Efectos de la fragmentación sobre la distribución de especies arbóreas en el Parque Nacional Fray Jorge : importancia de los atributos ecofisiológicos /
116
0
0
Texto completo
(2) II. PONTIFICIA UNIVERSIDAD CATÓLICA DE CHILE FACULTAD DE CIENCIAS BIOLÓGICAS PROGRAMA DOCTORADO EN CIENCIAS BIOLÓGICAS MENCIÓN ECOLOGÍA. EFECTOS DE LA FRAGMENTACIÓN SOBRE LA DISTRIBUCIÓN DE ESPECIES ARBÓREAS EN EL PARQUE NACIONAL FRAY JORGE: IMPORTANCIA DE LOS ATRIBUTOS ECOFISIOLÓGICOS. Por. BEATRIZ EUGENIA SALGADO NEGRET. Tesis presentada a la Facultad de Ciencias Biológicas de la Pontificia Universidad Católica de Chile para optar al grado académico de Doctor en Ciencias Biológicas mención Ecología. Dirigida por: Dr. Juan José Armesto Dra. Fernanda Pérez. Noviembre, 2013 Santiago, Chile.
(3) III. Agradecimientos Quiero comenzar agradeciendo a la Comisión Nacional de Investigación Científica y Tecnológica (CONICYT, Chile) y al Instituto de Ecología y Biodiversidad (IEB) por el apoyo financiero para realizar este doctorado y el trabajo de investigación. Quiero agradecer a mi tutor principal Juan Armesto, por apostar a ciegas y permitirme ser parte de su equipo, por sus invaluables enseñanzas y apoyo incondicional. A todos los miembros de su Laboratorio por todas las tertulias e increíbles discusiones. A Fernanda Pérez por ser una excelente guía, por las eternas discusiones teóricas, por todas las salidas de campo, pero sobretodo por convertirse en una gran amiga y confidente. Fefita, eres de los grandes regalos que me llevo de Chile…gracias por todo! A Fernando Valladares y su equipo por adoptarme por meses en su laboratorio y acogerme como un miembro más del equipo. Gracias por todos los análisis y discusiones que mejoraron este documento. A Pablo Marquet, Javier Figueroa y Martín Carmona por sus aportes y comentarios que mejoraron esta propuesta desde sus inicios. A Mylthon Jimenez-Castillo y su equipo por enseñarme el mundo de la hidráulica. Gracias especiales a Paulina Lobos. A Juan Monardez, por su fiel compañía, por las deliciosas cenas y discusiones en compañía del mejor vino. Juan mil gracias por presentarme un ecosistema maravilloso. A Aurora Gaxiola y Daniel Stanton por las múltiples charlas planteando hipótesis y discutiendo resultados…. sus comentarios mejoraron enormemente los manuscritos y su compañía fue un gran apoyo. A Felipe Albornoz, Rafaella Canessa, Carmen Ossa, Daniel Salinas, Patricio Valenzuela e Isabel Mujica por su invaluable apoyo en campo y laboratorio y por hacer de las salidas de campo paseos repletos de risas y complicidad. A Mariela Aguilera y Ximena Alvarez por todas las reuniones, discusiones y tertulias alrededor de la fisiología de las plantas…hubo momentos brillantes…gracias queridas! A mis compañeras de batalla y familia en Chile: Lidia Mansur, Sabrina Clavijo, Daniela Rivera y Carmencha Ossa…. no habría sido lo mismo sin ustedes. A Leo por todo su apoyo durante tantos años..
(4) IV. A Carolina Alcázar, Olga Caro y Carolina Useche por ser mis terapeutas en la distancia…gracias por todo el apoyo. A mi familia en Colombia….por darme la libertad de soñar y por estar al pie del cañón….mil gracias por estar siempre tan cerca a pesar de la distancia. A todas aquellas personas que no he nombrado pero que hicieron parte de este logro. Muchas gracias..
(5) V. Tabla de contenidos Lista de abreviaturas ............................................................VIII Resumen. .................................................................................IX. Introducción General……………….………………………..………1 Estructura de la tesis…………………………................................... 6. Área de estudio............................................................................. 7. Visión general…........................................................................... 9. Referencias................................................................................... 11. Capítulo I Estrategias divergentes de tolerancia a la sequía explican la distribución de especies arbóreas a través de un gradiente de humedad dependiente de neblina en un bosque lluvioso templado Abstract......................................................................................... 17 Introduction................................................................................... 18. Materials and Methods................................................................. 20 Results........................................................................................... 25. Discussion..................................................................................... 27. Acknowledgements………………………………………………31 References..................................................................................... 32. Tables............................................................................................ 38. Figures.......................................................................................... 41.
(6) VI. Capítulo II Variación en rasgos funcionales explica la distribución de Aextoxicon punctatum a través de un fuerte gradiente de humedad en un bosque fragmentado dependiente de neblina Abstract......................................................................................... 47. Introduction................................................................................... 48. Materials and Methods.................................................................. 51. Results............................................................................................ 54. Discussion...................................................................................... 56. Acknowledgements…………………………………………………….. 59 References...................................................................................... 60. Tables............................................................................................. 67. Figures........................................................................................... 70. Online supplemental materials……………………………………74. Conclusiones Generales Conclusiones................................................................................. 78. Anexo I Simetría de los parches de bosque depende de la dirección de los recursos limitantes Abstract......................................................................................... 84. Introduction................................................................................... 85. Materials and Methods.................................................................. 88. Results............................................................................................ 90. Discussion...................................................................................... 91. Acknowledgements……………………..……………………………….95.
(7) VII. References...................................................................................... 96. Tables........................................................................................100 Figures..................................................................................... 102.
(8) VIII. Lista de abreviaturas. AMAX = Photosynthetic rate; Tasa fotosíntesis gs = Stomatal conductance; Conductancia estomática Hv = Huber value; Valor Huber Ks = Sapwood-specific hydraulic conductivity LA = Leaf area; Área foliar LMA = Leaf mass area; Relación masa: área de la hoja PLC = Percentage of loss conductivity; Porcentaje de pérdida de conductividad RWCtlp = Relative water content at turgor loss point; Contenido relativo de agua al punto de pérdida de turgor SD = Stomatal density; Densidad estomática TD = Trichome density; Densidad de tricomas VD = Vessel density; Densidad de vasos VDi = Vessel diameter, Diámetro de vasos π0 = Solute potential at full turgor; Potencial de solutes a full turgor πtlp = Water potential at turgor loss; Potencial hídrico al punto de pérdida de turgor ɛ = Bulk modulus of elasticity; Modulo de elasticidad ψPD = Leaf water potentials predawn; Potencial hídrico al amanecer ψMD = Leaf water potentials at midday; Potencial hídrico al medio día.
(9) IX. Resumen El estudio de los rasgos funcionales y mecanismos fisiológicos que determinan la tolerancia de las especies a la sequía y su habilidad para competir por agua es fundamental para entender su distribución a través de gradientes de humedad y predecir su respuesta al cambio global, donde la fragmentación del hábitat y el cambio de uso del suelo son los principales motores de cambio. En este sentido, los bosques dependientes de neblina en las regiones semiáridas del mundo son un buen modelo de estudio para entender las respuestas de las especies al incremento en la aridez y la fragmentación del hábitat. En esta tesis se estudiaron los mecanismos fisiológicos que explican los patrones contrastantes de distribución observados a través de gradientes de humedad generados por la neblina costera en las tres principales especies arbóreas Aextoxicon punctatum, Drimys winteri y Myrceugenia correifolia que coexisten en los fragmentos de bosque del Parque Nacional Fray Jorge, en la región semiárida en Chile. Se identificó un continuo de estrategias en el uso de agua explicando la distribución de las especies a través del gradiente de humedad a pequeña escala. Drimys winteri, una especie restringida al núcleo húmedo, mostró rasgos que permiten un eficiente transporte de agua y ganancia de carbono; en contraste, Myrceugenia correifolia, especie que domina los bordes secos de sotavento, presentó rasgos que promueven la conservación del agua y menores tasas de intercambio de gases, así como menor potencial hídrico al punto de pérdida de turgor. La especie con amplia distribución Aextoxicon punctatum, mostró valores de rasgos intermedios, pero se observó variación de las medias, magnitud e integración fenotípica a través de las zonas dentro de los fragmentos. Así, árboles creciendo en los bordes secos presentaron mayor LMA, densidad de estomas y tricomas que los árboles del núcleo húmedo y el borde barlovento. En contraste, rasgos de la anatomía del xilema no variaron produciendo pérdida de la conductividad hidráulica en los bordes más secos. También se detectaron mayores niveles de integración fenotípica y variabilidad en los bordes secos. Los resultados mostraron que el particionamiento del pronunciado gradiente de humedad a pequeña escala entre las especies arbóreas está determinado por las tolerancias diferenciales de las especies a la sequía, y esas diferencias indican que las especies tienen habilidades contrastantes para lidiar con futuros cambios climáticos..
(10) I. Introducción General.
(11) 2 La disponibilidad de agua es el principal factor que determina la distribución de las especies arbóreas a través de gradientes de lluvia a gran escala así como en gradientes topográficos a pequeña escala (Gentry 1988, Wright 1992, Condit 1998, Bongers et al. 1999, Pyke et al. 2001, Condit et al. 2002, Engelbrecht et al. 2007). El estudio de los rasgos funcionales asociados al comportamiento de las especies bajo condiciones particulares de humedad del suelo ayuda a explicar la distribución de las especies (Poorter 2007, Markesteijn et al. 2011, Sterck et al. 2011), donde el éxito en el establecimiento y sobrevivencia en ambientes o épocas secas estará determinado por su habilidad para competir por agua y tolerar la sequía (Markesteijn et al. 2011). La capacidad de respuesta de las especies a la sequía y a cualquier variable ambiental está determinada por sus rasgos funcionales, los cuales son todas las características morfológicas, fisiológicas o fenológicas medidas a nivel individual (Viollé et al. 2007). Es bien conocido el trade off entre la adquisición y conservación de recursos que le permite a las especies especializarse a lo largo de esos gradientes ambientales (Reich et al. 2003, Diaz et al. 2004, Wright et al. 2004). Así, plantas que crecen en ambientes secos generalmente tienen hojas pequeñas, baja conductancia estomática, alta área foliar específica (e.j. Fahn 1986, Baldini et al. 1997, Niinemets 2001), pero presentan bajas tasas fotosintéticas y tasas de crecimiento (Reich et al. 2003). A nivel hidráulico también existen ciertos rasgos que determinan el establecimiento de las especies en determinados ambientes. Por ejemplo, especies que crecen en ambientes secos generalmente tienen vasos conductores más pequeños y densos con pequeños poros en las membranas que les permiten conducir agua bajo condiciones de baja disponibilidad hídrica disminuyendo el riesgo de embolismo. Estos rasgos incrementan la resistencia al flujo de agua y reducen la eficiencia hidráulica de las especies, afectando el suministro de agua a las hojas (Hacke et al. 2001, Choat et al. 2005, Markesteijn et al. 2011a,b). Según la combinación de rasgos funcionales, las plantas pueden estar ubicadas a través de un gradiente de estrategias (Reich et al. 2003, Díaz et al. 2004): en un extremo especies con rasgos que favorecen la conservación de los recursos (conservativas) a especies con rasgos que promueven la rápida captura de recursos (adquisitivas). Entender las estrategias y mecanismos que tienen las especies para sobrevivir a la sequía es crítico para predecir las consecuencias ecológicas de futuras alteraciones en la humedad del suelo debido a motores del cambio global como la fragmentación, el cambio de uso del suelo o el cambio climático..
(12) 3 La mayoría de los estudios relacionados con distribución de especies a través de gradientes de humedad, han examinado la variación de rasgos funcionales a nivel interespecifico (Pockman y Sperry 2000, Cornwell y Ackerly 2009, Engelbrecht et al. 2007, Choat et al. 2012, Salgado-Negret et al. 2013), considerando únicamente los valores promedio de los rasgos para cada especie, ignorado la importancia de la variabilidad intraespecífica. Esto puede responder a que las tendencias en comunidades diversas son principalmente el resultado del recambio de especies más que de la variación a nivel de especie (Cornwell y Ackerly 2009; Albert et al. 2010a,b; Hulshof y Swenson 2010). En ambientes con limitaciones hídricas, se ha propuesto una disminución de la variabilidad (coeficiente de variación) de los rasgos funcionales a nivel intraespecífico, debido a que solo individuos con un rango restringido de valores de rasgos es capaz de sobrevivir bajo esas condiciones ambientales (Cornwell y Ackerly 2009). A través de los gradientes de humedad del suelo también pueden variar las respuestas de los rasgos individuales (media y coeficiente de variación) y por lo tanto los patrones de correlación entre ellos (Pigliucci y Kolodynska 2002; Sardans, Penuelas y Roda 2006), y aunque en la literatura son bien conocidas las correlaciones entre rasgos foliares (Wright et al. 2004), rasgos hidráulicos (Chavé et al. 2009, Zanne et al. 2010) y entre ambos módulos vegetativos (Brodribb y Field 2000; Brodribb et al. 2002; Santiago et al. 2004; Wright et al. 2006; Meinzer et al. 2008; Baraloto et al. 2010), existe poca información acerca de cómo el ambiente puede alterar los patrones de correlación fenotípica entre rasgos de foliares y de madera (Nicotra et al. 1997, Wright et al. 2006). Uno de los ecosistemas con mayores limitaciones hídricas son los bosques dependientes de neblina encontrados en las regiones semiáridas del mundo (Hildebrandt y Eltahir 2006, del-Val et al. 2006, Katata et al. 2010). Estos bosques son relictos de periodos pasados cuando las condiciones fueron más húmedas, por lo cual son ecosistemas especialmente sensibles a los cambios actuales en la producción y distribución de la neblina. Se predice que alteraciones en la frecuencia e intensidad de la niebla ocurrirán debido a cambios en la temperatura superficial del mar y la altura de la capa de inversión térmica (Cereceda et al. 2002), pérdida de áreas de bosque y fragmentación o cambios en la estructura de los bosques afectando la captura de niebla (Hildebrandt y Eltahir 2006). En esos fragmentos de bosque, la intercepción de la niebla por las plantas es la principal o incluso la única fuente de agua durante la mayor parte del año (Dawson 1998, del-Val et al. 2006, Ewing et al. 2009). La intercepción por parte.
(13) 4 de la vegetación crea pronunciados gradientes de agua y nutrientes desde el borde barlovento (entrada de niebla) al borde sotavento de los parches (Weathers et al. 2000, del-Val et al. 2006, Ewing et al. 2009), con fuertes contrastes en cortas distancias (Ewing et al. 2009). Estudiar la respuesta de las especies a la variación en la humedad del suelo a cortas escalas espaciales generadas por gradientes topográficos o de fragmentación en estos ecosistemas, nos permite direccionar preguntas acerca las condiciones críticas para el mantenimiento de especies arbóreas bajo estrés por sequía debido a cambio climático. Un interesante ejemplo de bosques dependientes de neblina se encuentra en la región semiárida en Chile (30°S), donde un mosaico de más de 180 parches de bosque persiste en las montañas costeras rodeado por una matriz de vegetación xerofítica (Barbosa et al. 2010). Este bosque tuvo una distribución continua, pero el incremento en la aridez en el Terciario tardío dividió su distribución (Villagrán et al. 2004). Como consecuencia, este tipo de bosque quedó restringido al rango montañoso costero de la región Mediterránea en Chile inundado por niebla (Villagrán et al. 2004), la cual duplica la precipitación efectiva de esta zona (del-Val et al. 2006). Los fuertes gradientes de humedad generados por la intercepción de la neblina afectan la distribución y dinámica de las especies. Las especies arbóreas dominantes en esos parches son: Aextoxicon punctatum (Aextoxicaceae), que ocurre en todos los bosques pero prefiere el borde barlovento que recibe directamente la entrada de la neblina; Drimys winteri (Winteraceae) que tiende a estar agregada en el núcleo de los grandes parches de bosque; y Myrceugenia correifolia (Myrtaceae) que es más común en los parches pequeños y está normalmente confinada en los bordes sotavento más secos (del-Val et al. 2006; Gutiérrez et al. 2008). Estas distribuciones están determinadas por contrastantes patrones de regeneración y mortalidad dentro de los parches. El reclutamiento está concentrado en los bordes húmedos en barlovento y es tres veces mayor que en sotavento, mientras que la mortalidad es mayor en el borde sotavento opuesto al ingreso de la neblina costera (del-Val et al. 2006). Los contrastantes patrones de distribución de estas especies arbóreas ofrecen una gran oportunidad para valorar los mecanismos subyacentes a su habilidad para tolerar las condiciones secas y la variación en esos mecanismos a lo largo de gradientes de humedad espacial determinados por la entrada de la niebla. Las preguntas e hipótesis que se abordarán en esta tesis son las siguientes:.
(14) 5 1) ¿La variación de los rasgos foliares e hidráulicos relacionados con la tolerancia a la sequía explican los patrones de distribución contrastantes de tres especies arbóreas dominantes a través de gradientes de humedad a pequeña escala en los fragmentos de bosque del Parque Nacional Fray Jorge? Se espera que especies que crecen en sotavento bajo condiciones de déficit hídrico presenten un grupo de rasgos que favorezcan la conservación del agua (menor conductancia estomática) y que reduzcan el riesgo de cavitación (vasos angostos) con el costo de una menor efi ciencia hidráulica y fotosintética. 2) ¿Qué adaptaciones o mecanismos le permiten a los individuos de A. punctatum y M. correifolia crecer en los fragmentos pequeños o en los bordes secos de sotavento para lidiar con el déficit hídrico en comparación con individuos conespecíficos que creen en los núcleos húmedos de los fragmentos? Se espera que individuos que crecen en el borde seco en sotavento presenten rasgos fisiológicos que favorezcan la tolerancia a la sequía como menor πtlp y π0, en comparación con individuos conespecíficos que crecen en los núcleos húmedos de los fragmentos. Se espera que los individuos que crecen en fragmentos pequeños y en los bordes secos de sotavento tengan rasgos que favorezcan la conservación del agua como mayor densidad de tricomas y LMA y que reduzcan el riesgo de cavitación disminuyendo el diámetro de sus vasos conductores. 3) ¿La variabilidad e integración fenotípica incrementan en sotavento con mayor variabilidad ambiental y menor disponibilidad de agua? Se espera que la variabilidad e integración fenotípicas incrementen en los bordes de sotavento debido a la mayor variabilidad ambiental e incremento en el déficit hídrico. Las especies arbóreas que viven en los bosques de neblina del Parque Nacional Fray Jorge han estado expuestas a un incremento en la aridez debido a cambios climáticos ocurridos por periodos extendidos de tiempo (Villagrán et al. 2004; Gutiérrez et al. 2008), y han enfrentado cambios estacionales en la producción de la neblina que generan pronunciados gradientes de humedad dentro de los fragmentos (del-Val et al. 2006). Este estudio revela algunos de los mecanismos clave que explican el éxito de esas especies para coexistir dadas las variaciones pasadas y actuales en la disponibilidad de humedad del suelo. Los resultados se discuten a la luz de las posibles consecuencias de futuros cambios climáticos y sus efectos sobre la distribución y coexistencia de especies..
(15) 6. Estructura de la tesis En consideración a lo expuesto, en este proyecto de tesis se plantea como objetivo central estudiar los mecanismos fisiológicos que ayudan a explicar los patrones contrastantes de distribución y abundancia observados en las tres especies arbóreas dominantes en los fragmentos de bosque del Parque Nacional Fray Jorge: Aextoxicon punctatum, Drimys winteri y Myrceugenia correifolia. La intercepción de la niebla por parte de la vegetación en los bosques del Parque Nacional Fray Jorge genera fuertes gradientes de humedad del suelo, donde las zonas sotavento son más secas que los otros dos microhábitats, mientras que la humedad del suelo en las zonas barlovento (ingreso de la neblina) es comparable con los núcleos de los fragmentos (véase capitulo 1). Las especies arbóreas dominantes están distribuidas diferencialmente a través de este gradiente de humedad del suelo. Así, Aextoxicon punctatum (Aextoxicaceae), ocurre en todas las zonas de los parches pero prefiere el borde barlovento que recibe directamente la entrada de la neblina, Drimys winteri (Winteraceae) tiende a estar agregada en el núcleo de los grandes parches de bosque y Myrceugenia correifolia (Myrtaceae) es más común en los parches pequeños y está normalmente confinada en los bordes sotavento más secos (del-Val et al. 2006; Gutiérrez et al. 2008). Primero se estudiaron los rasgos foliares (área foliar, área foliar específica, tasa fotosintética, conductancia estomática) e hidráulicos (diámetro y densidad de vasos conductores, conductividad hidráulica específica de la madera y valor Huber) relacionados con la tolerancia a la sequía en las tres especies arbóreas (véase capitulo 1). Adicionalmente, se realizaron curvas presión-volumen para los individuos que crecen en el borde seco sotavento y en el núcleo húmedo, con el objetivo de entender cuáles eran los mecanismos de las especies para lidiar con el déficit hídrico (ajuste osmótico o incremento en la elasticidad celular) en sotavento en comparación con individuos conespecíficos que creen en los núcleos húmedos de los fragmentos (véase capitulo 1). Para entender la habilidad de Aextoxicon punctatum para sobrevivir a través del gradiente de humedad del suelo, primero se estudió la magnitud y variabilidad de los rasgos foliares (área foliar específica, densidad de estomas y de tricomas) e hidráulicos (diámetro y densidad de vasos conductores y conductividad hidráulica específica de la madera) relacionados con la tolerancia a la sequía a través de las tres zonas en los parches (ver capítulo 2); y segundo se estudió la integración fenotípica entre los rasgos.
(16) 7 funcionales evaluados en cada una de las zonas de humedad del suelo de los parches (ver capítulo 2). Esta tesis revela mecanismos fisiológicos clave que ayudan a explicar la distribución contrastante de las especies arbóreas a través de zonas y parches en el Parque Nacional Fray Jorge, y aporta información útil para intentar predecir la respuesta de estas especies a futuros cambios globales como la fragmentación y el cambio climático.. Área de estudio El área de estudio está ubicada en el Parque Nacional Fray Jorge, localizado en la región de Coquimbo (Chile) (30°40´S, 71°35´W) (Figura 1). El clima es Mediterráneo árido caracterizado por veranos cálidos y secos e inviernos húmedos y fríos (Di Castri y Hajek 1976). La temperatura promedio es de 13.6°C y la precipitación promedio es de 147 mm concentradas en los meses de Junio a Agosto (López-Cortez y López 2004). Durante los meses de Octubre a Enero hay mayor incidencia de la niebla costera, la cual puede aportar anualmente alrededor de 250 mm adicionales a las precipitaciones (del Val et al. 2006). Esta neblina está asociada con el agua fría generada por la corriente de Humboldt e inversión producidos por la subsidencia Anticiclón Pacífico Sur (Cereceda et al. 2002). Los fragmentos de bosque de neblina varían entre 0,1 y 36 ha (Barbosa et al. 2010) y se encuentran rodeados por una matriz de vegetación xerofítica y cactáceas. Están localizados entre los 400 y 600 m de altitud, representando el límite norte de distribución del bosque templado dominado por Aextoxicon punctatum, el cual tiene una distribución continua cerca de 1000 km hacia el sur del país (37°-43°S) (Smith-Ramírez et al. 2005). Florísticamente los fragmentos de bosque están dominados en su estrato arbóreo por Aextoxicon punctatum, género monotípico de una familia endémica de los bosques templados de Sudamérica (Aextoxicaceae), y otras especies como Myrceugenia correifolia (Myrtaceae), Rhaphithamnus spinosus (Verbenaceae), Drymis winteri (Winteraceae) y Azara microphylla (Flacourtiaceae) (Villagrán et al. 2004). Tienen importantes trepadoras y epífitas leñosas y herbáceas como Griselinia scandens (Griseliniaceae), Sarmienta repens (Gesneriaceae) y Mitraria coccinea (Gesneriaceae) (Villagrán et al. 2004), incluyendo helechos como Polypodium e Hymenophyllum..
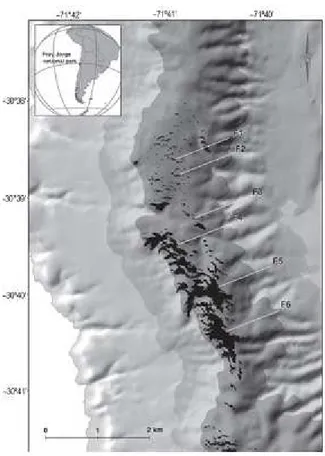
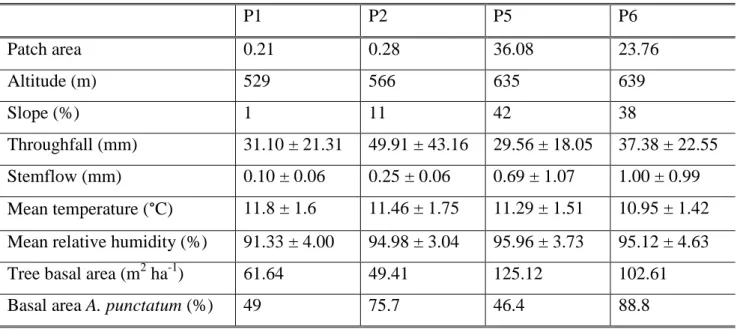
(17) 8. Figura 1. Ubicación de los fragmentos de bosque dependientes de neblina en el Parque Nacional Fray Jorge a los 30°S (Barbosa et al. 2010).. Para esta investigación se seleccionaron cuatro fragmentos: dos fragmentos pequeños (<0.5 ha) y dos fragmentos grandes (> 20 ha) (Tabla 1). Los fragmentos tienen similar exposición y edad (Gutiérrez et al. 2008), están separados por una distancia mínima de 400 metros entre sí y no están afectados por la presencia de otros fragmentos que pudieran alterar la captura de neblina (Barbosa 2005)..
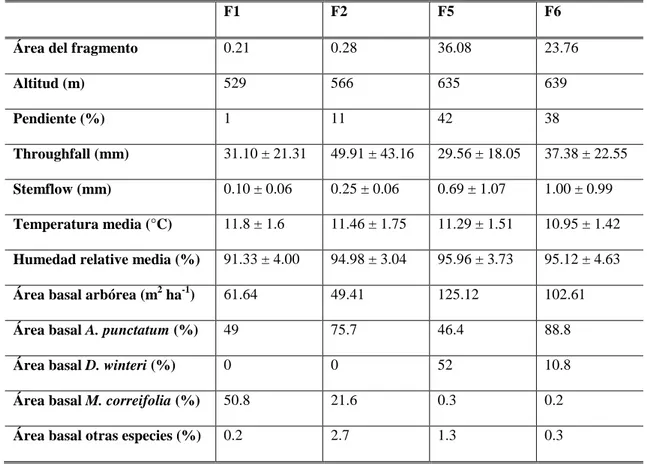
(18) 9 Tabla 1. Caracterización de los fragmentos de bosque, valores medios de las variables microclimáticas y área basal relativa para los individuos vivos (>5 cm dap) para los cuatro fragmentos estudiados (Gutiérrez et al. 2008, Barbosa et al. 2010). F1. F2. F5. F6. Área del fragmento. 0.21. 0.28. 36.08. 23.76. Altitud (m). 529. 566. 635. 639. Pendiente (%). 1. 11. 42. 38. Throughfall (mm). 31.10 ± 21.31. 49.91 ± 43.16. 29.56 ± 18.05. 37.38 ± 22.55. Stemflow (mm). 0.10 ± 0.06. 0.25 ± 0.06. 0.69 ± 1.07. 1.00 ± 0.99. Temperatura media (°C). 11.8 ± 1.6. 11.46 ± 1.75. 11.29 ± 1.51. 10.95 ± 1.42. Humedad relative media (%). 91.33 ± 4.00. 94.98 ± 3.04. 95.96 ± 3.73. 95.12 ± 4.63. Área basal arbórea (m2 ha-1). 61.64. 49.41. 125.12. 102.61. Área basal A. punctatum (%). 49. 75.7. 46.4. 88.8. Área basal D. winteri (%). 0. 0. 52. 10.8. Área basal M. correifolia (%). 50.8. 21.6. 0.3. 0.2. Área basal otras especies (%). 0.2. 2.7. 1.3. 0.3. Visión general Uno de los objetivos de la ecología es entender los procesos que estructuran las comunidades naturales, donde los estudios a través de gradientes ambientales han tenido gran relevancia. En las comunidades forestales de las regiones áridas del mundo, la disponibilidad de agua es uno de los principales factores que determina la distribución de las especies, y los patrones observados han sido frecuentemente atribuidos a las diferencias entre especies en sus tolerancias a la sequía y habilidades para competir por agua. Entender cómo los rasgos funcionales relacionados a la tolerancia a la sequía varían a través de gradientes a pequeña escala es importante para predecir la respuesta de las especies a futuros cambios climáticos. En esta tesis se estudiaron los mecanismos fisiológicos que explican los patrones contrastantes de distribución observados a través de gradientes de humedad generados por la neblina costera en las tres principales especies arbóreas Aextoxicon punctatum, Drimys winteri y Myrceugenia correifolia que coexisten en los fragmentos de bosque del Parque Nacional Fray Jorge, en la región semiárida en Chile. Se encontró un.
(19) 10 continuo de estrategias en el uso de agua que permitieron explicar la distribución de las especies a través del gradiente de humedad: en un extremo la especie Drimys winteri, con rasgos favoreciendo la eficiencia hidráulica y fotosintética; mientras que en el extremo opuesto la especie Myrceugenia correifolia, con rasgos favoreciendo la conservación del agua y reduciendo el riesgo a la cavitación. La especie con amplia distribución Aextoxicon punctatum, mostró valores de rasgos intermedios, con variación en los rasgos foliares y ausencia de variación en la anatomía del xilema a través de las zonas dentro de los fragmentos. En Aextoxicon punctatum se detectaron mayores niveles de integración fenotípica y variabilidad en los bordes secos..
(20) 11. Referencias Ackerly DD, Cornwell WK. 2007. A trait-based approach to community assembly: partitioning of species trait values into within- and among-community components. Ecology Letters 10: 135-145. Albert CH, Thuiller W, Yoccoz NG, Soudant A, Boucher F, Saccone P, Lavorel S. 2010a. Intraspecific functional variability: extent, structure and sources of variation. Journal of Ecology 98: 604-613. Albert CH, Thuiller W, Yoccoz NG, Douzet R, Aubert S, Lavorel S. 2010b A multitrait approach reveals the structure and the relative importance of intra- vs. interspecific variability in plant traits. Functional Ecology 24: 1192-1201. Baldini E, Facini O, Nerozzi F, Rossi F, Rotondi A. 1997. Leaf characteristics and optical properties of different woody species. - Trees 12: 73-81. Baraloto C, Paine CET, Poorter L, Beauchene J, Bonal D, Domenach AM, Herault B, Patiño S, Roggy JC, Chave J. 2010. Decoupled leaf and stem economics in rain forest trees. Ecology Letters 13: 1338-1347. Barbosa O, Marquet PA, Bacigalupe LD, Christie DA, del-Val E, Gutiérrez AG, Jones CG, Weathers KC, Armesto JJ. 2010. Interactions among patch area, forest structure and water fluxes in a fog-inundated forest ecosystem in semi-arid Chile. Functional Ecology 24: 909-917. Bongers F, Poorter L, Van Rompaey RSAR, Parren MPE (1999) Distribution of twelve moist forest canopy tree species in Liberia and Cote d’Ivoire: response curves to a climatic gradient. Journal of Vegetation Science 10: 371–382 Brodribb TJ, Feild TS. 2000. Stem hydraulic supply is linked to leaf photosynthetic capacity: evidence from New Caledonian and Tasmanian rainforests. Plant, Cell and Environment 23: 1381-1388. Brodribb TJ, Holbrook NM, Gutiérrez MV. 2002. Hydraulic and photosynthetic coordination in seasonally dry tropical forest trees. Plant, Cell and Environment 25: 1435–1444. Cereceda P, Osses P, Larraín H, Farías M, Lagos M, Pinto R, Schemenauer RS (2002) Advective, orographic and radiation fog in the Tarapacá region, Chile. Atmosphere Research 64: 261–271. Chave J, Coomes D, Jansen S, Lewis SL, Swenson NG, Zanne AE. 2009. Towards a worldwide wood economics spectrum. Ecology Letters 12: 351–366. Choat B, Marilyn CB, Luly JG, Holtum JAM. 2005. Hydraulic architecture of deciduous and evergreen dry rainforest tree species from north-eastern Australia. Trees-Struct Funct 19:305-311. Choat B, Jansen S, Brodribb TJ, Cochard H, Delzon S, Bhaskar R, Bucci SJ, Field TS, Gleason SM, Hacke UG, Jacobsen AL, Lens F, Maherali H, Martínez-Vilalta J, Mayr S, Mencuccini M, Mitchell PJ, Nardini A, Pittermann J, Pratt RB, Sperry JS, Westoby M, Wright IJ, Zanne AE. 2012. Global convergence in the vulnerability of forests to drought. Science 491: 752-756. Condit R. 1998. Ecological implications of changes in drought patterns: shift in forest composition in Panama. Climate Change 39: 413–427..
(21) 12 Condit R, Pitman N, Leigh EG Jr, Chave J, Terborgh J, Foster RB, Nuñez-V P, Aguilar S, Valencia R, Villa G, Muller-Landau HC, Losos E, Hubbell SP. 2002. Betadiversity in tropical forest trees. Science 295: 666–669. Cornwell WK, Ackerly DD. 2009. Community assembly and shifts in plant trait distributions across an environmental gradient in coastal California. Ecological Monographs 79: 109-126. Dawson TE. 1998. Fog in the California redwood forest: ecosystem inputs and use by plants. Oecologia 117: 476–485. Díaz S, Hodgson JG, Thompson K, Cabido M, Cornelissen JHC, Jalili A, MontserratMartí G, Grime JP, Zarrinkamar F, Asri Y, Band SR, BasconceloS1, CastroDíez P, Funes G, Hamzehee B, Khoshnevi M, Pérez-Harguindeguy N, PérezRontomé MC, Shirvany FA, Vendramini F, Yazdani S, Abbas-Azimi R, Bogaard A, Boustani S, Charles M, Dehghan M, de Torres-Espuny L, FalczukV, Guerrero-Campo J, Hynd A, Jones G, Kowsary E, Kazemi-Saeed F, MaestroMartínez M, Romo-Díez A, Shaw S, Siavash B, Villar-Salvador P, Zak MR. 2004. The plant traits that drive ecosystems: Evidence from three continents. Journal of Vegetation Science 15: 295-304. del-Val E, Armesto JJ, Barbosa O, Christie DA, Gutie´rrez AG, Jones CG, Marquet PA, Weathers KC. 2006. Rain forest islands in the Chilean semiarid region: fogdependency, ecosystem persistence and tree regeneration. Ecosystems 9: 598– 608. Di Castri F, Hajek E. 1976. Bioclimatología de Chile. Universidad Católica de Chile, Santiago Engelbrecht BMJ, Comita LS, Condit R, Kursar TA, Tyree MT, Turner BL, Hubbell SP. 2007. Drought sensitivity shapes species distribution patterns in tropical forests. Nature 447: 80–83. Ewing HA, Weathers KC, Templer PH, Dawson TE, Firestone MK, Elliott AM, Boukili VKS. 2009. Fog water and ecosystem function: heterogeneity in a California redwood forest. Ecosystems 12: 417–433. Fahn, A. 1986. Structural and functional properties of trichomes of xeromorphic leaves. Annals of Botany 57: 631-637 Gentry AH. 1988. Changes in plant community diversity and floristic composition on environmental and geographic gradients. Annals of Missouri Botanical Garden 75:1–34. Gutiérrez AG, Barbosa O, Christie DA, del-Val E, Ewing HA, Jones CG, Marquet PA, Weathers KC, Armesto JJ. 2008. Regeneration patterns and persistence of the fog dependent Fray Jorge forest in semiarid Chile during the past two centuries. Global Change Biology 14: 161–176. Hacke UG, Sperry JS, Pockman WT, Davis SD, McCulloh K. 2001. Trends in wood density and structure are linked to prevention of xylem implosion by negative pressure. Oecologia 126: 457-461. Hildebrandt A, Eltahir EAB. 2006. Forest on the edge: seasonal cloud forest in Oman creates its own ecological niche. Geophysical Research Letter 33: L11401..
(22) 13 Hulshof CM, Swenson NG. 2010. Variation in leaf functional trait values within and across individuals and species: an example from a Costa Rican dry forest. Functional Ecology 24: 217–223. Katata G, Nagai H, Kajino M, Ueda H, Hozumi Y. 2010. Numerical study of fog deposition on vegetation for atmosphere–land interactions in semi-arid and arid regions. Agricicultural Forest Meteoroly 150: 340–353 López-Cortés F, López D. 2004. Antecedentes bioclimáticos del Parque Nacional Bosque Fray Jorge. In: Squeo FA, Gutiérrez JR, Hernández IR (eds) Historia natural del parque nacional bosque Fray Jorge. Ediciones Universidad de La Serena, Chile, pp 45–60. Markesteijn L, Poorter L, Bongers F, Paz H, Sack L. 2011a. Hydraulics and life history of tropical dry forest tree species: coordination of species’ drought and shade tolerance. New Phytology 191:480-495. Markesteijn L, Poorter L, Paz H, Sack L, Bongers F. 2011b. Ecological differentiation in xylem cavitation resistance is associated with stem and leaf structural traits. Plant Cell Environ 34:137-148. Meinzer FC, Woodruff DR, Domec JC, Goldstein G, Campanello PI, Gatti MG, Villalobos-Vega R. 2008. Coordination of leaf and stem water transport properties in tropical forest trees. Oecologia 156: 31–41. Nicotra AB, Chazdon RL, Schlichting DS. 1997. Patterns of genotypic variation and phenotypic plasticity of light response in two tropical Piper (Piperaceae) species. American Journal of Botany 84: 1542-1552. Niinemets, U. 2001. Global-scale climatic controls of leaf dry mass per area, density and thickness in trees and shrub. Ecology 82: 453-469. Pigliucci M, Kolodynska A. 2003. Phenotypic plasticity and integration in resonse to flooded conditions in natural accessions of Arabidopsis thaliana (L.) Heynh (Brassicaceae). Annals of Botany 90: 199-207. Poockman WT, Sperry JS. 2000. Vulnerability to xylem cavitation and the distribution of Sonoran Desert. American Journal of Botany 87: 1287-1299. Poorter L. 2007. Are species adapted to their regeneration niche, adult niche, or both? Am Nat 169: 433–442 Pyke CR, Condit R, Aguilar S, Lao S. 2001. Floristic composition across a climatic gradient in a neotropical lowland forest. Journal of Vegetation Science 12: 553– 566. Reich PB, Wright IJ, Cavender-Bares J, Craine JM, Oleksyn J, Westoby M, Walters MB. 2003. The evolution of plant functional variation: traits, spectra, and strategies. International Journal of Plant Sciences 164: S143–S164. Salgado-Negret B, Pérez F, Markesteijn L, Jiménez-Castillo M, Armesto JJ. 2013. Diverging drought-tolerance strategies explain tree species distribution along a fog-dependent moisture gradient in a temperate rain forest. Oecologia. DOI 10.1007/s00442-013-2650-7 Santiago LS, Goldstein G, Meinzer FC, Fisher JB, Machado K, Woodruff D, Jones T. 2004. Leaf photosynthetic traits scale with hydraulic conductivity and wood density in Panamanian forest canopy trees. Oecologia 140: 543-550..
(23) 14 Sardans J, Peñuelas J, Rodá F. 2006. Plasticity of leaf morphological traits, leaf nutrient content, and water capture in the Mediterranean evergreen oak Quercus ilex subsp. ballota in response to fertilization and changes in competitive conditions. Ecoscience 13: 258-270. Smith-Ramírez C, Rovere AE, Núñez-Avila MC, Armesto JJ. 2007. Habitat fragmentation and reproductive ecology of Embothirum coccineum, Eucryphia cordifolia and Aextoxicon punctatum in southern temperate rainforests. In: (A.C. Newton, ed), “Biodiversity Loss and Conservation in Fragmented Forest Landscapes: The Forests of Montane Mexico and Temperate South America", pp. 102-119. Sterck F, Markesteijn L, Schieving F, Poorter L. 2011. Functional traits determine trade-offs and niches in a tropical forest community. Proceedings of the National Academy of Scienes 108: 20627-20632. Villagrán C, Armesto JJ, Hinojosa LF, Cuvertino J, Pérez C, Medina C. 2004. El enigmático origen del bosque relicto de Fray Jorge. In ‘Historia natural del parque nacional bosque Fray Jorge’. (Eds FA Squeo, JR Gutiérrez, IR Hernández) pp. 3–43. Weathers KC, Lovett GM, Likens GE, Caraco NFM. 2000. Cloud-water inputs of nitrogen to forest ecosystems in southern Chile: forms, fluxes and sources. Ecosystems 3: 590–595. Wright SJ. 1992. Seasonal drought, soil fertility and the species density of tropical forest plant communities. Trends in Ecology and Evolution 7: 260–263. Wright IJ, Reich PB, Westoby M, Ackerly DD, Baruch Z, Bongers F, Cavender-Bares J, Chapin T, Cornelissen JH, Diemer M, Flexas J, Garnier E, Groom PK, Gulias J, Hikosaka K, Lamont BB, Lee T, Lee W, Lusk C, Midgley JJ, Navas ML, Niinemets U, Oleksyn J, Osada N, Poorter H, Poot P, Prior L, Pyankov VI, Roumet C, Thomas SC, Tjoelklaas MG, Veneklaas EJ, Villar R. 2004. The worldwide leaf economics spectrum. Nature 428: 821-827. Wright IJ, Falster DS, Pickup M, Westoby M. 2006. Cross-species patterns in the coordination between leaf and stem traits, and their implications for plants hydraulics. Physiologia Plantarum 127: 445-456. Zanne AE, Westoby M, Falster DS, Ackerly DD, Loarie SR, Arnold SEJ, Coomes DA. 2010. Angiosperm wood structure: global patterns in vessel anatomy and their relation to wood density and potential conductivity. American Journal of Botany 97: 207-215..
(24) 15. CAPÍTULO I. II.. Estrategias divergentes de tolerancia a la. sequía explican la distribución de especies arbóreas a través de un gradiente de humedad dependiente de neblina en un bosque lluvioso templado Salgado-Negret B, Pérez F, Markesteijn L, Jimenez-Castillo M, Armesto JJ. 2013. Diverging drought tolerance strategies explain tree species distribution along a fogdependent moisture gradient in a temperate rain forest. Oecologia DOI 10.1007/s00442013-2650-7.
(25) 16 Diverging drought tolerance strategies explain tree species distribution along a fogdependent moisture gradient in a temperate rain forest Beatriz Salgado Negret1,2,*, Fernanda Pérez1,2, Lars Markesteijn3, Mylthon Jiménez Castillo4,5, Juan J. Armesto1,2 1. Departamento de Ecología, Pontificia Universidad Católica de Chile, Casilla 114-D,. Santiago, Chile; 2Instituto de Ecología y Biodiversidad, Casilla 653, Santiago, Chile; 3. Departamento de Biogeografía y Cambio Global, Museo Nacional de Ciencias Naturales,. Consejo Superior de Investigaciones Científicas (CSIC), Serrano 115 dpdo, E-28006, Madrid, Spain; 4Instituto de Ciencias Ambientales y Evolutivas, Universidad Austral de Chile, Casilla 567, Valdivia-Chile; 5Jardín Botánico Universidad Austral de Chile, Facultad de Ciencias, Universidad Austral de Chile, Casilla 567, Valdivia-Chile.. *Author for correspondence: Beatriz Salgado Negret [email protected] 56-2-3542637.
(26) 17. Abstract The study of functional traits and physiological mechanisms determining species´ drought tolerance is important to predict their responses to climatic change. Fog-dependent forest patches in semiarid regions are a good study system to understand species responses to increasing aridity and patch fragmentation. Here we measured leaf and hydraulic traits for three dominant species with contrasting distributions within patches in relict, fog-dependent forests in semiarid Chile. In addition, we assessed pressure-volume curve parameters in trees growing at dry leeward edge and wet patch core. We predicted species would display contrasting suites of traits according to local water availability: from one end favoring water conservation and reducing cavitation risk, to opposite end favoring photosynthetic and hydraulic efficiency. Consistent with our hypothesis, we identified a continuum of water use strategies explaining species distribution along small-scale moisture gradient. Drimys winteri, a tree restricted to the humid core, showed traits allowing efficient water transport and high carbon gain; in contrast, Myrceugenia correifolia, a tree that occurs in the drier patch edges, exhibited traits promoting water conservation and lower gas exchange rates, as well low water potential at turgor loss point. The most widespread species, Aextoxicon punctatum, showed intermediate trait values. Osmotic compensatory mechanism was detected in M. correifolia, but not in A. punctatum. We show that partitioning of the pronounced soil moisture gradients from patch cores to leeward edges among tree species is driven by differential drought tolerance. Such differences indicate that trees have contrasting abilities to cope with future reductions in soil moisture.. Keywords Climate change, fog-dependent forest, local water gradient, species distribution, plant hydraulic traits..
(27) 18. Introduction Water availability is a major factor influencing species distribution in forest communities across large-scale rainfall gradients as well as small-scale topographic gradients (Gentry 1988; Wright 1992; Condit 1998; Bongers et al. 1999; Pyke et al. 2001; Condit et al. 2002; Engelbrecht et al. 2007). Species’ distribution may be explained by functional trait divergence associated with performance under particular conditions of soil humidity (Poorter 2007; Markesteijn et al. 2011a; Sterck et al. 2011). Understanding the bases of such differentiation among forest trees may be critical for predicting the ecological consequences of future alteration of soil moisture gradients due to climate change. Fog-dependent forests, found in semiarid regions of the world (Hildebrandt and Eltahir 2006; del-Val et al. 2006; Katata et al. 2010), are thought to be relicts from past periods when conditions were more humid, and thus these ecosystems might be especially sensitive to current changes in fog water supply. Alterations in fog frequency and intensity are predicted to occur due to changes in sea-surface temperature and the height of the temperature inversion layer (Cereceda et al. 2002), loss of forest patch area and fragmentation, or changes in forest structure affecting fog capture (Hildebrandt and Eltahir 2006). In these patchy forests, fog interception by plants is the primary or even the only source of water during most of the year (Dawson 1998; del-Val et al. 2006; Ewing et al. 2009). The fog interception by trees creates pronounced water and nutrient gradients from windward to leeward edges in forest patches (Weathers et al. 2000; del-Val et al. 2006; Ewing et al. 2009), with strong contrasts over short distances, depending on wind direction (Ewing et al. 2009). Studying tree species responses to soil moisture variation at short spatial scales, due to topographic and/or patch fragmentation gradients in these fogdependent ecosystems, allows us to address questions about the critical conditions for sustaining tree species under increasing drought stress due to changing climate. Our study site, the Fray Jorge forest in central Chile, is a striking example of such a fog dependent ecosystem, where the strong water (and possibly nutrient) gradients inside the isolated forest patches affect the distribution and regeneration dynamics of tree species (del-Val et al. 2006). The patches are dominated by species characteristic of temperate and Mediterranean forests in Chile: Aextoxicon punctatum (in the monotypic family.
(28) 19 Aextoxicaceae) is found in all-size patches but it is more frequent in humid windward edges, directly facing the incoming fog; Drimys winteri (Winteraceae) tends to be aggregated in the interior of the largest forest patches and is not found in small patches; finally, Myrceugenia correifolia (Myrtaceae) is more common along the edges of small patches, including the drier leeward edge (del-Val et al. 2006; Gutiérrez et al. 2008). Such contrasting distribution patterns, and the pronounced short-distance, environmental gradients related to moisture supply by fog, offer a great opportunity to investigate the physiological mechanisms that explain tree species ability to respond to abrupt and pronounced changes in climate due global warming. Convergence in leaf traits reducing water loss by transpiration, as well as hydraulic traits favouring safety at the expense of hydraulic efficiency, has been reported for plants that are periodically exposed to severe water deficit (Mitchell et al. 2008; Markesteijn et al. 2011a,b). Such plants usually show narrower and shorter vessels with small pit pores, which are more resistant to drought-induced cavitation, but at the same time have an increased flow resistance and a lower hydraulic efficiency (Hacke et al. 2001; Choat et al. 2005; Mitchell et al. 2008; Markesteijn et al. 2011a,b), affecting leaf water supply. The capacity to maintain leaf turgor in response to decreasing soil moisture availability is also an important mechanism that favours drought tolerance (Kozlowski and Pallardy 2002; Baltzer et al. 2008; Kursar et al. 2009; Bartlett et al. 2012). Water potential at loss turgor point (πtlp) is a critical physiological determinant of a plant’s tolerance to water stress (Bartlett et al. 2012). Plants can reduce πtlp by accumulating osmotically active compounds in the cells (osmotic adjustment) or by increased cell wall flexibility (elasticity, ε). However, recently Bartlett et al. (2012) showed no direct role for ɛ in driving differences in πtlp across species, instead, elastic adjustments acted to maintain relative water content at turgor loss point (RWCtlp) despite very negative water potentials at full turgor (π0) and πtlp. Here, we measured leaf and hydraulic traits of the three main tree species occurring in fog-inundated rain forest patches of Fray Jorge (semiarid Chile), which show contrasting distribution patterns along the soil moisture gradient produced by fog influx. We also compared pressure-volume curves traits of individuals growing at windward and leeward edges of forest patches..
(29) 20 Specifically, we addressed the following questions: 1) How does the variation in functional traits related to drought tolerance explain species distribution along small-scale moisture gradients? 2) What mechanisms allow individuals growing along the drier leeward edges to cope with reduced water availability (such as osmotic adjustment or increased cell elasticity) in comparison with conspecific individuals growing in wetter patch core habitats? We expect that species growing in small patches and leeward patch edges would display a suite of leaf traits favoring water conservation (such a reduced stomatal conductance) and a suite of hydraulic traits reducing cavitation risk (such as narrow vessels), at the expense of photosynthesis and hydraulic efficiency. We also predict that individuals growing at leeward patch edge would have pressure-volume traits values favoring drought tolerance (such as lower πtlp and π0) in comparison with conspecific individuals growing in wetter patch core. Tree species occurring in the fog forest of Fray Jorge are exposed to increased aridity due to climatic changes over an extended period of time (Villagrán et al. 2004; Gutiérrez et al. 2008), facing seasonal changes in fog influx that drive pronounced moisture gradients within patches (del-Val et al. 2006). This study aims to reveal some of the basic mechanisms underlying the relative success of these species to coexist given past and current variations in moisture availability. Here, we will further discuss results in the light of the possible consequences of future climate change and its effects on species’ distribution and coexistence.. Materials and methods Study site and species Fray Jorge National Park (30°40´S. 71°30´W) comprises the northernmost patches of Chilean temperate rainforests, dominated by broad-leaved evergreen tree species, which exhibit remarkable floristic affinities with temperate forests located some 1000 km to the south (Villagrán et al. 2004). The area contains a mosaic of about 180 forest patches ranging in size from 0.1 to 36 ha, located on the summits of coastal mountains at an elevation of 450 to 660 m, surrounded by a matrix of semiarid scrub vegetation (Barbosa et.
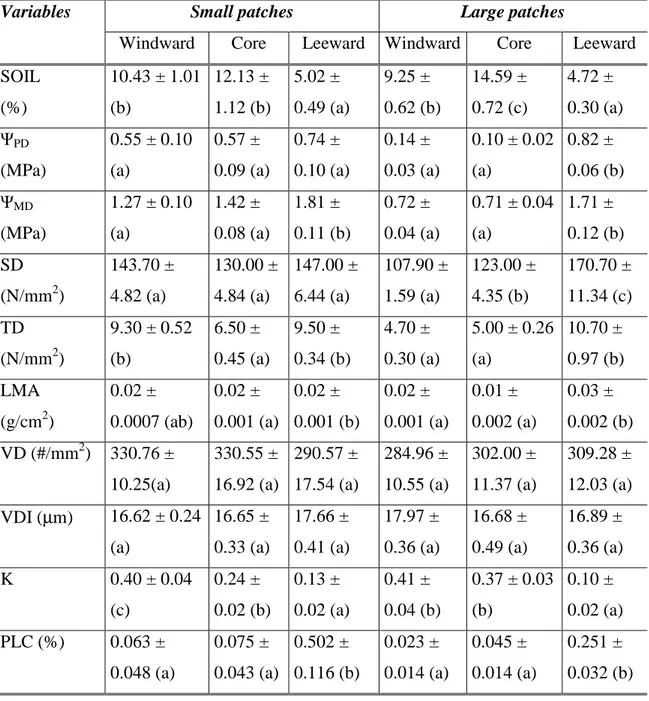
(30) 21 al. 2010). The regional climate is Mediterranean-arid with a mean annual rainfall of 147 mm concentrated during the cool winter months from May to August and a mean annual temperature of 13.6°C (López-Cortés and López 2004). Fog is a prominent and constant feature of the landscape above 400 m elevation especially during spring and summer months, when fragments can receive an additional input of at least 200 mm of cloud water annually via throughfall and stemflow (del-Val et al. 2006). A large 36 ha patch was selected for this study because it was the only one where all three focal tree species coexist. Additional details on the structure and physical gradients of patches are given by Barbosa et al. (2010). The forest patch studied was located at an altitude of 635 m, with average air temperatures inside the patch varying from 9.2°C in spring (October to December) to 13.3°C in winter (July to September) and relative air humidity varying between 83.6% in winter and 99.6% in spring-summer. The forest canopy is dominated by A. punctatum (Aextoxicaceae), with juveniles occurring more frequently along the edge directly receiving fog influx (windward), but adults found throughout patch, and co-dominated by D. winteri (Winteraceae), which tends to be aggregated inside the patch. M. correifolia (Myrtaceae) is occasionally represented in the canopy of the forest patch (0.3% basal area) but it is confined to the drier leeward edge (Gutiérrez et al. 2008). Volumetric soil moisture varies substantially in both small and large patches. Leeward edges are drier than the other two microhabitats, while soil moisture at the windward edges is comparable with patch core (25 measurements per zone in A. punctatum individuals): small patches; windward: 10.43% ± 1.01; core: 12.13% ± 1.12; leeward: 5.02% ± 0.49 and large patches; windward: 9.25% ± 0.62; core: 14.59% ± 0.72; leeward: 4.72% ± 0.30) (Salgado-Negret unpublished data). Volumetric soil moisture for our species measured at 20 cm depth, varied accordingly across sites occupied by the different tree species (30 measurements per species): D. winteri (22.9% ± 2.66), A. punctatum (13.4% ± 1.7) and M. correifolia (5.3% ± 0.53) (p<0.0001; F=23.01; d.f.=2) (Salgado-Negret unpublished data). The three species have a different phytoclimatic distribution in Chile: A. punctatum is a tree species endemic of western South America and it is broadly distributed in coastal forests from 30 - 43°S; D. winteri is distributed from Fray Jorge and central Chile to Sub-.
(31) 22 Antarctic forest in Tierra del Fuego at 55°S (Villagrán et al. 2004). Finally, M. correifolia is restricted to central Chile with a Mediterranean climate subjected to a cool rainy winter and a summer drought period of 2– 3 months (Di Castri and Hajek 1976).. Leaf traits We measured leaf traits for six individuals (dbh >10 cm) of each tree species using mature, fully expanded leaves without herbivore damage. All measurements were done on the same six individuals. CO2 assimilation curves were constructed using the CO2 reference concentration of 380 ppm, 50% relative humidity, and a temperature of 25° C. Photosynthesis (AMAX) and stomatal conductance (gs) were measured in M. correfolia, A. punctatum and D. winteri at 700, 500 and 700 umol m-2 s-1 respectively, with an open portable photosynthesis system (CIRAS-2 CRS068, PP Systems, Amesbury, USA) equipped with a LED light. Measurements were conducted between 10:00 and 13:00 h. After measurements of gas exchange, leaves were cut and leaf water potentials at midday were measured (ψMD, MPa) using a pressure chamber (Scholander-type, Model 1000 PMS). We also measured predawn leaf water potentials (ψPD, MPa) between 5:00 and 7:00 h for the same six individuals per species. After measurement, leaves were scanned (EPSON Stylus TX200) and analysed using ImageJ software (http://imagej.nih.gov/ij/) to determine leaf area (LA). Finally, leaves were dried for 48h at 65°C to obtain leaf dry mass (g) and calculate leaf mass per area (LMA; g cm-2) (Cornelissen et al. 2003).. Pressure-volume curves Pressure-volume curves were constructed for six individuals per species. One shoot was cut from each individual and the shoots were hydrated with distilled water in plastic bags to bring leaves to full turgor. Tissue rehydration is necessary to ensure that all samples are near saturation thus allowing for construction of the entire moisture release curve (Baltzer et al. 2008). After 24h of rehydration, we constructed pressure-volume curves following the Sack and Pasquet-Kok protocol (www.prometheuswiki.com). Water potentials of the leaves.
(32) 23 were measured with a Scholander-type pressure chamber (PMS, Model 1000) and the tissue was weighed immediately after measurement. The tissue was dehydrated slightly at room temperature, before re-weighing the leaf mass and re-measuring the water potential. This process was repeated until the tissue reached constant mass. When there was no further decrease in mass, leaves were dried for 48h at 80° C to determine dry mass. The following traits were estimated from the pressure-volume curves: solute potential at full turgor (π0; MPa), solute potential at turgor loss point (πtlp; MPa), relative water content at turgor loss point (RWCtlp; %), and the bulk modulus of elasticity (ɛ; MPa).. Hydraulic traits Maximum vessel length - One branch (2.5 – 10 mm diameter) was cut from the outer crown of each of six individuals per species and transported to the field station. Here, maximum vessel lengths were estimated cutting branches approximately 1 m from the distal apex and applying air pressure (approx. 60 Kpa) (cf. Ewers and Fisher 1989) to the cut end of the branch. The distal end of the branch was then trimmed back approximately 1 cm at a time until air bubbles were seen emerging from vessel ends (Brodribb and Feild 2000). The remaining branch length at this point was then measured as an estimate of Maximum vessel length (MVL; cm).. Sapwood-specific hydraulic conductivity - A second collection of branches was made from the same six individuals per species to measure hydraulic conductivity (water flux through a unit length of stem over a pressure gradient; Kh, in kg m-1 s-1 MPa-1) following Sperry et al. (1988). In the field station, branches were recut under water to avoid the induction of new embolisms. Distal ends were trimmed with a razor blade to clear any accidentally blocked vessels and about 1 cm of the bark at each side of the branch was removed. While submerged, the shaved end of the branch was wrapped in Parafilm. All branches used for hydraulic conductivity measurements were cut to the same length (approx. 30 cm). The branch was connected to a fluid column fed by a reservoir elevated to a height of 1 m, providing a constant pressure of 9.8 KPa. An electronic balance registered KCl solution flux as an increase in sample mass each 30 seconds. Measurements were taken when an.
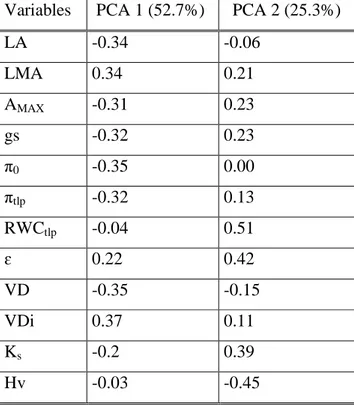
(33) 24 approximately constant flow was observed for at least 3 min. Afterwards, the stems were flushed with KCl solution at a pressure of ≈170 KPa for 10-15 minutes to remove emboli (Sperry et al. 1987) and hydraulic conductivity was measured again at its maximum capacity. We divided Kh by the cross-sectional area of the conductive xylem (see methods Hydraulic anatomy below), to standardise the flow of water per unit sapwood area and obtain sapwood specific hydraulic conductivity (Ks; kg MPa-1 m-1 s-1). As such, hydraulic conductivity was made comparable among segments of different diameters.. Hydraulic anatomy. The same stems were then perfused with safranin dye to visualize the conductive wood area. A cross-sectional area of the upper distal end of the stem was photographed with a digital camera mounted on a microscope, at 10x magnificacion and the image was processed using the imaging software SigmaScan Pro 5 (SPSS Inc.) to determine vessel diameter (VD; µm) and density (VDi; vessels mm-2). For each branch, we calculated the Huber value (Hv; cm2 cm-2) as the cross-sectional sapwood area of the upper distal end of the stem divided by the total supported leaf area. Finally, for each species, vessel diameters were divided into 5 µm size classes to construct frequency histograms. In line with the Hagan-Poiseuille law, the vessel ratios in each size class were raised to the fourth power and summed to determine the relative contribution of each vessel size class to overall hydraulic conductance (Choat et al. 2005).. Data analysis Differences in leaf traits (LMA, LA, gs and AMAX), hydraulic traits (vessel diameter and density, Ks and Hv), and traits derived from pressure-volume curves (π0, πtlp, RWCtlp and ɛ) were contrasted among three tree species using a multivariate analysis of variance (MANOVA). Because MANOVA showed significant species effects, we conducted a series of univariate ANOVAs followed by post-hoc Tukey´s tests to identify individual responses of each trait. Overall multivariate relations and trait differences among species were further explored using a principal components analysis (PCA). Differences in traits derived from pressure-volume curves between leeward and core zones from A. punctatum and M..
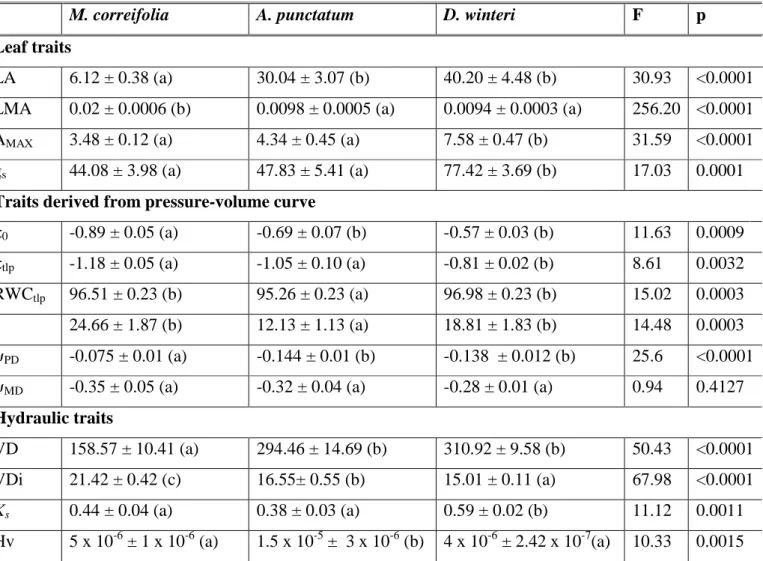
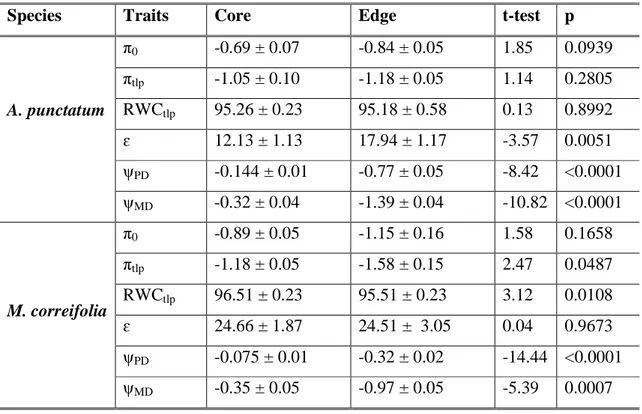
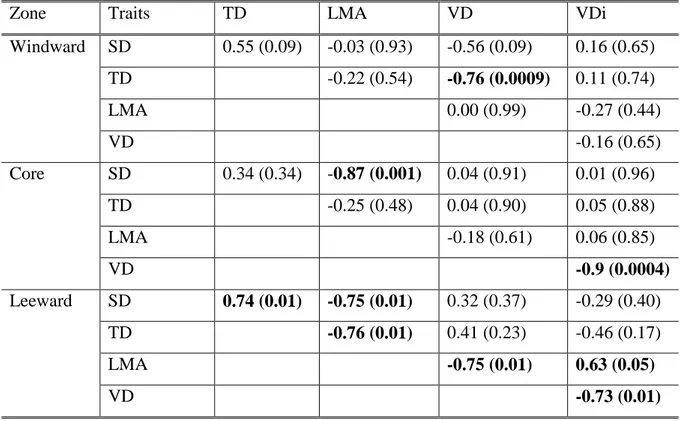
(34) 25 correifolia individuals were analysed with independent-samples t-tests. Statistical analyses were performed using InfoStat (Di Rienzo et al. 2011).. Results Species differences in leaf and hydraulic traits Leaf and hydraulic traits, as well pressure-volume curve related traits, differed substantially among the three coexisting tree species in Fray Jorge forest (MANOVA; Willk´s = 9.9 x E05; F = 33.11; p < 0.0001). Trait differentiation among species is best described by principal component analysis. The first component, which explained 53% of trait variation, showed an even contribution of variables with a magnitude of 0.3, and it clearly separated M. correifolia from D. winteri, placing A. punctatum at an intermediate position (Fig. 1). This component was negatively correlated with leaf traits that increased water transpiration and carbon gain (LA, gs, Amax), as well as with the solute potential at full turgor (π0) and the potential at turgor loss point (πtlp) (Table 1). Then, higher values along the first PCA component reflect stronger ability to conserve water and tolerate to drought, but lower gas exchange rates. PCA component 1 was also positively correlated with vessel diameter (VDi) and negatively correlated with vessel density (VD) (Table 1). The second component explained an additional 25.3% of the total variance and it separated A. punctatum from the other two species. This component was dominated by higher values of RWCtlp and lower values of Hv (Table 1). Significant differences in leaf traits among species were additionally detected using separate ANOVAs (Table 2). Accordingly, we found that D. winteri, a tree restricted to the moist cores of large patches, exhibited a higher stomatal conductance and photosynthetic rates than the other two species, although its average LMA did not differ from that of A. punctatum. In turn, we found that M. correifolia, a tree that occurs primarily in the drier leeward edges, had the smallest leaf area and lower stomatal conductance and photosynthetic rates. Finally, the most widespread tree species in these patches, A. punctatum, did not differ in stomatal conductance and photosynthetic rates from M. correifolia (Table 2, Fig. 2)..
(35) 26 Clear differences among the three species in traits derived from pressure-volume curves were also found (Table 2). The two species with more sclerophyllous leaves, A. punctatum and M. correifolia, showed the lowest πtlp and π0 values, and A. punctatum had the lowest RWCtlp (Table 2). The latter species also had the lowest ɛ, while values between the other two tree species did not differ. Predawn and midday leaf water potentials varied strongly among species (Table 2). In the summer season, presumably the warmer and drier period of the year, predawn leaf water potentials (ψPD) ranged from -0.075 MPa to -0.144 MPa, while midday water potential (ψMD) ranged from -0.28 to -0.35 MPa across the three species. Midday leaf water potentials never dropped below the turgor loss point, suggesting that species did not suffer from drought stress during the period of study. We found significant differences in hydraulic traits among tree species (Table 2). Hydraulic conductivity and vessel densities were higher and vessel diameters were smaller for D. winteri than for the other two species (Table 2, Fig 2). Contrary to our predictions, M. correifolia, the species that is most restricted to the semiarid Mediterranean-climate region, and presumably better adapted to summer drought, had larger vessel diameters than the other two species. A. punctatum, a predominantly coastal tree species, with a broad latitudinal distribution in Chilean forests and in the Fray Jorge forest patch mosaic, showed the lowest hydraulic conductivity, with intermediate vessel diameters and densities (Table 2, Fig. 2). According to the Hagan-Poiseuille law which states that in theory a vessel’s hydraulic conductance is proportional to the fourth power of its radius, D. winteri and A. punctatum hydraulic conductivity depended strongly on the lower vessel size classes (10 to 20 µm), 92.7% and 56.6% respectively (Fig. 3), while M. correifolia showed greater range of diameter classes and had 52% of its hydraulic conductivity accounted for by the wider vessel size class (20 to 30 µm) (Fig. 3).. Trait differences between patch core and leeward edge individuals We compared traits derived from pressure-volume curves between individuals growing in the patch core (away from edges) and in the leeward edge of the same patches; this comparison was only possible for A. punctatum and M. correifolia as these species co-.
(36) 27 occur in these two microhabitats. We did not have comparative data for D. winteri, because it was never found in patch edges. Most physiological traits obtained from the pressurevolume curves did not differ between A. punctatum trees in the core and leeward trees (Table 3), except for parameter ɛ. In the latter case, trees on the leeward edge of patches had a lower bulk modulus of elasticity than patch core trees. In contrast, M. correifolia showed clear differences in several attributes between trees sampled in the patch core and in the drier leeward edge. For this species, πtlp and RWCtlp values were lower at the leeward edge than at the patch core (Table 3). In the case of M. correifolia, ɛ did not vary between trees in the core and leeward edge of patches. Significant differences in ψPD and ψMD between trees in patch core and those in the leeward edge were found for both species, with the lowest values found for trees at the leeward edge (Table 3). In contrast to M. correifolia, for A. punctatum trees found at the leeward edge, ψMD dropped below πtlp.. Discussion Our results indicate that evergreen tree species were able to partition small-scale, but strong soil moisture gradients, fog-dependent forest patches, due to their differential ability to use soil water and tolerate drought-related habitat differences. For the three species dominating the canopy of fog-inundated patches in this semiarid region, we identified a continuous gradient of water-use strategies. Ecophysiological strategies varied between a set of plant traits that allows efficient water transport and high carbon gain, at the one end, to traits that enhance water conservation at the cost of lower gas exchange rates, at the opposite end. At one end of the continuum we find D. winteri, a tree species restricted to wet microhabitats in the core of large forest patches, which has high Ks, leaf area, photosynthetic rates and stomatal conductance. The opposite end of this gradient is occupied by M. correifolia, a species that is typically found in drier microhabitats of the leeward edges and in small forest patches, showing traits that imply increased drought tolerance, such as a small leaf area, reduced stomatal conductance and hydraulic conductivity, and low water potentials at turgor loss point. Finally, A. punctatum, the most abundant and widespread species in different microhabitats of Fray Jorge forest patches, displays intermediate values for the drought-tolerance traits investigated. The morphological and physiological differences.
(37) 28 detected among tree species in this ecosystem are likely to be important in shaping speciesspecific responses to future reductions in water availability as produced by reductions in fog frequency and rainfall, that are predicted for this and other semiarid regions in the coming decades (Johnstone and Dawson 2010). In this forest, D. winteri showed the broadest leaf area, highest photosynthetic rates and greatest stomatal conductance, which are associated with the highest KS. High conductivity contributes to a more efficient water supply to the leaves, supporting greater carbon assimilation (Meinzer et al. 1995; Sperry 2000; Brodribb and Feild 2000; Santiago et al. 2004). Still, in contrast with the former suite of traits, D. winteri had the smallest vessel diameters and the highest vessel density among species. D. winteri is an angiosperm, but belongs to the very primitive family Winteraceae, which does not have true vessels, but instead tissues that are very similar to the tracheids of coniferous species. Species with such vesselless wood are known to have up to 21 times lower inter-element pit resistance than eudicot vessels, and therefore their wood is highly conductive despite the short length and narrow diameter of tracheids (Hacke et al. 2006, 2007; Sperry et al. 2007). Despite its high Ks, large leaf area, and high stomatal conductance, D. winteri has a reduced ability to regulate water loss (Feild et al. 1998). Low stomatal control in D. winteri is probably associated with its hydrophobic granular plug, which consists of a porous, granular material that fills the stomatal cavity above the guard cells preventing them from fully closing (Feild et al. 1998; Feild and Holbrook 2000). This seems to be an adaptation to humid environments, where it precludes the formation of a permanent water film on the leaf surface that would obstruct CO2 diffusion into the leaf (Feild et al. 1998). Consequently, a reduced ability to regulate water loss in D. winteri implies a greater hydraulic demand that cannot be satisfied under the drier conditions that characterize small forest patches or patch edges in Fray Jorge. Species, such as D. winteri, will be more vulnerable to increased moisture stress at patch edges, as created by fragmentation. This will be further accentuated by the regional reductions in rainfall or fog inputs and will likely reduce the possibility that this species are able to maintain a viable population in the future. By contrast, M. correifolia, which is typically found in drier microhabitats in Fray Jorge, showed an opposite suite of traits compared to D. winteri, including smaller leaf areas, higher LMA, and a reduced stomatal and hydraulic conductance. The combination of.
Figure
+7

Outline
Documento similar
Relationships between plant water status and leaf structural traits with spectral indicators in Citrus species. Shaded boxes correspond to significant relationships according to
Three irrigation treatments were applied: control treatment (CTL) irrigated to ensure non-limiting soil water conditions; moderate water stress (MS) subjected to two drying cycles
For a short explanation of why the committee made these recommendations and how they might affect practice, see the rationale and impact section on identifying children and young
The expansionary monetary policy measures have had a negative impact on net interest margins both via the reduction in interest rates and –less powerfully- the flattening of the
Figure 5: Effect of the treatments applied on the chlorophyll index in the soybean plants grown in calcareous soil measured by Dualex in a) the third leaf level and b) fifth
midday leaf water potential (A) and midday stomatal conductance vs midday stem water potential (B) in response to different irrigation regimes in 3 year-old pear trees planted
No significant differences were seen in leaf and root total -SH contents between the control (33Mn+0Cd) and 0Mn+0Cd treatments, whereas they increased 6 and 4.9 times in the roots
Seasonal change in (A) leaf δ 15 N values, (B) leaf nitrogen total content and (C) area-based leaf nitrogen content for TW-FI (Transfer 281. water-Full Irrigation), TW-RDI
