Disponibleenwww.sciencedirect.com
Revista
Mexicana
de
Biodiversidad
www.ib.unam.mx/revista/ RevistaMexicanadeBiodiversidad88(2017)824–831
Ecología
Diversidad
de
briófitos
en
fragmentos
de
bosque
seco
tropical,
Montes
de
María,
Sucre,
Colombia
Bryophyte
diversity
in
a
tropical
dry
forest
fragment,
Montes
de
María,
Sucre,
Colombia
Stevens
García-Martínez
∗y
Jorge
D.
Mercado-Gómez
DepartamentodeBiologíayQuímica,GrupoEvoluciónySistemáticaTropical,UniversidaddeSucre,Cra28#5-267,Sincelejo,Colombia
Recibidoel16defebrerode2017;aceptadoel14dejuniode2017 DisponibleenInternetel6dediciembrede2017
Resumen
EnlosbosquessecostropicalesdelCaribecolombianoladiversidaddebriófitosesmuypococonocida,apesardeserconsideradocomouno delosfragmentosdebosqueenmejorestadodeconservación.Enelpresenteestudioseanalizóladiversidaddeestegrupodeplantasenlos MontesdeMaría,Sucre,Colombia.En6localidadesfueronidentificadosycuantificadoslosbriófitos,conbaseenestainformaciónseestimó sudiversidadbiológica(␣y)ytaxonómica(+y+).Seregistraronuntotalde1,125vástagospertenecientesa63especies,42génerosy
23familias.Lasfamiliasconmayorriquezafueron:Neckeraceae,Pottiaceae,Sematophyllaceae,LejeuneaceaeyPlagiochilaceae,mientrasqueen losgénerossobresalieron:Fissidens,NeckeropsisySematophyllum.ColosóyMorroafueronlaslocalidadesmásdiversasySanOnofrelamenos diversa,siendoafectadaporlafragmentaciónydestruccióndelbosque.Esteestudioconstituyeelprimeraportesobrelospatronesdediversidad debriófitosenestaregióndelpaís,asícomosusimplicacionesparaconocerlaestructuradelosensamblesenlosecosistemassecostropicales. ©2017UniversidadNacionalAutónomadeMéxico,InstitutodeBiología.EsteesunartículoOpenAccessbajolalicenciaCCBY-NC-ND
(http://creativecommons.org/licenses/by-nc-nd/4.0/).
Palabrasclave: Caribe;Composición;Hepáticas;Musgos;Patrones
Abstract
In the tropicaldry forestsofthe ColombianCaribbean bryophyte diversityislittleknown althoughthis area presentsthe bestpreserved forestfragments.Inthisstudy,weanalyzedbryophyte diversityinthe“MontesdeMaría”,Sucre,Colombia. Bryophyteswereidentifiedand quantifiedin6localities,onthebasisofthisinformationthetaxonomic(+and+)andbiological(␣and)diversitywerecalculated.Atotal
of1,125individualsbelongingto63species,42generaand23families,wereregistered.Species-richfamilieswere:Neckeraceae,Pottiaceae, Sematophyllaceae,LejeuneaceaeandPlagiochilaceae;whilethegenera:Fissidens,Neckeropsis,Sematophyllum,LejeuneaandPlagiochila,were important.ColosóandMorroaarethemostdiverselocalitiesandSanOnofrehastheleastdiversityofall,becauseofforestfragmentationand destruction.Thisstudyconstitutesthefirstcontributionon thediversitypatternsofbryophytesofthisregionofthe country;aswellastheir implicationsforthestructureoftheassemblagesindrytropicalecosystems.
©2017UniversidadNacionalAutónomadeMéxico,InstitutodeBiología.ThisisanopenaccessarticleundertheCCBY-NC-NDlicense
(http://creativecommons.org/licenses/by-nc-nd/4.0/).
Keywords: Caribbean;Composition;Liverworts;Mosses;Patterns
Introducción
Elbosquesecotropicalesunodelosecosistemasmás frag-mentadosydegradadosenelNeotrópicoproductodeldesarrollo
∗Autorparacorrespondencia.
Correoelectrónico:[email protected](S.García-Martínez). LarevisiónporparesesresponsabilidaddelaUniversidadNacional Autó-nomadeMéxico.
delaganaderíayagricultura(Marulandaetal.,2003;Pizanoy García,2014;Rodríguez,Banda,ReyesyEstupi˜nán,2012).En
Colombiaestebosquesehavistoafectadodesdehacemásde
50a˜nosporactividadeshumanasagresivasquereducensuflora
y fauna(Alvaradoy Otero,2015).Ademásdelaperturbación
antrópica,tambiénesotraproblemáticadegranimportanciala faltadeinformacióntaxonómicasobremuchosclados.Por ejem-plo,enplantas,ellinajeenelquesehandesarrolladogranparte de los análisis florísticoshansido lasangiospermas, dejando
https://doi.org/10.1016/j.rmb.2017.10.035
deladogruposcomolos pteridófitosy briófitos,deloscuales
actualmentese desconocesu diversidady distribución en los
bosquesestacionalmentesecosdelpaís.
EnColombia losestudios realizadosalafecha (Aguirre y Avenda˜no,2008;Aguirrey Ruiz,2001;Avenda˜noy Aguirre, 2007,2009;RuizyAguirre,2003;SantosyAguirre,2010;Van Reenen,GriffinyGradstein,1984)sehanenfocadoenla brio-floradezonasaltas(andinayparamuna),dejandoaunladolas tierrasbajasyvallesinterandinos(García,Basilio,Herazo, Mer-cadoyMorales,2016).Enelcasodelbosquesecotropicaldel
Caribe,másespecíficamenteenlosMontesdeMaría,área
con-sideradacomounadelaszonasdebosquesecoenmejorestado
deconservación(PizanoyGarcía,2014),soloseharegistradoel análisisflorísticorealizadoporGarcíaetal.(2016),que registra-ronmásde30especiesensolo3localidadesquecorrespondena 300m2,esdecir,menosdel10%delacoberturaboscosaenlos
MontesdeMaría.Enotraspalabras,esposiblequealampliar
eláreademuestreolariquezasepuedaincrementar
considera-blemente,conlocualsemejoraríaelestadodelconocimiento
sobreungruposubvaloradoenlosecosistemasestacionalmente
secosdeColombia.
Porotraparte,alanalizarlariqueza,composición y abun-danciadeespecies,losestudiossehanenfocadoeneldesarrollo
y aplicación de índices quemiden la entropía y no la
diver-sidadde unacomunidad (Jost,2006).Bajoeste contextoJost (2006)propusoanalizarladiversidadverdaderaorealatravés devariosnivelesqueincluyen:ordencero(0D),quecorresponde alariquezadeespecies(0D=S)yesinsensiblealaabundancia relativa; orden 1 (1D),donde lasespecies sonincluidas
pro-porcionalmentesegún su abundanciaen lacomunidad, y por
último,orden2(2D)quetomaencuentalasespecies
dominan-teso comunes.Estosnivelesposibilitanlacomparacióndela
diversidadde2omásáreasconcondicionesambientales
dife-rentes(Jost,2006),locualnoesposibleconlosíndicesclásicos (Moreno,Barragán,PinedayPavón,2011).
Asimismo, en las últimas décadas la diversidad ha sido
explorada a través de diferentes fuentes de información que
no incluyen necesariamente datos de riqueza y abundancia.
Warwick y Clarke (1995) propusieron calcular la distinción taxonómicapromedio(+)ysuvariación(+).Estosíndices
evalúanladistanciayvariación taxonómicaentrecadaparde
individuosapartirdellistadodeespeciescontodassus catego-ríasonivelestaxonómicos(ClarkeyWarwick,1998);esdecir,
permitenestimarelgradode relacióntaxonómicaentre
espe-cies donde a mayor número de linajes distribuidos entre los
nivelestaxonómicos,másaltosseránlosvaloresdedistinción
taxonómicapromedioy por endeladiversidad. Estos índices
sonimportantesparaconocerladiversidaddesdeotrospuntos
devistaysonrelevantesalmomentodefijaráreasprioritarias paralaconservaciónyparaelmanejodelosrecursosnaturales (García,Moreno y Bello,2014;Moreno etal.,2011),ya que sonsusceptiblesalaspresionesantropogénicas(Morenoetal., 2011;WarwickyClarke,1995).
Lapresenteinvestigacióntuvocomoobjetivoconocerla
com-posición de briófitos y determinar si existen patrones en su
diversidadatravés deíndicesde diversidadreal(␣y ), dis-tinciónyvariaciónenladistincióntaxonómicaen6localidades
quecorrespondenabosquesestacionalmentesecosenMontes
deMaría,departamentodeSucre,Colombia.
Materialesymétodos
LosMontesdeMaríasonconsideradoscomounasubregión
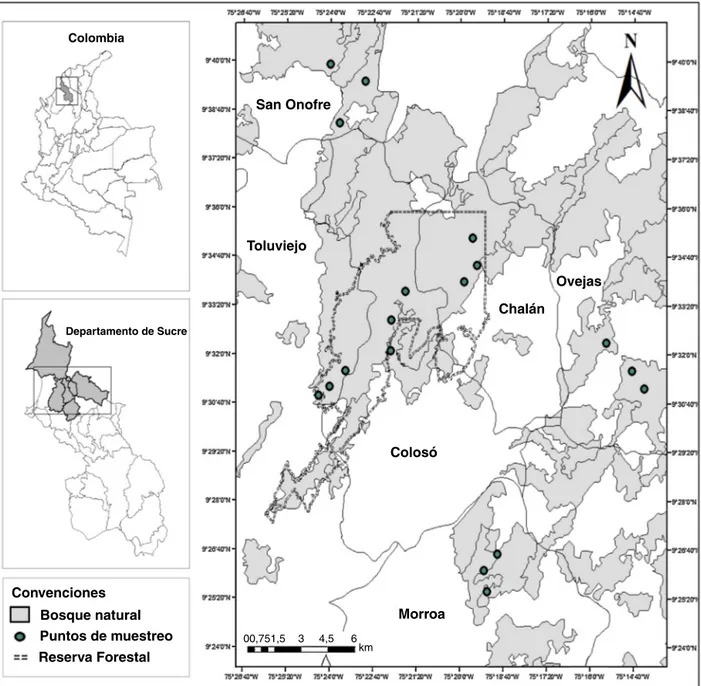
delCaribecolombianoyselocalizanenlapartecentraldelos departamentosdeBolívarySucre,abarcandounaextensióntotal de6,297km2(Aguilera,2013).Desdeelpuntodevista
fisiográ-ficoenellaseencuentraunsistemamonta˜nosoquepertenece
alasestribacionesdelaSerraníadeSanJerónimoyse
distri-buyeentrelosmunicipiosdeColosó,Chalán,Morroa,Ovejas,
partedeLosPalmitos,SanOnofreyToluviejo(Aguilera,2013
[fig.1]).
Los suelosenestaáreasonderivadosde mantosarenosos,
arcillososydecalizasdelMioceno(Halffter,1992).Asimismo, lasmonta˜nasycolinasestánfuertementeescarpadas,ademásde presentarafloramientosrocososyarcillolitasdeorigenmarino (Galván,Gómez,delaOssayFajardo,2009;Promontes,2003). Conrespectoalahidrologíaexisteunared pertenecienteala cuencadelGolfodeMorrosquillo(Verbel,2009)quejuntoala
topografíadelasubregiónformanunsistemademicrocuencas
quedaorigenanumerososarroyosycuerposdeaguadeduración estacionallimitadaporépocasdelluvias(Aguilera,2013).
De igual forma, en sitios como Morroa y Colosó se han
registradoreservorios de aguassubterráneas. En Morroa,por
ejemplo,estasaguassoncaptadasatravésdepozosprofundos
delosacuíferoslibresyconfinadosdelasdiferentes formacio-nesgeológicasdelárea(López,2003),asítambiénenColosó existenreservoriosdeaguassubterráneasquealafechanohan sidoestudiadosenprofundidad(Verbel,2009).
Respectoalclimaelrégimendelluviaesbimodaly
anual-mente se presentan valores desde 1,000 a 1,200mm con
incremento en elsegundosemestre del a˜no; latemporada de
sequíaestácomprendidaentrejunio-julioydiciembre-febrero
(Aguilera,2005,2013; Cuervo, Barbosay delaOssa,1986).
Finalmente,latemperaturavaríaentre26-30◦Cy lahumedad
relativa oscilaentre75% y 85%(Aguilera, 2013;Promontes,
2003).
Lavegetación estáconformada principalmenteporbosque
secotropical,segúnHoldridge(1967)entresuselementosmás
característicosresaltan especiesdeárbolescomo:Ampelocera
edentula, Aspidosperma polyneuron, Brosimum alicastrum,
MyrciafallaxySimiracordifolia(Herazo,Mercado-Gómez,& Mendoza,2017).Noobstante,históricamenteengranpartede lasubregiónladeforestación,laganaderíaextensiva,la
inten-sidadagrícolaylaurbanizaciónhanmodificadolamayoríade
las zonas boscosas, principalmenteen Ovejas y San Onofre.
Teniendo encuentaesta problemáticafueroncreadas2
reser-vasnaturalesprotectoras:ReservaforestalprotectoraSerranía
deCorazayMontesdeMaría,departamentodeSucre,yel
San-tuariodefaunayfloraLosColorados,departamentodeBolívar (Aguilera,2013).
Paralos muestreosse realizaronvisitas previasen
diferen-tes localidades de los Montes de María conel fin de ubicar
áreasconamplias coberturasde vegetación,sombra,
Colombia
San Onofre
Toluviejo
Ovejas
Chalán
Colosó
Convenciones
Bosque natural
Puntos de muestreo Reserva Forestal
Morroa
00,751,5 3 4,5 6
km Departamento de Sucre
Figura1.Ubicacióngeográficadeláreadeestudio;localidadesdemuestreoyposicióndelossitiosderecolectaenlosMontesdeMaría(Sucre-Colombia).
en otraspalabras, sitiosconlascondiciones mínimas para el
crecimientodelos briófitos(Garcíaetal.,2016).Seeligieron 6sitiosquesedistribuyeronenlosmunicipiosdeColosó, Cha-lán,Toluviejo,Morroa,OvejasySanOnofre(fig.1),loscuales fuerondenominadossegúnsumunicipio.Enestosseubicaronal azar3transectoslinealesde100×10myserecolectaron ejem-plaresdebriófitossegúnlasespecificacionesdadasporGarcía etal.(2016)yOrrego(2005);tambiénsetomarondatoscomo laaltitud,coordenadasgeográficasyserealizóunaestimación delacoberturaatravésdeunaplantillacuadriculadadeacetato
de30×20cm2 (cuadrosde1×1cm)modificadadeIwatsuki
(1960).
Unavezfinalizadoelprocesoderecolectaelmaterialfue her-borizadosegúnlosprocedimientosconvencionalesparabriófitos (Garcíaetal.,2016)yposteriormenteidentificadosmediantelas
claves dicotómicas en Churchill y Linares (1995),Gradstein
(1991), Gradstein, Churchill y Allen (2001), Sharp, Crum y Eckeckeckel (1994) y Uribe y Aguirre (1997). Adicional-mente,seutilizóbibliografíaespecializadaparaCalymperaceae (Reese, 1993),Frullaniaceae (Gradstein y Uribe, 2011), Pot-tiaceae (Zander, 1993) y Lejeuneaceae (Gradstein, 1994); de
igualforma, enlos casosdondefue necesario,losejemplares
fueron corroborados por expertos. Finalmente, fueron
con-sultadas bases de datos en línea como Bernal, Gradstein y
Celis 2016,Boyleetal.(2013) y MissouriBotanical Garden (http://www.tropicos.org/) para actualizar la nomenclatura de lasespecies.
deWhittaker(Moreno,2001)ylasimilitudpordistanciaa tra-vésdeunamatrizentrelocalidadesdemuestreo,alacualfue aplicadoelcoeficientedesimilitudBray-Curtis,ysobreéstese realizóunanálisisclústerparaagruparlasáreasdemuestreopor sucomposiciónflorística.Lamatrizdesimilitudseconstruyó apartirdelosdatosdeabundanciarelativa,conprevioajustea raízcuartaparadisminuirelefectodelasespeciesdominantes (comunes)encomparaciónconlasespeciesraras(Field,Clarke yWarwick,1982).Loscálculosparadiversidad␣serealizaron
conelprogramaPRIMER6.1.7(ClarkeyGorley,2006)ypara
conPAST(Hammer,HarperyRyan,2001).
Paraestimarla+ y+ se usaronlos sistemasde
clasifi-caciónparamusgosyhepáticasdeGoffinet y Shaw(2009)y
Söderströmetal.,(2015),conlocualseestableciólajerarquía
taxonómicaparacadaespecie;posteriormenteestainformación
fueanalizadapormediolafunciónTAXDTDESTdelsoftware
PRIMER6.1.7(ClarkeyGorley,2006).Paraestainvestigación laslocalidadesconvaloresaltosde+yconmenor+sonlas demayordiversidadtaxonómica(ClarkeyWarwick,2001).
Resultados
Seregistraron63especies,42génerosy23familias(tabla1).
Los musgosestuvieron mejorrepresentados con38 especies,
29génerosy18familias.Lashepáticaspresentaron25especies,
13géneros y 5 familias; de los taxonesidentificados en este
estudio21sonnuevosregistrosparaeldepartamentodeSucre
y9paraelCaribecolombiano(tabla1).
Con17especiesLejeuneaceaefuelafamiliamásrica,seguida porSematophyllaceae(6especies),FissidentaceaeyPottiaceae
(con 5 cada una). Los géneros sobresalientes fueron:
Lejeu-nea(8 especies),aestelesiguieronFissidens(6) Plagiochila
ySematophyllum(3cadauno);losdemáspresentarondeunaa 2especies.Enrelaciónconlaespecielasmáscomunesy domi-nantesfueronCalympereserosumyHyophilainvoluta,mientras que12especiesfueronrarasconunsoloregistroyexclusivas deunalocalidad(tabla1).
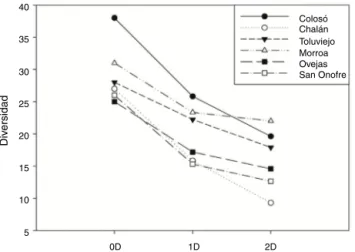
El análisisde diversidad ␣ reveló quelaslocalidades son significativamente diferentes en el número de especies (0D). ColosóyMorroapresentaronlosvaloresmásaltosderiquezade
especies,siendoColosó1.16vecesmásdiversoqueToluviejo
y1.5vecesmásqueOvejasalcompararlosvaloresde1D.En
cuantoaChalánySanOnofreladiversidaddisminuyóen∼61%
encomparaciónconColosó.Estamismatendenciaseobservó
encuantoa2D,dadoqueColosóyMorroaregistraronelmayor
númerodeespeciesdominantes,ySanOnofrees lalocalidad
conmenordominancia(fig.2).
Conrespectoalosresultadosdediversidad,enelanálisis declasificaciónsedistinguierongruposdelocalidadesconun niveldesimilitudmayoral55%(fig.3):elgrupo1conformado porMorroa-Ovejasyelgrupo2integradoporColosó-Toluviejo,
mientrasqueSanOnofreyChalánsemuestranindependientes
aestasagrupaciones.Porsuparte,alaplicarelíndicede Whit-takerseobtuvountotalde1.185comunidadesvirtualesdelas 6posiblesenelsitiodemuestreo.Enunanálisispareadoentre localidadesparaestablecerelporcentajederecambiodeespecies
necesario paraconformar unasolacomunidad, los resultados
Tabla1
Listadodeespeciesysudistribucióneneláreadeestudio.
Familias Especies Localidades
Musgos
Bryaceae Anomobryumconicum (Hornsch.)Broth.
Tv
Mielichhoferiamegalocarpum
(Arn.)Mitt.
Cl,Ch,Tv
Bartramiaceae Philonotisuncinata(Schwägr.) Brid.
Mr,Ov
Calymperaceae CalympereserosumMüll.Hal. Cl,Ch,Tv,Mr, Ov,So Hookeriaceae HookeriaacutifoliaHook.y
Grev.
Cl,Ch,Tv,Mr, Ov,So Fabroniaceae Fabroniaciliaris(Brid.)Brid. Cl,Tv,Mr,Ov
Fissidentaceae FissidensprionodesMont. Cl,Ch,Tv,Mr,Ov
FissidensflaccidusMitt. Ch,Tv,So
FissidensdissitifoliusSull. Cl,Ch,Tv,Mr, Ov,So
FissidenssteereiGrout Cl
FissidenspellucidusHornsch. Ch,Tv,Mr,Ov Hypnaceae Chryso-hypnumdiminutivum
(Hampe)W.R.Buck*
Cl,Tv,Mr,So
Leucomiaceae Leucomiumstrumosum
(Hornsch.)Mitt.
Cl
Meteoriaceae Meteoriumnigrescens(Hedw.) Dozy&Molk.*
Tv,Ov,So
Thuidiaceae ThuidiumtomentosumSchimp. Cl,Ch,Mr,So
Pelekiuminvolvens(Hedw.)A. Touwa
Cl,Ch,Tv,Mr,Ov
Pottiaceae TortellaalpicolaDixona,b Cl,Tv
Didymodonsp. Tv,Mr
Dolotortulamniifolia(Sull.)R.H. Zandera,b
Ov,So
Hyophilainvoluta(Hook.)A. Jaeger
Cl,Tv,Mr,Ch, Ov,So
Hyophiladelphusagrarius
(Hedwig)R.H.Zander
Cl
Sematophyllaceae Trichosteleumpapillosum (Hornsch.)A.Jaegera,b
Mr
Trichosteleumfluviale Cl,Mr
Sematophyllumsubsimplex
(Hedw.)Mitt.a
Cl,Mr
Sematophyllumswartzii
(Schwägr.)W.H.WelchyH.A. Cruma
Mr
Sematophyllumsubpinnatum
(Brid.)E.Brittona
Ch
Taxitheliumplanum(Brid.)Mitt.* Ch,So
Stereophyllaceae Eulacophyllumcultelliforme
(Sull.)W.R.BuckyR.R.Irelan
Cl,Tv,Mr,Ov
Neckeraceae Neckeropsisundulata(Hedw.) Reichardt
Cl,Tv,Ov,So
Neckeropsisdisticha(Hedw.) Kindb.a
Cl,Ch,So
Porotrichumsubstriatum
(Hampe)Mitt.
Cl,Tv,Ov,So
Porotrichodendronlindigiia Cl,Ch,Tv,Mr,Ov Octoblepharaceae OctoblepharumalbidumHedwa Cl,Tv,Mr,Ov
Pilotrichaceae Callicostellapallida(Hornsch.) Ångströma
Cl,Ch,Tv
Cyclodictyonalbicans(Hedw.) Kuntze
Cl,Ch
Rutenbergiaceae Pseudocryphaeadomingensis
(Spreng.)W.R.Buck
Ch,Mr,Ov,So
Splacnobryaceae Splachnobryumobtusum(Brid.) Müll.Hal.
Tabla1(continuación)
Familias Especies Localidades
Hepáticas
Frullaniaceae FrullaniacuencensisTaylor
a Cl,Mr,Ov,So
Frullaniariojaneirensis(Raddi) Sprucea
Cl,Ch,So
Lejeuneaceae Archilejeuneaconf.badia So
Blepharolejeuneaconf. Incongruaa
So
Bryopterisdiffusa(Sw.)Neesa Mr
Caudalejeunealehmanniana
(Gottscheetal.)A.Evansa,b
So
Ceratolejeuneaconf.Confusaa So
Cheilolejeuneasp. Ch,Tv,Mr,Ov
Mastigolejeuneaplicatiflora
(Spruce)Steph.
Cl,Ch,Mr
Mastigolejeuneaauriculata
(WilsonyHook.)Schiffner
Cl
Microlejeuneasp. Cl,Ch
LejeuneadeplanataNees Cl,Tv
Lejeuneaflava(Sw.)Nees Cl,Ch,Tv,Mr, Ov,So
LejeuneatrinitensisLindenb Cl,Ch,Tv,So
Lejeuneasp.1 Cl
Lejeuneasp.2 Cl,Ch,Mr,So
Lejeuneasp.3 Cl,Ch,Tv,So
Lejeuneasp.4 So
Lejeuneasp.5 Ch,So
Marchantia-ceae
MarchantiachenopodaL. Cl,So
MarchantiainflexaNeesy Mont.a,b
Cl,Mr,So
Plagiochila-ceae
PlagiochilafuscoluteaTaylor Cl,Mr,Ov
Plagiochilasp.1 Cl,Ch,Tv,Mr,Ov
Plagiochilasp.2 Ch
Targioniaceae TargioniahypophyllaL.a Ov
Ch:Chalán;Cl:Colosó;Mr:Morroa;Ov:Ovejas;So:SanOnofre;Tv:Toluviejo.
aNuevoregistroparaSucre
b NuevoregistroparaelCaribecolombiano.
Colosó Chalán Toluviejo Morroa Ovejas San Onofre 40
35
30
25
20
Div
ersidad
15
10
5
0D 1D 2D
Figura2.Diversidadalfa(0D,1Dy2D)debriófitosenlossitiosdemuestreos.
fueronsimilaresalosobservadosenlosanálisismultivariados.
MorroayOvejassolorequieren deun33%derecambiopara
establecersecomounasolacomunidad,esdecir,tienenun
por-centaje de67% de similitud.Enel casodeColosó-Toluviejo
presentanun61%desimilitudoun39%derecambiode
espe-cies.Porelcontrario,ChalánySanOnofresemuestrancomo
20
40
60
80
100
So Ch C1 T1 Mr Ov
Similar
idad de Bray-Curtis
Figura3.Dendrogramadeclasificación(clúster)basadoenlamatrizdesimilitud deBray-Curtisentrelocalidadesdemuestreo.
gruposindependientespormostrarelmayorporcentaje
reque-ridoparaconformarunacomunidad.SanOnofrenecesitaentre
53-57%derecambiodeespeciescontodaslaslocalidadespara
conformaralmenosunacomunidad,mientrasqueChalánentre
un47%a57%.
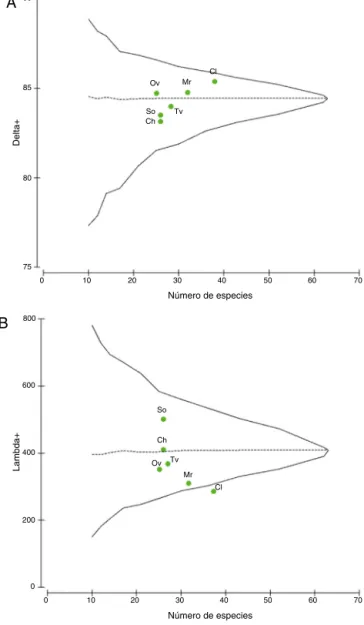
Enrelación conlosanálisisde distincióntaxonómica
pro-medio, la gráficade + muestra quelos valores se ubicaron
dentrodeloscontornosprobabilísticosdeladistribución
espe-rada (fig. 4A). Los valores de Colosó, Morroa y Ovejas se
encontraron cercanos a la media estimada; mientras que los
deToluviejo,ChalánySanOnofreseubicaronpordebajodel
límiteinferiordeconfianzaydistantesdelamediaesperada,a consecuenciadelabajacomplejidadtaxonómica(fig.4A).Las
localidadesconmayordistincióntaxonómicafueronColosóy
Morroa (tabla2),siendo Colosó elde mayorcobertura
taxo-nómica.UnresultadoimportanteesqueOvejas,apesardeser
unadelaslocalidadesconmenordiversidad,obtuvounamayor
distincióntaxonómicapromedioencomparaciónconToluviejo,
ChalánySanOnofre(tabla2).
En cuanto a la variación de la distinción taxonómica los
datosse localizaron dentrodelos contornosprobabilísticos y
pordebajodelamediaesperada,exceptoColosó,queseubicó
fueradelrangoprobabilístico,alpresentarlamenor variabili-dad ensu estructura taxonómica(fig.4B).El valor más alto
fue alcanzado por San Onofre y Chalán quienesse ubicaron
porencima delpromedioesperado,adiferenciadelasdemás
localidades(tabla2).
Discusión
Losresultadosmuestranquelascomunidadesdebriófitosno
sonhomogéneasensucomposición,abundanciaydominancia
de taxones. Los gruposse caracterizaronpor presentarpocas
especiesdominantes,conabundanciasintermediasyunnúmero
Mr
Mr Cl
Cl Ov
Ov Tv
Tv Ch
Ch So
So
70 60 50 40 30 20 200
400 600 800
10 0
70 60 50 40 30 20 10 0
0
Número de especies
Número de especies
90
85
80
75
Delta+
Lambda+
A
B
Figura4.Distincióntaxonómicapromedio(A)yvariacióntaxonómica(B)de briófitosenlaslocalidadesdemuestreo.Lalíneahorizontalsegmentada repre-sentalamediaesperadaylacontinua,loslímitesdeconfianza(95%).Lospuntos simbolizanlosvaloresobservadosparacadasitio.Convencionesenlatabla1.
Elanálisisdediversidad␣revelóqueColosóyMorroa
fue-ronmásdiversosencomparaciónconlasotraslocalidades,lo
cualpuedeserexplicadoporlossiguientesargumentos.Porun lado,estánlasdiferenciaslocales conrespectoalas variacio-nesambientalescomolahidrología,coberturavegetalsuperior, suelo,aguaydisponibilidaddeluz.Porejemplo,Colosóposee
7microcuencas comoresultadodelafiltracióndeaguas
sub-terráneas(Verbel,2009),permitiendoquelaofertahídricasea
permanentesobreestaszonas,proporcionandounhábitatideal
para su colonización. Delmismo modo, estas características
podríanestarcompartidasconMorroa,yaqueelrecursohídrico semantieneapartirdelosmantosacuíferosqueexistenenesta zona(Promontes, 2003).Empero,este factorno es constante paralasdemászonasysuduraciónesmuycorta,locual
deter-minalasespeciesquepuedensobreviviraperíodosdehumedad
largosocortos.También,estánlascolinasymonta˜nas(Serranía
deCoraza),quejuntoalosbancos denieblas (alamanecer y
Tabla2
Distinciónyvariacióntaxonómicadebriófitosenlossitiosdemuestreos.
Sitiosdemuestreo S + +
Colosó 38 85.38 234.93
Chalán 26 84.12 410.08
Toluviejo 27 83.42 339.55
Morroa 32 84.73 298.28
Ovejas 25 84.40 317.09
SanOnofre 26 83.14 500.92
+:distincióntaxonómicapromedio;+:variacióndeladistincióntaxonómica;
S:riquezadeespecies.
atardecer)queprovienendelgolfopuedenincrementarla
con-centracióndehumedad(Garcíaetal.,2016),yporlotantola ofertadeaguanecesariaparaelcrecimientodebriófitos.
Otrofactorquepuedeafectarladiversidaddeestosgruposde plantasesladeforestación,talcomofueobservadaenOvejasy SanOnofre,áreasconvaloresdediversidadinferioresaMorroa yColosó.Ladeforestaciónpuedeocasionarvariosprocesosque afectanpuntualmentealosbriófitos,talescomoreduccióndela
sombra,disminucióndelahumedadylaofertadeangiospermas
quesirven como forófitos, elementos queenconsideración a
estaszonasMorroayColosópresentansobregranpartedela˜no
y porlotanto unhábitatadecuado paraelcrecimientode un
mayornúmerodelinajesdiferentes.
En cuanto a diversidad  el recambio promedio (1.185)
entrelas6localidades mostrópoca variación,porlotanto,la diferenciadeespeciesentreunalocalidadaotranoesmuy repre-sentativa;estosresultadossoncoherentesconelanálisisclúster,
considerandoquelasimilitudfuemayora55%,indicandobajo
recambiodeespecies.Estabajadiversidadbetaconlleva
con-siderarquelasvariablesambientalestiendenaserhomogéneas
en el área de estudio y quepodría existir unasimilitud
eco-lógicaentremicrohábitats. Eneste sentido,lascaracterísticas ambientalessimilaresentrelaslocalidadesparecentenerunrol
importanteen estosresultados; porejemplo, lasemejanza de
Toluviejo-Colosópodríaestarrelacionadaporlacontinuidadde susbosques,yaqueambospertenecenalareservaforestal.En
elcasodeMorroa-Ovejassu similitudpuedeserelreflejo de
algúntipodeconexiónhistóricaantesdelallegadadelhombre aláreadeestudio.
Losanálisisde+ y+ indicanqueColosóyMorroason
lossitiosdemayordiversidadtaxonómica,demaneraquelos
ensamblajesestáncompuestosporunmayornúmerode
géne-rosyfamiliasqueestánmenosrelacionadostaxonómicamente
entresí(Webb,Ackerly,McPeekyDonoghue,2002).Esdecir, quelasvariablesabióticascomolaprecipitaciónylahumedad relativa,ylasbióticascomomayorcoberturadelavegetación,
han generado un ambiente apropiado para que coexistan un
mayor númerode especiesquenoestánrelacionadas
taxonó-micamente(Grass,Brandl,Botzat,NeuschulzyFarwig,2015). Porotraparte,losvaloresde+entreChalányToluviejofueron
muysimilares(tabla 2); sinembargo,en Chalánhubo mayor
variación de la distincióntaxonómica, loquesugiere que en
relacionados;demaneraque,haciaChalán,seincrementala
pro-babilidaddeencontrarespeciesmásemparentadas,delmismo
géneroofamilia,yconmayorhomogeneidadtaxonómica. Es
tambiénnotablequeOvejasobtuvomayordistinciónymenor
variación,portanto,suestructurataxonómicaesmásdiversay
susespeciesestánpocorelacionadastaxonómicamenteentresí
encomparaciónconotraslocalidades.
Bajo el contexto anterior, las localidades analizadas que
presentaron altos valores de distinción taxonómicapromedio
tienen mayor número de especies pertenecientes a distintos
taxones superiores. Por tanto, se podría sugerir que Colosó,
Morroa y Ovejas son los sitios que aumentan la
variabili-dad y diversidad taxonómica de este grupo en el área de
estudio.
EnrelaciónconSan Onofrelabajadiversidadtaxonómica
encontrada en esta localidad reflejó la presencia de
condi-ciones ambientales adversas y la distribución similar de las
especiesenlascategoríastaxonómicas(pocaamplitud
taxonó-mica),ocasionadaposiblementeporlaintervenciónantrópicay eldeterioroprogresivodelambiente.EnestesentidoClarkey Warwick(1998)afirmanquecuandounáreaostenta
alteracio-nesambientales lasespeciesse encuentranmuyrelacionadas,
pertenecientes almismo géneroo familia,y en consecuencia
loshábitatsposeenbajosvaloresdedistinción.Engeneral,los
índices taxonómicos coincidieroncon los índices ecológicos,
ya que el comportamiento de la + y + permiten sugerir
queladiversidadtaxonómicadebriófitosseveafectadatanto
porcondicionesdelhábitatcomoporfactoresantropogénicos
(González,Uma˜na,Álvarez,StevensonySwenson,2014; Prin-zingetal.,2008).
Enotrostérminos,laheterogeneidaddelhábitatactúacomo unfiltroambientalquedefineestructuralmentelosmecanismos deensamblajesdeestostaxones.Esdecir,quelasvariaciones
microambientalessonlasquedeterminan lospatrones
estruc-turalesdelascomunidadesde briófitoseneláreade estudio.
Sin embargo, la deforestacióny sus causas son factores que
ocasionanfuertesalteracionesenlacalidaddelhábitato
con-diciones ambientales que modifican los grupos de briófitos.
Ental sentido,se considera quelascondiciones de
perturba-ciónyvariacionesmicroambientalesenlasqueseencuentran
los fragmentos de bosque en la subregión Montes de María
sondeterminantesenlaestructuracióndelos gruposde estos
taxones.
Montes deMaría, talcomo ha sido descrito por Pizanoy
García (2014),es unazona altamente conservada,lo cualha
generadounagrandiversidaddeplantas,tales elcasodelos
briófitos,organismosque inicialmentehabían sidopoco
esti-madosenestaárea;noobstante,coneltrabajodeGarcíaetal. (2016)ylosnuevosreportesdadosenesteestudio,ademásde
incrementarmásdel50%conrespectoaloreportadopor
Gar-cía,tambiénreportavariasespeciescomoregistrosnuevospara
ColombiayelCaribe.
Segúnloanterior,esimportanteentenderqueotraszonasde
bosquesecoenelCaribecolombianotambiénpuedenteneruna
altariquezadeestosorganismospocoestudiadosenestos
bos-ques,dadoqueseasumíaquelascondicionesdeestosbosques
nogenerabanlasbasesparasucrecimiento.
Agradecimientos
Alproyecto“Análisisflorísticoyfitogeográficosobre
relic-tos de bosque seco tropical en el departamento de Sucre
(Colombia)”,financiadoporlaDivisióndeInvestigacionesde
laUniversidaddeSucre.AEdgarLinaresyalInstitutode
Cien-ciasNaturalesdelaUniversidadNacionaldeColombiaporel
soporterecibidoduranteeldesarrollodeestainvestigación.
Referencias
Aguilera,M.(2005).LaeconomíadeldepartamentodeSucre:ganaderíaysector público.EnDocumentosdetrabajosobreeconomíaregional.Bogotá:Banco delaRepública.
Aguilera,M.(2013).MontesdeMaría:unasubregióndeeconomíacampesina yempresarial.EnDocumentosdetrabajosobreeconomíaregional.Bogotá: BancodelaRepública.
Aguirre,J.yAvenda˜no,K.(2008).MusgosenlaregiónCaribe.EnO.Rangel (Ed.),Colombiadiversidadbióticavi:riquezaydiversidaddemusgosy líquenesdeColombia(pp.55–59).Bogotá:InstitutodeCienciasNaturales. Aguirre,J.yRuiz,C.(2001).Composiciónflorísticadelabriofloradela Serra-níadelPerijá(Cesar-Colombia): distribuciónyecología.Caldasia,23, 181–201.
Alvarado,D.yOtero,J.(2015).Distribuciónespacialdelbosquesecotropical enelValledelCauca,Colombia.ActaBiológicaColombiana,20,141–153. Avenda˜no,K.yAguirre,J.(2007).Losmusgos(Bryophyta)delaregiónde
SantaMaría-Boyacá(Colombia).Caldasia,29,59–71.
Avenda˜no,K.yAguirre,J.(2009).LabriofloradelaserraníadePerijá (Cesar-Colombia).EnO.Rangel(Ed.),Colombiadiversidadbióticaviii:mediay bajamonta˜nadelaserraníadelPerijá(pp.189–221).Bogotá:Institutode CienciasNaturales.
Bernal, R., Gradstein, R. y Celis, M. (2016). Catálogo de plantas y líquenes de Colombia [consultado 2 Nov 2016]. Disponible en: http://catalogoplantasdecolombia.unal.edu.co/
Boyle,B.,Enquist,B.,Raygoza,J.,Hopkins,N.,Lu,Z.,Matasci,N.,etal. (2013).Thetaxonomicnameresolutionservice:anonlinetoolforautomated standardizationofplantnames[consultado30Oct2016].Disponibleen: http://tnrs.iplantcollaborative.org/howcite.html.
Churchill,S.yLinares,E.(1995).ProdromusBryologiaeNovo-Granatensis. IntroducciónalaflorademusgosdeColombia.Bogotá:BibliotecaJosé JerónimoTriana.
Clarke,K.yGorley,R.(2006).Primerv6:usermanualandtutorial.Plymouth, UK:Primer-E.
Clarke,K.yWarwick,R.(1998).Ataxonomicdistinctnessindexandits statis-ticalproperties.JournalofAppliedEcology,35,523–531.
Clarke,K.yWarwick,R.(2001).Afurtherbiodiversityindexapplicableto specieslists:variationintaxonomicdistinctness.MarineEcologyProgress Series,216,265–278.
Cuervo,A.,Barbosa,C.ydelaOssa,J.(1986).Aspectosecológicosyetológicos deprimatesconénfasisenAlouattaseniculus(Cebidae),delaregiónde Colosó,serraníadeSanJacinto(Sucre),costanortedeColombia.Caldasia, XIV,68–70.
Field,J.,Clarke,K.yWarwick,R. (1982).Apracticalstrategyfor analy-singmultispeciesdistributionspatterns.MarineEcologyProgressSeries, 8,37–52.
Galván,S.,Gómez,H.,delaOssa,J.yFajardo,A.(2009).Biodiversidaden eláreadeinfluenciadelaestaciónprimatesdeColosó,Sucre,Colombia. RevistaColombianadeCienciaAnimal,1,98–121.
García,S.,Basilio,H.,Herazo,F.,Mercado,J.yMorales,M.(2016).Diversidad debriófitosenlosMontesdeMaría,Colosó(Sucre,Colombia).Colombia Forestal,19,41–52.
Gimingham, C. y Birse, E. (1957). Ecological studies on growth-form in bryophytes:I.Correlationsbetweengrowth-formandhabitat.Journalof Ecology,45,533–545.
Goffinet,B. yShaw,A.(2009). Bryophytebiology.NewYork:Cambridge UniversityPress.
González,S.,Uma˜na,M.,Álvarez,E.,Stevenson,P.ySwenson,N.(2014). Phylogeneticalphaandbetadiversityintropicaltreeassemblagesalong regional-scaleenvironmentalgradientsinnorthwestSouthAmerica.Journal ofPlantEcology,7,145–153.
Gradstein,S.(1991).AkeytotheColombianspeciesofholotipousLejeuneaceae (Hepaticae).Caldasia,16,429–438.
Gradstein,S.(1994).“Lejeuneaceae:Ptychantheae,Brachiolejeuneae”.Flora Neotropica.NewYork:NewYorkBotanicalGardenPress.
Gradstein,S.R.,Churchill,S.yAllen,N.(2001).GuidetotheBryophytesof TropicalAmerica.NewYork:NewYorkBotanicalGardenPress. Gradstein,R.yUribe,J.(2011).AsynopsisoftheFrullaniaceae
(Marchan-tiophyta)fromColombia.Caldasia,33,367–396.
Grass,I.,Brandl,R.,Botzat,A.,Neuschulz,E.yFarwig,N.(2015). Contras-tingtaxonomicandphylogeneticdiversityresponsestoforestmodifications: comparisonsoftaxaandsuccessiveplantlifestagesinSouthAfricanscarp forest.PlosOne,10,1–20.
Halffter,G.(1992).LadiversidadbiológicadeIberoaméricaI.Xalapa:Instituto deEcología,A.C.
Hammer,Ø.,Harper,D.yRyan,P.(2001).Past:paleontologicalstatistics soft-warepackageforeducationanddataanalysis.PalaeontologiaElectronica, 4,1–9.
Herazo,V.F.,Mercado-Gómez,J.D.y Mendoza, H.(2017).Estructuray composiciónflorísticadelbosqueseco tropicalenlosMontesdeMaría (Sucre-Colombia).CienciaenDesarrollo,8,79–90.
Holdridge,L.(1967).Lifezoneecology.SanJosé,CostaRica:TropicalScience Center.
Iwatsuki,Z.(1960).TheepiphyticbryophytecommunitiesinJapan.Journalof theHattoriBotanicalLaboratory,22,159–339.
Jost,L.(2006).Entropyanddiversity.Oikos,113,363–375.
López,A.(2003).EOT:Esquemadeordenamientoterritorialdiagnóstico inte-graldelterritorio(2009).MunicipiodeMorroa,DepartamentodeSucre: AlcaldíadeMorroa.
Marulanda,L.,Uribe,A.,Velásquez,P.,Montoya,M.,Idárraga,A.,López,M., etal.(2003).Estructuraycomposicióndelavegetacióndeunfragmentode bosquesecoenSanSebastián,Magdalena(Colombia).I.Composiciónde plantasvasculares.ActualidadesBiológicas,25,17–30.
Moreno,C.(2001).Métodosparamedirlabiodiversidad.Zaragoza,Espa˜na: ManualesyTesisSEAZaragoza.
Moreno, C., Barragán, F., Pineda, E. y Pavón, P. (2011). Reanalizando ladiversidad alfa: alternativasparainterpretary comparar información sobrecomunidades ecológicas.Revista Mexicanade Biodiversidad,82, 1249–1261.
Orrego,O.(2005).BriófitosdeCaldas:laReservadePlanalto.Boletín Científico-CentrodeMuseos-MuseodeHistoriaNatural,9,31–50.
Pizano,C.yGarcía,H.(2014).ElbosquesecotropicalenColombia.Bogotá: InstitutodeInvestigacióndeRecursosBiológicosAlexandervonHumboldt (IAVH).
Prinzing,A.,Reiffers, R.,Braakhekke,W.,Hennekens,S.,Tackenberg,O., Ozinga,W.,etal.(2008).Lesslineages–moretraitvariation: phylogene-ticallyclusteredplantcommunitiesarefunctionallymorediverse.Ecology Letters,11,809–819.
Promontes.(2003).ProgramadedesarrolloypazdelosMontesde María-Promontes”. Bogotá: PNUD, Corporación Territorios, Universidad de Cartagena,Cartagena.
Reese,W.(1993).“Calymperaceae”.InFloraNeotropica.NewYork:NewYork BotanicalGardenPress.
Rodríguez,G.,Banda,K.,Reyes,S.yEstupi˜nán,A.(2012).Listacomentada delasplantasvascularesdebosquessecosprioritariosparalaconservación enlosdepartamentosdeAtlánticoyBolívar(Caribecolombiano).Biota Colombiana,13,7–39.
Ruiz,C. yAguirre,J.(2003). Lascomunidades de briófitosysurelación con laestructura de lavegetación fanerogámica en elgradiente altitu-dinaldelaserraníadelPerijá(Cesar-Colombia).TropicalBryology,24, 101–113.
Santos,G.yAguirre,J.(2010).Losmusgosdelaregión delas Quinchas (MagdalenaMedio,Colombia).Caldasia,32,257–273.
Sharp,A.,Crum,H.yEckeckeckel,P.(1994).ThemossfloraofMéxico.Nueva York:NewYorkBotanicalGarden.
Söderström,L.,Hagborg,A.,VonKonrat,M.,Bartholomew-Began,S.,Bell, D.,Briscoe,L.,etal.(2015).Worldchecklistofhornwortsandliverworts. PhytoKeys,59,1–828.
Uribe,J.yAguirre,J.(1997).ClaveparalosgénerosdehepáticasdeColombia. Caldasia,19,13–27.
VanReenen,G.,Griffin, D.y Gradstein, S.(1984). Briófitosdeltransecto Buritacá-LaCumbre,SierraNevadadeSantaMarta,Colombia.EnT.Vander HammenyP.Ruiz(Eds.),LaSierraNevadadeSantaMarta(Colombia), transectoBuritaca-LaCumbre.Estudiosdeecosistemastropandinos(pp. 177–183).Berlín:Cramer.
Verbel,C.(2009).EOT:esquemadeordenamientoterritorialdiagnóstico inte-graldelterritorio(2009).MunicipiodeColosó,DepartamentodeSucre. (Versiónpreliminar).AlcaldíadeColosó.
Warwick,R.yClarke,R.(1995).New’biodiversity’measuresrevealadecrease intaxonomicdistinctnesswithincreasingstress.MarineEcologyProgress Series,129,301–305.
Webb,C., Ackerly,D.,McPeek, M.yDonoghue, M.(2002).Phylogenies andcommunityecology.AnnualReviewofEcologyandSystematics,33, 475–505.