FACULTAD DE CIENCIAS BIOLÓGICAS
Escuela de Formación Profesional de
Ciencias Biológicas
UNIVERSIDAD NACIONAL DE LA AMAZONÍA PERUANA
“ACTIVIDAD ANTIBACTERIANA DE EXTRACTOS FÚNGICOS
DE Ganoderma applanatum SOBRE CEPAS HOSPITALARIAS
DE Staphylococcus aureus y Pseudomonas aeruginosa
AISLADAS DEL HARDWARE DE COMPUTADORAS DEL
HOSPITAL CESAR GARAYAR – IQUITOS”
TESIS
Requisito para optar el título profesional de:
BIÓLOGO
AUTORAS:
BETSABETH TRINIDAD GUZMAN
EMPERATRIZ GUADALUPE MORALES AMARINGO
ii JURADO CALIFICADOR Y DICTAMINADOR
--- Blga. Julia Bardales García, MSc.
Miembro
---
Blga. Teresa de Jesús Del Águila Mori, MSc. Presidente
--- Blgo. Freddy Orlando Espinoza Campos, Mgr.
iii Asesores
--- Mblgo. Álvaro Benjamín Tresierra Ayala, Dr.
Asesor
--- Blga. María Elena Bendayán Acosta, MSc.
Asesora
--- Ing. Wilfredo Ruíz Mesía, Dr.
v DEDICATORIA
A mis padres por su gran apoyo incondicional y a mis hermanos por motivarme a ser mejor cada día.
Betsabeth Trinidad G.
A mi mamá, mis abuelos por su apoyo incondicional y a mis dos grandes amores por ser la inspiración y motivación de mi día a día para realizar este proyecto.
vi AGRADECIMIENTOS
Queremos expresar nuestro más grande agradecimiento a todas las personas que
hicieron posible la realización del proyecto.
Al Centro de Investigaciones de Recursos Naturales (CIRNA - UNAP), por permitirnos la
realización de esta tesis en su establecimiento.
A nuestros asesores: al Dr. Álvaro B. Tresierra Ayala, a la. Blga. María Elena Bendayan y
al Ing. Wilfredo Ruíz Mesia, por trasmitirnos sus valiosos conocimientos, por su paciencia
y su apoyo incondicional en todo este tiempo.
Al Laboratorio de Investigación de Productos Naturales Antiparasitarios de la Amazonía
(LIPNAA), en especial a las Ings. Lastenia Ruíz Mesia y Leonor Arévalo, así como
también al Sr. Cesar Tuesta, por darnos las facilidades y su apoyo. Y al mismo tiempo al
Laboratorio de Biotecnología por permitirnos y darnos las facilidades de usar sus equipos.
A los Biólogos Ronald Medina, Ricardo Abadie, Manuela Ruíz y Gavy Paredes por sus
apoyo y sus enseñanzas.
A los bachilleres José Luna V. y Aldo Alva V. por su apoyo en la salida de campo y
enseñanza estadística.
A mi jurado de tesis, por las observaciones, la evaluación de la presente investigación y
vii
3.2. Características de las bacterias en estudio --- 9
3.2.1. Género Pseudomonas --- 9
3.2.2. Género Staphylococcus --- 14
IV. MATERIALES Y MÉTODOS --- 20
4.1. Descripción del área de estudio --- 20
4.1.1. Lugar de recolección de muestras --- 20
4.1.2. Lugar de procesamiento de las muestras --- 21
4.2. Cepas bacterianas --- 21
4.3. Procedimiento --- 21
4.3.1. Recolección del hongo --- 21
viii
4.3.3. Ensayos Químicos --- 23
4.3.4. Evaluación de actividad antibacteriana por el método de difusión en agar ---23
4.3.5. Determinación de la concentración inhibidora mínima y la concentración bactericida mínima de los extractos fúngicos que muestren actividad antibacteriana 26 V. RESULTADOS --- 30
5.1. Sensibilidad de las cepas de P. aeruginosa y S. aureus frente a los extractos, mediante difusión en disco --- 30
5.2. Concentración inhibitoria mínima (CIM) y concentración bactericida mínima (CBM) de los extractos frente a las cepas de S. aureus --- 33
5.2.1. Concentración inhibitoria mínima de los extractos etanólicos frente a las cepas de S. aureus --- 33
5.2.2. Concentración Bactericida Mínima de los extractos etanólicos frente a las cepas de S. aureus --- 34
VI. DISCUSIÓN --- 35
VII. CONCLUSIONES --- 38
VIII. RECOMENDACIONES --- 39
IX. REFERENCIAS BIBLIOGRÁFICAS --- 40
ix LISTA DE CUADROS
Cuadro 1. Clasificación de las Pseudomonas según la homología de ARNr/ADN - 10
Cuadro 2. Coordenadas UTM del área de estudio --- 20
Cuadro 3. Actividad antibacteriana de los extracto de G. applanatum frente a las
cepas de S. aureus --- 30
Cuadro 4. Datos descriptivos de los halos de inhibición de los extractos con
actividad frente a S. aureus --- 31
Cuadro 5. Concentración inhibitoria y bactericida mínima de los extractos frente a
S. aureus --- 33
Cuadro 6. Concentración Inhibitoria Mínima de los extractos etanólicos al 31,25
mg/ml frente a S. aureus--- 33
x LISTA DE FIGURAS
Figura 1. Área de estudio --- 20
Figura 2. Lugar de procesamiento de las muestras --- 21
Figura 3. Especie Ganoderma applanatum --- 22
Figura 4. Ajuste y aplicación del inóculo --- 25
Figura 5. Concentración Inhibitoria Mínima (CIM) --- 28
Figura 6. Concentración Bactericida Mínima (CBM) --- 29
Figura 7. Halo de inhibicion frenta a las cepas de P. aeruginosa y S. aureus --- 30
Figura 8. Actividad antibacteriana de los extractos de G. applanatum frente a las cepas de S. aureus --- 31
xi LISTA DE ANEXOS
Anexo 1. Obtención del extracto etanólico --- 46
Anexo 2. Elaboración e impregnación de los discos con los extractos --- 46
Anexo 3. Sembrando las cepas bacterianas en medio TSB para la incubación ---- 47
Anexo 4. Inoculación de las cepas en agar MH para la prueba de sensibilidad ---- 47
Anexo 5. Colocación de los discos impregnados con los extractos en agar MH ---- 48
Anexo 6. Antibiograma de los extractos fúngicos frente a P. aeruginosa --- 48
Anexo 7. Antibiograma de los extractos fúngicos frente a S. aureus --- 49
Anexo 8. Determinación de la Concentración Inhibitoria Mínima (CIM) del extracto
fúngico (C) frente a S. aureus --- 49
Anexo 9. Determinación de la Concentración Inhibitoria Mínima (CIM) del extracto
fúngico (Ca) frente a S. aureus --- 50
Anexo 10. Siembra del inóculo proveniente de un tubo sin desarrollo bacteriano
en agar TSA para determinar Concentración Bactericida Mínima --- 50
Anexo 11. Determinación de la Concentración Bactericida Mínima (CBM) de los
xii RESUMEN
Los hongos del género Ganoderma se caracterizan por poseer metabolitos
secundarios que tienen propiedades antibacterianas, asimismo, la especie G.
applanatum presenta ciertos compuestos químicos que brindan beneficios a la
salud humana. Por esta razón, el presente trabajo se realizó con la finalidad de
determinar la existencia de actividad antibacteriana de los extractos fúngicos de
Ganoderma applanatum sobre cepas de Staphylococcus aureus y Pseudomonas
aeruginosa, aisladas del hardware de computadoras del Hospital Cesar Garayar -
Iquitos. Los hongos recolectados en la Reserva Nacional Allpahuayo Mishana (Km
28), fueron secados para obtener el extracto fúngico etanólico, luego, se evaluó la
actividad antibacteriana por el método de difusión en agar, y aquellos que
resultaron positivos, se determinó los valores de la concentración inhibidora mínima
y concentración bactericida mínima de los extractos fúngicos. Al evaluar la actividad
antibacteriana del extracto fúngico se observó que las cepas de S. aureus eran
sensibles al extracto fúngico. El CIM del extracto fúngico mostró actividad a
concentración de 31,25 mg/ml frente la cepa S. aureus, sin embargo, en el CBM no
se obtuvo resultados positivos. Se concluye que el extracto fúngico etanólico
presentó actividad inhibitoria frente a S. aureus pero no actividad bactericida; a
diferencia de P. aeruginosa que no mostro ninguna actividad.
Palabras claves: Actividad antibacteriana, extractos fúngicos, cepas
I. INTRODUCCIÓN
Desde hace milenios, los hongos han sido empleados por el hombre tanto para
la alimentación, como para el tratamiento de diferentes enfermedades, siendo
los países del lejano oriente, como Japón, China y Corea, los consumidores
más habituales (Smith, 2002), debido a la comprobación de las diversas
actividades biológicas exhibidas por sus metabolitos secundarios de gran
importancia por su amplio beneficio para la salud, éstos han sido aislados del
micelio y cuerpo fructífero de algunas especies de Ganoderma (Mattila, 2000).
Algunos son utilizados en medicina tradicional (Mata, 2003) y otros son empleados
en la producción de medicamentos (Smith, 2002); sin embargo, el número de
especies de macromicetos investigados es relativamente bajo en comparación al
conocimiento que se tiene de los micromicetos (Barros et al., 2007).
Los hongos del género Ganoderma, son considerados como importantes
organismos medicinales. A través de los años, el cuerpo fructífero de estos hongos
ha sido de gran utilidad en el mundo de la medicina tradicional y, el interés en su
estudio se viene incrementando debido, fundamentalmente, a que se han
encontrado metabolitos secundarios que podrían potencialmente ser utilizados en la
industria farmacológica para el tratamiento de diferentes enfermedades y
aplicaciones en el sector agroindustrial. Durante las dos últimas décadas, en estos
hongos se han identificado un buen número de compuestos caracterizados por
presentar diferentes tipos de actividad biológica, destacándose sustancias con
actividad cardiovascular, citotóxica, inmunomoduladora, analgésica, antidiabética,
antioxidante, insecticida y nematicida, entre otras (Zjawiony, 2004). Por otro lado,
son incapaces de sintetizar macromoléculas a partir del dióxido de carbono y la
energía procedente de la luz por el hecho de no poseer clorofila; debido a esto, su
2 conforman el sustrato donde se desarrollan, además de las condiciones climáticas
que los rodean, haciendo que los hongos estén presentes en diferentes regiones o
que crezcan en sustratos que varíen en su composición (García-Pajón y Collado,
2003). De igual forma, se ha documentado que Ganoderma applanatum biosintetiza
sustancias que generan importantes beneficios para la salud de animales y
humanos, por tener efectos hepatoprotectivos, antioxidantes, de protección
cardiovascular y antibacterianos, entre otros (Russell, 2006).
La especie Ganoderma applanatum crece en diferentes regiones del mundo, y se
han registrado diversos metabolitos secundarios de acuerdo al tipo de sustrato
donde se desarrolla y según las condiciones climáticas de los ecosistemas. La
Amazonía peruana alberga una amplia biodiversidad fúngica, debido a que su
biogénesis puede variar de acuerdo al lugar de donde procede y sintetizar nuevos
metabolitos secundarios, que presenten actividad antibacteriana, esto conllevó a la
realización del presente estudio, siendo así, un trabajo pionero en esta región.
Además, la realización de este estudio contribuiría a incrementar el conocimiento
generado en otras latitudes del planeta respecto a la actividad antimicrobiana de
esta especie de macromiceto y de esta forma incentivar al estudio de estos
organismos que posibilitarían generar una alternativa de solución para mejorar la
salud humana mediante el hallazgo de sustancias con capacidad antimicrobiana.
Teniendo en cuenta que los bosques amazónicos peruanos se caracterizan por
poseer una gran diversidad de hábitats, los que brindan las condiciones necesarias
para el crecimiento y desarrollo de diversos organismos (Mori et al., 2011), como
los hongos y entre ellos, la especie Ganoderma applanatum y que a pesar de sus
cualidades benéficas, es un organismo que en esta región ha sido muy poco
estudiado, el presente trabajo pretende determinar la actividad antibacteriana de los
3
aureus y Pseudomonas aeruginosa, aisladas del hardware de computadoras del
4 II. OBJETIVOS
2.1. General
Determinar la actividad antibacteriana de los extractos fúngicos de Ganoderma
applanatum sobre cepas de Staphylococcus aureus y Pseudomonas aeruginosa,
aisladas del hardware de computadoras del Hospital Cesar Garayar - Iquitos.
2.2. Específicos
Obtener el extracto etanólico de Ganoderma applanatum.
Evaluar in vitro, la susceptibilidad de las cepas bacterianas aisladas de
hardware de computadoras del Hospital César Garayar – Iquitos, frente al
extracto etanólico fúngico.
Determinar la concentración inhibitoria mínima y la concentración bactericida
5 III. REVISIÓN DE LITERATURA
3.1. Generalidades
Los hongos son organismos que se encuentran en todos los biomas y sobre los
más variados sustratos, registrándose así, la existencia de los hongos en áreas
desérticas y tropicales, en zonas templadas e inclusive en ambientes acuáticos
(Tello, 2010).
Los basidiomicetos producen una amplia gama de productos naturales que
abarca desde componentes estructurales con actividad antitumoral e
inmunológicamente activos hasta agentes antimicrobianos, antifúngicos,
antivirales, citostáticos, enzimas reguladores de crecimiento y aromas (Brizuela
et al., 1998).
3.1.1. Familia Ganodermataceae
Esta familia ha sido estudiada recientemente, cuyo interés se centra en el
aislamiento y caracterización de sus compuestos bioactivos, entre ellos los
polisacáridos, especialmente glucanos (Leskosek et al., 2010). Estos
polisacáridos poseen un grupo largo y diverso de sustancias que juegan un
importante rol en la estructura y función de la pared celular de los fúngicos.
Siendo así una fuente importante de polisacáridos (Osinska et al., 2014).
3.1.1.1. Género Ganoderma
El género Ganoderma P. Karst. Pertenece a la familia Ganodermataceae
del orden Polyporales (Basidiomycetes, Fungi) (Ryvarden, 1998). Donk,
6 los esporos truncados y ornamentados (Moncalvo y Ryvarden 1997). Es
uno de los géneros más amplios de los Polyporales y fue descrito por
Karsten (1881) basado en Polyporus lucidus de Europa, llamado también
Ganoderma lucidum.
Las especies de Ganoderma crecen sobre madera en descomposición
absorbiendo nutrientes a través de pequeños tubos conocidos como hifas y
desarrollan el cuerpo fructífero sobre troncos. Estas especies pueden
crecer bajo condiciones elevadas de calor y humedad y son usualmente
encontradas en regiones tropicales y subtropicales (Montalvan y Ryvarden,
1998). Poseen basidiocarpo perenne, sésil, leñoso, solitario, de aplanado a
forma de concha, en forma de repisa semicircular, superficie seca,
costrosa y ligeramente agrietada que crece en troncos de árboles muertos,
en diversos tipos de vegetación como selva alta perennifolia, selva
mediana subperennifolia, humedales, pastizales y en zonas urbanas, casi
todo el año (Ramos y Cappello, 1998).
Productos farmacéuticos importantes con aplicaciones médicas han sido
extraídos de diferentes hongos como G. lucidum y G. tsugae los cuales
están relacionados con un largo historial de uso en la medicina tradicional
que data hace más de cuatro milenios (Mizuno et al, 1995).
Se han aislado y purificado compuestos bioactivos del hongo Ganoderma
tales como polisacáridos, triterpenos, esteroides, lectinas y proteínas,
siendo mayormente investigado los dos primeros. Algunos de estos inhiben
el crecimiento de células cancerígenas in vitro, otros tienen actividad
antiviral in vitro o presentan otra actividad in vivo relacionada con aspecto
7 el desarrollo de agentes terapéuticos químicos en el tratamiento contra el
cáncer u otras enfermedades (Mizuno et al., 1995), como hipertensión,
bronquitis crónica, diabetes y arterioesclerosis y como un tónico sedativo
(Shiao, 2003).
Especie : Ganoderma applanatum (Pers. Ex Wallr.)
Descripción
El basidiocarpo de G. applanatum presenta píleo en forma de repisa,
semicircular de 6-20 cm de diámetro de color marrón a marrón oscuro, de
consistencia dura, superficie lisa y áspera por los bordes semicirculares en
tonalidades marrones a oscuras, al envejer la cutícula se despega del
contexto, sin olor y sabor especial, contexto de 1-4 cm de grosor, de color
castaño a marrón, se disponen en capas de 2–5 mm de ancho con tubos
de color marrón a castaño oscuro. En el himenio presenta poros redondos
de 5 – 6 poros por mm, de color blanco a cremas oscuras, al tacto o al
8 semilateral y al principio los primordios son negros en forma de dedos
(Espinoza, 2004). G. applanatum es una especie que presenta poca
información que está disponible sobre la producción de metabolitos
secundarios por estos hongos (Tawfik, 2011).
Hábitat y distribución geográfica
Es comúnmente encontrado en crecimiento sobre árboles muertos en
regiones templadas y tropicales del mundo. Es un macrofúngico perenne
con un cuerpo fructífero duro que está desprovisto de tallo (Volk, 1998).
Fructifica sobre la madera de árboles de hoja caduca, sobre todo robles y
hayas, pudiendo comportarse de forma parasitaria o saprofita. Como se
trata de carpóforos perennes, es obvio que se le puede encontrar en
cualquier época del año, siendo una especie frecuente en nuestros
bosques (Jung, 2011).
Su distribución geográfica Perú, España, México, es Cosmopolita
(Espinoza, 2004).
Estudios anteriores
Diferentes estudios se han llevado a cabo sobre la actividad antibacteriana
de esta especie de hongo tal como lo mencionan Guzmán et al. (2013),
quienes evaluaron la actividad bactericida de G. applanatum mediante el
método de difusión en agar, usando el extracto etanólico del hongo,
empleando concentraciones (200, 150 y 100 mg/ml), encontrándose
resultados positivos a 150 mg/ml frente a la bacteria Staphylococcus
9 ninguna de las concentraciones. Asimismo, estudios realizado por Nagaraj
et al. (2013), quienes evaluaron el potencial antibacteriano in vitro del
extracto metanólico de G. applanatum por el método de difusión en agar a
concentraciones de 70 mg/ml, 50 mg/ml y 25 mg/ml, encontraron que este
hongo tienen actividad antibacteriana significativa contra S. aureus (15mm)
y E. coli (13mm) y un efecto moderado contra Bacillus subtilis (12mm) y P.
aeruginosa (12mm) y efecto leve sobre el crecimiento de Klebsiella
pneumonia (8mm) y Salmonella typhi (8mm).
Otro estudio realizado por Zuluaga et al. (2007), quienes evaluaron la
actividad antimicrobiana del extracto etanólico de G. lucidum contra la
bacteria Staphylococcus aureus y Escherichia. coli, empleando el método
de dilución en caldo, dando resultados negativos a todas las
concentraciones estudiadas.
En un estudio realizado por Albino et al. (2007) sobre Ganoderma australe
en el que utilizaron ácido austrálico y metil austrálico, mediante el método
de microdilución demostraron que fueron activos contra hongos y bacterias
Gram (+) Bacillus cereus y S. aureus.
3.2. Características de las bacterias en estudio
3.2.1. Género Pseudomonas
Las bacterias del género Pseudomonas son bacilos gram negativos, móviles y
aerobios, alguno de los cuales producen pigmentos hidrosolubles (Jawetz et
al., 2012). Los miembros de este género son ubicuos y se encuentran en el
10 el agua. Se encuentran también en todo el ambiente hospitalario, en
reservorios húmedos como los alimentos, las flores cortadas, los lavatorios,
los baños, los equipos de diálisis y terapia respiratoria e incluso en las
soluciones desinfectantes. Es raro que las personas sean portadoras dentro
de la flora microbiana normal, salvo en pacientes hospitalizados y en
anfitriones inmunodeprimidos ambulatorios (Murray, 2010).
Este género presenta una sencilla exigencia para crecer y versatilidad
nutricional, debido a esto, tiene un amplio entorno de distribución. Puede
emplear muchos compuestos orgánicos como fuente de carbono y nitrógeno,
así como algunas cepas consiguen incluso crecer en agua destilada
empleando solo oligonutrientes. Además estos organismos poseen factores
estructurales, toxinas y enzimas que potencian su virulencia y los hace
resistente a la mayor parte de los antibióticos de uso habitual. (Murray, 2010).
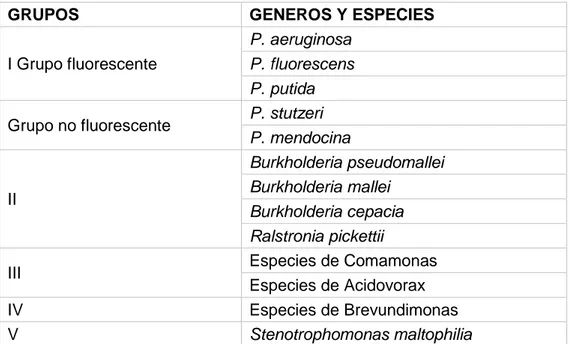
La clasificación de las Pseudomonas se basa en la homología de ARNr/ADN
y en las características de cultivo comunes.
Cuadro 1. Clasificación de las Pseudomonas según la homología de ARNr/ADN
GRUPOS GENEROS Y ESPECIES
I Grupo fluorescente
P. aeruginosa
P. fluorescens P. putida
Grupo no fluorescente P. stutzeri
11
3.2.1.1. Pseudomonas aeruginosa
P. aeruginosa tiene una amplia distribución en la naturaleza y suele estar
presente en medios húmedos en los hospitales. Puede colonizar al ser
humano normal, es un saprofito, causando enfermedad en personas con
defensas anormales. (Brooks et al., 2012). Constituye uno de los
microorganismos más importantes y problemáticos en las bacteriemias por
gramnegativos y se presenta con mayor frecuencia en pacientes
inmunodeprimidos con largos periodos de hospitalización, sometidos a
diversas manipulaciones, con antecedentes de infecciones graves y uso
previo de antibióticos de amplio espectro (Gómez et al., 2002). Es el
patógeno más importante dentro del género Pseudomonas, teniendo en
cuenta la cantidad y tipos de infecciones (invasivas y toxígenas) que
produce, así como la morbilidad y mortalidad que ocasiona. A partir de la
década del 60 se ha incrementado el interés médico por esta especie, al
convertirse en uno de los principales agentes causantes de enfermedades
adquiridas en el ámbito hospitalario, especialmente en pacientes
inmunocomprometidos. (Llop et al., 2001).
a. Morfología e Identificación
P. aeruginosa es móvil, tiene forma de bastón, mide casi 0.6 x 2 µm. Es un
gramnegativo y muestra una disposición en bacterias individuales, en
pares y a veces en cadenas cortas (Brooks et al., 2012).
b. Cultivo
P. aeruginosa es un aerobio obligado que se multiplica fácilmente en
12 dulzón, o de uvas. Algunas cepas producen hemólisis. Producen
pigmentos de fenazina en agar nutritivo, después de 24 horas de
incubación a 37°C y posteriormente a temperatura ambiente; los mismos
pueden ser azul (piocianina), amarillo verdoso (pioverdina), rojo
(piorrubina) y negro (piomelanina). Existe, aproximadamente, un 10% de
Pseudomonas aeruginosa que son apigmentadas (Llop et al., 2001).
P. aeruginosa en un cultivo puede producir múltiples tipos de colonias que
también tiene diferentes actividades bioquímicas y enzimáticas y diferentes
tipos de susceptibilidad antimicrobiana. A veces no está claro si los tipos
de colonia representan diferentes cepas de P. aeruginosa o si son
variantes de la misma cepa (Brooks et al., 2012).
c. Características de crecimiento
Se multiplica bien a una temperatura de 37 a 42 °C, su crecimiento a 42°C
ayuda a distinguirla de otras especies de Pseudomonas en el grupo
fluorescente. Oxida la glucosa pero no tiene acción sobre otros
carbohidratos. Son Oxidasa positivo, pero son Indol negativo (Brooks et al.,
2012).
d. Estructura antígena y toxinas
Las fimbrias se proyectan desde la superficie de la célula y favorecen la
adherencia a las células epiteliales del hospedador. El exopolisacáridos es
la causa de las colonias mucoides que se observan en los cultivos de
pacientes con fibrosis quística. El lipopolisacáridos, que existe en múltiples
13 microorganismo (Brooks et al., 2012). Presenta antígenos “O” del
lipopolisacárido y el antígeno “H” proporcionan varios serotipos. La
endotoxina del LPS actúa en igual forma que la de las enterobacterias.
Produce enzimas extracelulares como proteasas, elastasas y dos
hemolisinas: una fosfolipasa C termolábil y un glucopéptido termoestable
(Ramírez, 2003).
Muchas cepas de P. aeruginosa producen exotoxina A, esta toxina altera la
síntesis de proteínas al inhibir la elongación de la cadena peptídica en las
células eucariotas. Esta toxina probablemente participa en la
dermatonecrosis que tiene lugar en las quemaduras, el daño corneal en las
infecciones oculares y el daño tisular en las infecciones pulmonares
crónicas (Murray, 2010).
e. Patogenia
Pseudomonas aeruginosa, ataca principalmente la piel y las mucosas,
especialmente cuando se ha sufrido algún traumatismo como quemaduras,
infecta catéteres intravenosos y sondas urinarias; además, ataca a
pacientes inmunosuprimidos por drogas (quimioterapia del cáncer) o por
otras causas que se acompañan de neutropenia. Las meninges se infectan
cuando la toma de muestra del LCR se introduce al microorganismo con la
aguja. En el tejido infectado (piel o mucosas traumatizadas, etc.), la
bacteria coloniza y de allí se disemina por vía hematógena produciendo
sepsis con las manifestaciones propias de la endotoxina (fiebre, oliguria,
leucopenia y luego leucocitosis, CID, insuficiencia respiratoria, hipotensión,
choque y muerte). Las toxinas y enzimas excretadas por la P. aeruginosa
14 f. Resistencia a antibióticos
P. aeruginosa posee una resistencia inherente a muchos antibióticos y
puede mutar a cepas aún más resistentes durante el tratamiento. Aunque
se han identificado numerosos mecanismos de resistencia, la mutación de
las porinas constituye el principal mecanismo de resistencia. La alteración
de las proteínas que configuran la pared de estos poros con el fin de
restringir el flujo al interior de la célula conlleva a la aparición de resistencia
a numerosos grupos de antibióticos de manera simultánea. P. aeruginosa
sintetiza diferentes lactamasas que inactivan diversos antibióticos
β-lactámicos (Murray, 2010).
3.2.2. Género Staphylococcus
Las células del género Staphylococcus se desarrollan en un patrón que
recuerda a un racimo de uva, sin embargo, los microorganismos presentes en
muestras clínicas aparecen como células aisladas, en pares o en cadenas
cortas. La mayor parte de los estafilococcus tiene un diámetro de entre 0,5 y 1
µm y son anaerobios facultativos inmóviles capaces de crecer en un medio
con una elevada concentración de sal y a temperaturas de 18 a 40° C.
Actualmente, el género comprende 40 especies y 24 subespecies, muchas de
las cuales se encuentran en el ser humano. Los estafilococos conforman un
importante grupo de patógenos en el ser humano y originan un amplio
espectro de enfermedades sistémicas que pueden poner en peligro la vida,
infecciones de piel, los tejidos blandos, los huesos y el aparato genitourinario
15 Los estafilococos patógenos producen hemólisis de la sangre, coagulan el
plasma y producen enzimas y toxinas extracelulares. Un tipo común de
envenenamiento alimentario es el producido por una enterotoxina
estafilocóccica termoestable. Los estafilococos desarrollan con rapidez
resistencia a muchos agentes antimicrobianos, por lo que resulta difícil su
tratamiento. Las tres especies principales de importancia clínica son: S.
aureus, S. epidermidis y S. saprophyticus (Brooks et al., 2012).
3.2.2.1. Staphylococcus aureus
a. Morfología e Identificación
Staphylococcus aureus son grampositivas y agrupadas en racimos. En
cultivos líquidos se encuentran cocos aislados, en pares, tétradas y
cadenas cortas. Los cocos jóvenes se tiñen inmensamente con coloración
de Gram; al envejecer muchas células se vuelven gramnegativas.
b. Cultivo
Prosperan fácilmente en la mayoría de los medios bacteriológicos bajo
condiciones aerobias o microaerófilas a temperatura de 37°C, formando
mejor el pigmento a temperatura ambiente (20 a 25°C). Las colonias
desarrolladas en medios sólidos (agar-sangre), en un periodo de 18 a 24
horas tienen un diámetro de 1 a 3 mm y de 3 a 10 mm en incubación
prolongada durante 5 días; las mismas son redondas, lisas, elevadas y
resplandecientes. S. aureus suele formas colonias de color gris a amarillos
16 c. Características del crecimiento
Produce catalasa, fermenta lentamente muchos carbohidratos y produce
ácido láctico pero no gas. La actividad proteolítica varía mucho de una a
otra. Son relativamente resistentes a la desecación, al calor y al cloruro de
sodio al 9% pero inhibidas por determinadas sustancias químicas, como el
hexaclorofeno al 3%.
La resistencia que presenta a los antimicrobianos puede deberse
frecuentemente a la producción de β-lactamasa, que está sujeta a
plásmidos, haciendo que los microorganismos sean resistentes a muchas
penicilinas, estos plásmidos son transmitidos mediante transducción o
conjugación. La resistencia a la meticilina es independiente de la
producción de β-lactamasa, esta resistencia es codificada y regulada por
una serie de genes que se encuentra en una región del cromosoma.
d. Estructura antigénica
Contiene polisacáridos y proteínas antigénicas así como otras sustancias
importantes en la estructura de la pared celular. El peptidoglucano, un
polímero de polisacárido que contiene subunidades ligadas, proporciona el
exoesqueleto rígido de la pared celular, esto es destruido por acido potente
o por la exposición a lisozimas. Es importante en la patogenia de las
infecciones, el desencadenamiento de la producción de interleucina I y
17 e. Patogenia
La capacidad patógena de S. aureus está dada por mecanismos y factores
patogénicos que incluyen la presencia de cápsula, la cual inhibe la
fagocitosis (algunas cepas de S. aureus). La producción de enzimas y
toxinas permite su establecimiento, multiplicación y propagación a otros
órganos y sistemas, así como la colonización de sitios específicos del
hospedero y la elaboración de slime, el cual es un glicoconjugado
extracelular producido por S. aureus y ciertas cepas de estafilococos
coagulasa negativa, que desempeña en estos últimos un importante papel
en la ocurrencia de enfermedades (Llop et al., 2001).
S. aureus produce un gran número de factores de virulencia, entre los que
figuran alfa, beta, delta, gama y leucocidina de Panton-Valentine, toxinas
exfoliativas (A y B), enterotoxinas (A a E, G a I) y la toxina-1 del síndrome
del shock tóxicos (TSST-1). Las toxinas citolíticas conocidas como
hemolisinas, pueden provocar la lisis de los neutrófilos, lo que da lugar a la
liberación de las enzimas lisosomales que posteriormente dañan los tejidos
circundantes. Una citotoxina, la leucocidina de P-V, se ha relacionado con
infecciones cutáneas graves (Murray et al., 2007).
S. aureus causa enfermedad mediante la producción de toxina o a través
de la invasión directa y la destrucción del tejido. Las manifestaciones
clínicas de algunas enfermedades estafilocócicas se deben casi
exclusivamente a la actividad de la toxina (SPEE, intoxicación alimentaria
estafilocócica y SST), mientras que otras afecciones son consecuencia de
la proliferación de los microorganismos, la cual da lugar a la formación de
18 neumonía, etc.). La producción de enfermedad en presencia de un cuerpo
extraño requiere un número significativamente menor de estafilococos
(Murray et al., 2007).
f. Resistencia a antibiótico
Los estafilococos desarrollaron una rápida resistencia a los antibióticos
después de la introducción de la penicilina, en la actualidad una proporción
inferior al 10% de las cepas son sensibles a este antibiótico. Esta
resistencia está mediada por la enzima penicilinasa, la cual hidroliza el
anillo β - lactámico de la penicilina. La información genética que codifica la
producción de esta enzima se encuentra en un plásmido transmisible, lo
que facilita la rápida diseminación de resistencias entre los estafilococos.
Actualmente, una proporción comprendida entre el 30% y el 50% de las
cepas de S. aureus y más del 50% de los estafilococos
coagulasa-negativos son resistentes a las penicilinas semisintéticas (ej. meticilina,
nafcilina, oxacilina, dicloxacilina) (Murray et al., 2007).
Hasta hace poco tiempo, las cepas MRSA se restringían únicamente a los
entornos hospitalarios. No obstante, a lo largo de los últimos años se han
descrito algunos brotes comunitarios por MRSA. A pesar de la diversidad
geográfica de los brotes, las cepas parecen presentar una relación clonal y
difieren de las cepas de MRSA aisladas en los hospitales. Casi todas estas
cepas causantes de brotes comunitarios portan también la toxina
leucocidina P-V y se han relacionado con enfermedades graves, como la
19 Los estafilococos han demostrado gran capacidad para desarrollar
resistencia a la mayoría de los antibióticos, hasta la vancomicina, el
antibiótico de selección en la actualidad como tratamiento de los
estafilococos resistentes a meticilina, cepas de S. aureus han creado dos
mecanismos de resistencia a vancomicina. La diseminación de los
microorganismos resistentes a meticilina resulta, igualmente, difícil de
controlar debido a que el portador nasofaríngeo asintomático representa el
origen más frecuente de estos microorganismos. Sin embargo, el uso de
quimioprofilaxis basada en vancomicina y rifampicina ha logrado algunos
20 IV. MATERIALES Y MÉTODOS
4.1. Descripción del área de estudio
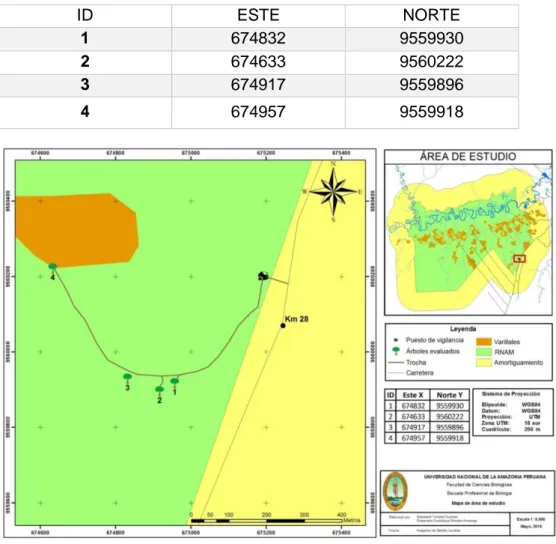
4.1.1. Lugar de recolección de muestras
La recolección de las muestras se realizó entre los meses de noviembre y
diciembre de 2014 en la Reserva Nacional de Allpahuayo Mishana Km 28 de
la carretera Iquitos-Nauta. Estas instalaciones se encuentran en el distrito de
San Juan Bautista, Provincia de Maynas, Región Loreto; ubicado en la
margen derecha del río Nanay, la altitud es de 122 msnm. La recolección se
realizó de 4 diferentes puntos, coordenadas UTM:
Cuadro 2. Coordenadas UTM del área de estudio
ID ESTE NORTE
1 674832 9559930
2 674633 9560222
3 674917 9559896
4 674957 9559918
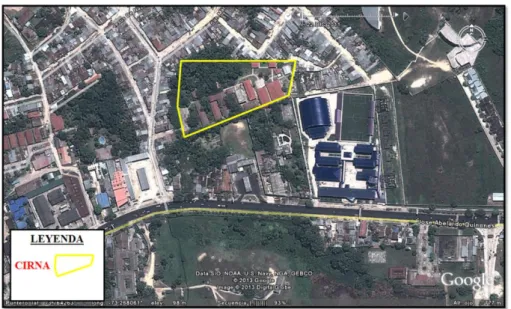
21 4.1.2. Lugar de procesamiento de las muestras
El procesamiento del hongo se llevó a cabo en los meses de enero a mayo
del presente año en el Laboratorio de Microbiología del Centro de
Investigación de Recursos Naturales de la UNAP (CIRNA-UNAP), ubicado en
el distrito de San Juan Bautista, Provincia de Maynas, Departamento de
Loreto.
Fuente: Google earth
Figura 2. Lugar de procesamiento de las muestras
4.2. Cepas bacterianas
Se utilizaron 5 cepas de Pseudomonas aeruginosa (Ps1, Ps2, Ps3, Ps4, Ps5) y 5
cepas de Staphylococcus aureus (St1, St2, St3, St4 y St5).
4.3. Procedimiento
4.3.1. Recolección del hongo
Para el registro de datos de campo en la recolección del hongo; se
anotaron en la ficha de colecta las características más resaltantes (color,
22
Se desprendió la seta entera utilizando una espátula y una navaja; con
mucho cuidado.
Se anotó las características del hábitat, tipo de bosque, tipo de substrato
en que crecen.
Se tomó fotografías con la finalidad de no perder sus características
originales.
4.3.2. Tratamiento de las muestras fúngicas
Los hongos colectados de los diferentes puntos fueron codificados como:
Ga1, Ga2, Ga3 y Ga4. Estos fueron transportados al laboratorio para ser
cortados en trozos pequeños, pesados y secados por un periodo de 8 días a
temperatura ambiente. Luego fueron pesados y molidos con un molinillo
manual de acero inoxidable. Posteriormente se pesaron y se colocaron los
pulverizados en diferentes frascos de vidrios previamente rotulados, junto con
el etanol para su posterior utilización.
23 4.3.3. Ensayos Químicos
4.3.3.1. Obtención del Extracto Etanólico
Para la preparación del extracto se empleó el método de maceración, con
etanol a temperatura ambiente, con renovación del solvente cada 4 días,
hasta agotamiento. El solvente fue eliminado mediante presión reducida a
55 °C. El extracto obtenido fue pesado y conservado en frasco de vidrio a
-10 °C para su posterior uso.
4.3.4. Evaluación de actividad antibacteriana por el método de difusión en agar
Trabajando por duplicado, se determinó la capacidad antibacteriana de los
extractos fúngicos frente a cinco cepas de Staphylococcus aureus y cinco
cepas de Pseudomonas aeruginosa, aisladas del hardware de computadoras
del Hospital Cesar Garayar - Iquitos.
4.3.4.1. Preparación de la solución stock de cada extracto fúngico
Se pesó 400 mg de cada extracto, se disolvió en 400 µl de agua destilada
estéril y así se obtuvo la solución stock (1 mg/µl), a partir de la cual, se
prepararon los discos de sensibilidad.
4.3.4.2. Preparación de los discos de sensibilidad
Empleando un perforador convencional, se confeccionaron discos de papel
de filtro Whatman Nº 3. Luego de ser esterilizados, a cada uno de ellos se
24 dejaron secar a 37 °C por espacio de 24 h. Se empleó como control, discos
de antibióticos de tetraciclina para S. aureus y cefepime para P.
aeruginosa.
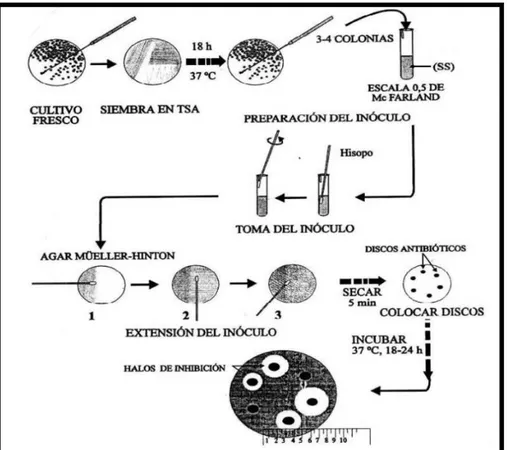
4.3.4.3. Ajuste y aplicación del inóculo
Se sembraron las bacterias en Caldo Tripticasa de Soya (TSB) y se incubó
a 37 °C por espacio de 18 a 24 horas. Luego, se repicó en placas con TSA
e incubaron por espacio de 18 a 24 h, para obtener cultivos jóvenes.
Luego, se procedió a preparar las suspensiones bacterianas en suero
fisiológico al 0,8 %, ajustándose la turbidez al tubo 0,5 de la escala de Mc
Farland (aprox. 1.5 x 108 UFC/ml). Se colocó 0.1 ml de la suspensión
bacteriana en la superficie de una placa con agar Mueller Hinton y se
diseminó sobre la superficie con la ayuda de un hisopo estéril. Se dejó
secar la placa a temperatura ambiente durante 3 a 5 minutos para que
cualquier exceso de humedad superficial sea absorbido. Luego, se
25 4.3.4.4. Aplicación de los discos
Discos impregnados con los extractos fúngicos y el disco de antibiótico
control, fueron colocados sobre la superficie del agar, a una distancia
mínima de 25 mm uno del otro para evitar la superposición de las zonas de
actividad. El disco no fue removido una vez que tomó contacto con la
superficie del agar. Se incubaron las placas en posición invertida a 37º C
por 18 a 24 horas. Transcurrido dicho periodo, se registró la actividad en
26 4.3.5. Determinación de la concentración inhibidora mínima y la concentración
bactericida mínima de los extractos fúngicos que muestren actividad
antibacteriana
4.3.5.1 Determinación de la concentración inhibitoria mínima (CIM)
Se empleó el método de macrodilución en caldo, para lo cual se
prepararon cultivos de 18 h, de las bacterias en estudio, en agar tripticasa
de soya. Luego, se preparó una suspensión bacteriana en solución salina
estéril a una turbidez equivalente al estándar 0.5 de la escala de Mc
Farland (aproximadamente 1.5 x 108 UFC/ml). A continuación, se hizo una
dilución al 1/100 (aproximadamente 1.5 x 106 UFC/ml) transfiriendo 100 µL
(0.1 ml) de la suspensión bacteriana a 9.9 ml de caldo tripticasa de soya
(TSB).
Se tuvo listo 10 tubos con 1 ml de caldo tripticasa de soya para cada
prueba (tubo N°1 al tubo N°10). Por otro lado, se preparó una solución
madre del extracto fúngico a una concentración de 500 mg/ml. Se añadió
1ml de la solución madre del extracto al tubo N° 1 que contiene 1ml de
caldo tripticasa de soya (concentración del extracto en este tubo es 250
mg/ml), se mezcló bien con la ayuda de un vórtex. A partir de este tubo, se
prepararon diluciones dobles seriadas, para lo cual se tomó 1 ml y se
transfirió al tubo N°2 (concentración del extracto = 125 mg/ml), después de
mezclar bien el contenido, se transfirió 1 ml al tercer tubo, del cual la
concentración de extracto fue 62.5 mg/ml y así sucesivamente hasta el
tubo N° 10, del cual se tomó y descartó 1 ml. De este modo, se obtuvieron
diluciones dobles de cada extracto desde 250 mg/ml hasta 0.49 mg/ml. A
27 del inóculo preparado de la cepa bacteriana que contiene
aproximadamente 1.5 x 106 UFC/ml, esto supone un inóculo final
aproximado de 7.5 x 105 UFC/ml y las concentraciones finales de los
extractos fueron desde 125 mg/ml hasta 0.24 mg/ml. Los tubos fueron
incubados a 37 °C durante 18 horas y luego se procedió a calcular la CIM,
considerándolo como la concentración correspondiente al tubo con menor
concentración del extracto donde no hubo desarrollo bacteriano,
29
4.3.5.2
Determinación de la concentración bactericida mínima (CBM)
A partir de cada uno de los tubos sin desarrollo bacteriano, se inoculó 0.1
ml en placas con agar tripticasa de soya y con una asa de vidrio se
dispersó en toda la superficie del agar en tres direcciones, las que se
incubaron a 37 °C durante 18 horas. Finalmente, para determinar la CBM
se contó el número de colonias en las placas, considerándose dicha CBM
como la menor concentración del extracto cuyo subcultivo produce un
número de colonias menor al 0.1% del inóculo original (7.5 x 105UFC/ml),
es decir, un número menor a 750 UFC/ml, y como se inoculó la décima
parte de 1 ml (0.1 ml), entonces se consideró la CBM al subcultivo que
produjera menos de 75 colonias.
30 V. RESULTADOS
5.1. Sensibilidad de las cepas de P. aeruginosa y S. aureus frente a los
extractos, mediante difusión en disco
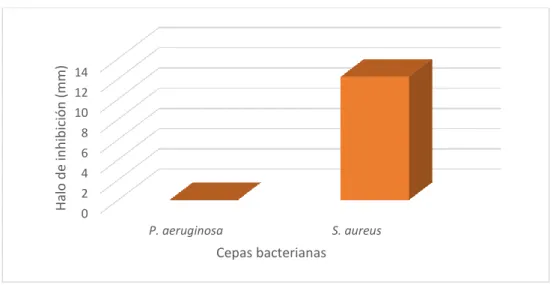
Figura 7. Halo de inhibicion frenta a las cepas de P. aeruginosa y S. aureus
En esta figura se puede observar que los extractos de G. applanatum no mostró
actividad antibacteriana frente a cepas de P. aeruginosa, debido a que no se formó
ningún halo de inhibición alrededor de los discos permitiendo el libre crecimiento de la
bacteria en toda la superficie del agar. Sin embargo, S. aureus mostró sensibilidad
frente a los extractos de G. applanatum.
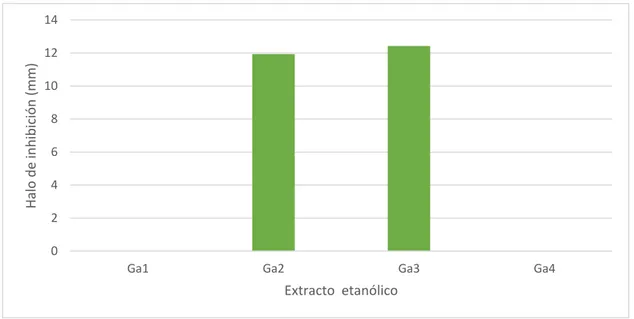
31 Figura 8. Actividad antibacteriana de los extractos de G. applanatum frente a las cepas
de S. aureus
En la prueba de sensibilidad se puede observar que los extractos de G. applanatum
Ga2 y Ga3 mostraron actividad antibacteriana frente a las 5 cepas de las
Staphylococcus aureus (Figura 8), siendo el extracto Ga3 el que obtuvo, en promedio,
el mayor halo de inhibición con 12,43 mm de diámetro, seguido del extracto Ga2 con
11,94 mm de diámetro de halo de inhibición, mientras que los extractos Ga1 y Ga4 no
mostraron ninguna actividad (Cuadro 3).
32 Figura 9. Datos descriptivos de los halos de inhibición de los extractos con actividad
frente a S. aureus
Mediante la estadística descriptiva podemos observar que el promedio del halo de
inhibición es de 11,94 mm para Ga2 y 12,43 mm para Ga3 con una desviación
estándar de 2,9 mm para Ga2 y 1,39 mm para Ga3 (Cuadro 4). Siendo el más preciso
el extracto Ga3, debido a que su desviación estándar está cerca al promedio. Además
el extracto Ga2 presenta un valor máximo de 16,3 mm y un valor mínimo de 8,7 mm; y
el extracto Ga3 muestra un valor máximo de 14,5 mm y un valor mínimo de 11mm
33
5.2. Concentración inhibitoria mínima (CIM) y concentración bactericida mínima
(CBM) de los extractos frente a las cepas de S. aureus
5.2.1. Concentración inhibitoria mínima de los extractos etanólicos frente a las
cepas de S. aureus
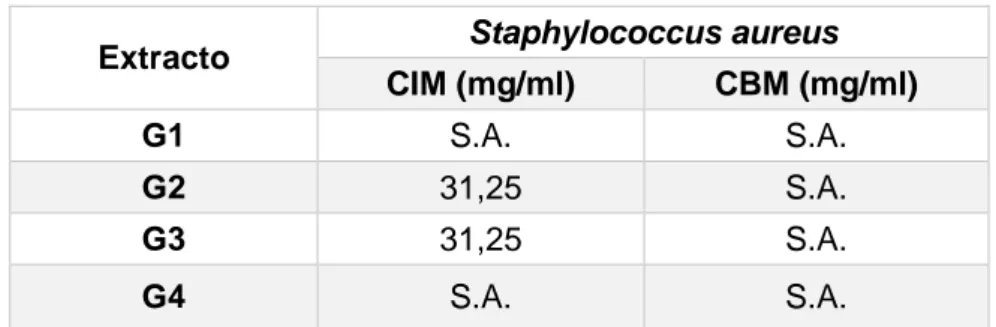
Cuadro 5. Concentración inhibitoria y bactericida mínima de los extractos frente a
S. aureus
S.A.: Sin Actividad
Después de evaluar cada uno de los extractos frente a las 5 cepas de S. aureus se
determinó que los extractos Ga2 y Ga3 mostraron una concentración inhibitoria
mínima (CIM) de 31,25 mg/ml, mientras que los extractos Ga1 y Ga4 no mostraron
resultado (Cuadro 5).
Cuadro 6. Concentración Inhibitoria Mínima de los extractos etanólicos al 31,25 mg/ml frente a S. aureus
0: Sin actividad. 1: Con Actividad
34 En el análisis estadístico se puede observar que el 40% de los extractos etanólicos al
31,25 mg/ml de concentración mostró inhibición en el crecimiento de las cepas de S.
aureus, mientras que el 60% no mostró inhibición (Cuadro 6).
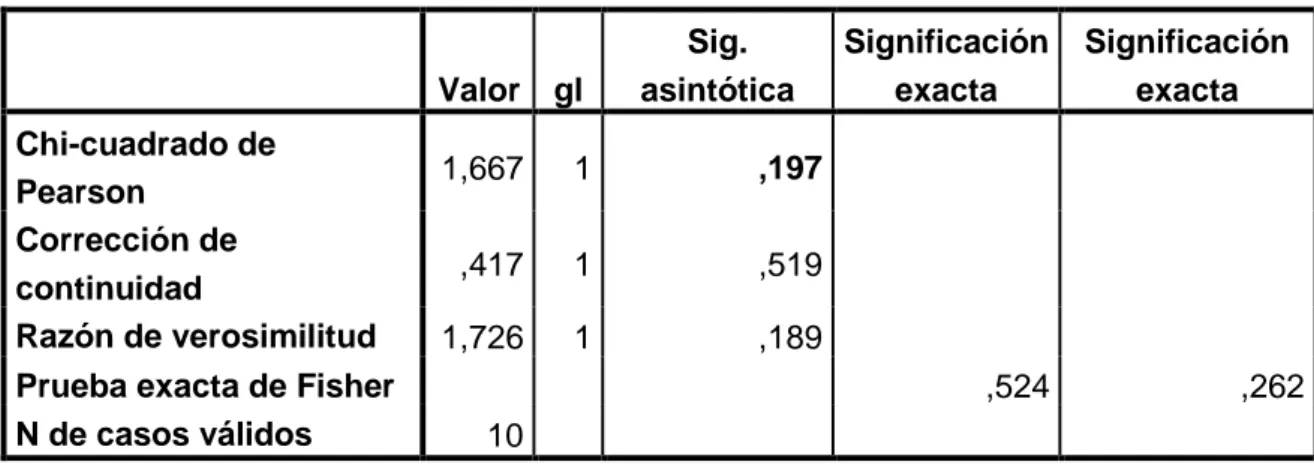
Cuadro 7. Prueba estadística de chi-cuadrado
Valor gl
Estadísticamente con la prueba de X2, no existe diferencia significativa (p = 0,519, con
corrección de Yates) de los extractos Ga2 y Ga3 sobre la actividad antibacteriana
(Cuadro 7).
5.2.2. Concentración Bactericida Mínima de los extractos etanólicos frente a
las cepas de S. aureus
En las pruebas de la concentración bactericida mínima, ninguno de los
extractos mostró actividad, ya que siempre se encontró crecimiento superior a
35 VI. DISCUSIÓN
Los hongos del género Ganoderma han sido empleados desde la antigüedad
como una fuente de la medicina natural, debido a la comprobación de las diversas
actividades biológicas que muestran sus metabolitos secundarios de gran
importancia por su amplio beneficio para la salud. Además, estos organismos han
sido estudiados por sus diferentes propiedades terapéuticas como antitumoral y
antiviral, pero muy pocas investigaciones han sido llevadas a cabo para
determinar su capacidad antibacteriana. Keypour et al. (2008) investigaron la
actividad antibacteriana del extracto de cloroformo de G. lucidum, logrando
demostrar un efecto inhibitorio sobre el crecimiento de B. subtilis y S. aureus. Así
mismo, Shekhar et al. (2010) demostraron que los extractos de sesquiterpenoides
de G. lucidum, G. chalceum, G. lipsiense y G. stipitatum presentaron una fuerte
actividad contra S. aureus. En concordancia con los resultados de los
investigadores realizadas por los grupos del Dr. Keypour y Dr. Shekhar, el hongo
G.applanatum, evaluado en este estudio también presentó acción antibacteriana
frente a S. aureus, lo cual indicaría que estas dos especies de Ganoderma podría
ser utilizadas para combatir las infecciones estafilocócicas. Sin embargo, su
actividad antibacteriana podría diferir dependiendo del tipo de extracto utilizado,
tal es el caso de los resultados registrados por Zuluaga et al. (2007), quienes
evaluaron la actividad antimicrobiana del extracto etanólico de G. lucidum
empleando el método de dilución en caldo, obtuvieron resultados negativos frente
a S. aureus y Escherichia coli en todas las concentraciones estudiadas.
Ameri et al. (2011), manifestaron que el extracto de acetato de etilo de G.
praelongum, G. resinaceum y G. lucidum exhibieron una gran actividad
antibacteriana contra S. aureus Resistente a Meticilina (MRSA) (19 mm; 14,2 mm
36 demostraron que la combinación del gel que contiene extractos acuosos de G.
praelongum posee capacidad para inhibir el crecimiento de MRSA y promover la
cicatrización de las heridas en ratones. Así mismo, Prasad et al. (2008), han
demostrado que el extracto metanólico de G. lucidum de la India posee una
actividad antibacteriana eficiente contra MRSA. De acuerdo con estos estudios
realizados por los grupos de investigadores del Dr. Ameri y Dr. Prasad, el hongo
G. applanatum podría ser una posible alternativa de solución para combatir a S.
aureus multidrogorresistente. No obstante, que en este trabajo se utilizaron cepas
drogorresistente MRSA negativo (Trinidad 2013), sería importante realizar este
tipo de investigación frente a cepas MRSA, debido a que en este estudio se
obtuvo resultados positivos del extracto etanólico mediante el método de difusión
en agar frente a S. aureus.
Guzmán et al. (2013) evaluaron la actividad antibacteriana del extracto etanólico y
de diclorometano de G. applanatum mediante el método de difusión en disco a
tres concentraciones de 200, 150 y 100 mg/ml, mostrándose buena actividad
antibacteriana a 150 mg/ml frente a S. aureus. De acuerdo con los estudios
realizados por Guzmán, en este presente trabajo también se observó que G.
applanatum presentaron actividad frente a S. aureus. Así mismo, estudios
realizados por Nagaraj et al. (2013) quienes evaluaron el potencial antibacteriano
in vitro del extracto metanólico del G. applanatum por el método de difusión en
agar, a concentraciones de 70 mg/ml, 50 mg/ml y 25 mg/ml; registraron actividad
antibacteriana significativa contra S. aureus (15mm) y Pseudomonas aeruginosa
(12mm). En contraste con este estudio, que encontraron actividad antibacteriana
frente a P. aeruginosa, el presente estudio no evidencio actividad antibacteriana
del extracto etanólico de G. applanatum frente a P. aeruginosa y esto podría
deberse a la diferencia en el tipo de extracto empleado o que las cepas
37 Osinska et al. (2014), mencionaron que la actividad antibacteriana de la fracción
de exopolisacáridos de G. applanatum (GpEPS), presentaron propiedades
antibacterianas frente a las cepas de S. aureus (ATCC 25923) con una zona de
inhibición de 17,9 mm y un valor de CIM de 1 mg/ml. El grupo de investigadores
del Dr. Keypour, quienes demostraron que el extracto de cloroformo de G. lucidum
presentó un efecto antibacteriano frente a B. subtilis y S. aureus, obtuvieron un
CIM y CBM de 8 mg/ml. A diferencia con lo mencionado por el grupo de
investigadores del Dr. Osinska y del Dr. keypour, en el presente estudio, en la que
también se observó actividad antibacteriana frente S. aureus, la zona de inhibición
promedio máximo que se mostró fue de 12,43 mm, con un CIM de 31,25 mg/ml, la
diferencia observada entre el valor del CIM del Dr. Osinska y de la presente
investigación, puede deberse a los exopolisacáridos de G. applanatum utilizados
38 VII. CONCLUSIONES
Ninguno de los extractos fúngicos ensayados mostró actividad contra
Pseudomonas aeruginosa.
Los extractos de Ga2 y Ga3 mostraron actividad frente a Staphylococcus
aureus.
Los extractos que mostraron actividad frente S. aureus obtuvieron una
Concentración Inhibitoria Mínima (CIM) de 31,25 mg/ml.
Los extractos fúngicos de Ga2 y Ga3 mostraron actividad inhibitoria, pero no
39 VIII. RECOMENDACIONES
Continuar la investigación utilizando otros tipos de extractos de la especie G.
applanatum, así como de otras especies fúngicas.
A fin de obtener mejores resultados con respecto a Staphylococcus aureus, se
sugiere realizar estudios similares con mayor número de cepas de dicha
40 IX. REFERENCIAS BIBLIOGRÁFICAS
ALBINO S.E.F., DELLE M.F., YUNES R.A., PAULERT R., SMANIA J.A. 2007.
Antimicrobial activity of methyl australate from Ganoderma australe. Rev.
Brasileira de Farmacognosia. 17(1): 14 - 16.
AMERI A. RAJIVE B.B., VAIDYA J.G., APTE K., DEOKULE S.S. 2008.
Anti-Staphylococcal and wound healing activities of Ganoderma praelongum and
Glycyrrhiza glabra formulation in mice. International Journal of Applied Research
in Natural Products. 6 (1): 27 - 31.
AMERI A., VAIDYA J.G., DEOKULE S.S. 2011. In vitro evaluation of
anti-staphylococcal activity of Ganoderma lucidum, Ganoderma praelongum and
Ganoderma resinaceum from Pune, India. African Journal of Microbiology
Research. 5(3): 328-333. En: http://www.academicjournals.org/ajmr
BARROS L., CALHELHA R.C., VAZ J.A., FERREIRA I.C.F .R., BAPTISTA P.,
ESTEVINHO L. M. 2007. Antimicrobial activity and bioactive compounds of
Portuguese wild edible mushrooms methanolic extracts. European Food
Research Technology. 225(2): 151–156.
BRIZUELA M. A., GARCÍA L., PÉREZ L., MANSUR M. 1998. Basidiomicetos: nueva
fuente de metabolitos secundarios. Rev Iberoam Micol 15: 69-74.
BROOKS F. G.; CARROLL C. K.; BUTEL S. J.; MORSE A. S.; MIETZNER A. T. 2012.
Jawetz, Melnick y Adelberg: Microbiología medica.25ª Edición. Editorial McGraw
41 ESPINOZA M. 2004. Determinación de hongos de la clase Basidiomycetes en el
Centro de Investigaciones Allpahuayo. Loreto – Perú. 59 – 61 pp.
GARCÍA-PAJÓN C.M., COLLADO I.G. 2003. Secondary metabolites isolated from Colletotrichum species. Natural Product Reports. 20(1): 426 – 431.
GÓMEZ J., ALCÁNTARA M., SIMARRO E., MARTÍNEZ B., RUÍZ J., GUERRA B.,
GÓMEZ V. J. Y VALDÉS M. 2002. Bacteriemias por Pseudomonas aeruginosa:
epidemiología, clínica y tratamiento. Estudio prospectivo de siete años.
GÓMEZ R. L., GIL J., CASTILLO J., RUBIO M. C. 1993. Impacto de los inhibidores de
beta-lactamasas en la susceptibilidad antibiótica de los patógenos más
frecuentes. En: Betalactamasas: su importancia para el clínico. Madrid: Smith
Kline& French S.A.E. 109-127.
expression from Ganoderma applanatum. 683 – 686.
KARSTEN P. A. 1881. Enumeratio boletinearum et polyporearum fenicarum.
Systemate novo dispositarum. Reviews Mycology 3: 101-134. Mencionado por
Tello S.I. 2010. Diversidad de los recursos genéticos mexicanos del hongo
funcional Ganoderma (Fungi, Ganodermataceae), conocido como Reishi en los
42 KEYPOUR S, RIAHI H, MORADALI MF, RAFATI H. 2008. Investigation of the
antibacterial activity of a chloroform extract of Ling Zhi or Reishi Medicinal
Mushroom, Ganoderma lucidum (W. Curt.: Fr) P. Karst. (Aphyllophoromycetideae) from Iran. Inter. J. Med. Mushrooms. 10(4): 345 – 349.
KIRK P.M., CANNON P.F., MINTER D. W., STALPERS J.A. 2008. Dictionary of the
Fungi. 10º Ed. CAB International.
LESKOSEK-CUKALOVIC I., DESPOTOVIC S., LAKIC N., NIKSIC V., TESEVIC V.
2010. Ganoderma lucidum — Medical mushroom as a raw material for beer with
enhanced functional properties. Food Research International. 43(1): 2262-2269.
LLOP H.A., VALDÉS–DAPENA V.M., ZUAZO S.J. 2001. Microbiología y Parasitología
Médicas. Edit. Ciencias Médicas, La Habana. Tomo I. 303 pp
MATA M. 2003. Macrohongos de Costa Rica. Instituto Nacional de Biodiversidad.
INBio, Edit. INBio, Santo Domingo de Heredia, Costa Rica. Vol. 1. 255pp
MATTILA P., SUONPÄÄ K., PIIRONEN V. 2000. Functional properties of edible
mushrooms. Nutrition 16: 694 – 6.
MONCALVO J.M., RYVARDEN L. 1997. A nomenclatural study of the
Ganodermataceae Donk. Oslo: Fungiflora.
MORI Á.T., BENDAYÁN A.M.E., TRESIERRA A.Á., GARCÍA D.M., RUÍZ S.E.,
BARDALES G.J., REÁTEGUI A.R., ESPINOZA C.F., DÁVILA F.C. 2011.
43 MURRAY R.P., ROSENTHAL K.S., PFALLER A. M.2010. Microbiología Médica.
Capítulo 21: Staphylococcus y cocos grampositivos relacionados. 6a Edición.
Editorial. Elsevier - Mosby España. 209-224.
MURRAY R.P., ROSENTHAL K.S., PFALLER A.M. 2007. Microbiología Médica.
Capítulo 22: Staphylococcus y microorganismos relacionados. 5a Edición.
Editorial. Elsevier - Mosby España. 221-236.
NAGARAJ K., MALLIKARJUN N., NAIKA R., VENUGOPAL T.M. 2013. Phytochemical
Analysis and In Vitro Antimicrobial Potential of Ganoderma applanatum (Pers.)
Pat. Of Shivamogga District – Karnataka, India. International Journal
Pharmaceutical Sciences Review and Research, 23(2); nᵒ 07. 36-41.
OSINSKA-JAROSZUK M., JASZEK M., MIZERKA-DUDKA M., BLACHOWICZ A.,
PIOTR R.T., JANUSZ G., WYDRYCH J., POLAK J., JAROSZ-WILKOLAZKA A.,
KANDEFER-SZERSZEN M. 2014. Exopolysaccharide from Ganoderma
applanatum as a promising bioactive compound with cytostatic and antibacterial
properties. BioMed Research International. 1: 1 – 10.
PRASAD Y., WESELY W.E.G. 2008. Antibacterial activity of the bio - multidrug (G.
lucidum) on multidrug resistant S. aureus (MRSA). Advanced Biotech. 6(9): 9 -
16.
RAMÍREZ G.G. 2003. Apuntes de Microbiología y Bacteriología. Edit. Unidad de
44 RAMOS S.D.O., CAPPELLO G.S. 1998. Revision del genero Ganoserma
(Basidiomycota) para el estado de Tabasco. 7 pp.
RIOS-CAÑAVATE, J.L. 2008. Ganoderma lucidum, un hongo con propiedades
inmunoestimulantes. Revista de Fitoterapia 8(2): 135-146.
RUSSELL R., PATERSON M. 2006. Ganoderma: a therapeutic fungal biofactory. Phytochemistry. 67: 1985–2001.
RYVARDEN L. 1998. African Polypores a review. Belgique Journal Botanique. 131(2):
150-155.
SHEKHAR R.B., GAURI B., JITENDRA G.V., SANDHYA A.G., HIRALAL B.S. 2010.
Antimicrobial activity of terpenoid extracts from Ganoderma samples. Revisado
en: www.ijplsjournal.com
SMITH J., ROWAN N., SULLIVAN R. 2002. Medicinal mushrooms: their therapeutic
properties and current medical usage with special emphasis on cancer
treatments. Reino Unido: University of Strathclyde.
TAWFIK M.M., ABDUL-HAFIZ A., AL-DUBOON, KAWTHER T. K. 2011. Bioactive
Compounds from a Polypore Fungus Ganoderma applanatum (Per s. ex Wallr.)
Pat. Jordan Journal of Biological Sciences. 4 (4): 205 – 212.
TELLO S.I. 2010. Diversidad de los recursos genéticos mexicanos del hongo funcional
Ganoderma (Fungi, Ganodermataceae), conocido como Reishi en los mercados
internacionales, y su relevancia para el desarrollo regional. Colegio
45 TRINIDAD G.B. 2013. Determinación del Perfil de Resistencia a antibióticos de cepas
de Staphylococcus aureus. Centro de Investigación de Recursos
Naturales-CIRNA Práctica Pre Profesional. UNAP. Iquitos, Perú. 17 p.
VOLK TJ. 1998. Why polyporus has been split into more than 100 genera. Mycophile. 39 (2): 1 – 3.
WACHTEL-GALOR S., TOMLINSON B., BENZOE I.F.F. 2004. Ganoderma lucidum
(“Lingzhi”), a Chinese medicinal mushroom: biomarker responses in a controlled
human suplementation study. Br J Nutr. 91: 263-269.
ZULUAGA V.J., PÉREZ C.C., ANGULO A. TORRES O., SANTAFÉ G. 2007. Química
y actividades antioxidante y bactericida del extracto etanólico del hongo
Ganoderma lucidum. Scientia et Technica Año XIII. N° 33. 329 – 332.
ZJAWIONY J.K. 2004. Biologically active compounds form Aphyllophorales (Polypore)
46 X. ANEXOS
Anexo 1. Obtención del extracto etanólico
Anexo 2. Elaboración e impregnación de los discos con los extractos
47 Anexo 3. Sembrando las cepas bacterianas en medio TSB para la incubación
48 Anexo 5. Colocación de los discos impregnados con los extractos en agar MH
49 Anexo 8. Determinación de la Concentración Inhibitoria Mínima (CIM) del extracto
fúngico (C) frente a S. aureus
50 Anexo 9. Determinación de la Concentración Inhibitoria Mínima (CIM) del extracto
fúngico (Ca) frente a S. aureus
51 Anexo 11. Determinación de la Concentración Bactericida Mínima (CBM) de los