CARTA DE AUTORIZACIÓN DE LOS AUTORES PARA LA CONSULTA, LA REPRODUCCIÓN PARCIAL O TOTAL, Y PUBLICACIÓN ELECTRÓNICA DEL TEXTO
COMPLETO.
Bogotá, D.C., 3 de Julio de 2009
Tesis doctoral
Trabajo de Grado
Señores
BIBLIOTECA GENERAL
Cuidad
Estimados Señores:
La suscrita
Ángela Maritza Cardoso Sandoval, con C.C. No. 1018411853, autor(es) de la tesis doctoral y/o
trabajo de grado titulado
RESISTENCIA DE
HELICOBACTER PYLORI
AL
METRONIDAZOL RELACIONADA CON LOS GENES
fdxA, fdxB, fldA, oorD, PorD, rdxA
y
frxA
presentado y aprobado en el año 2009 como requisito para optar al título de Bacteriólogo;
autorizo (amos) a la Biblioteca General de la Universidad Javeriana para que con fines
académicos, muestre al mundo la producción intelectual de la Universidad Javeriana, a través de
la visibilidad de su contenido de la siguiente manera:
Los usuarios puedan consultar el contenido de este trabajo de grado en Biblos, en los sitios
web que administra la Universidad, en Bases de Datos, en otros Catálogos y en otros sitios
web, Redes y Sistemas de Información nacionales e internacionales “Open Access”
y en las
redes de información del país y del exterior, con las cuales tenga convenio la Universidad
Javeriana.
Permita la consulta, la reproducción, a los usuarios interesados en el contenido de este
trabajo, para todos los usos que tengan finalidad académica, ya sea en formato CD-ROM o
digital desde Internet, Intranet, etc., y en general para cualquier formato conocido o por
conocer.
Continúo conservando los correspondientes derechos sin modificación o restricción alguna;
puesto que de acuerdo con la legislación colombiana aplicable, el presente es un acuerdo
jurídico que en ningún caso conlleva la enajenación del derecho de autor y sus conexos.
De conformidad con lo establecido en el artículo 30 de la Ley 23 de 1982 y el artículo 11 de la
Decisión Andina 351 de 1993,
“Los derechos morales sobre el trabajo son propiedad de los
autores”
, los cuales son irrenunciables, imprescriptibles, inembargables e inalienables.
Atentamente,
________________________________________
Ángela Maritza Cardoso Sandoval
C.C. 1018411853 Bogotá
NOTA IMPORTANTE: El autor y o autores certifican que conocen las derivadas jurídicas que
se generan en aplicación de los principios del derecho de autor.
FORMULARIO DE LA DESCRIPCIÓN DE LA TESIS DOCTORAL O DEL
TRABAJO DE GRADO
TÍTULO COMPLETO DE LA TESIS DOCTORAL O TRABAJO DE GRADO:
RESISTENCIA DE
HELICOBACTER PYLORI
AL METRONIDAZOL RELACIONADA
CON LOS GENES
fdxA, fdxB, fldA, oorD, PorD, rdxA
y
frxA
AUTOR O AUTORES
Apellidos Completos
Nombres Completos
Cardoso Sandoval
Ángela Maritza
DIRECTOR (ES) TESIS DOCTORAL O DEL TRABAJO DE GRADO
Apellidos Completos
Nombres Completos
Trespalacios Rangel
Alba Alicia
TRABAJO PARA OPTAR AL TÍTULO DE: Bacteriólogo
FACULTAD
: Ciencias
PROGRAMA:
Carrera _X_ Licenciatura ___ Especialización ____ Maestría ____ Doctorado
____
NOMBRE DEL PROGRAMA:
BACTERIOLOGIA
NOMBRES Y APELLIDOS DEL DIRECTOR DEL PROGRAMA:
Luz Amparo
Maldonado
CIUDAD:
BOGOTA AÑO DE PRESENTACIÓN DEL TRABAJO DE GRADO: 2009
NÚMERO DE PÁGINAS
77
TIPO DE ILUSTRACIONES:
-
Ilustraciones
-
Tablas, gráficos y diagramas
SOFTWARE requerido y/o especializado para la lectura del documento Adobe Reader, o programas que permitan leer documentos en .pdf
MATERIAL ANEXO (Vídeo, audio, multimedia o producción electrónica):
Duración del audiovisual: ___________ minutos.
Número de casetes de vídeo: ______ Formato: VHS ___ Beta Max ___ ¾ ___ Beta Cam ____ Mini DV
Otro. Cual? _____
Sistema: Americano NTSC ______ Europeo PAL _____ SECAM ______
Número de casetes de audio: ________________
Número de archivos dentro del CD (En caso de incluirse un CD-ROM diferente al trabajo de grado): _________________________________________________________________________
PREMIO O DISTINCIÓN(En caso de ser LAUREADAS o tener una mención especial):
_______________________________________________________________________________
DESCRIPTORES O PALABRAS CLAVES EN ESPAÑOL E INGLÉS:
Son los términos
que definen los temas que identifican el contenido. (
En caso de duda para designar estos descriptores, se
recomienda consultar con la Unidad de Procesos Técnicos de la Biblioteca General en el correo
[email protected]
, donde se les orientará).
ESPAÑOL
INGLÉS
Helicobacter pylori y resistencia antibiótica Helicobacter pylori and antibiotic resistance fdxA, fdxB, rdxA, frxA, fldA, oorD, PorD fdxA, fdxB, rdxA, frxA, fldA, oorD, PorD Resistencia al Metronidazol Metronidazole resistance
Nitroreductasas y oxidasas de Helicobacter Nitroreductases and oxidases Helicobacter
Activación y acción de Metronidazol Reductive Activation and action of metronidazole Biologia molecular, genes y Metronidazol Biology molecular, genes and metronidazole
RESUMEN DEL CONTENIDO EN ESPAÑOL E INGLÉS:
(Máximo 250 palabras -
1530 caracteres):
RESUMEN
Este trabajo de grado tiene como objetivo principal, realizar una revisión bibliográfica para
conocer la resistencia de Helicobacter pylori al metronidazol asociada a los genes rdxA, frxA,
fldA, fdxA, fdxB, oorD y PorD. Se realizó una búsqueda de literatura en las bases de datos de
EBSCO, Pubmed, Science direct, CDSR, Elsiver y Scielo de estudios de la resistencia
antimicrobiana de H. pylori al metronidazol, la actividad, diagnóstico y asociación de las
alteraciones de los genes que codifican las nitrorreductasas en dicha resistencia.
De los 64 estudios obtenidos en la primera búsqueda, se seleccionaron 31 artículos, identificando
que solamente 9 cumplían con los criterios de inclusión. Se observó que el 44.31% (117/264) de
los aislamientos son resistentes al metronidazol. De las 117 cepas resistentes, el 18.80% (22/117)
presentó un nivel de resistencia alto al metronidazol, el 76.92% (90/117) un nivel intermedio o
moderado de resistencia y el 0.85% (1/117) restante un nivel de resistencia bajo. Se encontró 326
mutaciones en las cepas resistentes, el 74.23% (242/326) correspondieron a alteraciones en el
gen rdxA, el 25.76% (84/326) en el gen frxA.
resistencia, la causa de la resistencia no implica la alteración del gen, encontrando en los estudios
mutaciones en cepas sensibles al metronidazol, lo cual implica que incluso en ausencia de un
rdxA funcional, una cepa de Helicobacter pylori puede ser sensible al metronidazol.
ABSTRACT
The main purpose of this graduation work is to do a bibliographic revision in order to know the
resistance of Helicobacter pylori at metronidazole associated to genes rdxA, frxA, fldA, fdxA,
fdxB, oorD and PorD. A search of literature from the data base of EBSCO was done, Pubmed,
Science direct, CDSR, Elsiver and Scielo of studies of the anti-microbial resistance of H. pylori at
metronidazole, the activity, diagnosis and association of the genes alterations which codify the
nitrorreductasas in such a resistance.
Out of the 64 studies obtained during the first search, 31 articles were selected, detecting that
only 9 of them met the inclusion criteria. It was observed that 44.31% (117/264) of the isolated
cases are resistant to metronidazole. Out of the 117 metronidazole resistance isolates, the
18.80% (22/117) presented a high level metronidazole resistance, the 76.92% (90/117) a
moderate level resistance and the remaining 0.85% (1/117) a low level metronidazole resistance.
326 mutations occurred in the resistant roots (bacterium), 74.23% (242/326) corresponded to
alterations in gene rdxA and 25.76% (84/326) in gene frxA.
RESISTENCIA DE HELICOBACTER PYLORI AL METRONIDAZOL RELACIONADA CON
LOS GENES fdxA, fdxB, fldA, oorD, PorD, rdxA y frxA
ÁNGELA MARITZA CARDOSO SANDOVAL
TRABAJO DE GRADO
Presentado como requisito parcial para optar al titulo de
BACTERIOLOGO
PONTIFICIA UNIVERSIDAD JAVERIANA
FACULTAD DE CIENCIAS
CARRERA DE BACTERIOLOGIA
BOGOTÁ D.C.
2
RESISTENCIA DE HELICOBACTER PYLORI AL METRONIDAZOL RELACIONADA CON
LOS GENES fdxA, fdxB, fldA, oorD, PorD, rdxA y frxA
ÁNGELA MARITZA CARDOSO SANDOVAL
DIRECTORA: ALBA ALICIA TRESPALACIOS RANGEL, MSc.
PONTIFICIA UNIVERSIDAD JAVERIANA
FACULTAD DE CIENCIAS
CARRERA DE BACTERIOLOGIA
BOGOTÁ D.C.
3
NOTA DE ADVERTENCIA
Artículo 23 de la Resolución N° 13 de Julio de 1946
4
RESISTENCIA DE HELICOBACTER PYLORI AL METRONIDAZOL RELACIONADA CON
LOS GENES fdxA, fdxB, fldA, oorD, PorD, rdxA y frxA
ÁNGELA MARITZA CARDOSO SANDOVAL
APROBADO
_______________________________________
Dra. Alba Alicia Trespalacios Rangel, MSc.
Directora
______________________________ _____________________________
Dr. William Otero, MD Dr. Hugo Diez, Ph. D
5
RESISTENCIA DE HELICOBACTER PYLORI AL METRONIDAZOL RELACIONADA CON
LOS GENES fdxA, fdxB, fldA, oorD, PorD, rdxA y frxA
ÁNGELA MARITZA CARDOSO SANDOVAL
APROBADO
_____________________________ _______________________________
Ingrid Schuler Ph D. Luz Amparo Maldonado
Decana Académica Directora Carrera de Bacteriología
6
DEDICATORIA
A Dios por acompañarme y guiarme siempre, a mis padres, por creer en mi, por apoyarme en todos mis sueños y por su gran esfuerzo para que culminará mi carrera, a mi hermana por ser mi cómplice, mi amiga, por su apoyo y su paciencia, a mi angelito hermoso por ser el
motor de mi vida, el origen de mi fuerza, la fuente de motivación y por darme la alegría de ser madre.
7
AGRADECIMIENTOS
A la Doctora Alba Alicia Trespalacios por haberme brindado la oportunidad de realizar esta investigación, por su apoyo, por su paciencia, por sus conocimientos y por su dedicación; a la Doctora Olga Raquel Villamizar por guiarme, por su paciencia y por sus consejos. A César por su apoyo incondicional, por creer siempre en mí, por nunca haber soltado mi mano, por cada sonrisa que me regala día tras día. A la Pontificia universidad Javeriana por la formación ética, profesional y por haberme acogido durante estos años.
8
TABLA DE CONTENIDOS
Pág.
Tabla de contenido
Lista de figuras
Lista de tablas
Lista de anexos
Resumen
Abstract
1. INTRODUCCIÓN 16
2. MARCO TEÓRICO 17
2.1. CARACTERISTICAS MICROBIOLOGICAS DE HELICOBACTER PYLORI 17 2.1.1. Morfología de Helicobacter pylori 17 2.1.2. Genoma de Helicobacter pylori 17 2.1.3. Sistema de transporte de solutos, vías metabólicas y cadena
transportadora de electrones 17 2.2. FACTORES DE PATOGENICIDAD DE HELICOBACTER PYLORI 20 2.3. DIAGNÓSTICO DE HELICOBACTER PYLORI 22
2.3.1. Métodos directos 22
2.3.2 Métodos indirectos 23
2.4. PRUEBAS DE SUSCEPTIBILIDAD ANTIMICROBIANA 24 2.4.1. Método de dilución en agar 24
2.4.2 E-TEST 25
2.5. ERRADICACIÓN DE HELICOBACTER PYLORI 26 2.6. RESISTENCIA ANTIMICROBIANA DE HELICOBACTER PYLORI 27 2.6.1. RESISTENCIA ANTIMICROBIANA DE HELICOBACTER
PYLORI AL METRONIDAZOL 28
2.6.1.1. Gen rdxA y frxA 31 2.6.1.2. Gen oorD y PorD 35 2.6.1.3. Gen fldA, fdxA y fdxB 37 2.7. TECNICAS DE BIOLOGIA MOLECULAR PARA LA IDENTIFICACIÓN
9
2.7.1. Reacción en cadena de la polimerasa (PCR) 38 2.7.2. Reacción en cadena de la polimerasa de transcriptasa
inversa (RT-PCR) 43
2.7.3 Amplificación aleatoria de DNA polimórfico 45 3. FORMULACIÓN DEL PROBLEMA Y JUSTIFICACIÓN 47 3.1. FORMULACIÓN DEL PROBLEMA 47
3.2. JUSTIFICACIÓN 47
4. OBJETIVOS 49
4.1. OBJETIVO GENERAL 49
4.2. OBJETIVOS ESPECIFICOS 49
5. METODOLOGIA 50
5.1. TIPO DE INVESTIGACIÓN 50 5.2. SELECCIÓN DE ARTICULOS 50 5.3. DEFINICIÓN DE CRITERIOS PARA LA INCLUSIÓN Y EXCLUSIÓN
DE LOS ARTICULOS 50
5.3.1. Criterios de inclusión 50 5.3.1.1. Tipos de artículos 50 5.1.3.2. Tipo de estudio designado para la investigación 50 5.1.3.3. Año de publicación de los artículos 51 5.3.2. Criterios de exclusión 51 5.4. IDENTIFICACIÓN Y SELECCIÓN DE ARTICULOS 51 5.4.1. Estrategia de búsqueda para la identificación de artículos 51 5.4.2. Selección de los artículos por mas de un observador 52 5.4.3. Discusión de los artículos 52 5.5. RECOLECCIÓN DE LA INFORMACIÓN 52 5.6. EXTRACCIÓN DE LOS DATOS 52 5.7. ELABORACIÓN Y ESCRITURA DEL DOCUMENTO 53
6. RESULTADOS 54
6.1. ANALISIS DE LA INFORMACIÓN 54 6.2. FRECUENCIA DE MUTACIONES DE LOS GENES rdxA Y frxA 61 6.3. TECNICAS DE BIOLOGIA MOLECULAR 66
7. DISCUSIÓN 68
7.1. SELECCIÓN DE LOS ARTICULOS 68 7.2. DISCUSIÓN DE LOS RESULTADOS 68
8. CONCLUSIONES 71
10
11
Lista de tablas
Pág.
Tabla 1. Esquemas de erradicación de Helicobacter
pylori 27
Tabla 2. Compuestos antimicrobianos modo de acción y
mecanismos de resistencia 28
Tabla 3. Mutaciones de los genes rdxA y frxA en cepas
resistentes al metronidazol relacionadas con la CMI 32
Tabla 4. Mutaciones de los genes rdxA en cepas resistentes de
metronidazol 34
Tabla 5. Parejas de primers utilizados para la amplificación de los
genes rdxA y frxA Helicobacter pylori 40
Tabla 6. Características de los estudios para la amplificación de los
genes rdxA y frxA 54
Tabla 7. Frecuencia de mutaciones en el gen rdxA 61
Tabla 8. Frecuencia de mutaciones en el gen frxA 65
Tabla 9. Comparación característica de las técnicas de biología molecular
12
Lista de figuras
Pág.
Figura 1. Modelo de la obtención de hierro 19
[image:17.612.109.525.150.644.2]Figura 2. Ciclo del ácido tricarboxílico en H. pylori 19
Figura 3. Prueba de E-TEST 25
Figura 4. Activación del metronidazol 30
Figura 5. Descarboxilación de piruvato en Helicobacter pylori 36
Figura 6. Metabolismo del piruvato y activación reductiva del metronidazol
por Helicobater pylori 37
Figura 7. Reacción en cadena de la polimerasa (PCR) 39
Figura 8. Secuenciación del DNA por PCR en el estudio de los genes
rdxA y frxA 42
Figura 9. Reacción en cadena de la polimerasa de transcriptasa inversa
(RT-PCR) 43
Figura 10. Amplificación aleatoria del DNA polimórfico 46
Figura 11. Resistencia antimicrobiana de los aislamientos de Helicobacter
pylori 60
Figura 12. Nivel de resistencia al metronidazol 61
Figura 13. Frecuencias de mutaciones gen rdxA 64
13
Lista de anexos
14
RESUMEN
Este trabajo de grado tiene como objetivo principal, realizar una revisión bibliográfica para conocer la resistencia de Helicobacter pylori al metronidazol asociada a los genes rdxA, frxA, fldA, fdxA, fdxB, oorD y PorD. Se realizó una búsqueda de literatura en las bases de datos de EBSCO, Pubmed, Science direct, CDSR, Elsiver y Scielo de estudios de la resistencia antimicrobiana de H. pylori al metronidazol, la actividad, diagnóstico y asociación de las alteraciones de los genes que codifican las nitrorreductasas en dicha resistencia. Se seleccionaron los artículos de acuerdo a los criterios de inclusión por los dos observadores, se obtuvo la frecuencia de las mutaciones de los genes rdxA y frxA recopilando y analizando la información reportada por cada uno de los documentos.
De los 64 estudios obtenidos en la primera búsqueda, se seleccionaron 31 artículos, identificando que solamente 9 cumplían con los criterios de inclusión. Se observó que el 44.31% (117/264) de los aislamientos son resistentes al metronidazol. De las 117 cepas resistentes, el 18.80% (22/117) presentó un nivel de resistencia alto al metronidazol, el 76.92% (90/117) un nivel intermedio o moderado de resistencia y el 0.85% (1/117) restante un nivel de resistencia bajo. Se encontró 326 mutaciones en las cepas resistentes, el 74.23% (242/326) correspondieron a alteraciones en el gen rdxA, el 25.76% (84/326) en el gen frxA.
La técnica más empleada para el estudio de las alteraciones genéticas en la resistencia de
H. pylori al metronidazol, fue la reacción en cadena de la polimerasa (PCR).
Se encontró que existe una gran variedad de alteraciones en los genes asociadas a la resistencia al metronidazol. Se observó, que las mutaciones de tipo cambio de marco que resultan en la truncación de la proteína y las mutaciones de sentido erróneo fueron las más frecuentes en el estudio de la resistencia por los genes rdxA y frxA, la presencia de alteraciones solo en el gen frxA confiere resistencia, encontrando que en una de las cepas, la mutación de una región del gen frxA generó resistencia al metronidazol sin estar acompañada de una alteración en otro gen, además, la causa de la resistencia no implica la alteración del gen, encontrando en los estudios mutaciones en cepas sensibles al metronidazol, lo cual implica que incluso en ausencia de un rdxA funcional, una cepa de
15
ABSTRACT
The main purpose of this graduation work is to do a bibliographic revision in order to know the resistance of Helicobacter pylori at metronidazole associated to genes rdxA, frxA, fldA, fdxA, fdxB, oorD and PorD. A search of literature from the data base of EBSCO was done, Pubmed, Science direct, CDSR, Elsiver and Scielo of studies of the anti-microbial resistance of H. pylori at metronidazole, the activity, diagnosis and association of the genes alterations which codify the nitrorreductasas in such a resistance. Articles were selected according to inclusion criteria by the two observers, mutations frequency of genes rdxA and frxA was obtained compiling and analyzing the reported information by each one of the documents.
Out of the 64 studies obtained during the first search, 31 articles were selected, detecting that only 9 of them met the inclusion criteria. It was observed that 44.31% (117/264) of the isolated cases are resistant to metronidazole. Out of the 117 metronidazole resistance isolates, the 18.80% (22/117) presented a high level metronidazole resistance, the 76.92% (90/117) a moderate level resistance and the remaining 0.85% (1/117) a low level metronidazole resistance. 326 mutations occurred in the resistant roots (bacterium), 74.23% (242/326) corresponded to alterations in gene rdxA and 25.76% (84/326) in gene frxA.
The most used technique for the study of genetic alterations in the resistance of H. pylori at metronidazole was the polymerase chain reaction (PCR).
It was found that there is a great variety of alterations in genes, associated with resistance to metronidazole. It was observed that frameshift mutations that end up in shortening of protein, and missense mutations, were the most frequent ones in the study of resistance for genes
rdxA and frxA. Presence of alterations was only in gene frxA causing resistance, finding that in one of the bacteria, mutation of a region of gen frxA caused resistance to metronidazole without being accompanied of an alteration in other gene, besides that, the cause of resistance does not imply alteration of gen, finding in studies, mutations in sensible isolates to metronidazole, which implies that even in the absence of an rdxA functional, a bacteria of
16
1. INTRODUCCIÓN
La bacteria Helicobacter pylori afecta a más de la mitad de la población mundial, la infección por esta bacteria es una de las principales causas de gastritis crónica, úlcera gástrica ó duodenal, linfomas de mucosas tipo MALT y adenocarcinoma de estómago. En la mayoría de los casos, la infección se adquiere durante la infancia y puede permanecer durante toda la vida si no se inicia un tratamiento para la erradicación de la bacteria. La prevalencia es alta en países subdesarrollados y se relaciona con las condiciones sanitarias, y el nivel económico de la población. En México, América Central, América del Sur y África, la prevalencia es cercana al 70-90%; en Europa oriental es aproximadamente del 70%, en Asia del 70-80%, Europa occidental, Canadá y Estados Unidos con cifras del 30% y Australia con un 20%. (Hunt RH, et al. 2006).
La resistencia antibiótica de la bacteria es un obstáculo para su erradicación y motivo de diversos estudios desde hace algunos años; la resistencia al Metronidazol, es la resistencia antibiótica más alta y frecuente en Helicobacter pylori, y se relaciona con el uso periódico de metronidazol y nitroimidazoles para el tratamiento de enfermedades ginecológicas, dentales y parasitarias.
Las mutaciones ó deleciones de genes que codifican nitrorreductasas presentes en microorganismos anaerobios y algunos microaerófilos, se relacionan con la resistencia al metronidazol; los principales genes involucrados son el gen rdxA que codifica la nitrorreductasa NAD(P)H insensible al oxigeno y el gen frxA que codifica la NAD(P)H Flavina Oxidorreductasa; la función principal dentro de la bacteria se encuentra en la oxidación del piruvato y en el transporte de electrones como donantes directos de electrones.
17
2. MARCO TEORICO
2.1. CARACTERISTICAS MICROBIOLOGICAS DE HELICOBACTER PYLORI
2.1.1. Morfología de Helicobacter pylori
Helicobacter pylori es una bacteria con forma espiral, gram negativa, microaerofílica, que requiere de niveles de dióxido de carbono para su crecimiento, mide 2.5 a 3.5 µm de largo y 0.5 a 1.0 µm de ancho; tiene de 4 a 6 flagelos unipolares que son esenciales para la motilidad bacteriana. Cada flagelo tiene aproximadamente 30 µm de largo y pueden cambiar a una forma cocoide especialmente después de tratamiento antibiótico ó después de un cultivo prolongado (Gerrits, 2004).
2.1.2. Genoma de Helicobacter pylori
El tamaño del genoma de Helicobacter pylori tiene un rango de 1.6 a 1.73 Mb, tiene un contenido promedio de 39% de G+C. Aproximadamente en el 40% de las cepas de H. pylori el tamaño de los plásmidos tiene un rango de 1.5 a 23.3 kb, pero estos plásmidos no posee factores de virulencia reconocidos, posee al menos dos copias cada uno de los genes rRNA 16S y 23S. La bacteria presenta diversidad de genes que codifican la ureasa estructural, proteínas, flagelina, citotoxina vacuolizante y CagA (Dunn et al., 1997).
2.1.3. Sistema de transporte de solutos, vías metabólicas y cadena transportadora de electrones
La bacteria tiene una capacidad limitada para la biosíntesis, por este motivo, se necesita de un gran número de sistemas de transporte para la adquisición de aminoácidos esenciales, iones y nutrientes del medio externo. Los sistemas de transporte primario dependen de la hidrólisis de ATP; los transportadores secundarios vinculados a iones son utilizados para la incorporación de solutos incluidos los aminoácidos, ácidos carboxílicos (lactato, 2-oxoglutarato, fumarato, malato) y varios iones (Kelly, 2001).
18
Helicobacter pylori expresa proteínas reguladoras de hierro en la membrana externa, que sirven como receptores para la absorción del grupo hemo. La lactoferrina humana (Lf) puede ser usada como única fuente de hierro, en la bacteria fue identificada una proteína putativa ligada a Lf de 70 kDa (Lpb), esta proteína solo se expresa cuando la bacteria crece en un medio que no contiene hierro, se cree que esta involucrada en el proceso de obtención de este elemento (Velayudhan et al., 2000).
Se ha encontrado que la bacteria codifica diversos sistemas de oxidación de hierro, posee homólogos de un número de genes fec de Escherichia coli que codifica un transportador de dicitrato férrico, tres genes ligados a fecA, el fecA1, fecA2 y fecA3, codifican un receptor de membrana externa para el dicitrato- Fe3+, el transporte de dicitrato férrico a través de la
membrana externa requiere un complejo de transducción de energía constituido por proteínas de la membrana citoplasmática TonB, ExbB y ExbD (Velayudhan et al., 2000).
Helicobacter pylori también posee un sistema vinculado a hierro ferroso (Proteína FeoB) que es importante en la asimilación de hierro bajo condiciones microaerofílicas (Kelly, 2001).
Bajo condiciones de depleción de hierro, la absorción ocurre principalmente por la vía FeoB, las flavinas como FMN, FAD y riboflavina, son reducidas en el citoplasma en forma de Fldred
y secretadas fuera de la membrana, el genoma de Helicobacter pylori contiene un homologo de NADP(H) flavina oxidorreductasa que es el responsable principal de dicha reducción, las Fldred median la transferencia de electrones de Hierro férrico Fe3+ a Hierro ferroso Fe2+. El
Fe2+ se difunde a través de las porinas de la membrana externa y transportado dentro del citoplasma por medio de la FeoB, la hidrólisis de ATP es inhibida por el vanadato. (Figura 1) El sistema Fec es utilizado cuando existen condiciones deficientes o bajas de hierro, por esta vía la absorción del Fe3+ es mediada por la FecA, proteína de la membrana externa, una proteína periplásmica de unión al soluto (PBP) y FecDE (Velayudhan et al., 2000).
Las principales vías de catabolismo de la glucosa que utiliza Helicobacter pylori son la vía de las pentosas fosfato y la vía de Entner-Doudoroff, además, la bacteria no posee enzimas que evidencien las reacciones anapleróticas para reposición del oxalacetato.
19
Figura 1. Modelo de obtención de Hierro (Velayudhan et al., 2000)
El primer paso de la vía oxidativa es catalizado por la citrato sintasa, el Acetil-CoA y oxalacetato son condensados para generar citrato y coenzimaA (CoA). Luego se genera isocitrato a partir de la isomerización del citrato vía cis-aconitato. La isocitrato deshidrogenasa cataliza la descarboxilación oxidativa dependiente NAD(P) de isocitrato para generar 2-oxoglutarato y CO2 (Mobley et al., 2001).
La descarboxilación de 2-oxoglutarato en presencia de Coenzima A (CoA), genera succinil-CoA con liberación de CO2, reacción catalizada por la enzima 2-oxoglutarato oxidorreductasa (OorD), compuesta de cuatro subunidades OorA, OorB, OorC y OorD. Se ha concluido que la ausencia del 2-oxoglutarato oxidorreductasa afecta la generación de succinil-CoA por medio de esta vía (Mobley et al., 2001).
La succinil CoA sintasa cataliza la conversión de Succinil-CoA a succinil y CoA, así, en estas reacciones interviene enzimas como la fumarasa y malato deshidrogenasa (Figura 2).
[image:24.612.195.428.516.682.2]20
El transporte de electrones cumple dos funciones; transferir los electrones de un donador a un aceptor y conservar parte de la energía que se libera en ese proceso para la síntesis de ATP. En este proceso de oxido-reducción están involucradas varias enzimas, entre las cuáles se encuentran NADH deshidrogenasa, flavoproteínas que contienen Flavín mononúcleotido (FMN) ó Flavín-adenina dinúcleotido (FAD), proteínas que contienen hierro y azufre, citocromos y transportadores no proteicos como las quinonas (Mobley et al., 2001).
Las quinonas son transportadores que median la transferencia de electrones entre la deshidrogenasa y compuestos oxidados ó reducidos de la cadena respiratoria. Las bacterias contienen dos tipos de quinononas: las benzoquininas como la ubiquinona y naftoquinonas como menaquinona; la quinona se reduce a quinol como resultado de la oxidación de substrato por las deshidrogenadas respiratorias; el quinol actúa como un reservorio de poder de reducción en la membrana citoplasmática, y puede ser utilizado como una fuente de electrones por varios complejos de proteínas rédox, el más importante es el complejo citocromo bc1, participando en la transferencia de la reducción de dos donantes de electrones (quinol) a un aceptor de electrones (citocromo c) y contribuyendo a la generación de una fuerza motriz de protones a través de la membrana citoplásmica (Mobley et al.,
2001).
Se ha identificado un único terminal oxidasa, el citocromo c oxidasa tipo bc; además hay tres tipos de deshidrogenasas que pueden ser donadores potenciales al pool menaquinona: glicerol-3-fosfato deshidrogenasa, D-lactato deshidrogenasa y una multisubunidad quinona oxidorreductasa. Los intermediarios de transporte de electrones entre el pool de menaquinona y la oxidasa son el citocromo BC1 y un complejo citocromo c553 (Kelly, 2001).
2.2. FACTORES DE PATOGENICIDAD DE HELICOBACTER PYLORI
La infección por H. pylori se adquiere de tres maneras; una transmisión oral-oral, mediante el desplazamiento de la bacteria del estómago a la boca provocado por reflujo gastroesofágico,
Gástrica-oral, transmisión del microorganismo a través del vómito; y por último la Fecal-oral,
contaminación con materia fecal (Rivas et al., 2000).
21
La adhesina BabA ha sido la más estudiada, facilita la unión de los microorganismos a los antígenos de Lewis b, las enzimas fosfolipasas A2 y C degradan los principales componentes lipídicos de la membrana.
Los lipopolisácaridos que poseen en su antígeno “O” constituyen otro factor de virulencia,
estos son los carbohidratos de Lewis “x” y Lewis “y” ó ambos. Los antígenos de Lewis participan en la patogénesis ayudando al microorganismo a evadir la respuesta inmune durante la colonización en el estómago, induce una respuesta autoinmune contra los antígenos de Lewis que expresa H. pylori y que son compartidos por las células de la célula contribuyendo a un daño directo o indirecto (Cervantes, 2006).
La citotoxina vacuolizante VacA está codificada por el gen vacA; es una proteína de la membrana externa, presente en todas las cepas; es una citotoxina vacuolizante, que causa alteraciones celulares, además bloquea la presentación antigénica y la activación de los linfocitos T, la proteína CagA está codificada por el gen cagA, y a diferencia de la VacA no está presente en todas las cepas de Helicobacter pylori; quienes la poseen, se adhieren al epitelio induciendo la liberación de interleucina-8 y la transcripción del factor AP-1 y la cascada de las cinasas, permitiendo la expresión de los proto-oncogenes; la OipA favorece la adherencia e induce la inflamación local, estimulando la secreción de la IL-8.
Además, tiene otros factores de virulencia que contribuyen a su protección y estadía en la mucosa gástrica y a la formación de ulceraciones durante la infección como la ureasa, enzima que cataliza la hidrólisis de urea para proporcionar amonio y carbamato, el cual se descompone para producir otra molécula de amonio y ácido carbónico, con lo cual neutraliza el microambiente; el efecto neto de esas reacciones hace que se aumente el pH, contrarrestando el ácido de la cavidad gástrica. El resultado del ión hidróxido generado por el equilibrio del amonio con el agua a partir del hidróxido de amonio, contribuye significativamente al daño histológico asociado con la infección; la urea indirectamente también es responsable del daño tisular mediante su interacción con el sistema inmune, porque estimula el estallido respiratorio de neutrófilos.
22
2.3. DIAGNÓSTICO DE HELICOBACTER PYLORI
El diagnóstico de H. pylori se puede realizar por dos métodos directos (invasivos) ó indirectos (no invasivos).
2.3.1. Métodos directos
1. Prueba de ureasa rápida (en una biopsia del antro): La bacteria posee la enzima ureasa que facilita la colonización y permanencia en la cavidad gástrica, cumple tres funciones principales en la protección frente al ácido de la mucosa gástrica, la provisión de nitrógeno en forma de amonio y como factor de virulencia en la patogenia de la úlcera gástrica; en general, en esta prueba la enzima hidroliza la urea provocando la liberación de CO2 y NH4,
con el consiguiente aumento de pH (Kelly, 2001).
2. Detección microscópica de la bacteria: La observación puede realizarse de improntas de biopsia ó en cortes histológicos introducidas en formaldehído al 10%, no hay tinciones especificas para H. pylori; en la muestra se observan infiltraciones de células inflamatorias con desarrollo de agregados linfocitarios intramucosa y en folículos; se utilizan tinciones como la de Hematoxilina-eosina para la identificación de gastritis y las secuelas clínicas de la infección como carcinoma gástrico o linfoma MALT, poco fiables para la identificación de la bacteria. Las tinciones Gram, Giemsa modificada y Warthin-starry, se utilizan para la identificación de la bacteria. La combinación de Hematoxilina-eosina, azul alcián y genta, permite identificar fácilmente a la bacteria y lograr una excelente visualización de la morfología gástrica (Mobley et al., 2001).
3. Cultivos microbiológicos: Para el cultivo se utilizan medios que contienen una base de agar, suplementos para el crecimiento y suplementos selectivos; la base de agar que se utiliza son el agar cerebro corazón, agar Columbia y agar Wilkins Chalgren, como suplementos de crecimiento, se agrega sangre o suero de oveja-caballo, que incluye numerosos nutrientes (vitaminas y oligoelementos); preferiblemente, la proporción de sangre es de 10%. Otros suplementos que se utilizan son la yema de huevo, carbón, almidón, albúmina sérica bovina y la catalasa. El sulfato ferroso y el piruvato de sodio proporcionan mucina que mejora el crecimiento de la bacteria; la temperatura óptima de incubación es de 37ºC, en condiciones microaerofílicas (10% de CO2), sin embargo, puede crecer a 30ºC ó
42ºC; las colonias aparecen 3 ó 4 días después de haber sido cultivadas (Mégraud et al.,
23
4. Reacción en cadena de la polimerasa (PCR): El DNA bacteriano puede ser utilizado como marcador de la infección, la PCR tiene como fin, el amplificar in vitro un número de copias de una región específica de DNA, para ser evaluadas, además, permite diferenciar y tipificar diversos genotipos (Mégraud et al., 2007).
2.3.2. Métodos indirectos
1. Prueba del aliento: La ureasa producida por H. pylori tiene la capacidad para hidrolizar una solución de urea marcado previamente con isótopo 13C ó 14C, la solución de urea
marcada es administrada al paciente; si está infectado, ocurrirá la hidrólisis, resultando el carbono marcado libre, que es absorbido por la sangre y exhalado en el aire aspirado, pero si no está infectado, no se producirá el carbono marcado y la mayoría de los isótopos serán eliminados por la orina. Se realizan las mediciones antes y 30 minutos después de haber ingerido la solución (Mégraud et al., 2007).
2. Detección de Anticuerpos contra Helicobacter pylori: La infección induce la respuesta inmune local y sistémica que se ven reflejadas en la producción de anticuerpos, en un principio se producen transitoriamente inmunoglobulinas de clase M (IgM), luego, a nivel de la mucosa predomina la producción de inmunoglobulina de clase A (IgA), mientras que en la sistémica se produce fundamentalmente inmunoglobulina de clase G (IgG). Para la detección de los anticuerpos se pueden utilizar muestras como: sangre, suero, saliva, orina ó heces; ésta prueba permite conocer si el paciente ha estado en contacto con la bacteria, la elevación de la concentración de anticuerpos indica la existencia de una infección activa o reciente. La técnica principal que se utiliza para la detección de los anticuerpos es la ELISA, la infección por H. pylori es crónica y predomina la IgG subclase 1 y 4 (Mégraud et al., 2007).
24
2.4. PRUEBAS DE SUSCEPTIBILIDAD ANTIMICROBIANA
Los métodos de susceptibilidad antimicrobiana son procedimientos que determinan la susceptibilidad de los microorganismos a agentes antimicrobianos, la sensibilidad de las bacterias a dichos agentes esta dada por la concentración mínima inhibitoria (CMI) que se define como la concentración más baja de antibiótico que inhibe completamente el crecimiento de la bacteria. La determinación In vitro de la susceptibilidad bacteriana a los antibióticos, se puede realizar por diferentes métodos como difusión en disco, dilución en caldo, dilución en agar y la prueba del epsilómetro (E-TEST), los últimos dos métodos son utilizados con frecuencia para identificar la resistencia de Helicobacter pylori al metronidazol (Coyle, 2000).
2.4.1. Método de dilución en agar
En el método de dilución en agar, el medio de cultivo tiene incorporado el agente antimicrobiano, cada placa de cultivo contiene una concentración diferente, ósea, diluciones definidas del agente antimicrobiano. La inoculación de la bacteria se realiza en la superficie del agar con un replicador, las placas son incubadas, finalizada la incubación, se examinan los cultivos observando si hubo crecimiento o no de la bacteria, de esta manera se determina la CMI (Coyle, 2000).
El Instituto de Estándares Clínicos y de Laboratorio (CLSI) recomienda en el método de dilución en agar para Helicobacter pylori como medio de cultivo, agar Mueller-Hinton suplementado con 5% de sangre de cordero envejecida, para el inoculo, se prepara una suspensión equivalente al tubo 2 de Mc Farland (1x107 a 1x108 UFC/mL) en solución salina
a partir de un subcultivo de 72 horas en agar sangre, con un replicador se aplica el inoculo (1 ó 3 µL) directamente sobre las placas de agar. La incubación se realiza a 35 + 2 °C por 3 días bajo condiciones microaerofílicas producida con un sobre generador de gas, para controlar la técnica se debe utilizar la cepa control H. pylori ATCC 43504 para la que existen límites aceptables de valor de CMI de metronidazol es de 64-256 mg/L. El punto de corte definido por el CSLI de resistencia al metronidazol es > 8 µg/mL (CSLI, 2008).
2.4.2. E-TEST
25
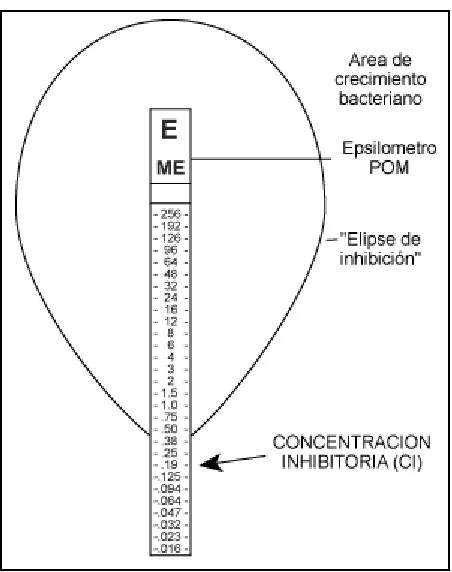
[image:30.612.204.430.251.537.2]exponencial del agente antimicrobiano inmovilizado en un lado de la tira y una escala de interpretación con las concentraciones crecientes de dicho agente en el otro lado. La inoculación de la bacteria se realiza en la superficie de una placa de agar, se coloca la tira sobre la superficie del agar y se incuba, el gradiente antimicrobiano es liberado inmediatamente en el agar, finalizada la incubación, se observa en la placa una elipse de inhibición de crecimiento. La intersección entre el borde de la zona de inhibición y la tira del soporte se produce en la concentración de agente antimicrobiano ó antibiótico que ya no es capaz de inhibir el crecimiento del microorganismo (Figura 3). La concentración inhibitoria es dada por el punto de corte de la intersección (Coyle, 2000).
Figura 3. Prueba de E-TEST (Coyle, 2000).
26
finalizada la incubación, se realiza la lectura, sobre la placa de agar se forma un área de inhibición en forma de elipse, determinando de esta manera la CMI (Piccolomini et al., 2007).
La Sociedad Británica de Quimioterapia Antimicrobiana (BSAC) recomienda en el E-TEST para Helicobacter pylori como medio de cultivo agar Mueller-Hinton ó Wilkings-Chalgren suplementado con 10% de sangre de caballo, para el inoculo, resuspender las colonias de un cultivo de 2 a 3 días de incubación en agua destilada estéril, se realiza una suspensión equivalente al tubo 3 de McFarland, inocular sobre la superficie de la placa de agar con una torunda empapada en esta suspensión, dejar secar, colocar la tira sobre la superficie del agar e incubar 35°C durante 3 a 5 días en condiciones microaerofílicas, se observa la placa y se lee la CMI, el punto de corte definido de la resistencia al metronidazol de H. pylori es de > 8 µg/mL (Alarcón et al., 2004).
2.5. ERRADICACIÓN DE HELICOBACTER PYLORI
In vitro, H. pylori es susceptible a la mayoría de los antibióticos, pero in vivo sólo unos pocos antibióticos pueden ser utilizados con éxito para su erradicación, la monoterapia no es lo suficientemente eficaz para erradicar la bacteria, por esta razón, requiere de una terapia que combine uno o dos antibióticos (Tabla 1), un inhibidor de la bomba de protones (PPI) y/o un componente de bismuto (Gerrits, 2004).
Los medicamentos supresores de ácido como los bloqueadores de H2 ó inhibidores de bomba de protones (IBP) aumentan el pH intragástrico, mejoran la actividad de los antibióticos y reducen lo efectos secundarios de los tratamientos, los IBP son inhibidores irreversibles de la enzima H+/K+ATPasa ubicada en la membrana apical de las células parietales del estómago, una vez activado, se une a un residuo de cisteína de la bomba de protones (enzima H+/K+ATPasa) mediante un enlace covalente y de esta forma bloquea en permanentemente la vía final común de la secreción de ácido gástrico, los bloqueadores de H2 bloquean el efecto de la Histamina, la cuál estimula la secreción de ácido (Gerrits, 2004).
27
Tabla 1. Esquemas de erradicación de Helicobacter pylori (Otero et al., 2007)
Esquema de tratamiento de primera línea
No alérgicos a la penicilina:
IBP o ranitidina-bismuto-citrato dos veces al día más amoxicilina, 1 g dos veces al día más claritromicina, 500 mg dos veces al día por 7 a 10 días; eficacia de 80% a 90%. Si la resistencia a la claritromicina es mayor de 20%, cambie este antibiótico por levofloxacina, 500 mg, 1 vez al día.
Alérgicos a la penicilina:
Subsalicilato de bismuto, 525 mg, por vía oral, cuatro veces al día. Metronidazol, 250 mg, cuatro veces al día.
Tetraciclina, 500 mg, cuatro veces al día.
Ranitidina o IBP, dosis estándar dos veces al día. Una o dos semanas; eficacia de 75% a 90%.
En Colombia, por la alta resistencia al metronidazol, las tasas de eficacia de este esquema son de 30% a 70%; por este motivo se utiliza el siguiente esquema:
Subcitrato de bismuto, 120 mg, cuatro veces al día. Tetraciclina o amoxicilina, 500 mg, cuatro veces al día. Furazolidona, 100 mg, cuatro veces al día.
Dos semanas; eficacia de 85% a 87%.
Si el esquema anterior ha fallado se recurre a tratamientos de rescate.
Esquema de rescate
Levofloxacina, 250 mg, 2 veces al día, ó 500 mg, 1 vez al día (durante 7 a 10 días).
Amoxicilina, 1 g, 2 veces al día o rifabutina, 500 mg, 1 vez al día IBP o IBP, dos veces al día. Tetraciclina, cuatro veces al día.
Amoxicilina, 500 mg, cuatro veces al día. Furazolidona, 100 mg, cuatro veces al día. 10 a 14 días; eficacia de 86%.
Rifabutina, 300 mg, una vez al día. Amoxicilina, 1 g, dos veces al día. IBP, dos veces al día.
7, 10, 14 días; eficacia de 70% a 79%.
Esquema de rescate después de dos fracasos previos
Esquema triple con levofloxacina o cultivar el H. pylori y, según su sensibilidad, elegir los antibióticos.
2.6. RESISTENCIA ANTIBIÓTICA DE HELICOBACTER PYLORI
28
[image:33.612.124.523.153.452.2]Los mecanismos moleculares de resistencia a los antibióticos se han caracterizado hasta el momento por mutaciones puntuales localizadas en el cromosoma (Gerrits, 2004).
Tabla 2. Compuestos antimicrobianos, modo de acción y mecanismos de resistencia
Compuesto
antimicrobiano Modo de acción Mecanismo de resistencia
Metronidazol
Reducción del metronidazol por nitrorreductasas que posee la bacteria para dar lugar a la formación de radicales nitro e intermediaros del metronidazol, causando daños estructurales en el DNA de la bacteria.
La ausencia de la reducción del metronidazol por mutaciones en genes
rdxA, fdxA, fdxB, fldA,
oorD, PorD y frxA que codifican dichas nitrorreductasas.
Claritromicina Unión a la subunidad ribosomal rRNA 23s, inhibiendo la síntesis proteica.
Mutaciones puntuales en la subunidad 23s
Amoxicilina
Unión a las proteínas de unión a la penicilina (PBPs), y, por tanto, interfiere con síntesis de la pared celular bacteriana.
Alteraciones estructurales de las PBPs, disminución de la permeabilidad de la membrana, disminución de la unión amoxicilina A PBP1A (resistencia) ó PBP-D (tolerancia).
Tetraciclina
Se acumula en el citoplasma donde se une a la subunidad ribosomal 30s, interfiriendo con la fijación de aminoacil-tRNA al ribosoma e inhibe la síntesis y crecimiento bacteriano.
Mutaciones puntuales en genes rrnA y rrnB que codifican la subunidad 16S rRNA.
2.6.1. Resistencia al metronidazol
Los mecanismos de resistencia al metronidazol de H. pylori que han sido estudiados son (Gerrits, 2004):
1. Deficiencia de la captación del antibiótico y el aumento del flujo de salida del medicamento:
29 Existen tres sistemas básicos de captación:
1. La captación de sustancias hidrofílicas a través del llenado de agua de los canales de porinas.
2. La captación de policationes a través de la vía que auto-promueve la absorción en sitios de unión de cationes divalentes en lipopolisácaridos.
3. La captación de sustancias hidrofóbicas a través de la membrana externa.
La resistencia antibiótica implica la permeabilidad limitada de la membrana externa con mecanismos de defensa secundarios como beta lactamasas periplásmicas o el flujo de salida del antibiótico.
Uno de los sistemas de flujo de salida asociado con la resistencia es el sistema de Resistencia-Nodulación-División (RND), este sistema depende de la fuerza protón motriz, la desactivación por mutaciones, hace a la bacteria hipersusceptible a los antibióticos, el aumento conduce a múltiples resistencias (Bina et al., 2000).
Aunque no se ha establecido que los sistemas de flujo de salida están implicados en la múltiple resistencia antibiótica, se encontró en el genoma de H. pylori 26695 la presencia de translocasas y proteínas de membrana externa (OEPs) ó proteínas vinculadas a TolC que pueden interactuar para formar el sistema de flujo de salida involucrado en la resistencia antibiótica (Van Amsterdam et al., 2005).
2. Aumento de la actividad de las enzimas que reparan el DNA:
El gen recA codifica una proteína que juega un papel esencial en la recombinación y reparación del DNA, esta proteína es capaz de inducir ó activar el sistema SOS, los efectos de SOS incluyen el incremento de la reparación del DNA, incremento en la tasa de mutación, inhibición de la división celular e inducción de profase (Thompson et al., 1995).
Los daños que se presentan en el DNA de la bacteria por acción de los antibióticos son reparados con mayor facilidad por acción de este sistema SOS lo que conlleva a un incremento en la resistencia.
3. Capacidades aumentadas en la captación de oxígeno:
30
La sensibilidad o resistencia a metronidazol depende de la ausencia o presencia de oxígeno en el lugar de la reducción de metronidazol. El metronidazol compite con el oxígeno por los electrones de la flavodoxina, y puede no ocurrir la activación del metronidazol, pero si el metronidazol es reducido por la flavodoxina y posteriormente oxidado al compuesto inactivo, no puede ejercer sus funciones, siendo un mecanismo de resistencia al antibiótico (Smith et al., 1997).
4. Disminución de la activación del antibiótico derivada de las enzimas que reducen el metronidazol:
[image:35.612.175.480.402.558.2]El Metronidazol es activado, por acción de las nitrorreductasas que lo reducen generando intermediarios nitrosos y el metabolito activo la hidroxilamina (radical libre), que daña el DNA, provocando la ruptura, desestabilización, desenrollamiento de la doble hélice y muerte celular (Figura 4) , el medicamento tiene un potencial rédox bajo y es efectivo en organismos con este mismo estado de rédox, como bacterias anaeróbicas y protozoarios, Trichomonas vaginalis y también microaerofílicos como Helicobacter pylori, que crecen mejor a tensiones bajas de oxígeno, este potencial rédox tan bajo hace al metronidazol un eficiente aceptor de electrones (Trend et al., 2001).
Figura 4. Activación del metronidazol (Gerrits, 2004).
La resistencia al metronidazol esta dada por la inactivación y cambios en estos genes que codifican dichas nitrorreductasas presentes en anaerobios y algunos microaerófilos, estudios han demostrado que los principales genes involucrados en esta resistencia son el gen rdxA
31
la oxidación del piruvato y en el transporte de electrones, como donantes directos de electrones (Trend et al., 2001).
Se ha sugerido que la resistencia al metronidazol esta involucrada con otros genes que codifican enzimas como ferredoxina (fdxA), proteína ligada a ferredoxina (fdxB), flavodoxina (fldA), 2-oxoglutarato oxidorreductasa (oorD), piruvato/ferredoxina oxidorreductasa (PorD) (Marais et al., 2003).
2.6.1.1. Gen rdxA y frxA
El gen rdxA codifica la enzima nitrorreductasa NAD(P)H insensible al oxígeno que actúa reduciendo componentes nitroaromáticos, a través de la reducción secuencial de dos electrones, generando intermediarios y productos finales, como la hidroxilamina que se genera por la reducción del metronidazol, produciendo daño al DNA y muerte celular (Aldana
et al., 2005).
Se ha relacionado la resistencia de Helicobacter pylori al metronidazol con la inactivación mutacional de este gen, Jenks et al., evaluó la contribución del gen rdxA en el desarrollo de resistencia a esta droga, usando un modelo de ratón para determinar la secuencia de este gen en cepas sensibles y resistentes al metronidazol, encontró que las cepas resistentes a esta droga presentan mutaciones por cambio de marco, siete de las doce mutaciones que se presentaron por cambio de marco fueron el resultado de la pérdida o ganancia de adenina (A) o de timina (T), se describieron mutaciones de sentido erróneo, tres de las cepas resistentes contienen la sustitución de aminoácidos como la prolina (P) por la leucina (L), la tirosina (Y) por la histidina (H) y la alanina (A) por la valina (V) (Jenks et al., 1999).
El gen frxA codifica la NAD(P)H Flavina Oxidorreductasa que cataliza la reducción de flavinas libres usando NADH o NADPH como donador de electrones. También, esta involucrada con la reducción del metronidazol para la formación de hidroxilamina.
32
mientras que las cepas sensible (MtzS) tienen un CMI menor de 0.016 a 4 ug/mL, la mayoría de la mutaciones en el gen rdxA son por cambios del nucleótido Guanina por Timina ó Adenina, (Han et al., 2006) pero las mutaciones de estos genes no son fijas (Tabla 3)
Tabla 3. Mutaciones de los genes rdxA y frxA en cepas resistentes al metronidazol relacionadas con la CMI
Cepa CMI (ug/mL) Gen Mutado Secuencia de nucleótido Cambio de
aminoácidos Referencia B-4 12.5 frxA AAC a AGC N124S Aldana et al.
2005
C-4 100 rdxA
frxA GTG a GCG AGC a AAC AAA a ATA ATG a AGG
S127N V69A K126I M149R
D-4 200 frxA TAT a GAT Y62D
E-4 100 frxA GCA a ACA
GTT a GCT TCA a GCA ACT a TTC
A16T V211A
S43A I44F
0844 8 rdxA
frxA G169 T A104C E35A V57F Han et al., 2006
1247 8 rdxA
frxA G184A G624T
V62I K208N
0937 12 rdxA
frxA G391T G56A E131stop C19Y
0888 16 rdxA
frxA
G223T C394T
E75stop E132stop
0871 32 frxA Inserción TT
160-1; A163T L54F; K55stop
1021 48 rdxA
frxA
G103T T540A
E35stop Y180stop
1221 128 rdxA
frxA C148T G482T G624T Q50stop C161F K208N
1202 128 rdxA Deleción T162
G163T V55stop F54L
1122 128 rdxA
frxA Deleción A54 G401A T116A
S134N K18N L39stop
1199 256 rdxA
frxA A187T Deleción T271 G295T K63stop Y91M V99stop
0940 >256 rdxA
frxA G401A C215T A217G
33
196 128 rdxA
frxA Y141stop L74stop Marais 2003 et al.
553 128 rdxA N73stop
166 128 rdxA
frxA L711, F72S, G73S S81L
548 128 rdxA
frxA T31E, R90K, A 108S A40V
220 64 rdxA
frxA Y141stop, R16H A135E
CAS 32 rdxA
frxA L132stop L39stop
313 32 frxA D62Y
RIG 32 rdxA
frxA A40stop R16C
Goodwin et. al., mostraron que la resistencia al metronidazol en H. pylori se debe a mutaciones nulas en el gen rdxA, propusieron que las células que contienen este gen mutado (inactivado), son incapaces de transformar el metronidazol a productos tóxicos como la hidroxilamina. Transformaron cepas sensibles a resistentes al metronidazol utilizando cósmidos, encontraron que E. coli resistente al metronidazol se hizo sensible por un gen
34
Tabla 4. Mutaciones de los genes rdxA en cepas resistentes al metronidazol
Número
de Cepa Nucleótido mutado Cambio de proteína Referencia
1 1 1 1 1 1
A G A G A G G A
A G, C T y otros A G, C T y otros
R200G Y47C, A143T
Q50R, K63E A80Y G145V
Cambio de 8 aminoácidos
Goodwin et. al.,1998
FP2 FR4 FR47 FP21 FP22 FR245 FR7 FP11 FP25 FP34 FR1 FR189 FR17 FR35 +1A +1T +GGCT +1A +1A +1A +CG - 206bp TG a CA
C a T Mutación de sentido
erróneo Mutación de sentido
erróneo Mutación de sentido
erróneo Mutaciones de sentido
erróneo 22stop 22stop 35stop 73stop 73stop 72stop
Cambio de C-terminal 81stop
C16H S43L S18F, H97T y G22
C187T K179R P180S y C184Y
Tankovic et. al., 2000
AR178 ARS AR13 APB APK AP208 AP120 AP81 AR27 AR9 +1T +1A G a T Insertion de IS605
-6 bp A a G C a T C a A Mutación de sentido
erróneo Mutación de sentido
erróneo 59stop 73stop 74stop 99stop M21F T58A A67V A187D A80T, P115L, G170S
G163V 350a2 7a 84a 342c2 136c2 Inserción A141 Sustitución T148 por
G
Sustitución T148 por G
Sustitución T223 por G
Sustitución523 T por G
TAA codón de parada 175 TAG condón de parada148 TAG condón de parada148 TAA condón de parada 223 TAG condón de parada 523
35 El nivel de resistencia al metronidazol se clasifica en:
1. Nivel alto de Resistencia: Contiene una mutación prematura de ambos genes rdxA y frxA.
2. Nivel Intermedio de Resistencia: Contiene una sola mutación prematura de cualquiera de los dos genes rdx ó frxA.
3. Nivel Bajo de Resistencia: Contiene una única mutación en el gen frxA pero no cambio específicos en rdxA. (Marais et al., 2003).
Como se mencionó anteriormente, los niveles de resistencia del metronidazol están relacionados con la concentración mínima inhibitoria (CMI), el punto de corte del metronidazol es de una CMI de 8 µg/mL ó mas, que corresponde al nivel bajo de resistencia (Kwon et al., 2000c, Aldana et al., 2005).
Kwon et al. y Jeong et al., demostraron en sus estudios que la inactivación de rdxA resulta de un nivel moderado de resistencia al metronidazol, con una CMI entre 16-32 ug/mL, mientras que la doble mutación de rdxA y frxA contienen un nivel alto de resistencia, CMI: >64 ug/mL (Kwon et al., 2000c, Jeong et al., 2000, Yamamoto et al., 2002).
2.6.1.2. Gen PorD y oorD
En microorganismos anaeróbicos y protozoarios, los electrones producidos por el complejo piruvato oxidorreductasa (POR) de la descarboxilación de piruvato son donados a la ferredoxina o flavodoxina (figura 5), estas proteínas transportadoras de electrones en un rédox bajo, reducen otro componente, usualmente un protón, que actúa como un aceptor final de electrones, el metronidazol, que tiene un potencial de rédox bajo, es activado para aceptar electrones provenientes de la reducción de ferredoxina o flavodoxina (Kwon et al.,
36
Figura 5. Descarboxilación de piruvatoen H. pylori (Hughes et al., 1997).
Otro mecanismo propuesto en estos microorganismos anaeróbicos, expuestos a condiciones atmosféricas aeróbicas, en presencia de oxígeno, se forman radicales nitro a partir de la reducción del metronidazol, también se generan aniones superóxido que son tóxicos para el microorganismo.
Se ha estudiado que POR es una enzima esencial en Helicobacter pylori, lo que implica que la inactivación de esta enzima es letal y la bacteria no sobrevive a través de la activación de vías metabólicas compensatorias. En H. pylori, in vivo el aceptor de electrones de esta enzima probablemente es una flavodoxina endógena y es reducida por POR, además, la actividad del 2-oxoglutarato oxidorreductasa, fue detecta en la bacteria, catalizando el análogo, reversible a la descarboxilación oxidativa de 2- oxoglutarato para la formación de succinil-CoA, uno de los principales intermediarios del ciclo del ácido tricarboxílico, la presencia de este 2-oxoácido oxidorreductasa es importante, porque, a diferencia de otras enzimas oxoácidas deshidrogenasas son muy lábiles al oxígeno, lo que en parte, soporta su naturaleza microaerofílica, POR esta implicado en la reducción de metronidazol (Hughes et al., 1997).
La generación de reductores con bajo potencial rédox, como ferredoxina o flavodoxina, por POR ó OOR, pueden ser utilizados para participar en diversos procesos, incluyendo la obtención de hidrógeno, en la descarboxilación el piruvato es unido por la piruvato ferredoxina oxidorreductasa a la coenzima A (CoA) para generar acetil-CoA, ferredoxina reducida y CO2, cuando no se generan reacciones de reducción, la ferredoxina es re-oxidada
y el hidrogeno es liberado a la célula, también son utilizados en procesos como la fijación del nitrógeno y el suministro de NADPH para la respiración en el transporte de electrones (Hughes et al., 1997).
37
El gen PorD codifica la enzima Piruvato/Ferredoxina oxidorreductasa como se mencionó anteriormente, cataliza la descarboxilación oxidativa del piruvato para generar acetil-CoA. Por su parte, el gen oorD codifica 2-oxoglutarato oxidorreductasa y participa en la descarboxilación oxidativa del 2-oxoglutarato en presencia de CoA para generar succinil-CoA y un aumento del CO2 (Hughes et al., 1997).
2.6.1.3. Gen fldA, fdxA y fdxB
Los genes fldA, fdxA y fdxB codifican las enzimas flavodoxina, ferredoxina y las proteínas ligadas a ferredoxina, respectivamente, que actúan como aceptor de electrones del complejo enzimático de la piruvato oxireductasa, que cataliza la descarboxilación oxidativa del piruvato.
Se ha propuesto que en anaerobios, la reducción del grupo nitro de nitroimidazoles es mediada por un complejo oxidorreductasa piruvato ferredoxina (flavodoxina), el piruvato puede ser transformado por la generación de electrones para la oxidación de piruvato como se muestra en la siguiente reacción:
Piruvato + CoA Acetil-CoA + CO2 + 2 e-
[image:42.612.147.489.472.627.2]Los electrones liberados en esta reacción son transferidos primero a la ferredoxina o flavodoxina (figura 6), para que ellos puedan ser capturados por el metronidazol, los productos formados reacción con el DNA, causando rompimiento de sus cadenas y la muerte celular (Kaihovaara et al., 1998).
Figura 6. Metabolismo del piruvato y activación reductiva del metronidazol por H. pylori
38
Kaihovaara et al., purificó la flavodoxina y confirmó que la flavoproteína actúa como un aceptor de electrones en la oxidación de piruvato, cambios espectrales de la proteína oxidada, por 300 nm a 700 nm, fueron medidos en la presencia de piruvato, la flavodoxina fue reducida a su forma neutral de flavina semiquinona por los electrones generados en la oxidación del piruvato, estos resultados indicaron la actividad piruvato flavodoxina oxidorreductasa esta presente en H. pylori, esta actividad en extractos bacterianos fueron adicionalmente detectados con la reducción de citocromo c en presencia de oxígeno, la flavodoxina fue esencial para la oxidación de piruvato, catalizando la transferencia de electrones por la piruvato flavodoxina oxidorreductasa a citocromo c (Kaihovaara et al.,
1998).
El principal producto final metabólico observado en la oxidación por H. pylori fue el acetato. La alta producción de acetato resulta por la aceleración de la actividad enzimática, de piruvato flavodoxina oxidorreductasa debido a la rápida reoxidación de la misma enzima o por la flavodoxina por oxígeno molecular (Kaihovaara et al., 1998).
Como se describió anteriormente, en la oxidación del piruvato, la producción de formato no se conoce detalladamente, pero es posible que en las cepas de H. pylori resistentes al metronidazol, la actividad de la piruvato flavodoxina oxidorrecutasa es reemplazada por la piruvato-formato liasa (Hoffman et al.,1996), en este caso se suministra el electrón a flavodoxina y la generación de CO2 puede ser reemplazada por el incremento de formato y de piruvato puede ser imparcial por flavodoxina y la concomitante reducción del metronidazol (Kaihovaara et al., 1998).
2.7. TECNICAS DE BIOLOGIA MOLECULAR PARA LA IDENTIFICACIÓN DE LA RESISTENCIA AL METRONIDAZOL EN HELICOBACTER PYLORI
Para el estudio de la resistencia de Helicobacter pylori al metronidazol se emplean técnicas de biología molecular, las mas utilizadas para la identificación de dicha resistencia son la reacción en cadena de la polimerasa (PCR), la reacción en cadena de la polimerasa de transcriptasa inversa (PCR-RT) y el DNA polimórfico amplificado al azar (RADP).
2.7.1. Reacción en cadena de la polimerasa (PCR)
39
muestras resultantes de la extracción y aislamiento de DNA, entre otras (Luque J. et al.2001). La amplificación del DNA se logra mediante la extensión de cebadores que son
complementarios a secuencias situadas hacia los dos extremos 3’ del segmento bicatenario
[image:44.612.116.527.165.382.2]que se pretende amplificar (Puerta et al., 2005).
Figura 7. Reacción en Cadena de la polimerasa (Enciclopedia Británica)
Este proceso consta de tres etapas:
1. Desnaturalización, el calentamiento del DNA para la separación de las dos hebras, se realiza mediante una incubación breve con una duración aproximadamente de 30 a 120 segundos, a una temperatura entre 68ºC y 97ºC, que debe ser superior a la temperatura de fusión de la región del DNA que se desea amplificar, como de cierto modo la desnaturalización es similar a la fusión de un sólido, se realizan curvas de fusión y a la temperatura central del intervalo de transición (50% de desnaturalización) se le llama temperatura de fusión Tm (Luque et al., 2001).
2. Hibridación ó anillamiento, enfriamento rápido por debajo de la Tm lo que permite la
hibridación de las hebras sencillas del DNA de los cebadores. Por lo general, se usan temperaturas de 37ºC a 65ºC que se mantienen entre 10 y 20 segundos (Luque et al., 2001).