B
26 Figura 9. Vista frontal de la lámina foliar de P. eryngioides, (A) detalle de los haces vasculares rodeados por fibras en forma de vaina o casquetes fibrosos; (B) detalle de la comisura del haz (CH); (C) detalle de los haces vasculares con anastomosis (AN). Barra = 100um
3.2 Caracterización anatómica de Puya maculata
Tejido epidérmico
Las epidermis adaxial y abaxial de la parte media están formadas por células rectangulares de paredes ligeramente engrosadas (Figura 9A, B), siendo las células epidérmicas más sinuosas en la cara abaxial (Figura 9B, D). La cutícula es delgada, sin embargo en la superficie abaxial y sobre la cutícula se observa una capa espesa, que podría ser cera (Figura 9B, E).
A B
C
27 Figura 9. Vista paradérmica de la parte media de la epidermis adaxial y abaxial P. maculata (A) superficie adaxial con paredes ligeramente engrosadas; (B) superficie abaxial, con escamas que se distribuyen en hileras longitudinales; (C) detalle de superficie adaxial mostrando las células rectangulares; (D) detalle de la superficie abaxial mostrando las células epidérmicas sinuosas; (E) detalle de la superficie abaxial mostrando una capa espesa sobre la cutícula. Barra = 1ooum
En los cortes transversales de la zona media se observó que ambas epidermis de la lámina foliar son uniestratificadas y ligeramente engrosadas. En la cara adaxial las células de la epidermis presentan lumen reducido, mientras que en la cara abaxial la forma de las células son poligonales y aparentemente con presencia de pequeños cuerpos de sílice. Se observa una hipodermis de tipo mecánico, que cumple su función estructural, presentando células rectangulares o poligonales, cuyas paredes se presentan uniformemente engrosadas y ligeramente lignificadas. En la superficie adaxial la hipodermis presenta de una a dos capas de células ligeramente esclerificadas (Figura 10A), mientras que en la superficie abaxial las células de la hipodermis presentan células esclerificadas, rectangulares a poligonales, paredes anticlinales, con un engrosamiento desuniforme, y presenta cuatro capas de células estando más desarrollada en comparación con la cara adaxial (Figura 10B). El contorno de la
E
[image:32.595.92.549.83.455.2]28
epidermis adaxial es lisa a ondulada y la epidermis abaxial es fuertemente ondulada (Figura 10). En ambas capas presenta cutícula delgada, sin embargo en la superficie abaxial se observan pequeñas proyecciones.
Figura 10. Vista frontal de la epidermis de la parte media de la lámina foliar de Puya maculata. (A) superficie adaxial con células poligonales y mostrando la hipodermis con paredes ligeramente esclereficadas; (B) epidermis de superficie abaxial mostrando cuerpos de sílice (s), e hipodermis desarrollada con células rectangulares a poligonales, además se observa la cámara subestomática; cámara subestomática =cs. Barra = 100um
En cortes transversales de la región basal de la hoja de P. maculata se observó que la
epidermis es uniestratificada y que las células epidérmicas son casi planas con lumen reducido (Figura 11A-B). La hipodermis adaxial y abaxial presenta una capa de células ovaladas a rectangulares con paredes tangenciales ligeramente engrosadas. La cutícula es gruesa presentando en el contorno de la superficie abaxial una capa de espesor fuera de lo epidérmico, que puede ser cera
A B
[image:33.595.92.545.151.364.2]29 Figura 11. Vista frontal de la epidermis de la parte basal de P. maculata (A) superficie adaxial mostrando la epidermis uniestratificada con lumen reducido e hipodermis con células ovaladas; (B) epidermis abaxial mostrando cutícula espesa e hipodermis con una capa de células. Barra 100um
Las hojas de P. maculata son hipoestomáticas, tienen los estomas al mismo nivel de las
células epidérmicas. El estoma está constituido por cuatro células subsidiarias dos laterales y dos polares (Figura 12A), la clasificación de éstas estructuras se describe como tetracítico. Los estomas se distribuyen en las crestas a lo largo de las hojas (Figura 12B). La cámara subestomática varía en profundidad y se comunica con el clorénquima.
B
A
A B
csl
cst
t
30 Figura 12. Vista paradérmica de la lámina foliar de P. maculata. (A) detalle del estoma tetracítico formado por cuatro células subsidiarias, csl = célula subsidiaria lateral, cst = célula subsidiaria terminal; (B) superficie abaxial, patrón de distribución de estomas (es) y tricomas (t), células epidérmicas de paredes rectangulares y sinuosas. Barra = 50um=A; 100um=B
[image:35.595.92.542.323.663.2]En la superficie superior de la lámina foliar se presentaron tricomas tipo escamas peltadas, los cuales se distribuyen en hileras longitudinales (Figura 12B), las escamas se insertan en los surcos y su porción peltada se extiende sobre las crestas, cubriendo completamente a los estomas. Las escamas están constituidos por un número muy variable de células (Figura 13A, B, C). El escudo que a menudo es asimétrico y aparentemente formado por dos células centrales, de tres a cuatro anillos de células con paredes de forma ovalada, ligeramente interconectadas y dispuestos concéntricamente con las células centrales. Estas escamas se insertan en filas paralelas al eje longitudinal de la hoja (Figura 12B).
Figura 13. Detalle de los tricomas de P. maculata. (A-B) vista paradérmica escama peltada, constituido por un número variable de células; (C) detalle de la base del tricoma en vista frontal. (D) detalle de las alas periféricas del tricoma en vista frontal. Barra = 100um
Sistema fundamental
A B
31
[image:36.595.87.259.153.263.2]En la parte media el parénquima acuífero está formado por células aparentemente isodiamétricas, que presentan paredes delgadas con rafidios (estructuras alargadas parecidos a un alfiler de oxalato de calcio) y sin espacios intracelulares (Figura 14).
Figura 14. Detalle del parénquima acuífero formado por células alargadas con presencia de rafidios (r)
El clorénquima está formado por células en empalizada que a su vez están formadas por tres a cinco capas de células (Figura 15B), y por células emparejadas que delimitan pequeños espacios de aire con forma isomorfica y con presencia de cloroplastos. Este está constituido por tres a ocho capas de células también llamadas canales de aire o aerénquima (Figura 15A). Estos canales de aire usualmente están continuos a la hipodermis en la superficie abaxial, con células isodiamétricas de proyecciones cortas. Se disponen intercaladamente con los haces vasculares y se extienden de vez en cuando con la hipodermis de la superficie abaxial. Las células del clorénquima se disponen de forma concéntrica alrededor de los haces vasculares. Se observó presencia de canales secretores (cs) dispersos (Figura 15A).
Figura 15. Vista frontal de la parte media de la lámina de Puya maculata; (A) detalle del parénquima aerífero con células isodiamétricas, presencia de canales secretores (cs), ae = aerénquima; (B) detalles del parénquima en empalizada formada por células alargadas que se dispone en forma concéntrica alrededor de los haces vasculares, pe= parénquima en empalizada. Barra = 100um.
A
pe ae
B
[image:36.595.92.546.457.692.2]32
[image:37.595.91.543.370.548.2]En la región basal el parénquima acuífero está formado por células redondeadas y ubicado en las superficies adaxial y abaxial, presentando rafidios (Figura 16A). El aerénquima está formado por células de forma isodiamétrica de brazos largos con cloroplastos, se ubican en cámaras casi redondeadas a los costados de los haces vasculares (Figura 16B).
Figura 16. Vista frontal de la parte basal de la lámina foliar de P. maculata. (A) detalle del parénquima acuífero formado por células redondeadas con presencia de rafidios (r). (B) detalle de los canales de aire con cámaras casi redondeadas (cac). Barra = 100um
Sistema vascular
En la región media los haces vasculares son colaterales cerrados, están conectados con la hipodermis abaxial por células alargadas, dispuestos en una única serie, estos difieren en tamaño y en general los más grandes se alternan con las más pequeños (Figura 15B). Los haces vasculares de mayor tamaño están rodeados parcialmente o completamente por una vaina. Los haces de menor tamaño tienen casquetes que son más alargados que anchos. Se observó haces vasculares transversales o comisuras, que cruzan entre los canales aéreos y
A B
33
parénquima en empalizada, conectando con los principales haces longitudinales (Figura 17 A).
[image:38.595.90.542.321.566.2]En la parte basal los haces vasculares están ubicados en el centro de la lámina, son de tipo colateral varían su tamaño, alternándose los mayores con los menores están rodeados por fibras en forma de una vaina, conectados entre sí (Figura 17B) Los más grandes están rodeados por fibras esclerenquimáticas y externamente por una vaina. Los más pequeños presentan prolongaciones hacia la superficie abaxial.
Figura 17. Vista frontal de los haces vasculares de la parte media y basal de la lámina foliar de P. maculata. (A) parte media, detalle de las comisuras del haz, que se conectan con los principales haces longitudinales; (B) parte basal, detalle de los haces vasculares formados por una vaina (v), conectados entre sí. Barra = 50um (B)
3.3 Caracterización anatómica de Puya nítida
Tejido epidérmico
La cara adaxial de la epidermis de la parte media presenta células casi isodiamétricas, polimórficas con paredes desigualmente engrosadas (Figura 18A, C), mientras que su cara
A
B
c
34
abaxial está formada por células rectangulares de paredes engrosadas, sinuosas(Figura 18B, D).
Figura 18. Vista paradérmica de la epidermis de la parte media de la lámina foliar de P. nítida. (A) superficie adaxial con células polimórficas con paredes desigualmente engrosadas; (B) superficie abaxial con células rectangulares con paredes engrosadas sinuosas; (C) detalle la epidermis adaxial; (D) detalle de la superficie abaxial. Barra = 100um
En los cortes transversales de la región media se observó que ambas epidermis (adaxial y abaxial) de la lámina foliar son uniestratificadas. Las células de la epidermis son redondeadas y más pequeñas en relación con las células de la hipodermis. Las células epidérmicas tienen un lumen reducido debido al engrosamiento de las paredes periclinales internas y anticlinales (Figura 19A). La hipodermis es de tipo mecánica y varia de uno a cuatro capas de células esclereficadas, de forma rectangular con paredes ligeramente engrosadas. En la superficie
A B
[image:39.595.91.546.184.587.2]35
[image:40.595.99.545.260.440.2]adaxial la capa de la hipodermis comprende una a tres células, las cuales tienen lumen reducido, menor al observado en las células epidérmicas (Figura 19A). La hipodermis de la superficie abaxial es más desarrollada y está formada por dos a cuatro capas de células (Figura 19B). En los bordes de la lámina se observó que la hipodermis está compuesta por ocho capas de células esclereficadas ligeramente engrosadas y de forma poligonal. Aunque la cutícula es delgada, el contorno de la lámina foliar muestra que es lisa tanto para la superficie adaxial como la abaxial.
Figura 19. Vista frontal de la parte media de la lámina foliar de P. nítida. (A) superficie adaxial mostrando una hipodermis con células esclereficadas compuesta por dos a cuatro capas de células; (B) superficie abaxial mostrando la hipodermis más desarrollada. Barra = 100um
En cortes transversales de la región basal de la hoja de P. nítida se observó una cutícula
delgada y además que la epidermis está formada por una capa de células rectangulares con paredes desigualmente engrosadas en ambas superficies y con lumen reducido. Aparentemente se observan cuerpos de sílice de forma esférica (Figura 20B). En la superficie adaxial las paredes son delgadas presentando una hipodermis no muy desarrollada (Figura 20A) como la de la superficie abaxial, además la superficie adaxial está formada por una capa de células, mientras que la abaxial está formada por una a dos capas de células de paredes esclereficadas.
36 Figura 20. Vista frontal de la parte basal de epidermis e hipodermis de la hoja de P. nitida (A) superficie adaxial mostrando las paredes delgadas; (B) superficie abaxial mostrando paredes ligeramente engrosadas y esclereficadas. Barra
La hoja de P. nítida es hipostomática, encontrandose sus estomas al mismo nivel de las
células epidérmicas. Se encuentran ubicados en las zonas costales e intercostales de la lámina. Están constituidos por cuatro células subsidiarias, dos laterales y dos polares, por lo cual se clasifican como tetracítico (Figura 21A). Presenta estomas expuestos, es decir que las escamas de los tricomas no los cubren totalmente. La cámara subestomática (Figura 21B) puede estar parcialmente ocluida debido a la ampliación de las células subsidiarias debajo de las células de guarda.
37 Figura 21. Vista paradérmica de la lámina foliar de P. nítida; (A) detalle del estoma tetracítico constituido por cuatro células subsidiarias; csl = células subsidiarias laterales, cst = células subsidiaria terminal; (B) detalle de la cámara subestomática (cs). Barra = 50um (A); 100um (B)
Los tricomas, formados de tres partes fundamentales (ala, pedúnculo y disco), son peltados y se encuentran en ambas superficies de la lámina, pero en la superficie abaxial son más abundantes. En ambas partes se distribuyen en hileras longitudinales y se disponen en ranuras en las zonas intercostales, asociados con los estomas. En vista frontal se observa que están localizados en una concavidad de la epidermis (Figura 22A), y sus células del disco aparentemente están formadas por 8 células, rodeadas por 16 células periféricas y una gran cantidad de células que conforman el ala (Figura 22B).
A
B B
A csl
cst
38 Figura 22. Tricoma escamoso de la lámina de P. nítida. (A) vista frontal; (B-C) vista paradérmica que muestra algunas de las células que lo conforman. Barra = 200um (Figura A); 50um (Figura B-C).
Sistema fundamental
En la zona media el parénquima acuífero está formado por células alargadas, de paredes delgadas y sin espacios intracelulares. Ocupa más de la mitad de la lámina foliar, sus células poseen un citoplasma claro muy vacuolizado y presentan rafidios (Figura 23A). El paso entre el parénquima acuífero a clorénquima en empalizada es abrupto. El clorénquima está formado por células en empalizada y por células que delimitan espacios de aire. El
parénquima en empalizada está formado por células alargadas de tres a cuatro capas que se disponen de forma concéntrica alrededor de los haces vasculares (Figura 23B). En el
parénquima aerífero los canales de aire están situados entre los haces vasculares. Estos canales están contiguos a la cámara subestomática y están atravesados por diafragmas formados por células braciformes (Figura 23C). A menudo constituyen un largo sistema de aireación (Figura 23D) que se extiende continuamente en casi toda la lámina; éste está formado por células estrelladas con brazos cortos que delimitan grandes espacios de aire o con brazos cortos que forman pequeños espacios, ambos presentan abundantes cloroplastos (Figura 23C). Se distribuyen intercalando con los haces vasculares y se extienden escasamente con la superficie abaxial de la lámina folia
r.
39 Figura 23. Sección transversal y longitudinal de la parte media de la lámina foliar de P nítida. (A) detalle del parénquima acuífero con rafidios (r); (B) detalle del mesófilo destacando el clorénquima y el aerénquima y la distribución en serie de los haces vasculares; (C) detalle del parénquima aerífero por canales de aire ocupados por diafragmas de células braciformes; (D) detalle del parénquima aerífero, vista longitudinal; barra = 100um
En la región basal el parénquima acuífero está formado por células de paredes delgadas anticlinales, además algunas veces constituido por dos tipos de células redondeas y alargadas. El parénquima acuífero está ubicado en ambas superficies, presenta idioblastos con rafidios (Figura 24A). En la superficie adaxial las células son redondeadas y sus paredes anticlinales y periclinales son delgadas; mientras que en la superficie abaxial las células del
B
C A
D
40
parénquima acuífero son más alargadas y adyacentes a la hipodermis abaxial, y además se observan redondas hacia el interior del mesófilo (Figura 24A). El clorénquima está formado por células de forma estrellada que delimitan espacios de aire con brazos largos, frecuentemente con cloroplastos, se ubican en columnas alargadas a los costados de los haces vasculares (Figura 24B). En ocasiones estos canales de aire se prolongan hacia la cara abaxial.
Figura 24. Vista frontal de la parte basal de la lámina foliar de P. nítida. (A) parénquima acuífero constituido por dos tipos de células; (B) detalle de las columnas alargadas del aerénquima, L = lumen, EI = espacios intracelulares. Barras: A = 200um; B=50um
Sistema vascular
La región media presenta haces colaterales cerrados, distribuidos en una serie única y de tamaños variables, unos pequeños y otros más grandes que están de forma alternada (Figura 23B). Están conectados con la hipodermis de la cara abaxial por células redondeadas. Los haces más grandes están parcialmente rodeados por fibras (y en ocasiones completamente rodeados) en forma de una vaina con células casi isodiamétricas y ligeramente engrosadas que también se observan entre el xilema y el floema, además se encuentran cubiertos por una
B A
[image:45.595.92.533.237.558.2]41
capa de fibras, que pueden extenderse hacia ambas superficies, presentando punteaduras. Los más pequeños están formados por fibras rodeados parcialmente por una vaina con alargadas extensiones hacia ambas superficies (Figura 25).
Figura 25. Detalle del haz vascular grande rodeados por en forma de una vaina con células isodiamétricas y ligeramente engrosadas que también se observan entre el xilema y el floema
[image:46.595.86.185.152.297.2]En la parte basal los haces vasculares son de tipo colateral cerrado de tamaños variables alternándose los más grandes con los más pequeños. Los de mayor tamaño tienen gran cantidad de fibras envolviendo a los haces y externamente están rodeados por una vaina con células casi isodiamétricas. Los de menor tamaño están completamente rodeados por una vaina y presentan extensiones de la vaina con paredes engrosadas hacia ambas superficies (Figura 25).
3.4 Diseño de una clave para la identificación de especies del género Puya orientado a
42
Con la ayuda de la combinación de las características anatómicas fue posible realizar la clave de identificación para las tres especies estudiadas del género Puya. Se encontraron
[image:47.595.79.562.526.714.2]características particulares en la epidermis de cada una de las especies, las cuales podrían ser utilizadas para su identificación (Tabla 1). En esta vemos que los tricomas son distintos en cada especie.
Tabla 1. Características de los estomas y de los tricomas de las tres especies estudiadas. Simbología: ad = adaxial, ab = abaxial; + = presencia, - = ausencia; AL = alargado, SC = semicircular, OB = oblongo
Cara cte rís ti ca s Es pe cie s Estomas al nivel de la epidermis Presencia de cuernos cuticulares Pres enc ia d e cá m ara sub es to m átic a
Presencia de escamas peltadas en la superficie adaxial o
abaxial
Forma de los tricomas
Puya eryngioides + - + ad AL
Puya maculata + + + ab SC
Puya nítida + - + ad/ab OB
Fuente: Elaboración de la autora
También se pudo observar diferencias en las células que conforman la hipodermis tanto en su estructura como en su composición, al igual que la forma del contorno de la epidermis (Tabla 2).
Tabla 2. Características del contorno de la epidermis y de la estructura y composición de la hipodermis foliar de las tres especies estudiadas. Simbología: (LO = ligeramente ondula; FO = fuertemente ondulada; L/O = lisa a ondulada; L=lisa; EG = engrosada; LG = ligeramente engrosada; + = presencia; - = ausencia)
Cara cte rís ti ca s Es pe cie s
Contorno de la superficie
adaxial
Contorno de la superficie
abaxial
Hipodermis esclereficada
adaxial esclereficada abaxial Hipodermis
Media Basal Media Basal
Número de capas
de células Paredes
de la células
Número de
capas de células Paredes de las células
media basal media Basal
Puya
eryngioides L LO L L 1 - LG 2-3 1 EN
Puya maculata L/O LO FO L/O 1-2 1 EN 4 1 EN
Puya nítida + - + - 1-3 1 EN 2-4 1-2 EN
Fuente: Elaboración de la autora
43 Tabla 3. Características anatómicas observadas en el sistema fundamental y el sistema vascular de la lámina foliar de las tres especies estudiadas (Simbología: + = presencia)
Cara cte rís ti ca s Es pe cie s Parénquima en empalizada Parénquima
aerífero aeríferos Canales vasculares Haces
rodeados completamente
por una vaina
Haces vasculares
rodeados parcialmente por una vaina
Número de capas de
células
Forma de las células Can al es lo ng itu di na le s Cám ara s c as i re do nd as Col um na s al arg ad as
Media Basal Media Basal
Puya
eryngioides 3-4
Irregulares/
alargadas + - - + + - -
Puya
maculata 3-5 Isodiamétricas - + - + + + +
Puya nítida 3-4 Estrelladas - - + + + + +
Fuente: Elaboración de la autora
Del análisis y combinación de caracteres morfoanatómicos se determinó la siguiente clave para la identificación de las tres especies del género Puya.
1. Estomas al mismo nivel de las células epidérmicas
2. Parénquima acuífero con idioblastos portadores de rafidios 3. Parénquima en empalizada con 3-4 capas de células
………..Puya nítida
3. Parénquima en empalizada con 2-3 capas de células 4. Escamas solo en una superficie
………..Puya maculata
2. Parénquima acuífero sin idioblastos portadores de rafidios
5. Células del canal aerífero con proyecciones cortas isodiamétricas; canales longitudinales
44
45
A continuación se realiza un breve análisis de los aspectos más importantes de este trabajo investigativo. La discusión se centra por un lado en el proceso metodológico seguido en este estudio; y por otro lado en el análisis y evaluación de las implicaciones de los estudios morfológicos en estudios más aplicados.
4.1. Aspectos metodológicos para estudios morfoanatómicos
En general la mayoría de estudios anatómicos inician con la generación de información que sirve como línea de base para futuros estudios aplicativos (Benzing, 1990; Pita y Meneses, 2002; Horres et al., 2007); es decir se orientan a realizar descripciones generales de las
estructuras morfológicas de grupos de especies vegetales de interés particular.
Los estudios anatómico-descriptivos de especies vegetales han tomado mayor relevancia durante las últimas décadas, especialmente en el neotrópico (Tomlinson, 1969; Brighigna et al., 1984; Benzing, 1990; Pita y Meneses, 2002; Horres et al., 2007), sin embargo en el
Ecuador este tipo de investigaciones son todavía incipientes. En la mayoría de estos estudios ha sido muy común utilizar la metodología propuesta por Johansen en 1940, que se denomina cortes a mano alzada, la misma que es muy económica y no requiere de instrumentos sofisticados; pero, también existen otras metodologías que han incorporado instrumentos tecnológicos para mejorar los estudios anatómicos; por ejemplo dentro de estas se menciona la propuesta por D´Ambrogio (1981) quien usa el micrótomo para realizar los cortes; también se menciona las técnicas de fotometría utilizada por Tomlinson (1969), la para la identificación de tricomas, estomas y mesófilos.
En este estudio, se usó la metodología propuesta por Johansen en 1940, a pesar de ser una técnica desarrollada a mediados del siglo anterior, sigue siendo una de las mejores opciones por los buenos resultados que se obtienen con ella, y sobre todo por lo sencilla y económica que resulta.
4.2. Implicaciones de los estudios morfológicos