ESCUELA DE INGENIARÍA DE LA INDUSTRIA FORESTAL,
AGRONÓMICA Y DE LA BIOENERGÍA
Dpto. de Ciencias Agroforestales
INSTITUTO UNIVERSITARIO DE INVESTIGACIÓN EN GESTIÓN
FORESTAL SOSTENIBLE
TESIS DOCTORAL:
From flower to fruit: Ecological and evolutionary implications of abiotic
and biotic factors affecting
Juniperus thurifera
L. cone development.
Presentada por Erik Rodríguez García para optar al grado de doctor
con Mención Internacional por la Universidad de Valladolid
Dirigida por:
Diseño de cubiertas y portada: Hugo Rodríguez García
Fotografías: Erik Rodríguez García. 2010 - 2016.
Portada y capítulo 2: Megastigmus thuriferana ovopositando Capítulo 1: Conos femeninos de sabina albar
Capítulo 3: Microscopía óptica de semilla de sabina: nucela con tubos polínicos Capítulo 4: Agujero de salida de M. thuriferana
Capítulo 5: Sabinar albar en Cabrejas del Pinar (Soria) Agradecimientos: Aegithalos caudatus
“Creo que la naturaleza une a las culturas del mundo. Un árbol tiene raíces en el suelo y ramas que rozan el cielo, y nos recuerda que para prosperar tenemos que saber de dónde venimos.”
Wangari Muta Maathai
CONTENTS
Resumen 11
Capítulo 1 De la flor al gálbulo: Implicaciones biológicas y evolutivas de los factores abióticos y bióticos que controlan la producción de gálbulos
en Juniperus thurifera L. Una visión general de la tesis 15 ¿Qué ocurre entre la floración y la dispersión?: el marco general de la tesis 16
¿Estoicismo o resignación? El caso de la sabina albar 19
Objetivos 22
Metodología 23
Principales resultados 27
Principales conclusiones 29
Referencias 31
Chapter 2 Ripe fruits may not be optimal: green fruits improve the detection of
traits associated to predispersal predation risk 39
Abstract 40
Introduction 41
Material and methods 42
Natural history 42
Study area 43
Sampling design and fruit analysis 43
Statistical analysis 44
Results 46
Phenological sequence of events 46
Fruit and full seed production 48
Fruit damage 49
Discussion 52
References 57
Supplementary material 60
Capítulo 3 Deciphering the role of reproductive investment, pollination success
and predispersal seed predation on reproductive output in Juniperus thurifera 65
Abstract 66
Introduction 67
Material and methods 69
Natural history 69
Study area 70
Sampling design and sample analyses 70
Statistical analysis 72
Results 73
Discussion 76
Capítulo 4 Efficiency of pollination and satiation of predators determine
reproductive output in Iberian Juniperus thurifera woodlands. 89
Abstract 90
Introduction 91
Material and methods 93
Natural history of the system 93
Population variation in fruit production and seed set 94
Population variation in fruit damage by arthropods 97
Variation in seed output across populations 98
Results 98
Variation in seed number 98
Variation in the proportion of full, empty and aborted seeds 98
Variation in fruit damage by arthropods 100
Seed output across populations 100
Discussion 101
References 107
Supplementary material 113
Capítulo 5 You’d better walk alone: Changes in forest composition affect pollination
efficiency and predispersal fruit damage in Iberian Juniperus thurifera forests. 115
Abstract 116
Introduction 117
Material and methods 119
Study area and sampling design 119
Study species 119
Fruit production and seed set 120
Predispersal fruit damage 122
Seed output 122
Statistical analysis 123
Results 124
Discussion 127
References 131
11
Resumen
El desarrollo de frutos y semillas es un factor crítico en la demografía de las plantas. La can-tidad de frutos y semillas que se desarrollan correctamente depende de múltiples factores: disponibilidad y uso de los recursos para la reproducción, éxito en la polinización y fertili-zación y depredación predispersiva. Estos factores actúan a escala individual o poblacional conjuntamente con otros factores como las condiciones climáticas, la densidad de individuos o la composición específica de los bosques.
La sabina albar (Juniperus thurifera L.) muestra una alta variabilidad interanual en la produc-ción de gálbulos (denominados indistintamente frutos, aunque desde un punto de vista botánico sean pseudofrutos). Por otra parte, las semillas muestran una viabilidad baja lo que puede estar relacionado tanto con la disponibilidad de recursos, como con la eficiencia de la polinización. Comprender los mecanismos que determinan el éxito reproductivo de la sabina albar y cómo se ven afectados por diferentes motores del cambio global es crítico para conocer el futuro de esta especie.
La presente tesis se estructura en cinco capítulos en los que se exploran los diferentes fac-tores que limitan la producción de gálbulos y semillas viables en la sabina albar a diferentes escalas temporales y espaciales. En el capítulo 1 se expone el marco teórico de la tesis, se muestra su diseño, así como los principales resultados y conclusiones obtenidos en la misma.
En el capítulo 2 se monitoreó el proceso de maduración de los gálbulos de sabina albar y la incidencia y selección de gálbulos de sus depredadores predispersivos durante tres cohortes de frutos entre los años 2013 y 2016. La mayor pérdida de gálbulos ocurre durante los cinco primeros meses de su desarrollo. Esta pérdida ocurre simultáneamente con la entrada de los principales depredadores predispersivos (ácaros, polillas, cochinillas y avispas). La identifica-ción de los criterios de selecidentifica-ción de gálbulos por parte de los artrópodos fue más eficiente si se realizaba en frutos inmaduros, que en maduros tal y como suele realizarse.
pes-12
ticidas que fueron aplicados en la cohorte de frutos de 2014. El experimento mostró cómo los niveles de recursos y la exclusión de los artrópodos depredadores fueron los principales factores que determinan el éxito reproductivo, afectando tanto a la cantidad de gálbulos que finalizan su desarrollo, como a la calidad de sus semillas. En este contexto de un sabinar denso, la polinización no resultó ser un factor limitante, y de hecho la adición de polen tuvo efectos negativos en la cantidad de gálbulos.
En el capítulo 4 se estudió a una escala geográfica amplia el impacto combinado de las con-diciones climáticas y de la depredación predispersiva sobre la producción de gálbulos y la calidad de las semillas. Se analizaron muestras recogidas en los años 2007 y 2008 proceden-tes de catorce sabinares ibéricos. La producción de gálbulos y su calidad estuvo favorecida por condiciones de humedad superiores a la media durante el primer año de formación del gálbulo. A nivel de árbol una producción elevada tuvo un efecto de saciado en ácaros, pero resultó en una atracción para polillas y avispas.
En el capítulo 5 se evaluó de qué modo la configuración específica de los sabinares puede afectar a la producción de gálbulos y niveles de depredación predispersiva. Se realizó un di-seño pareado con sabinares puros y mixtos en nueve localidades del centro de la península Ibérica. Los sabinares puros tuvieron una mayor producción de gálbulos, que pudo estar li-gado tanto a mayores niveles de recursos, un mayor éxito polinizador (como indica la menor tasa de aborción) y a la reducción global de la predación predispersiva en sabinares mixtos.
De la flor al gálbulo: Implicaciones biológicas y evolutivas
de los factores abióticos y bióticos que controlan la producción
de gálbulos en Juniperus thurifera
L.
Una visión general de la tesis
CAPÍTULO 1
“Es la hora de mover lo soñado de recurrir a aquello que parecía imposible”
Capítulo 1
16
¿Qué ocurre entre la floración y la dispersión?: el marco general de la tesis
Las plantas necesitan desarrollar correctamente sus frutos y semillas para una dispersión de los mismos y obtener así una reproducción satisfactoria (Herrera 1991, Pías et al. 2007). Sin embargo, desde que se desarrollan las estructuras destinadas a la reproduc-ción (flores en angiospermas, conos en gimnospermas) hasta que un fruto o semilla es dispersado existe una serie de factores, como el éxito de la polinización y fertilización o la depredación predispersiva, que controlan la cantidad de frutos que obtienen el nivel de desarrollo óptimo para su dispersión (Hainsworth 1984; Price et al. 2008; Boieiro et al 2012). Si bien, discernir la importancia relativa de cada uno de estos factores sobre el éxito reproductor es difícil debido a que varios componentes pueden interactuar simul-táneamente y afectarse mutuamente (Hainsworth 1984; Herrera 1991; Gruwez et al. 2013; Walsh et al. 2014), las plantas deben adaptar su ciclo vital a las diferentes condiciones y presiones para optimizar su éxito reproductor. Por otra parte, los resultados de estos procesos dependen en gran medida de su fenología (Ehrlén 2015). De esta forma, la inclusión de la historia natural en la investigación juega un papel decisivo a la hora de comprender procesos ecológicos como las relaciones planta – artrópodo (Östergård et al. 2007; Espelta et al. 2009, Xia et al. 2016) o como afectan las condiciones climáticas al éxito reproductor (Gruwez et al. 2013).
Para lograr una correcta reproducción, las plantas deben invertir recursos en la forma-ción de flores, frutos y semillas (Obeso; 2002, Hirayama et al. 2008; Teitel et al. 2016) de manera que a mayor inversión mayor éxito reproductivo. Sin embargo, la disponibilidad de recursos no es ilimitada, y su uso en reproducción promueve una competencia de recursos con otras funciones vegetativas, como el crecimiento (Obeso; 2002, Hirayama et al. 2008; Teitel et al. 2016), o incluso con los recursos disponibles para la reproducción futura (Karlsson et al 1990; Lyles et al. 2015; Pessendorfer et al. 2016).
Una visión general de la tesis
17
participan factores ambientales, por ejemplo: abundantes precipitaciones reducen el éxito de la polinización a través del viento o unas temperaturas cálidas que favorezcan la actividad de los insectos polinizadores (Knops et al. 2007; Rech et al. 2016).
Los frutos y semillas acumulan nutrientes durante su desarrollo. Este alto valor nutricional resulta atractivo para diversos depredadores especialistas, fundamentalmente artrópodos, que reducen la cantidad de frutos y semillas que llegan a la madurez (Fig 1.). Además, los artrópodos que se desarrollan en el interior de frutos y semillas también encuentran un refugio idóneo frente a sus enemigos e inclemencias climáticas (Sallabanks and Courtney 1992). Los artrópodos eligen los frutos por unas determinadas características, lo que genera una presión evolutiva sobre las plantas y su capacidad reproductora (Janzen 1971; Kolb et al. 2007). Como respuesta a esta presión, las plantas han desarrollado diferentes estrategias defensivas. Por un lado, presentan estrategias que actúan a corto plazo, como las defensas químicas y físicas (Janzen 1969; Schoonhoven et al. 2005; Boivin and Auger-Rozenberg 2016), la aborción selectiva (Bonal et al. 2010; Meyer et al. 2014; Boivin and Auger-Rozen-berg 2016) o el retraso en la fertilización de óvulos hasta después la puesta de huevos para evitar la pérdida de embriones (Rouault et al. 2004; Aderkas et al. 2005). Por otro lado, presentan mecanismos que actúan a largo plazo entre los que se incluye la modificación de
Capítulo 1
18
las características de los frutos para hacerlos menos atractivos a los depredadores (Janzen 1969; Espelta et al. 2009; Beckman y Muller-Landau 2011) o la alta variabilidad interanual en la producción de frutos y semillas, que obliga a los depredadores a ajustar sus ciclos demo-gráficos a los ciclos de producción de frutos, aumentando las plantas su éxito reproductivo durante los años de alta producción (Turgeon 1994).
El éxito reproductivo es modulado, además, por factores que actúan a mayores escalas es-paciales, como las condiciones climáticas (García et al. 2000; Obeso 2002), la fragmentación del hábitat (Sork et al. 2002; Knight et al. 2005), la densidad de individuos y su estructura (Sork et al. 2002; Knight 2003; Sanz and Pulido 2015) o la presencia de otras especies (Mugnaini et al. 2007; Aderkas et al. 2012). De hecho, a una escala geográfica amplia es el clima quien determina la variabilidad en la producción de frutos que existe entre diferentes poblaciones (García et al. 2000; Obeso 2002; Montesinos et al. 2010), mientras que, a una escala local, variaciones temporales del clima determinan la inversión reproductiva (Lee y Bazzaz 1982; Herrera 1991; Crone y Lesica 2006). Para especies con una fuerte variación interanual en su producción de frutos hay una clara correlación entre las condiciones climá-ticas y una alta producción (Kelly and Sork 2002).
Una visión general de la tesis
19
de la dispersión (Santos and Tellería 1994, González-Varo et al. 2009). Por el contrario, una densificación de los bosques puede resultar también en una mayor depredación predisper-siva de frutos y semillas (Sholes 2008; Guyot et al. 2016) y en el caso de bosques mixtos, la presencia de otras especies puede reducir la eficiencia en la polinización, especialmente por el viento, debido a un efecto barrera o por interferencia polínica entre especies que solapen sus periodos reproductivos (Mugnaini et al. 2007, Aderkas et al. 2012, Millerón et al. 2012).
¿Estoicismo o resignación? El caso de la sabina albar
La sabina albar (Juniperus thurifera L.; Fig. 3) es un endemismo mediterráneo occidental con sus principales poblaciones en la Península Ibérica y Marruecos, presentando relic-tos en Francia, Italia y Argelia (Costa et al. 1997). Se trata de una especie que aparece en zonas de clima mediterráneo de marcada continentalidad y sequía estival, generalmente en suelos pobres predominantemente calcáreos, aunque también en suelos silíceos (p.e. Sierra de Guadarrama en Segovia o Barrios de Luna en León) (Costa et al. 1997). Además, los sabinares albares están incluidos en el anexo IV de la directiva hábitats (código 9560) y si bien la especie está catalogada como preocupación menor (LR/lc) según la Unión
Capítulo 1
20
Internacional para la Conservación de la Naturaleza (UICN) sus poblaciones se encuentran severamente fragmentadas y en continuo decrecimiento (Farjon 2013). La estructura actual de los sabinares albares es consecuencia de un manejo tradicional ligado a la ganadería ovina y caprina (Olano et al. 2008; Fig. 3 A y B), uso que aún persiste en el Atlas marroquí si bien en España no ha desaparecido por completo todavía. Durante la segunda mitad del siglo XX los usos tradicionales de los sabinares han ido cesando su actividad lo que está generando cambios en la estructura de estos bosques (Fig. 3C). Así, están entrando nuevas especies arbóreas como encinas, robles o pinos y se está produciendo una densificación de los sabinares (Olano et al. 2012).
La dispersión de la sabina albar se basa en el consumo de sus pseudofrutos carnosos, gál-bulos o conos por un amplio grupo de vertebrados, principalmente aves del género Turdus
(Tellería et al. 2011) y carnívoros como el zorro (Vulpes vulpes (Linnaeus 1758)) o la garduña (Martes foina (Erxleben 1777)) que son muy eficientes para la dispersión en espacios abier-tos (Escribano-Ávila et al. 2012). Sin embargo, la cantidad de gálbulos viables lisabier-tos para la dispersión está afectada por múltiples factores y puede ser muy baja (Montesinos 2010). Parte de esta limitación reside en la fuerte variabilidad interanual, ligado a la estrategia de reducción de las elevadas tasas de depredación predispersiva por artrópodos (Mezquida y Olano 2013). Mientras que, por otra parte, la sabina albar presenta una reducida viabilidad de las semillas (Montesinos et al. 2010) lo que puede estar relacionado tanto con la efi-ciencia de la polinización y la disponibilidad de recursos. En general, estos efectos podrían agudizarse en un futuro cercano, como consecuencia de los cambios de uso y del cambio climático. En otras especies de Juniperus se ha predicho un aumento en la tasa de pérdida de gálbulos por factores bióticos y abióticos, reduciendo el número de semillas viables como se ha indicado para otras especies del género (Gruwez et al. 2013), además de una posible disminución de las tasas de dispersión de frutos y semillas. Aunque la sabina albar muestra una gran capacidad de adaptación a variaciones en las condiciones climáticas, haciéndola muy resiliente frente a un futuro escenario de clima más cálido y seco (Camarero et al. 2010), las tasas de depredación predispersiva por artrópodos y aborto de semillas son especialmente sensibles a las condiciones ambientales (Montesinos et al. 2010), pudiendo ser un cuello de botella para el futuro de la especie.
Una visión general de la tesis
21 Fig. 3. A: Sabinas albares en Siguero (Segovia). B: Sabinar albar en Calatañazor (Soria). C: Sabinar albar mixto en Megina (Guadalajara)
A
B
Capítulo 1
22
de ellos que llegan a un estado óptimo para su dispersión (Roques 1984; ver pág. 69-70). En el caso de la península Ibérica este grupo de artrópodos está formado principalmente por: los ácaros Trisetacus quadrisetus (Thomas 1889) (Acari, Phytoptidae), las polillas Pammene juniperana (Millière 1858) (Lepidoptera, Tortricidae) y Mesophleps oxycedrella (Milière 1871) (Lepidoptera, Gelechiidae) y la avispa Megastigmus thuriferana Roques & El Alaoui 2006 (Hymenoptera, Torymidae). Estas especies atacan secuencialmente los gálbulos durante su desarrollo mostrando preferencias por determinadas características de los gálbulos. Esta depredación de gálbulos secuencial lleva a una aborción selectiva de los gálbulos infesta-dos en las fases tempranas de su desarrollo. Además, cuando los gálbulos son atacainfesta-dos en fases de desarrollo similares a la madura, los artrópodos depredadores podrían seleccionar gálbulos con características similares a las preferidas por los dispersores, generando presio-nes selectivas conflictivas (Siepielski y Benkman 2007).
Objetivos
El objetivo principal de esta tesis es comprender los diferentes factores que limitan la producción de gálbulos y semillas viables en la sabina albar, así como comprender el im-pacto y selección de frutos por parte de los depredadores predispersivos, determinando su respuesta a diferentes escalas temporales y ambientales. Los trabajos de investigación llevados a cabo para esta tesis se organizan en cuatro capítulos cuyos objetivos específicos son:
Una visión general de la tesis
23
Capítulo 3. Discernir el papel que juegan los niveles de recursos, el éxito en la polinización y la depredación predispersiva y cómo interactúan entre ellos para determinar el éxito reproductivo final en la sabina albar. Con ello se pretende comprender mejor cómo interac-túan entre sí los diferentes filtros que determinan el número final de gálbulos que llegan a madurar. Este trabajo modificó experimentalmente los niveles de polen y de depredadores predispersivos.
Capítulo 4. Evaluar a una escala geográfica amplia el impacto combinado de las condiciones climáticas y de la depredación predispersiva sobre la producción de gálbulos y la calidad de las semillas. Con ello se trata de comprender cómo se comportan la sabina y sus depre-dadores bajo un gradiente de condiciones climáticas y cómo estas condiciones interactúan con los sucesos locales.
Capítulo 5. Evaluar cómo la configuración de los sabinares puede afectar a la producción de gálbulos y niveles de depredación predispersiva. Se realizó un diseño muestral com-prendiendo bosques puros y mixtos de sabina en diferentes regiones geográficas. En este trabajo se trata de observar de qué modo la tendencia de los sabinares a convertirse en bosques mixtos pueden afectar el éxito reproductor de la especie.
Metodología
Esta tesis aborda la cuestión con un enfoque observacional, combinado con un diseño experimental en el capítulo 3. Se recolectaron gálbulos en diferentes fases de desarrollo de diversos sabinares de la península Ibérica en los que se ha realizado trabajo de campo entre los años 2012 y 2016 durante la duración de la tesis. Además, en el capítulo 4 se dispuso de muestras recolectadas en años anteriores por el equipo de trabajo.
Capítulo 1
24
artrópodos en diferentes fases fenológicas. Estos datos se compararon con los resultantes de los gálbulos maduros que se recogían en octubre de su segundo año de desarrollo.
En el capítulo 3 se trató de discernir experimentalmente el papel que juegan los niveles de recursos, el éxito en la polinización y la depredación predispersiva en el desarrollo de gálbulos. Para ello se realizó un experimento con un diseño factorial completo que incluía diferentes tratamientos: A) control, B) aporte extra de polen, C) aporte de pesticidas y D) aporte extra de polen + pesticidas. Se realizó en el mismo sabinar que el estudio anterior, en Villaciervos (Soria). Para ello se seleccionaron al azar cuarenta árboles. En cada árbol se seleccionaron 8 ramas y se aplicó cada uno de los tratamientos a dos ramas por árbol. El experimento se realizó para la cohorte de gálbulos del año 2014. Se realizó además un seguimiento mensual de la pérdida de frutos. En octubre de 2015 se recolectaron todos los gálbulos maduros de las ramas tratadas para estimar la depredación por artrópodos y analizar la calidad de las semillas. Con el fin de observar el efecto del aporte extra de polen sobre la fertilización en junio de 2015 se recolectaron 100 gálbulos inmaduros (50 control y 50 con aporte extra de polen). Estas muestras fueron procesadas para la observación, en microscopio óptico, de la formación de tubos polínicos (ver metodología en Capítulo 3). La influencia del esfuerzo reproductivo inicial, el aporte extra de polen y la aplicación de pesticidas sobre el desarrollo de gálbulos, la calidad de las semillas y la depredación predispersiva se testó mediante modelos mixtos.
En el capítulo 4 se evalúo cómo influyen las condiciones ambientales en el desarrollo de gálbulos y su efecto combinado sobre la calidad de las semillas y la depredación predis-persiva a una escala geográfica amplia. Para ello se analizaron muestras provenientes de catorce sabinares a lo largo de su distribución en la península Ibérica recolectadas durante los años 2007 y 2008. En cada sabinar fueron seleccionados al azar treinta sabinas en las que se estimó la producción de gálbulos y se recolectaron treinta gálbulos para la estima de la depredación predispersiva y diez para el análisis de sus características y de la calidad de sus semillas. La influencia combinada de las condiciones ambientales y la producción de gálbulos sobre la calidad de las semillas y la depredación predispersiva se evaluó mediante modelos mixtos.
Una visión general de la tesis
25
clara dominancia de la sabina albar (> 90 % de representación en pies) y bosques con baja representación (< 50 %) obviando los estados intermedios (51 – 89 %). Con este esquema, durante otoño de 2014 se realizó un diseño pareado con sabinares puros y mixtos en diferentes localidades del centro de la Península Ibérica: Megina, Pinilla de Molina y Valher-moso (Guadalajara); Arcones, Prádena y Siguero (Segovia); Bayubas de Abajo, Calatañazor y Cabrejas del Pinar (Soria). En cada sabinar fueron seleccionadas al azar treinta sabinas en las que se estimó la producción de gálbulos y se recolectaron treinta gálbulos para la estima de la depredación predispersiva y diez para el análisis de sus características y calidad de sus semillas. El efecto de la composición del bosque sobre la producción de gálbulos y la calidad de sus semillas se evaluó mediante modelos mixtos.
Capítulo 1
26
experimentador, sino que interesa conocer en qué medida la estimación de los valores de la variable respuesta pueden variar en función de la población (niveles del factor) que se seleccione de un modo aleatorio. De esta forma el factor “población” representaría el efecto aleatorio en el modelo mixto. Estos modelos pueden ser empleados tanto para variables que siguen una distribución normal (modelos mixtos lineales; capítulo 4 y 5) como para variables que siguen otro tipo de distribuciones, como la binomial o la Poisson (modelos mixtos lineales generalizados; capítulos 2 a 5).
En cada capítulo de la tesis, los efectos fijos se eligieron en base a los conocimientos biológicos previos. En el capítulo 2 con el fin de simplificar la comparación de modelos
1 mm
1 mm
1 mm
1 mm
Fig 4. Diferentes estados identificados en las semillas de sabina albar. A: Semilla llena, mostrando un embrión y megagametofito bien desarrollados. B: Semilla vacía con embrión y megagametofito sin desarrollar por completo. C: Semilla vacía sin embrión ni megagametofito desarrollados. D: Semilla abortada sin desarrollo post-fertilización..
A B
Una visión general de la tesis
27
obtenidos en las diferentes fases fenológicas sólo se consideraron modelos saturados. En el capítulo 3 se realiza una selección de modelos eliminando secuencialmente las variables menos explicativas hasta obtener un modelo óptimo sencillo. En el capítulo 4 con el fin de obtener información general de los posibles efectos fijos a una escala geográfica amplia sólo se consideraron modelos saturados. Finalmente, en el capítulo 5 se realizó primero una selección de la estructura de efectos aleatorios sobre los modelos saturados y, segundo, una selección de estructura de efectos fijos tal como se indica en Zuur et al. (2009). Esta selección de estructuras se realizó en base al criterio de información bayesiana (BIC) reco-mendable cuando el objetivo es evaluar hipótesis (Aho et al. 2014).
Principales resultados
En el capítulo 2 se describió el ciclo vital, asociado al desarrollo de los gálbulos, de los de-predadores. Los diferentes artrópodos depredadores atacaron secuencialmente los conos y semillas de la sabina albar. Los ácaros colonizaron las flores a la espera, tras la poliniza-ción, que se inicie el desarrollo de semillas para su infestación. Salieron del interior de las semillas durante el invierno en busca de nuevas flores que colonizar. Se identificaron las dos especies de polillas, M. oxycedrella y P. juniperana. Estas realizaron la puesta de huevos sobre los gálbulos, de primer y segundo año, desde finales de mayo a mediados de junio, entrando sus larvas en los gálbulos desde mediados de junio a finales de julio. Las larvas se desarrollaron en el interior del gálbulo y salieron de su interior a partir de finales de octu-bre. Las cochinillas colonizan nuevos gálbulos en su fase de ninfa durante el mes de junio. Una vez seleccionado el lugar idóneo sufren la metamorfosis a la fase sedentaria y adulta completando su ciclo vital. Finalmente, las avispas depositaron huevos en el interior de las semillas desde finales de julio hasta mediados de septiembre. Las larvas se desarrollaron en el interior de las semillas a lo largo de doce meses, saliendo de su interior desde finales de julio hasta finales de agosto.
Capítulo 1
28
gálbulos de mayor tamaño, característica que se relaciona con un mayor contenido de semillas viables; las cochinillas no seleccionan gálbulos en base a su tamaño y/o contenido en semillas. Durante el segundo año de formación del cono: la polilla M. oxycedrella selec-cionó los gálbulos de mayor tamaño, pero con un menor número de semillas en su interior. Por el contrario, cuando la evaluación de la selección se basó en gálbulos maduros no fue detectado ningún efecto para entrada de las dos especies de polillas, mientras que la avispa seleccionó gálbulos con un mayor contenido en semillas.
Los resultados correspondientes al capítulo 3 muestran que existe una asociación entre un mayor nivel de recursos estimado como número de flores por rama y éxito reproductor. Estimado tanto en términos de número final de gálbulos, como de tasa de éxito por flor y de número de semillas llenas por gálbulo. La polinización no resultó ser un facor limitante, además, la adición manual de polen no mejoró el éxito reproductivo e incluso supuso un menor número de gálbulos que completaron su desarrollo. Sin embargo, las flores que fueron tratadas con extra de polen mostraron una menor tasa de aborción en sus semillas. La exclusión de los artrópodos depredadores mediante biocidas resultó en un aumento en el cuajado de gálbulos, así como una mejor calidad de las semillas.
Los resultados correspondientes al capítulo 4 muestran que las condiciones ambientales favorables se asocian con una mayor eficiencia en la polinización, incrementando la pro-ducción de gálbulos y la calidad de sus semillas. Así, condiciones de humedad superiores a la media durante el primer año de formación del gálbulo incrementó su producción total. A su vez, los árboles con una mayor producción de gálbulos presentaron más semillas llenas y menos vacías y abortadas. Árboles con una alta producción de gálbulos generaron un efecto de saciado en los depredadores predispersivos menos móviles como los ácaros mientras que polillas y avispas, con una capacidad de dispersión mayor, buscan árboles con una mayor producción. Además, las polillas y avispas seleccionaron árboles con gálbulos más grandes y con un mayor contenido en semillas a la vez que evitaron árboles con una mayor incidencia de otros depredadores.
Una visión general de la tesis
29
viables por gálbulo resultó similar en ambos tipos de sabinar, los puros presentaron una menor cantidad de semillas abortadas y mayor de vacías lo que indica que en sabinares puros hay una mayor cantidad de semillas que inician su desarrollo. Por otra parte, los artró-podos que consumen los frutos y semillas mostraron una respuesta más compleja. Ácaros y polillas mostraron mayores consumos de gálbulos y semillas en sabinares puros, mientras que la depredación por avispas fue mayor en bosques mixtos.
Principales conclusiones
La inclusión de la historia natural y fenología de los procesos de formación del gálbulo en sabina albar y su interacción los artrópodos depredadores de gálbulos proporciona una mejor comprensión de las relaciones planta-artrópodo. En primer lugar, nos permite ob-servar cómo se ajustan entre sí los ciclos biológicos de la sabina albar y de los artrópodos depredadores. A la vez, nos permite discernir en qué momento se producen las respuestas defensivas por parte de la sabina albar, principalmente en forma de aborto selectivo. Si bien tras la polinización la cantidad de gálbulos que comienzan su desarrollo es elevada (ver Capítulo 2) hay una gran pérdida de frutos entre mayo y septiembre del primer año de formación. Esta aborción parece asociada a la entrada de ácaros y polillas (ver capítulo 2 y 3).
Por otra parte, en frutos cuyo proceso de desarrollo es largo, como es el caso de los gálbulos de sabina albar, el aborto selectivo en etapas tempranas de formación puede implicar que se subestime la capacidad depredadora de los artrópodos (ver capítulo 2). Es más, como la aborción selectiva no es igual para todas las especies, esto puede provocar cambios en la importancia relativa de los diferentes agentes implicados (ver capítulo 3). En tercer lugar, en esta tesis hemos visto cómo los modelos de selección de fruto basados en fases fenológicas cercanas a la ovoposición proporcionan una mejor comprensión de las presiones selectivas en aquellos depredadores que infestan los gálbulos en etapas tempranas (ver capítulo 2).
Capítulo 1
30
indicios que sugieren una limitación polínica.
En este trabajo diferentes indicios sugieren la existencia de una interacción entre diferentes factores. Así, una mayor inversión en reproducción puede llevar a una saturación de los ácaros (ver capítulo 3), resultando en un mayor éxito reproductivo. Sin embargo, los de-predadores con mayor capacidad dispersiva (polillas y avispas) son atraídos por los árboles que muestran una mayor producción de gálbulos (ver capítulos 3 y 4). Por otra parte, los diferentes depredadores interactúan entre sí (ver capítulos 3 y 4), de forma que la exclusión de ácaros y polillas conllevó una mayor tasa de gálbulos atacados por avispas (ver capítulo 3).
Cuando se abre el foco y se analiza a escalas espaciotemporales mayores, se observa cómo las condiciones ambientales en el año de floración e inicio de los frutos modulan la forma-ción de gálbulos (ver capítulo 4). Así, hay una relaforma-ción positiva entre la cantidad de gálbulos y de semillas viables con condiciones de mayor humedad con respecto de la media en las etapas tempranas de formación del gálbulo. Además, la depredación predispersiva está determinada por las condiciones ambientales de cada zona y por la producción y características de los gálbulos (ver capítulo 4). Así, por ejemplo, los ácaros depredan menor porcentaje de los gálbulos cuando los años son más húmedos.
Una visión general de la tesis
31
Referencias
Aderkas P., Rouault G., Wagne R., Rohr R., Roques A. (2005) Seed parasitism redirects ovule develop-ment in Douglas fir. Proceedings in Biological Sciences / The Royal Society 272: 1491–1496.
Aderkas P., Nepi M., Rise M., Buffi F., Guarnieri M., Coulter A., Gill K., Lan P., Rzemieniak S., Pacini E. (2012) Post-pollination prefertilization drops affect germination rates of heterospecific pollen in larch and Douglas-fir. Sex Plant Reproduction25: 215-225.
Aho K., Derryberry D.W., Peterson T. (2014) Model selection for ecologist: the worldviews of AIC and BIC. Ecology 95: 631-636
Albert M.J., Escudero A., Iriondo J.M., (2001) Female reproductive success of narrow endemic Erodium paularense in contrasting microhabitats. Ecology82: 1734-1747
Améztegui A., Brotons L., Coll L. (2010) Land-use changes as major drivers of mountain pine (Pinus uncinata Ram.) expansion in the Pyrenees. Global Ecololy and Biogeography19: 632-641.
Beckman, N.G., & Muller-Landau, H.C. (2011). Linking fruit traits to variation in predispersal vertebrate seed predation, insect seed predation, and pathogen attack. Ecology92: 2131–2140.
Boieiro M., Rego C., Serrano A.R.M., and Espadaler X. (2012) Seed production and pdispersal re-productive losses in the narrow endemic Euphorbia pedroi (Euphorbiaceae). Plant Ecology213: 581–590.
Boivin T., Auger-Rozenberg M., Native fruit, cone and seed insects in the Mediterranean Basin. In Paine T.D., Lieutier F. (2016) Insects and diseases of mediterranean forest systems. Pp 47-88. Switzerland: Springer International Publishing.
Bonal R., Muñoz A., Espelta, J.M. (2010) Mismatch between the timing of oviposition and the seasonal optimum. The stochastic phenology of mediterranean acorn weevils. Ecological Entomology 35: 270-278.
Camarero J.J., Olano J.M., Parras A. (2010). Plastic bimodal xylogenesis in conifers from continental Mediterranean climates. New Phytologist185: 471-480
Costa M., Morla C., Sainz H. (1997) Los bosques ibéricos: una interpretación geobotánica. Barcelona (Spain): Planeta. 597p.
Chauchard S., Carcaillet C., Guibal F. (2007) Patterns of land-use abandonment control tree-recruit-ment and forest dynamics in Mediterranean mountains. Ecosystems10: 936–948.
Crone E.E., Lesica P. (2006) Pollen and water limitation in Astragalus scaphoides, a plant that flowers in alternate years. Oecologia150: 40–49.
Ehrlén, J. (2015). Selection on flowering time in a life-cycle context. Oikos124: 92–101.
Capítulo 1
32
Assemblage and Environmental Heterogeneity. PLoS ONE 7: e46993. doi:10.1371/journal. pone.0046993
Espelta, J.M., Bonal R., Sánchez-Humanes B. (2009) Pre-dispersal acorn predation in mixed oak forests: Interspecific differences are driven by the interplay among seed phenology, seed size and preda-tor size. Journal of Ecology97: 1416–1423.
Farjon, A. 2013. Juniperus thurifera. The IUCN Red List of Threatened Species 2013: e.T42255A2967372. http://dx.doi.org/10.2305/IUCN.UK.2013-1.RLTS.T42255A2967372.en. Downloaded on 28 March 2017.
García D., Zamora R., Gómez J.M., Jordano P., Hodar J.A. (2000) Geographical variation in seed produc-tion, predation and abortion in Juniperus communis throughout its range in Europe. Journal of Ecology88: 435–446.
Getzin S., Dean C., He F., Trofymow J.A., Wiegand K., Wiegand T. (2006) Spatial patterns and competi-tion of tree species in a Douglas-fir chronosequence on Vancouver Island. Ecography29: 671-682
Gibbs H.K., Ruesch A.S., Achard F., Clayton M.K., Holmgren P., Ramankutty N., Foley J.A. (2010) Tropical forests were the primary sources of new agricultural land in the 1980s and 1990s. Procedures of the Natural Academy of Sciences107: 16732-16737.
Gimeno T.E., Pías B., Martínez-Fernández J., Quiroga D.L., Escudero A., Valladares F. (2012) The de-creased competition in expanding versus mature juniper woodlands is counteracted by adverse climatic effects on growth. European Journal of Forest Research131: 977-987.
González-Varo J.P., Arroyo J., Aparicio A. (2009) Effects of fragmentation on pollinator assemblage, pollen limitation and seed production of Mediterranean myrtle (Myrtus communis). Biological Conservation142: 1058–1065.
Gruwez R., Leroux O., De Frenne P., Tack W., Viane R., Verheyen K. (2013) Critical phases in the seed development of common juniper (Juniperus communis). Plant Biology15: 210–219.
Guyot V., Castagneyrol B., Vialatte A., Deconchat M., Jactel H. (2016) Tree diversity reduces pest damage in mature forest across Europe. Biology Letters12: 20151037. http://dx.doi.org/10.1098/ rsbl.2015.1037.
Hainsworth F.R., Wolf L.L., Mercier T. (1984) Pollination and Pre-Dispersal Seed Predation: Net Effects on Reproduction and Inflorescence Characteristics in Ipomopsis aggregate. Oecologia63: 405-409
Hansen A.J., Neilson R.P., Dale V.H., Flather C.H., Iverson L.R., Currie D.J., Shafer S.C.R., Bartlein P.J. (2001) Global change in forests: Responses of Species, communities and biomes. Bioscience51: 765-779
Herrera C.M. (1991) Dissecting factors responsible for individual variation in plant fecundity. Ecology 72: 1436–1448.
Una visión general de la tesis
33
Janzen D.H. (1969). Seed-eaters versus seed size, number, toxicity and dispersal. Evolution23: 1–27.
Janzen D.H. (1971) Seed predation by animals. Annual Review of Ecology and Systematics2:465–492
Karlsson P.S., Svensson B.M., Carlsson B.Å., Nordell K.O. (1990) Resource investment in reproduction and its consequences in three Pinguicula species. Oikos59: 393–398.
Kenkel N.C. (1988) Pattern of self-thinning in Jack pine: testing the random mortality hypothesis.
Ecology69:1017-1024.
Koenig W.D., Knops J.M.H., Carmen W.J., Pearse I.S. (2015) What drives masting? The phenological synchrony hypothesis. Ecology 96: 184–192.
Knight T.M. (2003) Floral density, pollen limitation, and reproductive success in Trillium grandiflorum.
Oecologia137: 557–563.
Knight T., Steets J., Vamosi J., Mazer S., Burd M., Campbell D.R., … Ashman T.L. (2005) Pollen limitation of plant reproduction: pattern and process. Annual Review of Ecology Evolution and Systematics36: 467–497.
Knops J.M.H., Koenig W.D., Carmen W.J. (2007) Negative correlation does not imply a tradeoff be-tween growth and reproduction in California oaks. Procedures of the Natural Academy of Sciences 104:16982–16985
Kudo G., Harder, L.D. (2005) Floral and inflorescence effects on variation in pollen removal and seed production among six legume species. Functional Ecology 19: 245–254.
Labouche A.M., Richards S.A., Pannell J.R. (2016) Effects of pollination intensity on offspring number and quality in a wind-pollinated herb. Journal of Ecology105: 197–208.
Lee T.D., Bazzaz F.A. (1982) Regulation of fruit and seed production in an annual legume, Cassia fasci-culata. Ecology63: 1363–1373.
Lyles D., Rosenstock T.S., Hastings A. (2015) Plant reproduction and environmental noise: How do plants do it? Journal of Theoretical Biology371: 137-144
Meyer K., Soldaat L., Auge H., Thulke H. (2014) Adaptive and Selective Seed Abortion Reveals Complex Conditional Decision Making in Plants. American Naturalist183: 376–83.
Mezquida E.T., Olano J.M. (2013) What makes a good neighborhood? Interaction of spatial scale and fruit density in the predator satiation dynamics of a masting juniper tree. Oecologia173: 483-492.
Millerón M, Heredia U.L., Lorenzo Z., Perea R., Dounavi A., Alonso J., Gil L., Nanos N. (2012) Effect of canopy closure on pollen dispersal in a wind-pollinated species (Fagus sylvatica L.). Plant Ecology 213: 1715-1728
Capítulo 1
34
Montesinos D., Fabardo J. (2015) Changes in land use and physiological transitions of Juniperus thuri-fera forest: from decline to recovery. Canadian Journal of Forest Research45: 764–769
Mugnaini S., Nepi M., Guarnieri M., Piotto B., Pacini E. (2007) Pollination drop in Juniperus communis: response to deposited material. Annals of Botany100: 1475-1481
Obeso J.R. (2002) The costs of reproduction in plants. New Phytologist155: 321–348.
Olano J.M., Rozas V., Bartolomé D., Sanz D. 2008. Effects of changes in traditional management on hei-ght and radial growth patterns in a Juniperus thurifera woodland. Forest Ecology and Management 255: 506-512
Olano J.M., Zavala M.A., Rozas V. 2012. Disruption of Juniperus thurifera woodland structure in its nor-thwestern geographical range: potential drivers and limiting factors. European Journal of Forest Research131: 563-570.
Östergård H., Hambäck P.A., Ehrlén, J. (2007). Pre-dispersal seed predation: The role of fruit abortion and selective oviposition. Ecology88: 2959–2965.
Pessendorfer M.B., Koenig W.D., Pearse I.S., Knops J.M.H., Funk K.A. (2016) Individual resource limita-tion combined with populalimita-tion-wide pollen availability drives masting in the valley oak (Quercus lobata). Journal of Ecology104: 637–645.
Phelps J., Carrasco L.R., Webb E.L., Koh L.P., Pascual U. (2013) Agricultural intensification escalates future conservation costs. Procedings of the Natural Academy of Sciences110: 7601-7606.
Pías B., Salvande M., Guitián P. (2007) Variation in Predispersal Losses in Reproductive Potential in Rowan (Sorbus aucuparia L. Rosaceae) in the NW Iberian Peninsula. Plant Ecology188: 191–203.
Price M.V., Campbell D.R., Waser N.M., Brody A.K. (2008) Bridging the generation gap in plants: pollina-tion, parental fecundity, and offspring demography. Ecology89:1596–1604.
Rech A.R., Dalsgaard B., Sandel B., Sonne J., Svenning J.C., Holmes N., Ollerton J. (2016) The macro-ecology of animal versus wind pollination: ecological factors are more important than historical climate stability. Plant Ecology & Diversity9: 253–262.
Rey Benayas J.M., Martins A., Nicolau J.M., Schulz J.J. (2007) Abandonment of agricultural land: an overview of drivers and consequences. CAB reviews: perspectives in agriculture, veterinary sciences, nutrition and natural resources 2. Wallingford (UK): CABI Publishing. 14p.
Roques A., Raimbault J.P., Goussard F. (1984). La colonisation des cônes et des galbules des Genévriers Méditerranéens par les insectes et les Acariens et son influence sur la possibilité de régénération Naturelle de ces essences. Ecologia MediterraneaX: 147–169.
Rouault G., Turgeon J., Candau J.N., Roques A., von Aderkas P. (2004) Oviposition strategies of conifer seed chalcids in relation to host phenology. Naturwissenschaften91: 472–480.
Una visión general de la tesis
35
Santos T., Tellería J.L. (1994) Influence of forest fragmentation on seed consumption and dispersal of Spanish juniper Juniperus thurifera. Biological Conservation70: 129–134
Sanz R., Pulido F. (2015). Pollen limitation and fruit abortion in a declining rare tree, the Eurasian yew (Taxus baccata L.): A reproductive cost of ecological marginality. Plant Biosystems149: 818–826.
Schoonhoven L.M., van Loon J.J.A., Dicke M. (2005). Insect-Plant Biology. Oxford (U.K.): Oxford Univer-sity Press.
Siepielski A.M., Benkman C.W. (2007). Selection by a predispersal seed predator constrains the evolu-tion of avian seed dispersal in pines. Functional Ecology21: 611-618.
Sholes O.D.V. (2008) Effects of associational resistance and host density on woodland insect herbivo-res. Journal of Animal Ecology77: 16-23.
Sork V.L., Davis F.W., Smouse P.E., Apsit V.J., Dyer R.J., Fernandez J.F. Kuhn B. (2002). Pollen movement in declining populations of California Valley oak, Quercus lobata: Where have all the fathers gone?
Molecular Ecology11: 1657–1668.
Teitel Z., Pickup M., Field D.L., Barrett S.C.H. (2016). The dynamics of resource allocation and costs of reproduction in a sexually dimorphic, wind-pollinated dioecious plant. Plant Biology18: 98–103.
Tellería J.L., de la Hera I., Ramirez A., Santos T. (2011). Conservation opportunities in Spanish juniper
Juniperus thurifera woodlands: the case of migratory thrushes Turdus spp. Ardeola 58: 57-70
Turgeon J. (1994) Insect exploiting seed cones of Larix spp., Picea spp., and Pinus spp.: species richness and patterns of exploitation. In: Turgeon J.J., de Groot P. (eds). Biology, damage and management of seed orchard pests. Ontario (Canada): Canadian Forestry Service. pp 15–30
Valladares F., Benavides R., Rabasa S.G., Díaz M., Pausas J.G., Paula S., Simonson W.D. (2014) Global Change and Mediterranean forests: current impacts and potentias responses in Forest and Global Change. In Coomes D.A., Burslem D.F.R.P., Simonson W.D., editors. Forest and Global Change. Cam-bridge (U.K.): CamCam-bridge University Press. p47-75.
Vayreda J., Martinez-Vilalta J., Gracia M., Canadell J., Retana J. (2016) Anthropogenic-driven rapid shifts in tree distribution lead to increased dominance of broadleaf species. Global Change Biology doi: 10.1111/gcb.13394
Walsh R.P., Arnold P.M., Michaels H.J. (2014). Effects of pollination limitation and seed predation on female reproductive success of a deceptive orchid. AoB PLANTS6: 1–12.
Wang Y., Pederson N., Ellison A.M., Buckley H.L., Case B.S., Liang E., Camarero J.J. (2016) Increased stem density and competition may diminish the positive effects of warming at alpine treeline. Ecology 97: 1668-1679.
Capítulo 1
36
Ripe fruits may not be optimal: green fruits improve the detection
of traits associated to predispersal predation risk
“Weeks in the field can save you minutes in the library”
Thomas Eisner
Capítulo 2
40
Abstract
Ripe fruits may not be optimal
41
Introduction
Arthropods and plants are among the largest taxa of living organisms in number of spe-cies and in terrestrial ecosystem biomass, respectively. Spespe-cies in both groups interact in different ways with each other, and close interactions has been traditionally considered as the promoters of the current diversity of terrestrial life forms (Ehrlich and Raven 1964). In fact, 90 % of flowering plants are estimated to be pollinated by animals, mainly by insects (Ollerton et al. 2011) and 26 % of the insect species feed on vascular plants (Schoonhoven et al. 2005). Interactions between plants and insects had critical economic implications especially in agricultural systems (Rechcigl and Rechcigl 2000; Schoonhoven et al. 2005, Badii et al. 2015) and are basic for the functioning of natural systems from an ecological and evolutionary point of view (Janzen 1971; Kolb et al. 2007; Suchan and Alvarez 2015).
Capítulo 2
42
Our aim was to explore whether the common practice of exploring predator-plant selection using ripe fruits provides similar information to evaluating this relationship at the pheno-logical stage corresponding to the timing of fruit use by each seed predator. To perform this task, we selected the conifer Spanish juniper (Juniperus thurifera), which produces pseudo-fruits (hereafter fruits) that mature over a 20 months period when they are attacked by a wide array of specialized arthropods, including mites, moths, chalcid wasps and scale insects (Roques et al. 1984). The ecological and evolutionary implications of seed predation in this species and other junipers has received an intense research effort which has been based on the evaluation of ripe fruits (Mezquida and Olano 2013; Roques et al. 2013; El Alaoui et al. 2013; Mezquida et al. 2016). We hypothesized that plants having long seed maturiation periods, levels of seed predation might be underestimated due to selective fruit losses of infested fruits with different abortion rates determining changes in their rela-tive infestations ranks. Moreover, we predicted that the set of traits associated to predator preferences may differ from the traits evaluated on ripe fruits, and that these differences would be more intense for seed predators showing higher losses of damaged fruits.
Material and methods
Natural history
Spanish juniper (Juniperus thurifera L.) is a dioecious juniper species, endemic to the Western Mediterranean where it inhabits continental summer-dry environments. Wind pollination occurs in late winter-early spring. Pollen germination and megagametophyte formation starts after pollination, but fertilization is delayed until late spring. Embryo development begins after fertilization and seeds are almost fully grown by mid-summer, when they start to harden their seed coat. Megagametophyte and embryo final maturation takes 12-13 ad-ditional months (Gruwez et al. 2013). Juniper fleshy cones (hereafter fruits) ripe 20 months after pollination in next year autumn (Montesinos et al. 2012). Thus, individuals display two fruit cohorts from March to November.
Ripe fruits may not be optimal
43
Mites, Trisetacus quadrisetus (Thomas 1889) (Acari, Phytoptidae), uses seeds as a growth chambers where they feed and reproduce (Roques et al. 1984). The larvae of two moth species: Mesophleps oxycedrella (Millière 1871) (Lepidoptera, Gelechiidae) (yellow juniper moth hereafter) and Pammene juniperana (Millère 1858) (Lepidoptera, Tortricidae) (brown juniper moth hereafter) feed on juniper fruit pulp and seeds (Roques et al. 1984). The larva of a chalcid wasp, Megastigmus thuriferana Roques & El Alaoui 2006 (Hymenoptera, Torymi-dae) feeds on well-developed megagametophyte and embryo (Rouault et al. 2004). Other frequent arthropods that feed on Spanish juniper fruits include the pulp sucker scale insect
Carulaspis juniperi (Bouché 1851) (Hemiptera, Diaspididae) and the pulp eater juniper fly
Rhagoletis zerny Hendel 1927 (Diptera, Tephritidae).
Study area
Sampling site was located in Villaciervos, Soria province, Central Spain (41º 44’ N, 2º 40’ W; 1150 m.a.s.l.). Climate is Mediterranean continental, mean annual precipitation is 533 mm. Mean monthly temperatures range from 2.8 ºC in January to 20 ºC in July. The rock parent material is Cretaceous limestone covered by deep soil with small areas of exposed bedrock. Vegetation is dominated by a J. thurifera woodland with open areas covered by xeric grass-lands with shrubs, including Cistus laurifolius L. J. communis L. and Thymus zygis Loefl.
Sampling design and fruit analysis
Capítulo 2
44
ripe, all second-year fruits remaining in the branches were collected to characterize fruits and quantify pre-dispersal fruit damage.
Second monitoring was aimed to assess the timing of attack by different seed predator, quantify their impact and to characterize fruits and seeds during seed development. We sampled fruits at different stages of seed development for the 2013 and 2014 cohorts. We collected 20 fruits from all around the canopy from 10 randomly selected female juniper trees. In 2013, sampled trees were different from the trees where we followed flower and fruit development. In 2014, fruits were collected from a subset of ten trees from those twenty used to monitor fruit success, taking care of gathering fruits from branches away from those monitored. During the first and second year of each cohort, fruits were collected monthly from June to September. Sampled fruits were measured to the nearest 0.01 mm with a digital caliper and opened in the laboratory under a dissecting microscope. We counted the total number of seeds and the number of ‘full’ seeds (i.e., those showing a completely developed embryo and megagametophyte). Fruits were also inspected to detect arthropod activity. Arthropods attacking Spanish juniper fruits can be identified through their aspect and/or feeding behavior (see Appendix 1 for details). Mites’ colonies deform the seeds causing an elongation of their tip that usually stick out of the fruit surface. Yellow juniper moth larva is white with a brown cephalic capsule and a black post-cephalic mark. Brown juniper moth larva is white with a dark cephalic capsule and four black post-cephalic marks. Both moth species showed a different feeding pattern on seed and fruit pulp. Scale insects are found on the fruit surface: nymphs are yellow with red eyes and sedentary adults have circular and ellipsoidal shapes, and are white-colored. Chalcid wasp larva is white and develops inside the seeds, making a circular hole in the apical region of the fruit when emerging as adults. The larvae of juniperflies are apod, flat and yellow, and they feed only on fruit pulp.
Statistical analysis
Ripe fruits may not be optimal
45
We used linear mixed models with a binomial error structure to analyze the effect of di-fferent factors on transition success between the didi-fferent stages. We consider as success the number of fruits per branch that persisted from between two consecutive stages. Firstly, we analyzed the full transition from ‘flowers’ to ‘ripe fruits’. Besides, we analyzed the three transitions between the critical stages (flower to initial fruit, initial to unripe fruit and unripe to ripe fruit). Fixed term for all models included initial flower number per branch. All transitions from initial fruits onwards included also number of fruits per branch in the previous stage. Random component for all models included branches nested within trees and cohort.
We used fruit success monitoring dataset to test whether the initial reproductive effort affected the final number of full seeds per fruit. We used linear mixed models with Poisson error structure. Fixed terms in the model included fruit size and the number of flowers and ripe fruits per branch. Random components for the model included branches nested within trees and cohort.
Capítulo 2
46
To compare results from models using fruits collected shortly after arthropod entry with those from models based on predation rates on ripe fruits as is the common practice, we performed linear mixed models for each predispersal predator using ripe fruits to quantify damage rates and fruit traits from the branches of the 20 trees of our fruit success moni-toring. All models included the same fixed and random terms as analyzed previously: fruit size, seed number per fruit and competing predators’ infestation rates at tree level accor-ding to entry order. Tree and year were included as random factors. Moths entering during the first and second year of fruit development were analyzed separately, albeit statistical analysis was only performed for yellow juniper moth second entry. Model calculations were performed using the nlme packages in R environment (R Development Core Team, 2015).
Results
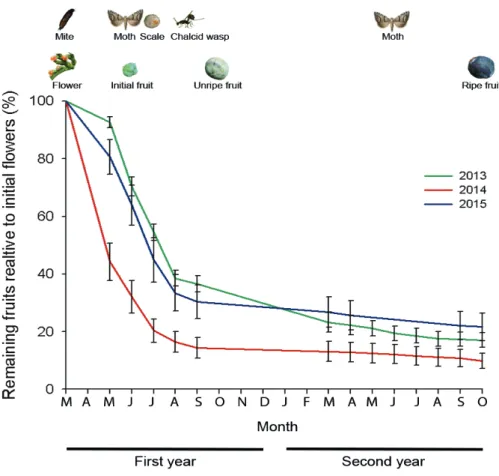
Phenological sequence of events
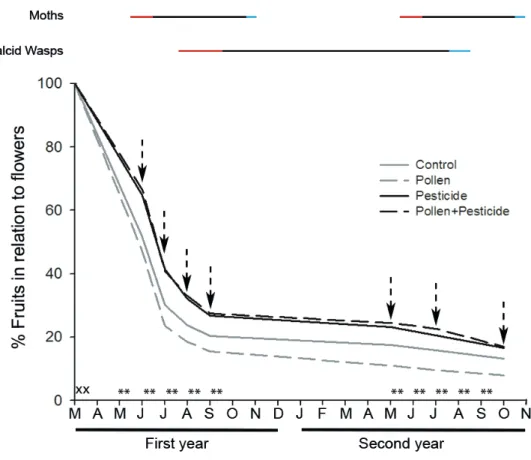
The small, scale-shaped, J. thurifera female flowers started to grow in September and fi-nished their development in next late February to early March. Pollination took place in March. Fruit growth after pollination was fast and fruits reached their final size by Septem-ber of the first year (Fig. 1). Seed fertilization took place during May and June. Seeds gained size relatively fast after fertilization and reached their final size in September of the first year. Seed coat hardening was initiated in September. Fruits and seeds matured during the second year and in late September, green fruits turned to dark blue.
Different arthropods attacked fruits during their development (Fig. 1; see Appendix 2 for details). Mites were the first species to colonize the fruits by entering the flowers during winter, waiting for seed development after pollination. As a result, mites were already inside the seeds in first year June. Mites used the seed as a growth chamber feeding on the nucellus. They had several generations and in first year September, mites ceased their reproduction (no eggs could be found within the colonies). Adults exited the seed in mid-winter and searched new female flowers to colonize.
Ripe fruits may not be optimal
47
Hatching occurred from mid-June to late July. Larvae entered the fruit leaving the egg corion glued to the fruit surface. When larva was infesting a first-year fruit, it made a gallery and entered into the seed, feeding first on seed corion and later on the megagametophyte. Larvae exited the seed in September and fed on pulp. Eggs laid in second year fruits showed a distinct behavior with larvae feeding only on pulp. In both cases, larvae left the fruit at mid-autumn of their respective year by making an irregular hole in fruit surface.
Brown juniper moth is also an annual species with only one generation per year. They had the same activity period as yellow juniper moth but they differed in feeding behavior. Fe-males laid one egg on the fruit surface during June and hatching occurred from mid-June
Capítulo 2
48
to late July. Larvae fed on egg corion after hatching. Larvae that hatched in a first years’ fruit, usually ate one seed (in some cases two). Afterwards they built a nest with seed remains, silk and fecal deposits, resting during August. The larvae reactivated in September and fed on pulp. Larvae hatching in second year fruits only fed on pulp. In both cases, larvae exited the fruit from late October to early November of their respective year.
Scale insects are annual species with only one generation per year. Adult males flied during late-May and June in search of sedentary females for mating. Females produced one to forty eggs that were laid under the scale cover. Nymphs hatched in July, colonizing first and second year fruits. Once the nymph selected the feeding site they started to suck pulp and became sedentary. Individuals passed the winter as an adult.
Chalcid wasp is an annual species with only one generation per year. Females selected first year fruits and laid one egg per seed from late July to late September. Larvae fed on the seed and stayed within the seed until late June when they pupated during a month. Adults emerged from the fruit in late July.
Fruit and full seed production
A total of 11,480 flowers were monitored in the 100 branches from 20 female juniper trees of fruit success monitoring. All the tagged branches in 2013 presented flowers. However, the number of re-flowering branches was 93 for 2014 and 52 for 2015. Flower number diffe-red between years: 2013 (3,215), 2014 (6,086) and 2015 (2,390). Largest number of flowers per branch (64.3 ± 9.8; mean ± standard error) occurred in 2014 with decreasing flowering effort in 2015 (46.0 ± 8.1) and 2013 (32.2 ± 3.3).
Ripe fruits may not be optimal
49
Probability of a flower becoming a ripe fruit increased with branch flower number (Table 1). This effect depended on the transition between flowers and initial fruits, since transi-tion probabilities between initial to unripe fruits and from unripe to ripe fruits showed no relation with either the number of flowers or fruits per branch. Fruit success monitoring revealed a higher number of full seeds within the fruits in 2014 cohort (0.51 ± 0.02 seeds per fruit) than in 2015 (0.27 ± 0.02) and 2013 (0.16 ± 0.02). Number of full seeds per fruit in ripe fruits was positively associated to fruit size and flower number per branch (Table 2).
Fruit damage
A total of 3,740 immature fruits collected from 2013 to 2015 were used to evaluate monthly damage during fruit development and 2,342 ripe fruits were used to evaluate the final fruit damage by arthropods. As much as 60.6 ± 3.5 % of the 2013 cohort fruits were damaged by any of the arthropods in September 2013. Damage rates decreased during winter, but recovered at slightly lower levels due to arthropods second entry to 54.5 ± 4.0 % in August 2014. Damage rates were lower for the 2014 cohort (39.2 ± 3.6 % in August 2014 and 37.2 ±
Fruiting % Fixed terms Estimate SE P Random SD Flower to ripe fruit Intercept -2.389 0.359 <0.001 Tree:Branch 1.267 March 1st year to Flowers per branch 0.007 0.001 <0.001 Branch < 0.001
October 2nd year Year 0.562
Flower to initial fruit Intercept 1.331 0.807 0.099 Tree:Branch 1.262 March 1st year to Flowers per branch 0.005 0.001 <0.001 Branch 2.9·10-5
June 1st year Year 1.375
Initial to final size fruit Intercept -0.627 0.241 0.009 Tree:Branch 0.881 June 1st year to Flowers per branch 3.8·10-5 0.002 0.982 Branch 1.3·10-5
September 1st year Fruits per branch 0.003 0.002 0.156 Year 0.375
Final size to ripe fruit Intercept 0.220 0.242 0.364 Tree:Branch 1.282 September 1st year Flowers per branch 0.003 0.002 0.074 Branch 3.0·10-5
to October 2nd year Fruits per branch 0.001 0.004 0.851 Year 0.298
Capítulo 2
50
3.6 % in August 2015). Estimation of damage rates using ripe fruits was considerably lower for the 2013 cohort (25.4 ± 1.8 %) but of similar magnitude for the 2014 cohort (32.4 ± 1.4 %).
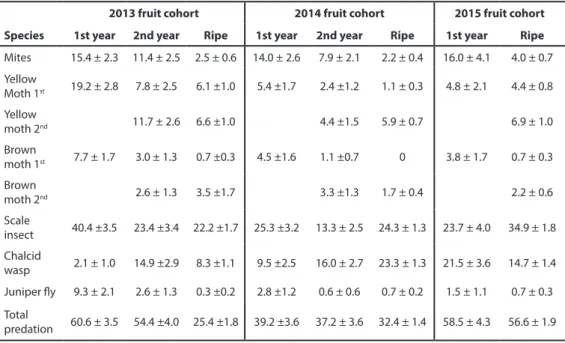
Scale insects showed the highest incidence, reaching a maximum of 40.4 ± 3.5 % in Sep-tember 2013 (Table 3). Yellow juniper moth was the next in importance, reaching 19.2 ± 2.8 % in July 2013. Mites (15.4 ± 2.3 % in June 2013) and chalcid wasps (16.0 ± 2.7 % in September 2014) showed damage rates in the same range. Brown juniper moths had lower, but still significant damage rates with a peak in 2013 August with 7.7 ± 1.7 %. Juniper flies had a moderate impact in 2013 (9.3 ± 2.1 % in September 2013), but was negligible for the other cohorts. Comparison of these values with predispersal damage rates based just on ripe fruits from fruit success monitoring revealed large underestimations of predispersal damage rates for all species but chalcid wasps.
Fruit damage monitoring revealed variation in damage rates at seasonal and inter-annual scales. At seasonal scale percentage of fruit damage peaked one or two months after ar-thropods entry and then declined for all species except for chalcid wasp that maintained similar damage levels along fruit maturation cycle. In species with two entries, a secondary damage peak occurred during second year of fruit development. Differences between maximum damage rates in first year of fruit development and second year September at the end of fruit damage monitoring were rather high especially for both moths first year entry (Yellow juniper moth: from 19.2 ± 2.8 % to 0.6 ± 0.6 %; Brown juniper moth: from 7.7 ± 1.7 % to 0.6 ± 0.6 %) scale insects (from 40.4 ± 3.5 % to 5.6 ± 1.7 %) and for mites (from 15.4 ± 2.3 % to 7.5 ± 2.4 %) and much lower for second entry of arthropods (Yellow juniper moth: from 11.7 ± 2.6 % to 2.8 ± 1.2 %; Brown juniper moth: from 2.6 ± 1.3 % to 3.3 ± 1.3
Fixed terms Estimate SE P Random SD # Full seeds in ripe fruits Intercept -2.775 8.5·10-4 <0.001 Tree:Branch 0.990
Flowers per branch 0.004 5.8·10-4 <0.001 Branch 3.2·10-4
Fruits per branch -4.2·10-4 8.3·10-4 0.615
Fruit size 0.068 8.5·10-4 <0.001
Ripe fruits may not be optimal
51
%; scale insects: 3.9 ± 1.4 % to 0.8 ± 0.8 %) and chalcid wasps (from 16.0 ± 2.7 % to 12.5 ± 3.0 %). Predispersal damage rate also showed some degree of variability between both monitored years with higher level of infestation for first year moths’ and scale insect entry in 2013 cohort (Table 3).
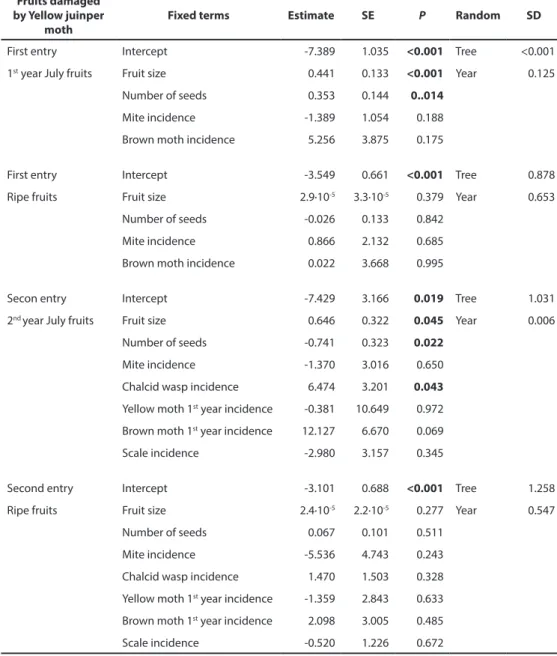
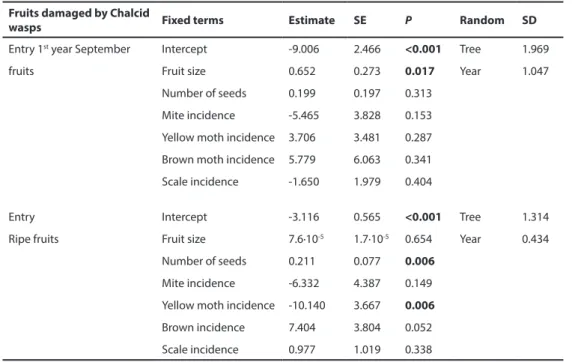
Linear mixed models for fruit preferences for the different taxa differed when the analyses were based on fruit characteristics measured shortly after oviposition compared to models based on ripe fruits. In its first entry, yellow juniper moth preferred larger fruits with more seeds (Table 4). However, the model yellow juniper moth preferences based on ripe fruits did not find any effect of either fruit size or number of seeds per fruit, even with a much larger sample size. In its second entry, yellow juniper moth showed a positive effect of fruit size, but a negative effect of number of seeds per fruit. Interestingly, a positive effect of the incidence of chalcid wasps was also found. In contrast, analysis of yellow juniper moth second entry preferences based on ripe fruits detected no effect of fruit size, seed number or the incidence of chalcid wasps (Table 4). When coexistence at fruit level was evaluated presence of moth yellow juniper moth was marginally positively associated to chalcid wasp
2013 fruit cohort 2014 fruit cohort 2015 fruit cohort Species 1st year 2nd year Ripe 1st year 2nd year Ripe 1st year Ripe
Mites 15.4 ± 2.3 11.4 ± 2.5 2.5 ± 0.6 14.0 ± 2.6 7.9 ± 2.1 2.2 ± 0.4 16.0 ± 4.1 4.0 ± 0.7 Yellow
Moth 1st 19.2 ± 2.8 7.8 ± 2.5 6.1 ±1.0 5.4 ±1.7 2.4 ±1.2 1.1 ± 0.3 4.8 ± 2.1 4.4 ± 0.8
Yellow
moth 2nd 11.7 ± 2.6 6.6 ±1.0 4.4 ±1.5 5.9 ± 0.7 6.9 ± 1.0
Brown
moth 1st 7.7 ± 1.7 3.0 ± 1.3 0.7 ±0.3 4.5 ±1.6 1.1 ±0.7 0 3.8 ± 1.7 0.7 ± 0.3
Brown
moth 2nd 2.6 ± 1.3 3.5 ±1.7 3.3 ±1.3 1.7 ± 0.4 2.2 ± 0.6
Scale
insect 40.4 ±3.5 23.4 ±3.4 22.2 ±1.7 25.3 ±3.2 13.3 ± 2.5 24.3 ± 1.3 23.7 ± 4.0 34.9 ± 1.8 Chalcid
wasp 2.1 ± 1.0 14.9 ±2.9 8.3 ±1.1 9.5 ±2.5 16.0 ± 2.7 23.3 ± 1.3 21.5 ± 3.6 14.7 ± 1.4 Juniper fly 9.3 ± 2.1 2.6 ± 1.3 0.3 ±0.2 2.8 ±1.2 0.6 ± 0.6 0.7 ± 0.2 1.5 ± 1.1 0.7 ± 0.3 Total
predation 60.6 ± 3.5 54.4 ±4.0 25.4 ±1.8 39.2 ±3.6 37.2 ± 3.6 32.4 ± 1.4 58.5 ± 4.3 56.6 ± 1.9
Capítulo 2
52
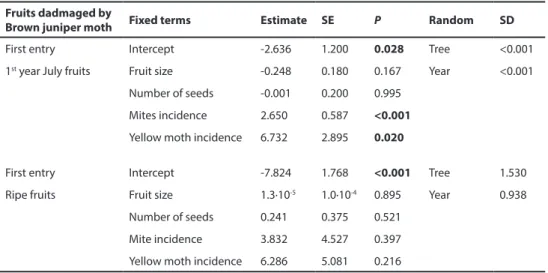
presence (Χ2 = 7.81; P = 0.0501). The linear mixed model for brown juniper moth first entry
revealed that their incidence increased in trees with higher incidence of mites and yellow juniper moths. When this analysis was performed on ripe fruits the effects of the incidence of mites or yellow juniper moths were not detected (Table 5). Interestingly, whereas the presence of mites was strongly positively associated to brown juniper moth infestation at fruit level (Χ2 = 29.13; P < 0.001), both moths showed a random pattern (Χ2 = 0.20; P =
0.978). No significant effects were detected for scale insects’ preferences during their first entry (Table S2). However, when this analysis was performed on ripe fruits a positive effect of number of seeds per fruit was found (Table S2). Finally, predation monitoring revealed the preference of chalcid wasps for larger fruits. The same analysis performed on ripe fruits found a positive signal of seed number per fruit and of yellow juniper moth fruit damage rate (Table 6).
Discussion
Monitoring of Spanish juniper fruit development revealed that fruit set was largely determi-ned by initial reproductive effort. Initial investment in flowers enhanced ripe fruit set levels and number of full seeds per fruit. Several arthropods damaged fruits and depredated the seeds during the developing period, greatly diminishing ripe crop size. The sequential attack of mites, moths and scale insects exerted a strong pressure during the first five mon-ths of fruit development. Selective abortion of damaged fruits caused that an important proportion of damaged fruits were not detected in ripe fruits. Moreover, fruit preferences by arthropods differed when analyses were based on developing compared to ripe fruits.