División de Ciencias Biológicas y de la Salud
CLONACIdN
Y CARACTERIZACIóN DEL CANDIDATO
A
PORINA OmpF
DE
Salmonella typhi
9,12
Vi:d
COORDlNAClm DE SE!W!Girk :
l)ocuAlENrALEs
-
fWOTEGJ4T E S I S
QUE
PARA
OBTENER EL GRADO DE:
MAESTRA EN
BIOLOGÍA
EXPlERIMENTAL
PRESENTA:
MARIA
TERESA
MATA
GONZÁLEZ
TUTOR: DR. VIANNEY FRANCISCO ORTÍZ NAVARRETE
M6xic0, D.F., 1999
Índice de figuras
indxe de tablas
Principales Abreviaturas
INTRODUCCT~N
PLANTEAMIENTO DEL PROBLEMA
OBJETIVOS
DESARROLLO EXPERIMENTAL
A. Cepas bacterianas
B. Plásmidos
C. Preparación de bacterias competentes
D. Amplificación del gen candidato a la porina OmpF
D. 1 Diseño de los cebadores
D.2 Obtención del DNAc de S. @phi 9,12 Vi:d
D.3 PCR para la amplificación del probable gen OmpF
E. Purificación del producto de PCR
F. Clonación del producto de PCR
G. Detección del probable gen OmpF
G. 1 Aislamiento y transferencia de colonias
G.2 Hibridación con sondas radiactivas
G.3 Detección de las colonias portadoras del candidato OmpF
I. Secuenciación del gen
J. Extracción de porinas mediante el método de Nikaido
K. Purificación de la proteína recombinante
L. Caracterización del producto del gen OmpF
L. 1 Bioquímica
L.2 Tnmunoquímica
L. 3 Topológica
M. Análisis comparativo de la estructura primaria
RESULTADOS
l. Obtención del gen candidato a porina OmpF de S. typhi
Y,
12 Vi:d2. Clonación del candidato OmpF
3. Tdentificación del probable gen OmpF
4. Análisis del producto del probable gen OmpF
5. Análisis comparativo de la estructura primaria
INDICE DE FIGURAS
Figura 1.
Figura 2.
Figura 3.
Figura 4.
Figura 5.
Figura 6.
Figura 7.
Figura 8.
Figura 9.
Figura 10.
Figura 11.
Figura 12.
Figura 13.
Figura 14.
Organización de la envoltura de las bacterias gram-negativas
Estructura del monómero de la porina OmpF de Escherichia coli
Regulación de la porina OmpF en E. coli
Mapa del plbmido pUC19
DNA cromosomal de S @phi 9,12 Vi:d
Gen OmpC y candidato a gen OmpF
Plhsmido pUC19 digerido con Sma I
Hibridación de colonias acarreadoras del gen candidato OmpF de
.S.
typhiClonación del gen probable OmpF en pUC 19
Secuencia del gen probable OmpF
Homología y patrón de restricción del probable OmpF y OmpC
Proteína recombinante
Expresión de la proteína recombinante OmpF de S. typhi en b'. coli UH302
Caracterización de la proteína recombinante
iNDICE
DE TABLAS
Tabla 1. Homología entre la probable porina OmpF de S. typhi y la porina OmpF de S.
typhimurium.
Ab’ s BSA DLso Kda LPS M.E. NIH
a m p C
rOmpF Pb PBS PCR P.G. PhoE PME SDS SDS-PAGE Y.P.
PRINCIPALES ABREVIATURAS
Anticuerpos
Albúmina Sérica Bovina
Dósis letal al 50%
Kilodaltons
Lipopolisacárido
Membrana externa
Cepa abierta de ratones
Proteína OmpC recombinante
Proteína probable OmpF recombinante
pares de bases
Amortiguador de fosfatos-salina
Reacción en Cadena de la Polimerasa
Péptido glicana
Proteína de membrana externa, inducible a concentraciones bajas de fosfatos
Proteínas de membrana externa
Dodecil sulfato de sodio
Electroforésis en geles de poliacrilan?ida con dodecil sulfato de sodio
INTRODUCCION
Las fiebres entéricas continúan causando considerables morbilidad y mortalidad
en naciones en las que existe una relación inversa entre el incremento de sus poblaciones
y urbanización, y la viglancia microbiológca del agua potable’. En estos países la
causa más frecuente y seria de fiebres entéricas es Salmonella typhi, agente etiológico
de la fiebre tifoidea2. El control epidemiológico de la enfermedad depende del
mejoramiento de la infraestructura sanitaria, del saneamiento básico y de la inspección
de portadores tifoídicos3, para lo cual es necesario no solo mejorar el nivel económico de
la población, sino también elevar el grado educacional en salud. Debido al tiempo y
dificultades que implica el logro de estos objetivos, y a pesar de sus imperfecciones
técnicas, las vacunas continúan siendo una buena alternativa para su prevención4.
Para la profilaxis de la fiebre tifoidea en nuestra país se cuenta con una vacuna
elaborada con bacterias muertas por calor, cuyas principales limitaciones son: que la
protección conferida e s a en función de la dosis infectante a la que el individuo esté
expuesto, que la inmunidad es de corta duración y que presenta efectos colaterales
adversos dados por la toxicidad del lipopolisacárido bacteriano5.
En los últimos &os, han sido estudiados diversos antígenos de la superficie de
Salmonella typhi como probables canddatos a vacunas contra la fiebre tifoidea. Entre
tanto en animales de experimentación como en personas con fiebre tifoidea, sin
embargo estos no correlacionan con un estado de inmunidad6.
Por otra parte Robbins y Robbins7 desarrollaron una vacuna a partir del antígeno
Vi de Salmonella, la cual, h e evaluada en estudios de campo en Nepal y Africa del
Surgv9, con resultados de protección parcial, muy probablemente por la naturaleza T-
independiente del antígeno polisacarídico, lo que hace suponer que no se genere
respuesta inmune celular ni se desarrollen efectivamente cklulas de memoria para
inmunizaciones o retos posteriores.
El estudio de las proteínas de membrana externa (PME’s) de las bacterias gram-
negativas como inmunógenos ha cobrado mucha importancia, ya que por su naturaleza
proteica y su localización habitualmente inducen anticuerpos de alta afinidad y
favorecen una respuesta celular, la que genera una inmunidad prolongada que es crucial
en la defensa contra patógenos intracelulares como S. typhi 10,11,12,13
En 1988 fue demostrada la importancia de las $ME de S. typhi. en la inducción
de inmunidad protectora al reto con la bacteria viva virulenta en un modelo murino de
fiebre tifoidea. La inmunización de ratones NIH con una dosis de 30pg de PME conf~rió
sobrevivencia al 100% de los ratones al reto hasta con 500 DL50 tanto de S. typhi 9,12 Vi:d aislada de un paciente con fiebre tifoidea, como de la cepa Ty2, que es empleada
para la producción de la vacuna tifoídica parenteral; mientras que la inmunización con
PME de S. typhimurium confirió protección del 60% y 30% respectivamente. Ademhs,
protector al reto con S. typhi, efecto que desapareció cuando el suero fue absorbido con
PME14. Mediante ensayos de inmunoelectrotransferencia demostraron que este suero
reacciona principalmente con un par de PME de P.M. entre 38 y 4 1 Kda,
correspondientes a las porinas15.
Por otro lado, la observación de que los pacientes con fiebre tifoidea y en fase de
convalecencia generan anticuerpos de clase IgG que reconocen principalmente porinas
de S. typhi, sugiere que también en la infección natural por S. typhi las porinas son blanco importante de la respuesta inmune e indica que la inmunidad por anticuerpos
contra porinas juega un papel esencial en el control de la infección, aunque, ello no
descarta la participación de otros mecanismos de la respuesta inmuneI6.
Estudios posteriores evaluaron en el modelo murino la participacidn de las
porinas de S. typhi 9,12 Vi:d en la inducción de una respuesta inmune protectora: la
inmunización de ratones NIH con la mezcla de 2 porinas extraídas de la membrana externa de la bacteria, y purificadas por métodos bioquímicos tradicionales, indujo
protección hasta del 80% al reto de 500 DL50 de S. typhi, en ratones que recibieron dos
inmunizaciones con 5pg de proteínaI7.
Finalmente, mediante ensayos in vitro se ha evidenciado que las porinas aisladas
tanto de S. typhi, S. typhimurium y E. coli son capaces de inducir una respuesta
proliferativa de linfocitos T provenientes de ratones, inmunizados con PME o con
porhas’*. Datos similares se han obtenido en linfocitost provenientes de individuos con
Todos estos resultados han permitido proponer a las porinas de S. typhi como
las proteínas candidatos en la elaboración de una vacuna contra la fiebre tifoidea.
Las porinas, corresponden al grupo de las proteínas matrices de membrana externa de las bacterias gram-nagativas y por sus niveles de expresión en la membrana
pertenecen al grupo de proteínas principales.
Sus
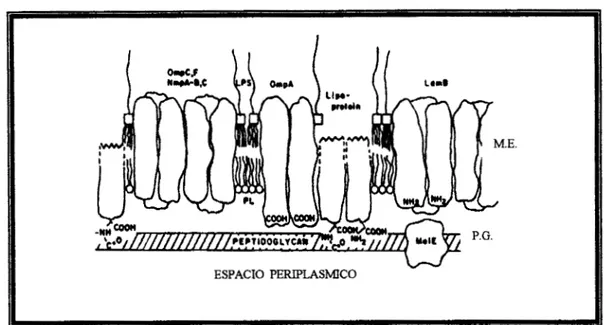
nombres fueron acuiiados por sucaracterística asociación a la peptidoglicana mediante enlaces iónicos (figura 1) y por su
capacidad de formar poros que participan en permeabilidad de moléculas hidrofilicas
de bajo peso molecular como aminoácidos, iones inorgánicos y sachridos con un límite
de exclusión de 600 Da2', con características particulare cada una. Algunos miembros de
la familia de porinas (lamB) funcionan como receptores para fagos2'. Estudios
bioquímicos y moleculares indican que hay una extensa homología entre las porinas de
bacterias gram-negativas, aunque de hecho existen secuencias de aminoácidos
Fig l. Organización de la envoltura de las bacterias gram-negativas”.
La membrana externa (M.E.), contiene lipopolisacárido (LPS) y a las porinas OmpC, OmpF, OmpA, LamB, que se asocian a la peptidoglicana (P.G.)
Los análisis por espectrofotometría en la región de infkarojo y por dicroismo
circular de las porinas de E. coli, muestran que las formas funcionales son trímeros
compuestos de subunidades monoméricas idénticas cuya estructura secundaria está
conformada por 16 hojas P-plegadas antiparalelas y largas asas con algunas a-hélices
intercaladas (figura 2). Cada monómero expone 8 asas (“loops”) cortas hacia el espacio periplásmico y 8 asas largas situadas hacia el exterior de la membrana externa. Estos
monómeros, forman un cilindro, al unirse mediante: un enlace iónico el extremo
El mayor número de modificaciones en la secuencia proteica entre una y otra
porina se localiza en las asas externas24 y por lo tanto son éstas las responsables de la
diferencia en el tamaño de la entrada de los poros. La estructura del trimer0 se forma
mediante el entrecruzamiento de las regiones hidrofóbicas de las vueltas p-15 y p-1,
donde los h e z residuos carboxilo de la región p-16, en particular el último (una Phe) y
la hoja pl, son esenciales para el correcto ensamblaje del. monómero de la porina. En el
centro del barril se forma un canal inclinado con respecto al eje longitudinal, que está
rodeado por dos cinturones de aminoácidos aromáticos que por su interacción con la
membrana le permite anclarse a la misma.
Las porinas de la membrana externa de E. coli han sido l a s más caracterizadas; a
la fecha se han clonado y secuenciado los genes de las 3 porinas principales: Om*, OmpF y PhoE, cuyos productos proteicos representan arriba del 2% de la masa total de
la bacteria. En Salmonella typhimuriumz5, además se expresa la porina OmpD.
I1
Fig. 2. Estructura del mondmero de la porina OmpF de E. coli 28
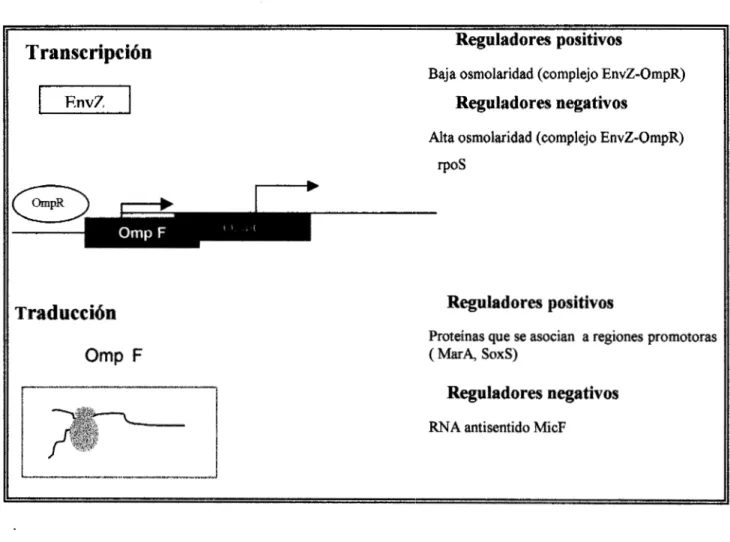
La regulación transcripcional y traduccional de :las porinas de bacterias Gram-
negativas es multifactorial (figura 3).
PhoE es inducida a bajas concentraciones de fosfato, sus poros son
particularmente eficientes en el transporte de fosfato inorgánico, compuestos
fosforilados y algunos otros solutos cargados negativamente. OmpF es catión selectivo,
y su expresión decrece en cultivos bacterianos en fase estacionaria de crecimiento,
debido a un gen que codifica para la proteína rpoS, la cual compite con la subunidad
sigma (o) de la RNA polimerasa e impide la transcripción de OmpF.
OmpF se expresa preferencialmente en medio de baja osmolaridad, mientras que
la expresión de OmpC se incrementa en medio de alta osmolaridad. Dicha regulación es
mediada por un sistema de dos componentes OmpR y Env Z, cuyos genes se encuentra
en el locus OmpB. OmpR es una proteína citoplasmática que se une a los promotores de
las porinas OmpC y OmpF, mientras que EnvZ es una proteína de la membrana interna,
de la que se propone que modula la h c i ó n de OmpK a través de fosforilaciones y
desfosforilaciones.
Otro factor bien definido en la regulación de porinas es el efecto de la
temperatura de crecimiento, en donde parhcipa una secuencia de RNA antisentido
denominada MicF, regulando negativamente a la porina OmpF a nivel post
trancripcional. El gen de MicF se localiza río arriba de la secuencia de Om@ y se
transcribe en dirección opuesta, codifica para dos distintos RNA, uno de 93 nucleótidos
que se une al ribosoma en el sitio de unión al RNAm de la OmpF y otra de 174
secuencias iniciales del
RNAm
de la OmpF. La trancripción del gen MicF es estimuladaa temperaturas de 37°C o mayores, y juega también un papel importante en la respuesta
a agentes tóxicos como el etanol, ácidos y antibióticos, así como el estrés oxidativo.
Además se han descrito proteínas que se unen al promotor de MicF capaces de regular
positivamente su expresión, como: MarA, SoxS, cuyos extremos carboxilo presentan
homología con la familia de reguladores transcripcionales XylS-AraC. El incremento
en la síntesis de MicF resulta en un decremento en la producción de 0 m ~ F ~ ~ . Por otro
lado, las propiedades del transporte fisiológxo mediado por porinas en S. typhi se
desconocen.
Reguladores positivos
Transcripcibn
Baja osmolaridad (complejo EnvZ-OmpR) Reguladores negativos
Alta osmolaridad (complejo EnvZ-OmpR)
vos
Omp F
Reguladores positivos
Proteínas que se asocian a regiones promotoras
( MarA SOXS)
Reguladores negativos RNA antisentido McF
PLANTEAMIENTO DEL PROBLEMA
1.
2.
3.
4.
5.
Como fue descrito en las propiedades inmunogénicas de las PME , se han
aislado de la membrana dos proteínas de 38-41
Kda.
que se expresansimultáneamente en cultivos bacterianos a 37OC y condiciones de osmolaridad
específicas. S610 una de ellas ha sido identificada y corresponde a la porina
0mpC2*. Considerando:
Que alsporinas son proteínas conservadas filogenéticamente entre las bacterias29.
Que presentan unas homologías hasta de un 90% a nivel de DNA entre ellas, y hasta
de un 70% a nivel de aminoácidos.
Que la porina OmpC caracterizada en: E. coli (38 Kda3') Salmonella typhimurium
(36 Kda31) y Salmonella @phi (36 Kda2*) presentan un peso semejante.
Que el polipéptido maduro de la porina OmpC de Salmonella typhi, presenta una
homología de un 79% a nivel de aminoticidos con su análoga en E. Coli3'
Que en corrimiento electroforético las porinas de Salmonella typhimurium
presentan un patrón electroforktico muy parecido al de las porinas de Salmonella
typhi.
OBJETIVO GENERAL
A. 1 Obtención y caracterización de la porina OmpF de Salmonella typhi.
OBJETIVO PARTICULARES.
B. 1 hslamiento y secuenciación del gen de la porina OmpF de Salmonella typhi.
B.2 Producción de la proteína recombinante.
DESARROLLO EXPERIMENTAL
2 2 5 7 1 2
A. CEPAS BACTERIANAS
A.l Salmonella typhi 9,12 Vi:d, aislada de un paciente con fiebre tifoidea y
conservada en la U.I.M. en Inmunoquímica del
C M N
desde 1985.A.2 E. coli DHSa (GIBCO B E ) , cepa derivada de la
DH51;
caracterizada poruna deleción en el operón lac y en un profago 080 que controla la síntesis del
fragmento omega de la P-galactosidasa, gyrA (Nal'), lo que le confiere
resistencia al ácid0 nalidíxico.
A.3 E. coli UH302 derivada de la cadena p400 mediante selección de resistencia a los fagos Tulb, Tula, TuII, y carente de las proteínas de membrana externa
Om@, OmpF y 0 m ~ A ~ ~ .
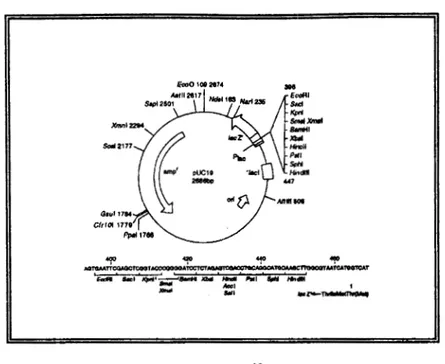
B. PLASMIDOS
B.l Plásmido de clonación pUC 19, que contiene el sitio de clonación múltiple en la
~~
Figura 4. pUC19= I 2686 pb
C. PREPARACION DE BACTERIAS COMPETENTES.
Una colonia de E. coli DH5a (o UH302) se cultivó durante toda la noche a 37°C
con agitación constante a 200 r.p.m. en 30 ml de medio LB (Luria Bertani, 1%, tiptona ,
0.5% de extracto de levadura y 1% de NaCl pH 7.5). Posteriormente se hzo una
dilución 1:200 del cultivo y se continuó hasta alcanzar una D.O. A 600 nm de 0.6,
después de la cual se le aiiadieron 600 m1 de medio LB y se continúo el cultivo por 15
min más. La bacteria se cosechó por centrifugación a 6000 r.p.m. durante 10 minutos. La pastilla se resuspendió en 0.01 M de Cae12 fiío y se incubó en hielo durante dos
horas. Al término la bacteria se cosechó por centrifugación, se resuspendió en 10 rnl
D. AMPLIFICACION DEL GEN DE LA PORINA OmpF
D.l DiseAo de los cebadores.
Los cebadores sintetizados para la amplificaciih del gen OmpF de S. typhi
corresponden a los extremos 5’ y 3’ de la secuencia publicada de la porina OmpF de
Salmonella typhimuriurn3’ y fueron denominados 2474 (posición 48-64 pb) y 2475 (posicibn 1285-1301 pb),
2474
5’ GCG AAT TCA CGG TAG CGA AAC G 3’ 2475
5’ TTC TAG AGG TGT GCT ATT AGA AGT GG 3’
D.2
Obtenci6n del DNA cromosomal de S. typhi 9J.2 Vi:dEL DNA cromosomal se obtuvo mediante Isotiocianato de G ~ a n i d i n a ~ ~
(TRIZOL GIBCO-BRL), para lo cual 1x1 O’ bacterias se resuspendieron en 1 m1 de trizol y se incubaron 5 minutos a temperatura ambiente. Posteriormente se le daderon
2 0 0 ~ 1 de cloroformo, se homogenizó e incubó 3 minutos a temperatura ambiente; al
término la solución se centrifugó a 6000 r.p.m. por 15 minutos a 4°C y se recuperó la
El DNA se recuperó centrifugando a 6000 r.p.m. durante 5 minutos a 4°C. y se lavó con una solución O. 1 M de Citrato de Sodio en etanol al 10%. Se dio un último
lavado con etanol al 75% y se secó por centrifugación al vacío (Speed vacuum)
[Hetovac Modelo VR-1 Heto Lab Equipment].
D.3 Reacci6n de Polimerizaci6n en cadena (PCR) para amplificaci6n del gen candidato a porina OmpF
El DNA genómico obtenido se utilizó como templado para amplificar por PCR
el gen de la pori^ OmpF de S. typhi, empleando el aparato Robocycler 40 ( Strategene)
y la enzima Pwo polimerasa (Lakeside).
Condiciones de la reacción.-
T 1 t l T 2 Num:Ciclos
VENTANA 1
45°C 1' 72°C 90°C 1' VENTANA
3
1'10'' 72°C
90°C l'lO''45"C VENTANA 2
45°C 2'
Concentraciones de los reactivos.-
COMPONENTE
~~~~~ ~ ~
Amortiguador de la enzima
Mezcla de dNTP's
Oligonucléotidos iniciadores
Polimerasa Pwo
Dimetil sulfóxido (DMSO)
~ ~~ ~~~~
CONCENTRACION FINAL
I
20 mM Tris-HC1, pH 8.4 con 50 mM de KC1,20 mM MgS04
100 pM de cada nucleótido
20.8 pM
2 u
3%
I
E. PURIFICACI~N DEL PRODUCTO DE PCR (CANDIDATO
AL
GENOmPF)
El producto obtenido de la PCR se sometió a corrimiento electroforético en gel
de agarosa al 1%. La banda de 1200 pb se cortó y se incubó con NaI a 56°C hasta licuar
completamente el gel. El DNA se adsorbió a perlas de vidrio por 30 min en agitación
constante y se recuperó mediante centrifugación, se lavó con una solución de (50 m1 de
etanol por cada 100 ml. que contenía 2OmM de Tris pH 7.4, 1mM EDTA y 0.1 M de NaC1). Finalmente el DNA se eluyó de als perlas a 56OC y se disolvió en agua
F. CLONACION DEL PRODUCTO DE PCR
El candidato a gen OmpF purificado (inserto) se ligii en el plásmido P U C ~ ~ ~ ’ , que
previamente se había digerido con la enzima Smal, en relación molar 3: 1 inserto:vector
mediante la catálisis con la enzima T4 DNA ligasa. El producto de la ligación se utilizó
para transformar bacterias competentes mediante un choque térmico a 42°C durante 1
min, después de lo cual se sembraron las bacterias en medio LB-Ampicilina (50 pg/ml)
y se incubaron 16 H a 37°C. Se analizaron las colonias resultantes (resistentes a
Ampicilina ) para identificar a aquéllas que portaban el inserto (OmpFP7.
G. DETECCION DEL GEN OmpF
G.l Aislamiento y transferencia de colonias. Las colonias probables positivas (seleccionadas mediante corrimiento
electroforético en gel de agarosa por presentar un peso molecular de 3.8 Kb, correspondiente al plásmido + el inserto ) se aislaron en placas de LB-agar y se transfirieron por contacto a papel de nitrocelulosa. Las bacterias se lisaron mediante el
contacto del papel con NaOH 0.5 M y NaCl 1.5 M por 2 min, seguido del tratamiento con (Tris-HC1 0.5 M a pH 8.0 y NaCl 1.5 M) y lavado con SSC (Citrato de sodio,
Cloruro de sodio) 2 veces por 45 segundos cada uno. Se secaron brevemente en papel
Whatman 3 MM y se entrecruzaron para fijar el DNA bacteriano a los papeles de
6.2 Hibridacidn con sondas radiactivas
Los papeles de nitrocelulosa (que contenían las, colonias) fueron previamente
bloqueados con leche al 5% en PBS y se hibridaron con una sonda de DNA de OmpF
marcada con citocina [a-32P] durante 24 hrs. a 68°C. El marcaje radiativo de la sonda
de DNA se realizó aiiadiendo dCTP32p y dGTP, dTTF’ y dATP no marcados en una
reacción de polimerización catalizada por la fiacción Klenow de la DNA polimerasa, la
sonda fue purificada por filtración en gel (Dupont’s Random Primers Extension kit)38
G. 3 Deteccih de las colonias portadoras del candidato OmpF
Los papeles de nitrocelulosa se lavaron a una astringencia de (0.1 Wa7) y la
sefial positiva de hibridación se reveló a través de la exposición a una placa para rayos
X.
H. CARACTERIZACION MOLECULAR DEL PROBABLE GEN OmpF
El DNA de los genes OmpF y el gen de OmpC , los cuales previamente se
habían sometido a digestión con diversas enzima
(Nu.
I, Cla I, NCO I, Nru I, Taq I yDde I) se sometieron a electroforésis en un gel de agarosa al 1%, durante 45 min. a 60 volts. El DNA se transfirió a papel de nitrocelulosa mediante capilaridad con Buffer
SSC 1OX durante 12 Hrs. El papel de nitrocelulosa previamente entrecruzado y
bloqueado con PBS-Leche 5% se hibridó con la sonda de OmpC marcada con citocina
I. SECUENCIACION DEL GEN
El plásmido pUC 19 que contenía el inserto candidato al gen OmpF se purificó
mediante lisis alcalina y se sometió a una amplificación por PCR en las siguientes
condiciones, empleando un secuenciador automático (Perkin-Elmer), y los cebadores
2474 y 2475.
T1 t l T2 t2
VENTANA1 96 30” 50 20’ 7
La secuenciación del producto de PCR se realizó por el método de Fan y cols 39 (Perkin
Elmer)
J. EXTRACCION DE PORINAS MEDIANTE EL METODO DE NIKAIDO
Las colonias que demostraron contener el plásmido con el probable gen OmpF se
expandieron en cultivo para de ellas extraer y caracterizar las proteínas (método de
Nikaido4’). Para ello se transformaron bacterias UH302:’2 lo que garantiza la expresión
de la proteína recombinante. Los cultivos bacterianos se crecieron en un medio mínimo de sales (Medio mínimo A) conteniendo 5% de extracto de levadura, sulfato de
magnesio al 25% y 12.5% de glucosa. Se incubaron a 317°C en agitación hasta alcanzar
sonicación y se centrifugó a 7000 r.p.m. por 15 min. a 4°C para remover las células
intactas y detritus. El sobrenadante se centrifugó a 40,000 r.p.m. por 45 min a 4°C para
obtener las membranas celulares. La cubierta de peptidoglicana se disoció mediante una
solución de Tris-HC1 lOmM pH 7.7, SDS al 2% incubándose 45 min a 32°C y
centrifugándose a 40,000 r.p.m. a 20°C. Las proteínas :insolubles se resuspendieron en
Buffer de Nikaido.(Tris 50 m M pH 7.7, SDS al 1%, NaC1 0.4 M, EDTA 5 m M ,
p-
mercaptoetanol al 0.05%) incubhndose 2 horas a 37°C. La suspensión se centrifugó a
40,000 r.p.m. durante 45 min a 20°C y las porinas se recuperaron del sobrenadante.
K. PURIFICACI~N DE LA PROTEÍNA RECOMBINANTE
Se llevó a cabo en una columna de exclusión molecular en gel de Sephacryl S -
200 (Pharmacia Chemical Co.), la cual se estabilizó previamente con 3 veces su
volumen total con Buffer de Nikaido pH 7.7. La muestra se inyectó en la columna con
una velocidad de flujo de 0.65 mVmin y se recolectaron fracciones de 3 m1 en un
colector autómatico (LKB Instruments) determinando su absorbancia a 280 nm.
L. CARACTERIZACION DEL PRODUCTO DEL GEN OmpF
L.l. Bioquimica (Electroforbis)
Las muestras de porinas se precipitaron con acetcma, se resuspendieron en buffer
verticales y sistema de amortiguador discontinuo, según la técnica descrita por
Laemmli4'. El gel introductor contenía 4% de acrilamidahis acrilamida y el gel
separador 12.5% acrilamidal bis acrilamida.
El corrimiento se llevó a cabo a 30 mA por placa. Concluido esto, los geles se
tifieron lhr con colorante azul de Coomassie R-250 y se decoloraron (Metano1 30% y
Ác. Acético 7%) hasta apreciar las bandas proteicas.
L. 2. Inmunoquímica (Western Blot)
La transferencia electroforética de las porinas contenidas en el gel de
poliacrilamida al papel de nitrocelulosa se realizó en una unidad de transferencia
( B L O W ) . L a s proteínas se transfirieron al papel a un voltaje constante de 75 V h . 5 h a
4OC, empleando el buffer T ~ w b i n ~ ~ . Los papeles se bloquearon con PBS-leche al 5%
durante toda la noche, se lavaron con PBS-tween 0.02% y se incubaron 1 hora a 25°C
con suero antiporinas hecho en conejo e 1 6 ' s purificadas del suero antiporinas.
L.3. TopoMgica (Citometría de Flujo)
Los cultivos bacterianos (UH302 ,SuZmoneZiu yphi , y Bacterias UH302 transformadas con el plásmido pST13 que contiene a la porina OmpC y con el plásmido pUC19 que
contiene a la porinas OmpF) se crecieron en medio LB (con o sin antibiótico) y se incubaron a 37°C en agitación hasta alcanzar su fase de crecimiento logarítmico (6 Hrs).
Cosechándose mediante centrifkgación a 3,000 r.p.m. y resuspendiéndose en PBS para
(540 m) a 1x108 por tubo eppendorf. Se cosecharon las bacterias mediante
centrifugación a 14,000 r.p.m por 5 minutos y la pastilla se resuspendió en 100 pl del
primer anticuerpo (suero antiporinas o IgG’s anti porinas) y se incubaron en agitación
por 45 minutos a T.A. Posteriormente se lavaron las bacterias y se incubaron 100 p1 del
segundo anticuerpo (anti-conejo conjugado a isotiocimato de fluoresceína FITC.)
durante 45 minutos en agitación a T.A., se lavaron con PBS y se les añadió yoduro de
propido (Y .P 4 mg/L). Después de agregar el Y .P. las muestras de inmediato fueron analizadas en el citómetro de flujo FACSort (BECTON-DICKINSON). Los anticuerpos
fueron diluidos en PBS con BSA al (O. 1 % W / V ) ~ ~ .
M. ANÁLISIS COMPAIUTNO DE LA ESTRUCTURA
PRIMARIA
Utilizando el programa Blast National center Biotechnology
(www.ncbi.nlm.nih.gov), se alinearon las secuencias de nucleótidos correspondientes a la regiones 5’ y 3’ del candidato a porina OmpF con la porina OmpF de Sulmonellu
RESULTADOS
1. OBTENCION DEL GEN CANDIDATO A PORZNA OmpF DE Styphi 9,12 Vi:d
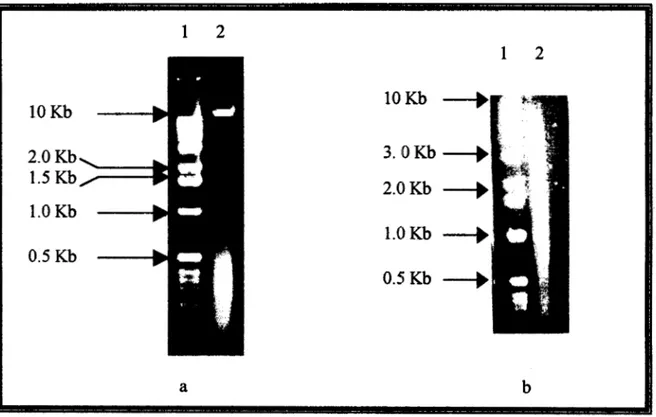
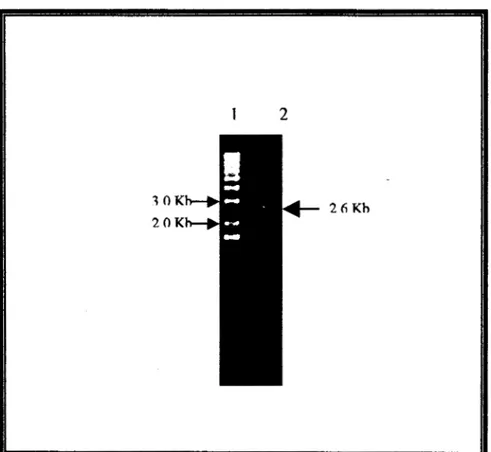
El DNA cromosomal de S. typhi 9,12 Vi:d que h e purificado por la técnica de
Isotiocianato de Guanidina mostró un patrón de alta calidad (integridad y pureza) (Fig
5a), se sometió a una digestión enzimática con Eco RV ( Fig 5b) y fue utilizado como
templado en la Reacción de Polimerización en Cadena
(PCR),
que utilizando a loscebadores 2474 (extremo 5’) y 2475 (extremo 3’), se llevó a cabo con el fin de amplificar un probable candidato a porina OmpF en S. typhi 9,12 Vi:d.
10 Kb
2.0 Kb 1.5 Kb
1.0 Kb
0.5 Kb
1 2
10 Kb
3.10 Kb
2.0 Kb
1.0 Kb
0.5 Kb
1 2
a b
Fig 5 DNA cromosomal de S. typhi 9,12 Vi:d integro (a) y digerido con Eco RV (b).
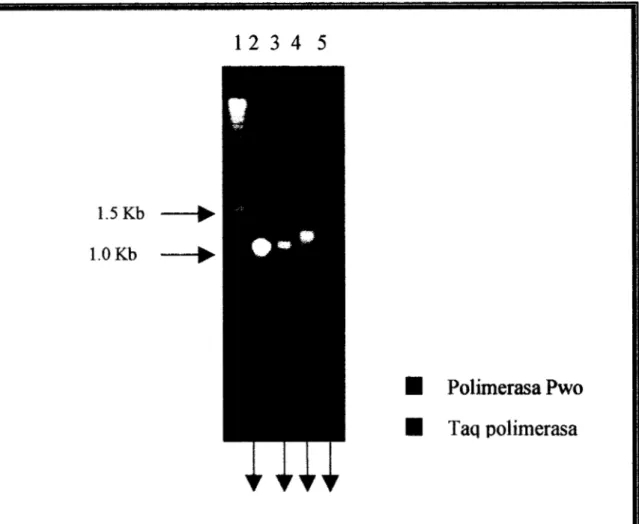
El producto de la reacción (PCR) corresponde a un segmento de DNA de aproximadamente 1,200 pb, y como se observa en la Fig 6, tiene
un
peso mayor peso algen OmpC (1,040 pb) de S. typhi
1.5 Kb 1 .O Kb
1 2 3 4
5
I
I
Polimerasa Pwo
Taq polimerasa
2. CLONACION DEL CANDIDATO OmpF
Una vez purificado el producto de PCR (candidato OmpF) fue clonado en el
plásmido pUC 19, previamente purificado y digerido (Fig 7)
3 O K h - 2 O K h -
1 2
2 h K h
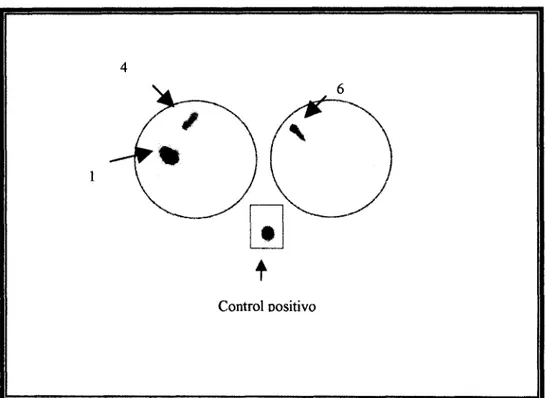
Una réplica de las colonias obtenidas tras la transformación de E. coli DH5a fue
hbridada con una sonda correspondiente al producto de PCR marcado con 32P. Las colonias positivas (Fig 8) se sometieron a expansión, extracción y digestión del DNA
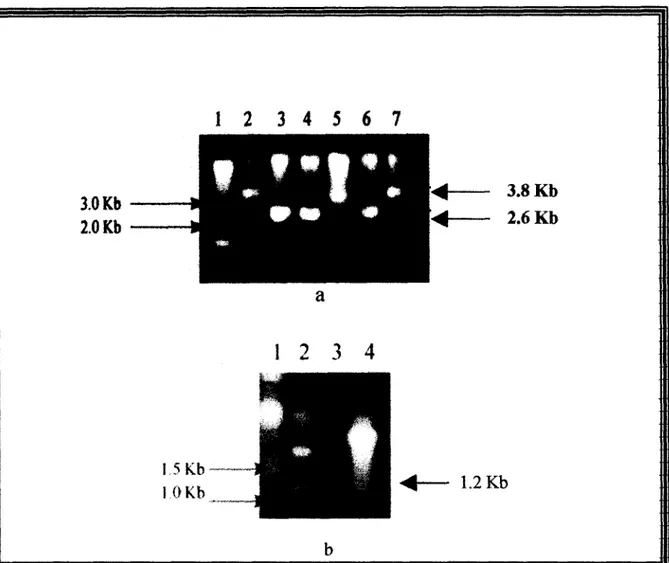
plasmídico, lo que permitió corroborar la inserción del gen candidato en el vector, tanto
por el aumento del P.M. (3.8 Kb) de la construcción (Fig 9a), como por la liberación de
un segmento de DNA de 1,200 pb, correspondiente al gen canddato a OmpF (Fig 9b).
4
t
Control oositivoFig 8. Hibridación de colonias acarreadoras del gen candidato
1
2
3
4
5
6
7
a
.+"
1.2KbFIG. 9. Clonaci6n del probable gen OmpF en pUC19.
(a) Corrimiento etecZroforEttico de M A ptasmidico dlgendo
con
Kpn 1 proveniente de las colonias 1,4,6 (carriles 2,5,7 respectivamente), y DNA del pldsmido pUC19 digerido con Kpn I, carriles 3,4, y 6.(b) Ljberacjbn del inserto de 3200 pb, mediante digestion con Sac I/ Hind 111, Carriles
3. IDENTIFICACION DEL GEN PROBABLE OmpF
La secuencia de los extremos terminales 5’ y 3’ del gen clonado se
determinó automáticamente a través del espectro de fluorescencia emitidas por los
nucleótidos marcados e incorporados en una reacción de polimerización del gen
probable OmpF, con la enzima Amplitaq (Perkin-Elmer), lográndose secuenciar 450 pb
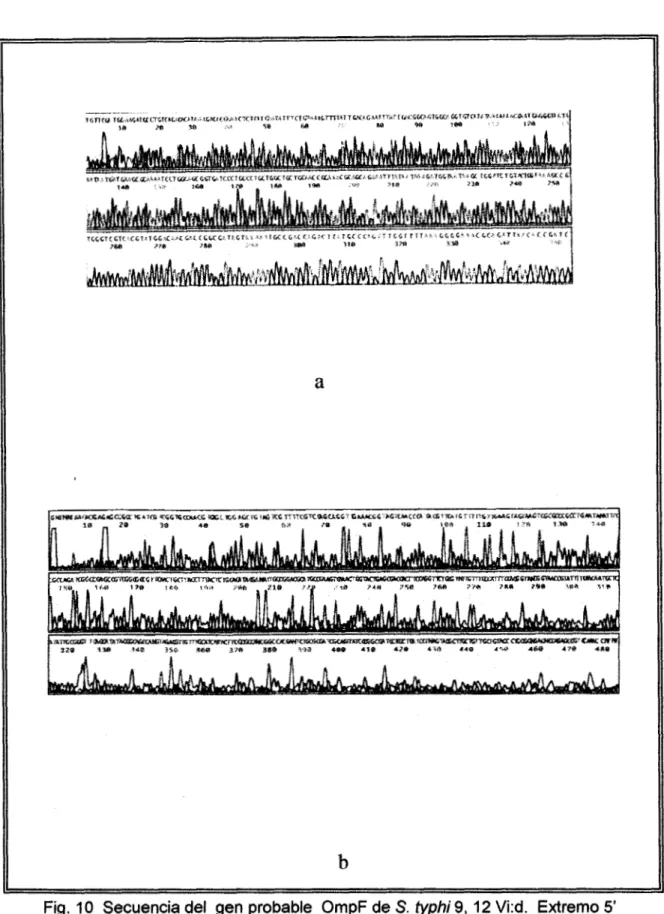
de cada extremo. Del extremo 5’ terminal 430pb de las 450 pb secuenciadas (Fig loa) mostraron una homología de secuencia del 97% con el correspondiente extremo en el
gen OmpF de S. typhimurium . y sólo 45 pb presentaron homología con OmpC,
mientras que 439 pb del extremo 3’ terminal (Fig lob) mostraron una homología del
99% con el 3’ del gen análogo y sólo 114 pb mostraron homología signrficativa con
Om$. Las homologías se determinaron con el programa (Blast. National Center
Biotecnology).
a
b
El patrón de restricción enzimática (utilizando las enzimas Taq 1, Alu I, Dde I,
Nru I, Nco I y Cla I) permitió establecer diferencias entre el candidato OmpF y OmpC,
como se observa en la (Fig 1 la), la porina OmpC sólo fue digerida con la enzima Cla I,
mientrás que el candidato OmpF fue digerido con las emimas: Taq I, Nru I, Nco I y Cla
I Sin embargo la similitud entre ambos genes, se puso de manifiesto por la hibridación
I SKk"-+
1 OKh"---,
05Kh--"+
OmpF 0mPc
a
1 2 3 4 5 6
Carriles: 1, 8 P.M. 2,9 Taq 1 3.10 Alu I 4,11 Dde I 5,12 Nru I 6,13 Nco I 7, 14 Cla I
DNA OmpC
Sonda
OmDC bORDEN 1 A l u I 2 Cia 1 3NcoI 4 Nru I
5 Taq 1 6 Dde I
DE LOS CARRILES
Fig 11 Patrdn de restricci6n del probable OmpF y OmpC de S. typhi 1 la: 2-7
4. ANALISIS DEL PRODUCTO DEL GEN OmpF
Las clonas acarreadoras del probable gen OmpF se utilizaron para transformar
E.coZi UH302 (deficiente de las porinas OmpA, OmpC y OmpF), y los cultivos masivos de estas bacterias transformadas para obtener porinas de las membranas mediante la
técnica Nikaido4'. Las proteínas se purificaron por cromatografia de exclusión
molecular en una resina de Sephacril S-200 (Fig 12). Las fiacciones con mayor D.O. a
280 nm fueron colectadas y sometidas a electroforésis vertical en condiciones reductoras
(Fig 13), observándose que el producto del probable gen OmpF clonado, corresponde a
una proteína de un PM similar al gupo de porinas, ligeramente más alto que el de
OmpC de S @phi (Fig 13b, carriles 3 y 2, respectivamente). Cuando se hace una mezcla
de OmpC recombinante (rOmpC) y la probable OmpF recombinante (rOmpF), el
comportamiento electroforético corresponde al de las prinas expresadas naturalmente
en S. typhi (carriles 5 y 2 de 13a, respectivamente), lo que sugiere fuertemente que la identidad del producto del gen clonado es OmpF.
0.1
o .o
Purificacion de la porina OmpF
columna sephacryl
S-200
dona 1 -UH302
1 2 3 4 S
66 Kda
45 Kda
34 Kda
24 Kda
18 Kdn
__+
-b
-b
-b
-b
a
b
La proteína recombinante OmpF purificada de E coli UH302 y desnaturalizada
fue reconocida en un ensayo tipo Western por las IgG's de un conejo inmunizado con la
mezcla de porinas de S. typlzi expresadas naturalmente, las cuales reconocen con la
misma intensidad a ambas proteínas recombinantes (Fig 14a, 14b).
Por otro lado, mediante Inmunofluorescencia Indirecta utilizando los anticuerpos
policlonales antiporinas, se demostró que la proteína probable OmpF se expresa en la
superficie de la E. coli UH302 transformadas con el plásmido que acarreaba el gen, lo que demuestra que su exporte y plegamiento es análogo al que sufren las porinas nativas
en S. typhi (Fig 15). El nivel de expresión es similar al de OmpC, como se pudo
observar por la intensidad de fluorescencia que se obtuvo con la m 3 0 2 transformada
a
1 2 3 4 5 6
5 ESTRUCTURA PRIMARIA DE LA PROBABLE
OmpFDE
S.typhi
El análisis comparativo de la probable OmpF de S. @phi
con
otras
porinas
descritas,
se
hizo
utilizando
el programa Blast National Center Biotechnology(www.ncbi.nlm.nih.nov), alineando l a s secuencia de nucleótidos de las regiones 5’ y 3’.
Se
demostró una identidad en 5’ y 3’ del 97% y 99%, respectivamente, con lasecuencia OmpF de Salmonella typhim~rium’~,de acuerdo al modelo estructural de la porina
OmpF de E. coli26
.
La región 5’, aquí alineada codifica parae1 codón de inicio, el péptido lider, la hojap
plegada(p)
16, el asa corta (T) T1, lap
1, el asa largas (L) L1, la p2, el T2, lap3, el L2 y la p4. La 3’ alineada codifica para la hoja
p
9, el ct hélice, L5, pl0, T6,pll,L6, p12, T7, p13, L7, p14, T8, pl5, L8 y p16. En la tabla 1 se muestra la identidad
entre la estructura primaria de la probable prim OmpF de S. typhi y la porina OmpF de S. typhimurium.
En la tabla 2 se muestra la identidad entre la probable porina OmpF y OmpC de S.
typhi, en las que la homologia sustancialmente se encuentra en 3 regiones: la primera que
comprende p2, T2 y p3 del extremo 5’, la segunda situada en el extremo 3’ y que abarca del
L6 a
p
13, y la tercera, adyacente a la región 2 en 3’, comprende delp
14 al L8; de acuerdoOmpF de S. typhimurim 1 2
Probable OmpF de S. &phi
Secuencia Homología Regiones Sustituciones
"KRKILAAVIPALLAAATAAAIE C.lnicio, Peptido Lider, p 16
IYNKDGNK~~LDLYGKAVGR~@HVWTTT
LVRLAF~OAGL
T I, p I, L I, p 2, T 2, p 3 1 GDSK30NADQTYAQlG40FKGETQ1NTD50 L 2, p 4
LTGFGQWEYR60TKADRAEGEQ70QNSN
V G A K Y D A N N V Y L A A V Y A E T S f3 11, L 6 , p 12, T 7 , 813,
16
, . . , . . ,
2 1 V 2 % N T W T W 3 " T Q N L E W A Q L 7,p 14, T 8, p 15, L 8, p YQFFDFGLRPAISWQSK280GQLNGAG287
GSADLAKYIQAGAT-
WVDyRFNLLD'2%NDYSSSYV329
4301440 (97%)
4391443 (99%) G -K)267
I
Tabla 1. Homología entre la porina OmpF de Salmonella typhimurium y el candidato a
OmpC de S. typhi 1 2 3
5’ 3’
Probable OmpF de S. typhi
Secuencia
P 2
KGETQP
l d e n t l
Regibn
1
ND~Q~LTGPGQ
T 2 43/53 (8 1 %)WEY p 3 Y -+F=
52/60 (88%) F- Lm
DISCUSION
Por su localización y naturaleza, las PME han sido consideradas como
antígenos importantes en la inducción de una respuesta inmune protectora contra
infecciones por bacterias gram-negativas . Desde 1988, fue demostrada la
importancia de las mismas en un modelo murino de fiebre tifoidea, donde las porinas de
,SuZmoneZZu typhi son el principal blanco de la respuesta inmune h ~ m o r a l ' ~ y ~elular'~, lo que ha permitido proponerlas como proteínas candidatas vacunales contra la fiebre
tifoidea.
22 y 45
10,11,12 y 13
Las porinas que codleren protección son una mezcla de dos proteínas, de las
que una de ellas ha sido caracterizada como la porina OmpC y la identidad de la segunda
permanecía sin establecer, sin embargo por las semejanzas bioquímicas existentes entre
las porinas, se podía tratar de OmpF, lo que nos llevó a diseiiar cebadores
correspondientes a los extremos 5' y 3' del gen probable OmpF de SulmoneZZu lyphimurium", para poder clonar al correspondlente gen de S. Yphi: que hasta la fecha
no había sido identificado.
El gen amplificado de Styphi presentó un peso aproximado de 1,200 pb, el cual es ligeramente mayor que el gen OmpC de S.typhi de aproximadamente 1,040 pb, y
presenta un patrón de digestión enzimática dlferente a OmpC
.
El gen OmpF de S. typhifue clonado en el plásmido P U C ~ ~ ~ ~ , y una vez caracterizadas las construcciones se
procedió a transformar E. coZi UH30232 (deficientes de porinas OmpC, OmpF y OmpA) con la finalidad de expresarla como proteína recombinante. La purificación de
la proteína recombinante por el método de Nikaido4', permite establecer que la OmpF
enlaces iónicos a la peptidoglicana. La purificación por cromatografia de exclusión
molecular en Sephacryl S-200 permite establecer que se asocia en forma de
homotrimeros, ya que eluye en la fracción en la cual se obtienen las porinas de S. typhi,
es decir con un peso molecular mayor de 1 10 Kda.
La electroforésis en poliacrilamida-SDS determin6 que se trata de la porina con
mayor peso molecular presente en la mezcla nativa, y su similitud antigénica quedó de
manifiesto por los ensayos de Westem-blot, en los que se observó la reactividad con anticuerpos policlonales dirigidos contra las porinas nativas de S. lyphi. La adecuada
expresión de la proteína sobre la superficie de la bacteria se corroboró con los ensayos
de Citometría de Flujo, donde los anticuerpos policlonales contra las porinas nativas
identificaron una población de m 3 0 2 que expresa la proteína en la superficie de manera
semejante a la porina OmpC.
La secuenciación parcial del gen y el análisis de la homología con la porina
OmpF de S typhimurium y OmpC de S typhi permite corroborar que el gen amplificado
del genoma de S. lyphi y expresado como proteína recombinante se identifica como el
BIBLIOGRAFIA
1. Christie, A.B. 1987. Typhoid and Paratyphoid fever in: “Infection diseases”, A.B.
Christie 4* de. De. Churchill Livingstone. New York p. 100.
2. Jones B.D. 1996. Salmonellosis: Host Immune Responses and Bacterial Virullence
Determinants. Annu. Rev. Immunol. 14:533-61.
3. Nastasi A., Mammina C., Villafrate N.R. 1993. Epidemiology of Salmonella
typhimurium Ribosomal DNA. Analysis of strains fiom Human and Animal source
Epidemiol. Tnfect. 1 10:553-565.
4. Pegues David A. and S.I. Miller. 1994. Salmonellosis, includmg typhoid fever.
Current Opinion in Infectious Diseases. 7:6 16-623.
5. Joó I. 1970. International conference on the application of vaccines against viral,
rickettsial and bacterial desease of man. Washngton D.C. p 339-341. PAHO/WHO
Scientific publication 226. Pan American Health Organization. Washington D.C:
6. Hornick, R.B., S.E. Greisman, T.E. Woodward H.L. Dupont, A.T. Dawkins, and
M.J. Snyder. 1970. Typhoid fever., pathogenesis and immunological control. Parts 1
and I1 N. Engl. J. Med. 283:686-691. 739-746.
7. Robbins, J,D., Robbins, J.B. 1984. Re-examination of the inmunopathogenic role of
the capsular polysaccharide (vi antigen) of Salmonellcr typhi. J. Infect. Dis. 150:436-
8. Acharya IL. Love, C.L., Thapa,R., et al. 1987. Prevention of typhoid fever in Nepal
with the Vi capsular polysaccharide of Salmonella typhi. N. Eng. J. Med. 3 17: 1 1 O 1
-
1 104.
9. Klugman, K.P., Koornhof. H. J., Gilbertson, Y.T, et al. 1987. Protective activity of
Vi capsular polysacharide vaccine against typhoid fever Lacet. 2: 1 165-1 169.
10. Wang, L. Y. y Frash,
C.
E. 1984. Development of a .Neisseriu meningiticlis group Bserotype 2b protein vaccine and evaluation in a mouse model. Infect. Tmmun. 46:
408-414.
11. Buchanan, T. M. Pearce, G. K. y Arko, R. J. 1977. Protection against infections with
Neisseria gonorrhoeae by immunization with outer membrane proteins complex and
purified pili. J Infect Dis. 136: 1342-1347.
12. Guling, P. A. , McCracken, G. H. , Frich, C. F. , Johnston, K. H. y Hansen, E. J.
1982. Antibody response of infants to cell surface exposed outer membrane proteins
of Haemophilus disease. Infect. Tmmun. 37: 82-88.
13. Kussi, N. J. ; Nurmien, M. ; Saxén, H. y Miikela,
F'.
H. 1981. Immunization withmajor membrane protein F (porin). Preparation in experimental murine
Salmonellosis; effect of lipopolysaccharide. Infect. Immun. 34: 328-332.
14. Isibasi, A. , Ortíz, V., Vargas, M., Paniagua, J., Gonzhlez, C., Moreno, J., y Kumate,
J. 1988. Protection against Salmonella typhi infection in mice after immunization
with outer membrane proteins isolated from Sulmonellu typhi 9, 12 d, Vi. Infect.
Tmmun. 56: 2953-2959.
15. Isibasi, A., Paniagua, J., Rojo, M. P., Martin, N., Rarmírez, G., González, C., López-
Macias, C., Kumate, J. & Ortíz, V. 1994. Role of pori.ns from Salmonella typhi in the
16. Ortíz, V., A. Isibasi, E. Garcia-Ortigoza and Jesus Kumate. 1989. Immunoblot
Detection of class-specific Humoral Immune Response to Outer Membrane Proteins
isolated from Salmonella typhi in Human with typhoid fever. J. Clin. Microbiol. 37:
1640-1645.
17. Isibasi, A., Ortíz-Navarrete, V., Paniagua, J., Pelayo, Rosana., G o d l e z , C. R.,
Garcia, J. A., & J. Kumate. 1992. Active protection of mice against Sulmonellu vphi
by immunization with strain-specific porins. Vaccine. 1 O: 81 1-81 3.
18. González C., Isibasi A., Ortíz-Navarrete, V., Paniagua J., Garcia J.A., Blanco F &
Kumate J. 1993. Lymphocitic proliferative response to outer membrane proteins
isolated from Salmonella typhi. Microbiology & Immunology 37:793-799.
19. Blanco, F., A. Isibasi, C.R. González, V. Ortíz, J. Paniagua, C. Arreguín and J.
Kumate. 1993. Human cell mediated lmmunity to prins from Salmonella typhi.
Scand J. Infect. Dis. 25: 73-80
20. Jap B.K. & Waiian P.J. 1996. Structure and Functional Mechanism of Porins.
Physiol. Review. 76 (4): 1073-1088.
21. Nikaido, H. & Vaara, M. 1985. Molecular Basic of bacterial outer membrane
permeability. Microbial. Rev. 49: 1-32.
22. Osborn, M. J. y Wu, H. C. 1980. Proteins of the outer membrane of g a m negative bacteria. Annu. Rev. Microbiol. 34: 369-422.
23. Schulz, G. E. 1993. Bacterial Porins. Structure and function. Current Opinion in
Structure Biology. 5: 701 -707.
24. Nikaido H. 1994. Porins and Specific Difussion Channels in Bacterial Outer
25. Nikaido,H. and Vaara M. 1987. Outer Membrane. En “Escherichiu coli and
Sirln~onellu typhimurium: Celular and Molecular Biology” Iingraham,J.L., Low,
K.B., Magassanik, B., Schrechter, M. y Umbarger., H.E. Eds). Vol 1. American
Society for Microbiology. p.p. 7-22.
26. Cowan, S . W., T. Schnnger., G. Rummel., M. Steiert.:, R. Ghosh., R.A. Pauptit., J.N.
Jansonius & J. P. Rosenbusch. 1992. Crystal Structures explain functional properties
of two E. coli porins. Nature. 358: 727-733.
27. Pratt L.A., W. Hsing, K. E. Gibson and T. J. Silhavy. 1996. From acids to
osmZ:multiple factors influences synthesis of the OmpF and OmpF porins in
Escherichia coli.
28. Agüero J., G. M0ra.M.J. Mroczenslu-Wildey, M.E. Fernández-Beros. L. Aron and F.C. Cabello. 1987. Cloning, expression and characterization of the 36 Kdal.
Sulmonellu typhi porin gene in Escherichiu coli. Microbial. Pathogenesis. 3 :399-407.
29. Mizuno T., M.Y Chou and M. Tnouye. 1983. Acompartive Study on the Genes for
Three Porins of the Escherichia coli Outer Membrane. J.Bio1. Chem. 258 (1 1): 6932-
6940.
30. Puente J.L. et al. 1995. The Salmonella OmpC gene: structure and use as a cairier for
heterologous sequences. Gene 156 1-9.
31. Meyer P. et al. 1998. Virulence of a Salmonella typlumurium OmpD Mutant.
Infection and Immunity. 66: 1 387-390
32. Cole S.T., Sonntag T. Henning U. 1982. Cloning nad ’Expression in Escherichia coZi
K-12 of the genes for major outer membrane protein OmpA form shigella
dysenteriae, Enterobacter aerogenes and Serratia marcescens. J. Bacteriol. 149: 145-
33. Yanisch-Perron, C., Viera J., and Messing J. 1985. Gene 33 103
34. J. Sambrook., E.F. Fritsch, T. Maniatis. 1989. Molecular Cloning a laboratory
Manual. Second ed. Cold Spring HarborLaboratory Press.
35. Venegas A., Gomez, I., Bruce E. and Martinez, M. PCR amplification and cloning of
the Salmonella typhimurium OmpF porin gene. DNA. sequencing and expression in
Escherichiu coli. Unpublished. Acces. 2 3 1594
36. Chomczynski P., and Sacchi N. 1987. Single step method of RNA isolation by acid guanidium tyocianate phenol chloroform extraction. Annal. Biochem 162: 156-159. 37. Weiss, B., Jacquemin-Sablon, A., Live, T.R., Fareed, G.C., and Richardson, C.C.
1968. J. Biol. Chem. 2434543
38. Feinberg, A.P. and Volgelstein, B. 1983. A technique for radiolabeling DNA restriction endonucleases fiagments to hrgh specific activity. Anal Biochem, 132,6-
13.
39. Fan J., R.S. Ranu, C. Smith., C. Ruan, C.W. Fuller. 1996. DNA Sequencing with
[alpha p”]-labeled ddN’TP terminators: a new approach to DNA Sequencing with
Thermo-Sequenace DNA polimerase. Biotechniques 2 1 (6): 1 132-7.
40. Nikaido H. 1983. Proteins Forming Large Channels form Bacterial and mitochondrial
Outer membranes: Porins and Phage Lambda Receptor Proteins. Methods in
En~~m010gy 97: 85-101
41. Laemmli, U.K. 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227:680-685.
42. Towbin, H., Staehelin,
T.,
Gordon, J. 1979. Electrophoretic transfer of proteins form polyacrilamide gels to nitrocellulose sheets: procedures and some applications. Proc.43. Van der Waaij L.A. G. Mesander, P.C. Limburg, and 13. van der Waaij. 1994. Direct
Flow Cytometry of Anaerobic Bacteria in Human Feces. Cytometry 16: 270-279 44. Gottlieb, M. and Chavko, M. 1987. Analytical Biochemistry,l65:33-37
45. Lugtenberg, B. and Van Alpen, L. 1983. Molecular nrchtecture and functioning of the membrane of Escherichia coli and other gram-negative bacterias. Biochem.
Los integrantes del jurado designados por la división de Ciencias Biológicas y de la
Salud, de la Universidad Autónoma Metropolitana, Unidad Iztapalapa aprobaron la
presente tésis el día 22 de Noviembre de 1999.