Uncertainty in stock assessments, spatial distribution and habitat modelling of two small pelagic fish species, sardine (Sardina pilchardus) and anchovy (Engraulis encrasicolus), in the Mediterranean from late autumn spanish acoustic surveys
187
0
0
Texto completo
(2)
(3) D DOCTO ORAL THESIS T S 2015 Doctorral Pro ogramm me of Biology B y UN NCERTA AINTY IN STOC CK ASSES A SMENT TS, SP PATIAL L DIS STRIBUTION AN ND HABIT TAT MO ODELLING OF O TW WO SM MALL PELAG GIC FISH SP PECIES S, SAR RDINE (Sardi dina piilchard dus) AND A AN NCHOV VY (En ngrauliis enc crasico olus), IN THE T ME EDITER RRANE EAN FROM M LA ATE AUTUM MN SP PANISH H ACOU USTIC SURVE EYS Vollume I of I M Pilar Tugorres Ferrà M.. Dirrector/a a: Magdalena a Iglesiias Marrroig Tuttor/a: Antoni A Martín nez Tab berner. Do octor by y the Univers U sitat de e les Ille es Bale ears.
(4)
(5) A la meva família, als meus amics, i a tots els que d’alguna manera han fet aquest treball posible.
(6)
(7)
(8) “We shall s not ceaase from ourr exploration n And the t end of all our explorring Will be b to arrive where w we staarted And know k the plaace for the fiirst time” (T.S.. Eliot). “I alm most wish I haadn’t gone down that rabbit-hole r - and yet - and ye et it’s ratther curious, you know, this so ort of life!” ( (Alice, in Alicce in Wonde erland, by Lewis Carrol) C.
(9)
(10)
(11) Agradecimientos/Agraïments Esta tesis no habría sido posible sin la participación de tanta gente que de alguna manera han aportado su granito de arena tanto al trabajo y a la persona que soy. Así que estas líneas son para agradecerles el haber estado en algún momento ahí, en mi camino. En primer lugar, cómo no darle las gracias a mi madre. Una GRAN persona en mayúsculas a la que además de la vida le debo tantas cosas!! Entre ellas, el haber podido introducirme en el mundo de la investigación (con sus más y sus menos pero siempre bonita) y haber conocido a tanta gente extraordinaria que han formado o forman parte de esta historia. A mis compañeras/os de trabajo, de café y a mis amigas/os de mis primeros años en el mundo científico en el IMEDEA, a los que guardo tanto cariño. Gracias a todas/os por tantos momentos, gracias por haber estado ahí. Besos a todas/os, os deseo lo mejor! También quiero darle las gracias al grupo de acústica al completo del Centro Oceanográfico de las Islas Baleares – Instituto Español de Oceanografía (COB-IEO). En especial, a Juan Miquel y a Dolors por su trabajo como técnicos de acústica, sin los cuales nada habría sido posible. Agradezco a Juan, con especial cariño, sus lecciones de teoría acústica durante mi primera campaña de acústica, sin las cuales el McLennan & Simmonds, piedra filosofal de la acústica, habría sido un hueso duro de roer!!! A Marilin, como directora de tesis y jefa de grupo, por su paciencia, sus consejos y su apoyo. Compartir siempre es enriquecedor. Así pues, gracias a los acústicos y geoestadísticos de diferentes países con los que me topado en reuniones internacionales el haber podido intercambiar quebraderos de cabeza en momentos difíciles y me han hecho descubrir que había alguien ahí fuera con “dilemas” parecidos a los míos. A Marco Barra, Roberto Gramolini, Marianna Giannoulaki, Fabio Campanella…Un gracias enorme a Marianna y a su equipo y colaboradores en Creta, por su hospitalidad y su colaboración (ευχαριστώ! #efaristó!). A Pierre Petitgas, por ser un referente en geoestadística. Gracias al IEO por haber hecho todo esto posible! Quiero agradecer a las tripulantas/es, científicas/os y marineras/os que han participado y posibilitado las campañas ECOMED, en las cuales he basado mi trabajo. No quiero olvidarme del equipo de administración del COB, duro y no siempre reconocido trabajo el suyo pero indispensable para que todo esto funcione. Esto no habría sido lo mismo sin mis compañeras/os de viaje, en el sentido más literal y en el más literario de la palabra. Gracias por las pequeñas y a las grandes enseñanzas. A mis compañeras/os y amigas/os del COB-IEO, a las horas de cuerda, a las charlas de café y diversión.Gracias por demostrarme vuestro apoyo en tantos momentos y por vuestros consejos. A todas las personas que han formado parte de las horas de diversión y de los momentos de estrés. A mis compañeras/os de despacho, del presente y del pasado y sobre todo a las del presente por soportar mis últimos y más duros meses. Gracias por vuestra inconmensurable paciencia. I, per suposat, a sa meva família, a sa pradina, a mon pare, en es meu germà, a sa meva germana, i a en Lleonet i a en Felipe, per estar al meu costat en tot moment. Sense oblidar a meva familia peninsular-illenca, n’Ainarona, na Chispa, na Loa i en Jesús, sempre aprop! A ses amigues i germanes d’adopció, na Marga i na Mar, n’Aina i n’Antònia. No fa falta que vos digui quant vos estim. Gràcies a tots!. i.
(12) A Esporles, es rentadors des Jonquet y Santa Catalina, por diversos y entrelazados motivos. A Santa Bárbara y a las latas de sardina.. ii.
(13) Derived publications Chapters 2 to 5 presented in this thesis have been partially published or submitted for publishing in the form of scientific papers. In particular, Chapter 2 is based on the paper: “Tugores, M.P., Iglesias, M., Díaz, N., Oñate, D., Miquel, J. and Giráldez, A. 2010. Latitudinal and interannual distribution of the European anchovy (Engraulis encrasicolus) and sardine (Sardina pilchardus) in the western Mediterranean, and sampling uncertainty in abundance estimates. ICES Journal of Marine Science, 67: 1574-1586” A reduced version of Chapter 3 has been sent for publication as: “Tugores, M.P., Iglesias, M., Oñate, D. and Miquel, J. Spatial distribution, sampling precision and survey design optimization with non-normal variables: the case of anchovy (Engraulis encrasicolus) recruitment in the Spanish Mediterranean waters. Progress in Oceanography. Submitted” Chapter 4 have been partially published within a broader paper, co-authored with people from other Mediterranean countries and including data from these other countries as: “Tugores, M.P., Giannoulaki, M., Iglesias, M., Bonanno, A., Ticina, V., Leonori, I., Machias, A., Tsagarakis, K., Díaz, N., Giráldez, A., Patti, B., De Felice, A., Basilone, G., and Valavanis, V. 2011. Habitat suitability modeling for sardine Sardina pilchardus in a highly diverse ecosystem: the Mediterranean Sea. Marine Ecology Progress Series, 443: 181-205” Chapter 5 have as well been partially published within a broader paper, coauthored with people from other Mediterranean countries and including data from these other countries as: “Giannoulaki, M., Iglesias, M., Tugores, M.P., Bonanno, A., Patti, B., De Felice, A., Leonori, I., Bigot, J.L., Ticina, V., Pyrounaki, M.M., Tsagarakis, K., Machias, A., Somarakis, S., Schismenou, E., Quinci, E., Basilone, G., Cuttitta, A., Campanella, F., Miquel, J., Oñate, D., Roos, D., and Valavanis, V. 2013. Characterizing the potential habitat of European anchovy Engraulis encrasicolus in the Mediterranean Sea, at different life stages. Fisheries Oceanography, 22 (2): 6989”. iii.
(14) iv.
(15) Llist of acronyms and abreviations AIC - Akaike’s Information Criterion ANOVA - Analysis Of Variance AUC - Area Under the ROC CFP – European Common Fisheries Policy CHLA - sea surface chlorophyll concentration CI – confidence interval CUFES – Continuous Underway Fish Egg Sampler CV – Coefficient of Variation DSP – Digital Signal Processing EDSU – Elementary Distance Sampling Unit EU – European Union EVA – Estimation of the VAriance FAO – Food and Agriculture Organisation of the United States GAMs - Generalised Additive Models GCV - Generalized Cross Validation GFCM - General Fisheries Commision for the Mediterranean Sea GLOBEC - Global Ocean Ecosystem Dynamics programme GSA - Mediterranean Geographical Subarea I2D – intrinsic geostatistics in 2 dimensions ICES – International Council for the Exploration of the Sea IEO – Instituto Español de Oceanografía MDT - maximization of the specificity-sensitivity sum MPA – Marine Protected Area NASC – Nautical Area Scatering Coefficient NC – Northern Current NS – Northern subarea (defined in Fig. 2.1) PAR - photosynthetically active radiation PDF – probability distribution function PRV – prevalence. v.
(16) PSH – potential spawning habitat ROC - Receiver Operating Characteristic curve RSH - realised spawning habitat SGMED - Study Group on Mediterranean Fisheries SLA - sea level anomaly SS - Southern subarea (defined in Fig. 2.1) SSS - sea surface salinity SST - sea surface temperature T1D – transitive geostatistics in 1 dimension TS – target strength Tukey HSD - Tukey’s Honestly Significant Difference test. vi.
(17) Contents Agradecimientos/Agraïments................................................................................................ i Derived publications ............................................................................................................... iii Llist of acronyms and abreviations ..................................................................................... v Summary/Resumen/Resum ................................................................................................... 1 Chapter 1. General Introduction ........................................................................................... 5 1.1 Motivation and definition of the problem ................................................ 7 1.2 The Spanish Mediterranean waters: geomorphological and hydrographical characteristics ..................................................................................... 9 1.3 Sardine (Sardina pilchardus Walb.) and anchovy (Engraulis encrasicolus L.): notes about the species ecology, biology and geographical distribution ...................... 11 1.4 Echointegration acoustic surveys for pelagic fish stock assessment: the case of the Spanish Mediterranean surveys .......................................................... 14 1.5 Structure and objectives of the present work ......................................... 19. Section I. Spatial distribution, random error and the sampling scheme ............ 21 Chapter 2. Latitudinal and inter‐annual distribution of European anchovy (Engraulis encrasicolus) and sardine (Sardina pilchardus) and sampling uncertainty in the Western Mediterranean .................................................................. 23 2.1 Introduction ................................................................................. 25 2.2 Materials and methods .................................................................... 26 2.3 Results ........................................................................................ 29 2.4 Discussion .................................................................................... 37. Chapter 3. Two dimensional spatial distribution of anchovy (Engraulis encrasicolus) in the Spanish Mediterranean waters to estimate sampling precision and optimize sampling design ........................................................................ 41 3.1 Introduction ................................................................................. 43 3.2. Material and methods ..................................................................... 45 3.3 Results ........................................................................................ 50 3.4 Discussion .................................................................................... 61. Section II. Environmental variables and habitat ......................................................... 66 Chapter 4. Identifying the potential habitat of sardine (Sardina pilchardus) spawners and sardine eggs in the Mediterranean Sea ............................................. 68 4.1 Introduction ................................................................................. 70 vii.
(18) 4.2 Materials and Methods ..................................................................... 71 4.3 Results ........................................................................................ 76 4.4 Discussion .................................................................................... 88. Chapter 5. Identifying the potential habitat of European anchovy (Engraulis encrasicolus) recruitment in the Mediterranean Sea ................................................ 96 5.1 Introduction ................................................................................. 98 5.2 Materials and methods .................................................................... 99 5.3. Results ...................................................................................... 101 5.4 Discussion ................................................................................... 110. Chapter 6. General Discussion ..........................................................................................117 6.1 Stock assessment, spatial distribution and fisheries management ............... 120 6.2 Habitat identification, suitability and overlap ....................................... 122. Chapter 7. General conclussions ......................................................................................125 7.1 Stock assessment, precision and spatial distribution of sardine and anchovy in the Northern Spanish Mediterranean continental shelf waters ........................ 127 7.2 Habitat identification of sardine and anchovy in late autumn .................... 128. Bibliography ...........................................................................................................................129 ANNEX. Main erros associated to abundance estimates of pleagic fish stocks the case of late autumn Spanish Mediterranean acoustic surveys (2003‐2006) ..129. viii.
(19) Summary/Resumen/Resum. Summary Small pelagic fish are species that live in the water column and have little relationship with the sea bottom. During the day they may form schools with feeding, defence or energetic efficiency purposes and disperse during the night. The importance of small pelagic fishes within the marine ecosystem rely in the proportion of biomass they represent and the clue function they develop as intermediate links in the energy transfer between lower and upper levels of the trophic chain. Their populations are particularly sensitive to environmental fluctuations (Cole and McGlade, 1998; Lloret et al., 2004) and frequently highly exploited by commercial fisheries. These may occasionally collapse the stocks affecting both the marine ecosystem and the fisheries they sustain. In the Mediterranean, almost 50% of the total annual landings are attributable to small pelagic fishes (Lleonart and Maynou, 2003). In the Western Mediterranean Sea, sardine (Sardina pilchardus) and anchovy (Engraulis encrasicolus) are the two most important species in terms of landed biomass and commercial interest (Lleonart and Maynou, 2003). Despite their importance little is known about the spatial distribution of these stocks or about the relationship between the spatial distribution and environmental variables. Since the ‘90s acoustic surveys are annually performed in the Spanish Mediterranean continental shelf in late autumn (Abad et al., 1998 a,b) coinciding with anchovy’s recruitment and the beginning of the spawning season for sardine (Abad and Giráldez, 1993; Giráldez and Abad, 1995). Although the main goal of these surveys is to estimate abundance and biomass of sardine and anchovy, data about the whole pelagic community has also been gathered. Furthermore, in the last decade it was detected an increasing appearance of other small and medium-sized pelagic species and the application of a multi-species approach to Mediterranean fisheries assessment was advised (Lleonart and Maynou, 2003). The present work is structured in two sections. The first section of the work will analyse the spatial distribution (1D and 2D) of sardine and anchovy by means of geostatistical techniques, both transitive and intrinsic methods (Matheron, 1969). Special attention will be paid to the estimation of the uncertainty associated to abundance estimations and, concretely, the uncertainty caused by sampling scheme which is thought to be one of the main contributors to random error (ICES, 1998). The second section will explore the environmental factors that drive the presence or absence of anchovy and sardine in late autumn. Satellite environmental data as sea surface temperature, salinity, chlorophyll-a, photosintethic active radiation, sea level anomalies or bottom depth will be related to the presence-absence of sardine and anchovy stocks through Generalised Additive Models (GAM) to try to depict the relationships that may be found between both (Bellido et al., 2008; Giannoulaki et al., 2008). The spawning area of sardine will also be studied in order to model the presence-absence of sardine eggs and try to understand the evolution of their stocks.. 1.
(20) Resumen Los pequeños peces pelágicos son especies que viven en la columna de agua y que tienen poca relación con el fondo marino. Durante el día forman bancos de peces con el objetivo de alimentarse, defenderse de potenciales depredadores o por motivos de eficiencia energética, dispersándose durante la noche. La importancia de los pequeños pelágicos en el ecosistema marino radica en la proporción de biomasa que representan y en la función clave que desempeñan como eslabones intermedios, transfiriendo energía entre los niveles tróficos inferiores y superiores de la cadena trófica. Sus poblaciones son particularmente sensibles a las fluctuaciones ambientales (Cole and McGlade, 1998; Lloret et al., 2004) y frecuentemente se encuentran en un régimen de explotación elevado por parte de las pesquerías comerciales. Esto puede comportar de manera ocasional el colapso de sus poblaciones explotables, afectando al ecosistema marino y a las pesquerías que las mantinen. Casi el 50% del total de la biomssa anual desembarcada en los puertos del Mediterráneo provienen de los pequeños peces pelágicos (Lleonart and Maynou, 2003). En el Mediterráneo Occidental, la sardina (Sardina pilchardus) y el boquerón (Engraulis encrasicolus) son las dos especies más importantes, tanto en términos de biomasa como capturada como por su interés comercial (Lleonart and Maynou, 2003). A pesar de su importancia, la distribución espacial de las poblaciones explotables es poco conocida así como también lo es la relación de la distribución espacial con los parámetros ambientales. Desde principios de los años 90, se llevan a cabo campañas acústica en la plataforma continental española del Mediterráneo a finales de otoño (Abad et al., 1998 a,b), coincidiendo con el reclutamiento de la anchoa y la época de puesta de la sardina (Abad and Giráldez, 1993; Giráldez and Abad, 1995). A pesar de que el principal objetivo de estas campañas es la estimación de la abundancia y de la biomas de la sardina y el boquerón, datos de la comunidad pelágica en conjunto también se recogen periódicamente. Además, a lo largo de la última década se ha detectado una aparición incremental de otras especies de pequeños y medianos pelágicos y por tanto, se aconsejó la aplicación de un enfoque multiespecífo en la evaluación de pesquerías del Mediterráneo (Lleonart and Maynou, 2003). El presente trabajo se estructura en dos seciones. La primera sección del trabajo analizará la distribución espacial (1D y 2D) de la sardina y el boquerón por medio de técnicas geoestadísticas, tanto transitivas como intrínsecas (Matheron, 1969). Se prestará especial atención a la incertidumbre asociasda a las estimaciones de abundancia y, concretamente, a la incertidumbre asociada al diseño de muestro que se considera uno de los factores que contribuyen con mayor intensidad al error aleatorio (ICES, 1998). La segunda sección explorará los factores ambientales que condicioinan la presencia o ausencia de sardina y boquerón a finales de otoño. Datos ambientales obtenidos de satélite, como la temperatura superficial del mar, la salinidad, clorofila, radiación fotosintéticamente activa, anomalía del nivel del mar y la profundidad del fondo marino se relacionarán con la presencia-ausencia de las poblaciones explotables de sardina y boquerón mediante modelos aditivos generalizados (GAM, del acrónimo inglés) para determinar las relaciones existentes (Bellido et al., 2008; Giannoulaki et al., 2008). El hábitat de la puesta de sardina será igualmente estudiada para modelar la presencia-ausencia de huevos de sardina e intentar entender la evolución de las especies explotables.. 2.
(21) Resum Els petits peixos pelàgics són espècies de peixos que viuen a la columna d’aigua i que tenen poca relació amb el fons marí. Durant el dia formen bancs de peixos amb l’objectiu alimentar-se, defensar-se en front potencial depredadors o per eficiència energètica, dispersant-se durant la nit. La importància dels petits pelàgics a l’ecosistema marí radica en la proporció de biomassa que representen i en la funció clau que exerceixen com esglaons intermedis, transferint energia entre les nivells inferiors i superiors de la cadena tròfica. Les seves poblacions són particularment sensibles a les fluctuacions ambientals (Cole and McGlade, 1998; Lloret et al., 2004) i freqüentment es troben baix un règim d’explotació elevat per part de les pesqueres comercials. Això pot comportar de manera ocasional el col·lapse de les seves poblacions explotables, afectant a l’ecosistema marí i a les pesqueries que sostenen. Gairebé el 50% del total de la biomassa anual desembarcada als ports del Mediterrani prové dels petits peixos pelàgics (Lleonart and Maynou, 2003). Al Mediterrani Occidental, la sardina (Sardina pilchardus) i l’aladroc (Engraulis encrasicolus) són les dues espècies més importants, tant en termes de biomassa capturada com pel seu interès comercial (Lleonart and Maynou, 2003). Malgrat la seva importància, la distribució espaial de les poblacions explotables és poca coneguda així com també ho és la relació d’aquesta distribució espaial amb variables ambientals. D’ençà de principis dels anys 90, es duen a terme campanyes acústiques a la plataforma continental del Mediterrani espanyol a finals de la tardor (Abad et al., 1998 a,b), coincidint amb el reclutament de l’aladroc i amb l’època de posta de la sardina (Abad and Giráldez, 1993; Giráldez and Abad, 1995). Tot i que el principal objectiu d’aquestes campanyes és l’estimació de l’abundància i de la biomassa de la sardina i l’aladroc, dades del conjunt de la comunitat pelágica també es recolleixen. A més a més, al llarg de la darrera dècada s’ha detectat una aparició incremental d’altres espècies de petits i mitjans pelàgics i per tant, es va aconsellar l’aplicació d’un enfoc multiespecífic a l’avaluació de les pesqueres del Mediterrani (Lleonart and Maynou, 2003). El present treball s’estructura en dues seccions principals. La primera secció del treball analitzarà la distribució espaial (1D i 2D) de la sardina i l’aladroc per mitjà de tècniques geoestadístiques, tant transitives com intrínseques (Matheron, 1969). Es prestarà especial atenció a la incerteza associada a les estimacions d’abundància i, concretament, a la incertesa originada pel disseny de mostreig que és considerat un dels factors que contribueixen amb més intensitat a l’error aleatori (ICES, 1998). La segona secció explorarà els factors ambientals que condicionen la presència o absència de sardina i aladroc a finals de tardor. Dades ambientals obtingudes de satèl·lit, com la temperatura superficial de la mar, la salinitat, clorofil·la, radiació fotosintèticament activa, anomalies del nivell de la mar i la profunditat del fons marí es relacionaran amb la presència-absència de les poblacions explotables de sardina i aladroc mitjançant models additius generalitzats (GAM, de l’acrònim anglès) per determinar les relacions existents (Bellido et al., 2008; Giannoulaki et al., 2008). L’hàbitat de la posta de la sardina serà igualment estudiada per tal de modelar la presència-absència d’ous de sardina i intentar entendre l’evolució de les seves poblacions explotables.. 3.
(22)
(23) Chapter 1. General Introduction.
(24)
(25) 1. General IIntroduction n. Ch hapterr 1. Geenerall Intro oductio on 1.1 Motivaation and d definittion of th he probllem The pelagiic ecosystem is the co ombination of physical and biological interac ctions that occur in th he water co olumn of se eas and oce eans worldw wide, with little conta act or depe endence with the sea bottom. The T pelagic ecosystem is the most productiive in the oceans o and d representss around a 95 % of th he world occean production (Marg galef, 1993). Its explo oitation playys an imporrtant sociall and econo omical role,, contributiing to up to o a 50% of the world landings l of marine spe ecies (Fréon et al., 20 005). Altogether with aquaculturre, fisherie es provide direct d and indirect em mployment to over 660 0–820 millio on people, or about 10–12 percent of the world d’s populattion (Food and Agricculture Orga anisation; FAO, F 2012).. Within the pelagic ecosystem,, small pelagic fishes play a kkey role in n the functtioning of the ecosystem as the ey transfer energy fro om plankto on to the upper u levells of the trophic chain n (Cury et al., a 2000). They also sustain imp portant fish heries world dwide mak king them highly h valua able in eco onomical te erms as we ell (Cury et al., 2000 0; Shannon et al., 200 00; Daskalovv, 2002). Their T relativve low position in the food web,, their reprroductive sttrategy of producing p la arge quantiities of egggs over exte ended areass and spaw wning period ds, as long as their sh hort life sp pan make th heir popula ations highlly dependen nt on the su uccess of th heir recruitment and to t environm mental varia ability and changes c (Co ole and McG Glade, 1998 8; Lloret et al., 2004). Since the industrialissation of th he ‘tradition nal’ small-sscale fisherries, collapsses of small pelagic fish fisherie es’ have offten been reported, r e e.g. the Pe eruvean anc chovy ns) in the 70 0s or the European E anchovy (Eng graulis encrasicolus) in the (Engrraulis rigen North hern Spain n, where the fishe ery was closed be etween 20 006 and 2010. 2 Overrexploitatio on was poin nted out ass a threat for fisherie es in 1945--46 for the e first time in the fra amework of o FAO (Ca addy and Cochrane, C 2 2001) and it has bee en an interrnational co oncern for many m years since then. Marine fiisheries are threate ened worldwide ma ainly thro ough incre easing overe exploitation n but also by climatte change which stro ongly intera act (FAO, 2010; 2 Brander, 2007)). Fishing reduces th he age, sizze, and th he biodiverrsity of marine m ecosyystems ma aking popullations and ecosystem ms more vu ulnerable a and sensitivve to clima ate change e (Branderr, 2007). Climate ch hange has effects o on demogra aphy, distribution and d stock stru ucture as well w as varriations in the t productivity, structure and composition c n of the eco osystems on n which the e species de epend (Bran nder, 2007). Marine ressearchers and managers have the e challenge to develop p and imple ement mech hanisms tha at are able to guaranttee a sustainable explo oitation of marine fish heries (Pauly et al., 2002) 2 while ensuring the persiste ence of the population ns. Actuallyy, the European Common Fishe ery Policy (CFP) hass as a main goal tthe sustain nable explo oitation of aquatic liviing resources in econo omic, sociall and enviro onmental te erms, while e applying the preccautionary principle and progre essively im mplementing an ecosyystem apprroach to fish heries mana agement. A successsful manag gement nee eds to be based on n a good knowledge e and unde erstanding of o the reality that is trying to deal with. In this contexxt, one firstt step is to know the state of th he population that is to be mana aged. Acou ustic surveyys are used worldwide e as a stand dard method d to perform m annual asssessments of the stoc cks of pelag gic fish spe ecies (Simm monds and MacLennan, M 2005). One e importantt reason for this 7.
(26) 1. General IIntroduction n is tha at acoustic surveys pro ovide high quality data to estima ate abundan nce and bio omass of sm mall pelagicc fish specie es at a high h spatial re esolution en nabling as w well high qu uality analyysis of the spatial s distrribution of the speciess under stud dy. In the recent yearss ecosystem m-based managementt has been n given gro owing atten ntion (e.g. García et al., 2003;; Pikitch et al., 2004 4); notwithstanding siinglespecies stock assessments a s remain the base fo or any postterior attempt to ma anage wholle ecosystems. The un ncertainty on o the relia ability (or precision) p o of single-sp pecies stock k assessmen nts has som metimes bee en used to excuse the e implementation of fiishing restrrictions tha at can be economical e ly painful (Pauly et al., a 2002). Thus, routtinely estim mation of th he precision n of single-sspecies stoc ck assessme ent is crucia al to accom mplish a conscious ma anagement and is of special s inte erest in the e internatio onal comm munity since e late 80s in the fra amework of the Interrnational Council C of the Sea (ICES). Furth hermore, asssessing the e uncertain nty in the esstimates ca an become clue to pro operly unde erstand the e temporal evolution of the po opulation. Obviously, O the intere est in imprroving the accuracy a an nd precision n of those abundance and bioma ass estimates by reducing the asssociated errrors cannott be ignored d. Besides ab bundance estimates e a and its asso ociated erro ors, the kn nowledge of o the ns and the influence i o environmental facto of ors on spatiial distributtion of fish population the stock s variab bility is as well w importtant for con nservation biology b and manageme ent of any threatened d species. Knowing K wh here a spec cies is foun nd is crucia al to design n, for insta ance marine e protected d areas (M MPAs) or clo osed areas. Acoustic surveys provide high quality sp patial data of pelagicc fish spec cies that can c be app plied to sp patial distribution mo odelling or,, in combin nation with h environm mental data a, to mode el the habittat of the species (e.g g. Giannoula aki et al., 2007; 2 Giann noulaki et a al., 2011; Be ellido et all., 2008). The ability to pre edict the effects off measuress and policies for stock mana agement is an interestting subjectt to be developed. Changes in the distributiion of the species, s in its abunda ance and biomass b tha at might be e driven byy environm mental forcing and/or climate c cha ange would ideally nee ed to be kn nown in ord der to be ab ble to elabo orate a dyn namic mana agement which w potentially would facilitate e to achieve e the main n goal of susstainability. Modelling the habitatt of a species might be b useful in n order to p predict pote ential chan nges in the distributio on of a spe ecies origina ated by ch hanges in th he habitat (e.g. incre ease/reducttion, displa acement) which w migh ht be drive en by the cclimate cha ange. Thuss, above and beyond d the mere e stock asssessment and a the esstimation of o its preciision, habittat modelling becomes more and d more inte eresting. P Potential ha abitat mode elling (or suitability s m maps) can offer o some insight in this topic; in spite of o the limittations of any a particular modelling approac ch. Further, gaining kknowledge about a the suitable s ha abitats for a species might m be useful to ide entify area as that could be interresting to be protected d for the sp pecies, for instance i byy establishin ng MPAs. In the Me editerranea an Sea, tw wo small fissh pelagic species Eu uropean sa ardine (Sard dina pilcha ardus Walb b.) and European E anchovy a (E Engraulis e encrasicoluss L.) domiinate the fiisheries’ catches. Theyy representted 32-44% of the tota al landings in the Mediterranean between 20 003 and 20 009 (FAO, 2013). 2 In th he Western Mediterran nean, sardiine and an nchovy are as well th he most im mportant sp pecies in tterms of la anded biom mass and co ommercial interest (L Lleonart an nd Maynou, 2003) beiing anchovyy the main n target spe ecies of the e purse seiiners in the e area (Perrtierra and Lleonart, 1996; Abad d et al., 199 98a). Beside larrge pelagicss, sardine and a anchovvy are the most m abund dant fish sp pecies in th he pelagic communityy. In the Sp panish Mediterranean continenta al shelf anc chovy 8.
(27) 1. General IIntroduction n and sardine represented between 38 3 and 67 % of the estimated biomass in n the acou ustic surveyys between 2003 and 2009 2 (unpublished datta, IEO). In the contin nental shelff of the Ba alearic basin (Northerrn Spanish Mediterran nean waterrs), sardine e and anch hovy repressented 65--92 % of the t small and medium sized pelagic sp pecies comm munity estimated in th he acoustic surveys in the period 2003-2006 (Tugores et e al., 2010 0). Their sto ocks have been declining since the 90s, although anchovy show wed a slight recover in n 2001 and 2003 (Girá áldez et al.., 2006a,b; Palomera et al., 2007 7). In addittion, there e is increasing conccern aboutt the posssible over--exploitatio on of anch hovy’s recru uitment sin nce an imp portant fraction of th he fishing effort relie es on imma ature fish (Pertierra and Lleonartt, 1996). Sizze at first maturity m occcurs at 12.5 and 12.7 cm for ma ales and fem males respe ectively wh hile the min nimum land ding size is 9 cm (Perttierra and Lleonart, L 19 996). The prese ent study focus f the attention to t the pre ecision of tthe abunda ances estim mates of th hese two species s pro ovided by late l autum mn acoustic surveys in n the Span nish contine ental shelf,, analyse the t spatial structure of their stocks and relate r theirr habitat to o the enviro onmental ch haracteristic cs present in the samp pled area in n late autumn (mid No ovember-miid Decembe er).. 1.2 The Sp panish Mediter M rranean waters: geomo orpholog gical and d hydrog graphicaal characcteristicss The study area is loca ated in the Western Mediterrane M an basin an nd comprise es the Span nish Mediterranean co ontinental shelf s along the Iberia an Peninsula, between n the Strait of Gibralttar and the Spanish-Frrench borde er (Figure 1.1a). The ccontinental shelf is na arrow, often n less than 6 nauticall miles (nm m, 1 nautica al mile = 18 852 m) between the Strait of Gibraltar G an nd the Cap pe of Paloss. Northwa ards, the co ontinental shelf wide ens till the surrounding gs of the Eb bro River, where w the maximum w width is rea ached (33 nm). In the Catalan coast, con ntinental sh helf is narrow (less tthan < 14 nm), inden nted by su ubmarine ca anyons. Th he total are ea of the continental shelf is about a 8,513 3 nm2 (29 199 Km2). aters, the North N The circullation is dominated byy the entrance of the Atlantic wa Curre ent (NC) a cyclonic c alo ong-slope frront in the north-westtern Medite erranean and the outfllow of large e rivers. Th he NC flow ws along the e continenttal shelf fro om northea ast to south hwest (Font et al., 19 988), carryying water from f the Gulf G of Lion n to the Ca atalan coastt, and eve entually rea aches the Alboran Se ea (Millot, 1999; Fontt et al., 1988). Stron ng northerly winds, tyypical in wiinter, may intensify the t NC (Pin not et al. 2002). 2 The wide contiinental she elf and fressh-water ru un-off from m the Rhon ne and the Ebro riverrs further characteris c e the area a. In the Alboran A Sea a the wate er circulation is domiinated by less saline Atlantic water w entering the ba asin througgh the Stra ait of Gibra altar. The surface s Atla antic waterrs, although h relativelyy nutrient-p poor, are re elated with mesoscale features, such s as turbulent mixiing, anticycclonic gyress, meanderrs and eddie es (Estrada a, 1996) gen nerating up pwelling alo ong the narrow contin nental shelf and resullting in a local enricchment of nutrients and a primarry productiion in the area (Champalbert, 1997). Two o anticyclon nic gyres are generate ed in the A Alboran Sea a, the mostt western gyyre is quasii permanen nt while the e eastern on ne is more variable (M Millot, 1999 9). The photicc zone, or volume v of the t water column c whe ere the ligh ht penetrattes, is thought to be the most pro oductive in seas and oceans o and is generallyy between 0 and 100-2 200 m dep pth. Moreo over, prima ary producttion is high her in coa astal areas with riverrine outflow ws and alsso in place es where water w circu ulation gen nerates nuttrient upwe elling from the sea bo ottom to th he sea surfa ace. In Wesstern Meditterranean, these 9.
(28) 1. General IIntroduction n areass of increa ased producctivity are found in th he vicinity of the maiin rivers, Rhône R and Ebro, E and in the Alborran Sea due e to water masses m circu ulation. Mean chloroph hyll-a conccentration can be ussed as a proxy p to reveal r the areas of higher priimary production (Figu ure 1.1b).. Figurre 1.1. Span nish Mediterrranean wate ers: (above) main wate er circulation n patterns in i the studyy area (NC: Northern N currrent; solid liines showing g the most sttable circula ation; dashed d lines showing more va ariable curre ents) and (b bottom) mean chloroph hyll-a concen ntration (mg g m-3) D 20 008 from Se eaWiFS satelllite data (A Acker & Lepttoukh, between Januaryy 2003 and December ason.gsfc.nasa.gov/Giova anni) 2007)). http://rea. 10.
(29) 1. General IIntroduction n. 1.3 Sardin ne (Sarrdina pilchardu p us Wallb.) and d anch hovy (En ngraulis encrasico e olus L.): notes about a th he speciies ecolo ogy, biollogy and d geograaphical distribut d tion Pelagic fisshes are known to perrform daily vertical miigrations, u usually gath hering in large schoolss during the e day and re emaining diisperse during the nigh ht (Fréon et e al., 2005 5). Small pe elagic fishess, considering as such h species with adults’ length between 10 an nd 30 cm, are commo only foragin ng species of o high eco ological valu ue, due to their functtion as botttom-up ene ergy transfe erors in the trophic cha ain (Cury ett al., 2000). The biolo ogy of thesse species, i.e. short life-span and a reproductive stra ategy (producing large e quantitie es of eggs over exte ended area as), makes these spe ecies especially sensiible to envvironmentall changes (Bakun, ( 199 96). Their recruitment is particu ularly sensiible to enviironmental variability and change es (Cole and McGlade,, 1998; Llorret et al., 2004). 2 Abru upt increase es and decreases of the populatiions of sma all pelagic fishes f are frequent f an nd may afffect both the fisheries they susttain and th he structure e and functtioning of the ecosysstem (Cury et al., 20 000; Shanno on et al., 2000; Dask kalov, 2002 2), triggerin ng the colla apse of the fisheries, e.g. the Pe eruvean ancchovy (Engraulis rigen ns) in the 70 0s or, more e recently, European anchovy a (En ngraulis enccrasicolus) in the North hern Spain,, where the e fishery wa as closed be etween 200 06 and 2010 0. In the ca ase of anch hovy, their populationss have been n recently recognised r as vulnerab ble to long term incre ease in tem mperature by the Global Ocea an Ecosyste em Dynamics program mme, GLOB BEC (Barang ge et al., 2010). Geog graphical an nd bathyme etric distrib bution European sardine (Sardina pilcchardus) ha as a less extended e ge eographical and bathyymetric disstribution than t Europe ean anchovvy (Engrauliis encrasico olus). Sardiine is found d in the No orthern hem misphere fro om Norwayy to Senegal while ancchovy is fou und in the both b hemisspheres, fro om Iceland and Norw way to South Africa. B Both specie es are distributed in the Easterrn Atlanticc and the Mediterran nean (Figurre 1.2). In n the Mediterranean they are widespread through the w e Western and Centra al Basins ass well as in the Northe ern Aegean Sea (Figure e 1.2). The bathymetrric distributtion of Euro opean sardiine is betw ween 25 and d 100 m de epth (White ehead, 1990 0) whereas anchovy ca an be found d up to 400 0 m depth (Schneider, 1990). (a). (b b). graphical disstribution off the main small s pelagicc fish specie es in the We estern Figurre 1.2. Geog Meditterranean: (a) sardine,, Sardina pilchardus p and (b) anchovy, Engra aulis encrassicolus (Modiified from Fiishbase; sourrce: Whitehe ead et al., 1988 and Whiitehead, 199 90).. 11.
(30) 1. General IIntroduction n Spaw wning Sardine sp pawns from m autumn to spring with a peak in wintter, in Jan nuaryFebruary (Palom mera and Olivar, 199 96; Olivar et al., 20 001, 2003) with maxiimum abun ndance of spawners s a sea surfa at ace temperatures of about 18ºC C (Oliver, 1957; Oliva ar et al., 2001). Sp pawning occcurs main nly in insh hore waterrs with highest conccentration of o sardine eggs at de epths below w 100 m (O Olivar et al., 2001, 2003). 2 Conttrarily, anch hovy spawn ns in the wa armest mon nths (Whitehead et al., 1988; Tsikliras et al., a 2010). In the We estern Mediiterranean anchovy spawns s between Aprill and Octo ober with a peak in summer, in June-A August, in waters wiith sea su urface temp perature be etween 15ºº and 22ºC (Palomera a, 1989, 19 992; García a and Palom mera, 1996 6). The spaw wning of sa ardine and d anchovy is associatted to wa ater enrich hment proce esses. In th he Western n Mediterra anean, watter enrichm ment in win nter occurs at a globa al scale and is relate ed to verticcal mixing, which pro oduces the recirculatio on of nutriients from the bottom m to the euphotic e zo one. Verticcal mixing is enhance ed by wind ds and thus it is though ht to favourr sardine re ecruitment (Palomera et al., 2007 7 and referrences therrein). During summer water w enric chment occcurs mainly at a local scale and is related to river ru unoff, fronttal systemss and upwe ellings (Fon nt et al., 1987; Estra ada, 1996; Arnau A et al., 2004). An nchovy spaw wning and early-life e sttages are cllosely relatted to the ese areas, especially to river outflows o as anchovy spawns in n the proxiimity of rivvers, even in regions of low disc charge (Palo omera, 198 89, 1992; García G and Palomera, 1996; Oliva ar et al., 2001). 2 Durin ng the spaw wning seaso on anchovyy may ntually ente er lagoons, estuaries or lakes, as it has good d tolerance to low salinities even (5-41 1ppt); conttrarily, sard dine larvae do not to olerate low salinities (Sabatés et al., 2001). Matu urity The length at first maturity m (L L50) of sardiine ranges from 10.9 cm to 16.8 cm (mainly age 0 groups and 1) 1 across itss distributio on area. L500 is generally smaller in the Mediterranean Sea than in i the Atla antic Ocean n (Silva et al., 2006 and refere ences there ein). In the e Spanish Mediterrane M ean waters,, L50 of sarrdine was e estimated to t be 15.00 0 cm in the e coasts of Valencia (A Andreu et al., a 1950) and a betwee en 12.6 and d 13.8 cm in n the Malag ga Bay, Albo oran Sea (A Abad and Giiráldez, 199 93). Regard ding anchovvy, L50 varie es between 5.6 and 13 3.7 cm across its distribution area a (Koranten nk, 1993; Urriarte el al., 1996). In n the Weste ern Mediterrranean, it was w estimatted to be 12.5 and 12..7 cm for males m and females resp pectively, while w the minimum m leg gal size in tthe area is 9 cm (Girá áldez and Abad, A 1995; Pertierra and a Lleonarrt, 1996). Feed ding Small pe elagic fishe es are planktivorou p us speciess, generally feeding g on phyto oplankton and/or a miccro- and me eso-zooplan nkton (Freó ón et al., 2 2005). Euro opean anch hovy is know wn as a zoo oplanktivoro ous fish mosstly eating copepods, but in the Black and Azov A Sea itt has also be een found to t feed spo oradically on phytoplan nkton (Palo omera et al., 2007 an nd referencces therein). European sardine has h a more e heterogen neous diet preying on zooplankto on (e.g. cla adocerans or o decapodss) as well a as phytoplan nkton like diatoms (C Costalago and a Palome era, 2014). Both small pelagic ffish species are capa able of sw witching between filte er- and pa articulate-ffeeding, ussing partic culate feeding for bigg ger preys. This abilityy, known fo or long for European a anchovy an nd for sardiines over th he world, was w only re ecently described for European ssardine (Bod de et al., 2004; 2 Garrido et al., 2007, 2 2008; Nikoliouda akis et al., 2012). 2. 12.
(31) 1. General IIntroduction n In the Bayy of Biscayy, it has be een suggessted that anchovy a hass higher trophic plastticity than sardine, byy foraging more offsh hore and a wider rangge of prey sizes (Chouvelon et al., 2014, 2015). How wever, labo oratory exp periments indicate tha at, in osition to anchovy, sardine s can n retain ve ery small preys, i.e.. nanoplankton, oppo allow wing them to reprodu uce under lower productivity conditions c ((Garrido ett al., 2007 7). Differencces have be een observe ed in the nuber of gilll rakers bettween anch hovies and sardines in n the Atlan ntic and be etween sard dines from the Atlanttic and sarrdines from m the Medite erranean, suggesting s differencess in their diiets that sh hould further be invesstigated (C Costalago and Palo omera, 201 14). Nevertheless, the tradittional chara acterization n of sardine e as a phyto oplanktofag gous fish ha as been put into questiion as long as, in term ms of preyy contributiion to carb bon intake, zooplankton seems to t be more e importantt in sardine e’s diet (Garrido et al., a 2008a; Nikoloudakkis et al., 2012; 2 Costa alago and Palomera, P 2 2014). Although in i other are eas, anchovvy eggs are frequently found in sa ardine’s stomach (Garrrido, 2003;; Bachiller, 2012; Garrrido et al.., 2008) an nd although h cannibalism is frequ uent in othe er areas forr other speccies of the genus Engrraulis, in the e Mediterra anean Sea intraguild i p predation an nd cannibalism have not n been reported neitther for anc chovy nor for f sardine or has been considere ed negligiblle (Tudela and a Palome era, 1997; Fréon F et all., 2005; an nd referencces therein;; Costalago and Palom mera, 2014).. In fact, almost no fish f eggs are a found in sardine and anchovy stomachs in the e Mediterra anean (Palo omera et all., 2007 and d reference es therein; Bacha B and Amara, A 200 09; Borme et e al., 2009 9; Garrido and a van derr Lingen, 20 014 and refferences th herein). The e only exce eption in th he 1940s an nd 1950s, when w ancho ovy eggs we ere found in i Mediterrranean sard dine’s stom machs (Massutí and Olivver, 1948; Andreu and d Rodríguezz-Roda, 195 51), suggestting a control role of sardine s on anchovy’s a p population (Palomera et e al., 2007 7 and refere ences there ein). A density-depen ndent mechanism and d/or differrential disttribution in n the wate er column may explain the alm most absenc ce of fish eggs sardin ne and anc chovy stom machs in the Mediterrranean (Valldés et al., 1987; Ga arcía and P Palomera, 1996; Palom mera et al., 2007; Garrrido and va an der Linge en, 2014). Ecolo ogical role in the ecossystem In the Wesstern Meditterranean the bulk of the t marine biomass co omes from these two small pelagic speciess, sardine and a anchovvy. As forag ging specie es, they pla ay an impo ortant role in transferrring energyy between the t lower and a the upp per layers of o the troph hic chain. In the NW Mediterran nean sardin ne plays a ‘wasp-waisst’ flow con ntrol, affeccting upperr and lowerr levels of the t trophic chain, while anchovy would disp play a ‘botttom-up’ co ontrol thus having a higher h impa act on the upper leve els (Coll ett al., 2008 8). In the Catalan C Sea adult anch hovy consumed up to a 20% of tthe zooplan nkton production acccessible to o anchovy aggregatio ons (Tudella and Pa alomera, 1997), highllighting the e importantt role of th his species in channe eling energyy of the pe elagic food webs to higher trophiic levels. Migra ations Pelagic fisshes are kn nown to pe erform long g migrationss for spawn ning, feeding or winte ering. In th he Iberian Peninsula sardine s mig grates in sp pring from Portuguese e and Canttabrian watters to Gallician coastt (Carrera and Porteiiro, 2003). In the case of anch hovy, it overwinters in the North East of the e North Sea and move to the Soutth for the rest r of the year. In the Gulf of Vizcaya, anchovy a mo ove from so outh to norrth in summ mer (Huret et al., 200 07). In the Eastern Me editerranea an, anchovyy from the Azov Sea overwinter o in the Black Sea (Borssa et al., 20 004). However, migration is highly va ariable and might dep pend of the e species de ensity as w well as on peculiaritie p s of the arrea where the speciess is found. In the We estern 13.
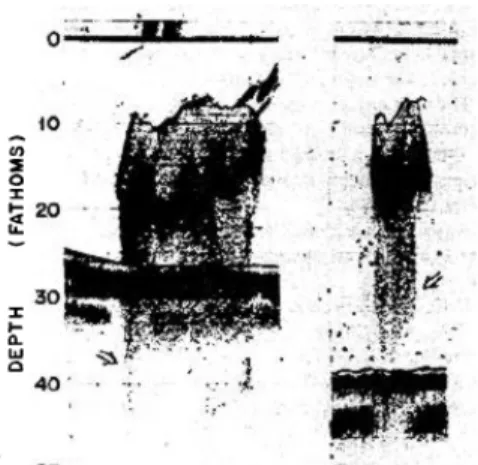
(32) 1. General IIntroduction n Mediterranean large l migra ation patterrns have not been desccribed eithe er for sardiine or for anchovy. a Sa ardines werre relativelly sedentarry and remain in refugee areas when they are scarcce while th hey can be b highly migratory m w when they are abundant, colon nizing coo oler waterss (Lluch-B Belda et al., a 1992). Semi-ind dependent subpopu ulations of sardine seem s to coexist c in Western Mediterrane M ean with small migrations betw ween them m and no significant migration along sho ore (Ramon n and Castrro, 1997). In the case of anchovyy, although the season nality of the e landings might m be in ndicative off migratory movementts it could also a be indiicating a re eduction in fleet accessibility to the stock by, e.g. a seasonal re eduction of school size e (Fréon, 1989), or an n increase in catchabiility during spawning season like e occurs witth other pe elagic species (Jorgensen et al., 2004; Skare et et al., 20 005). Seaso onality of landings and d relationsh hips with environmenttal factors In the We estern Mediterranean,, sardine landings pe eak in autu umn and sp pring, when n the population is mainly m comp posed of 1--2 year old individualss (Lloret et al., 2004 4) while an nchovy land dings peak in spring-ssummer (Pa alomera, 1992), when n the popu ulation is mostly m com mposed of 1-year old individualss (Pertierra a and Lleo onart, 1996 6). Anchovy landings arre positively correlate ed with rive erine outflow ws, cold wiinters and intense Norrth-westerlly winds, att one year lag (Martín n et al., 200 08; Palome era et al., 2007 2 and re eferences therein). t A positive co orrelation between b wiind and anc chovy recru uitment we ere also fo ound in other areas of the world, w e.g. South-We estern Atlan ntic, Northe ern Chile and a the Black Sea. In the case of o sardine, w wind seeme ed to have e a positive e influence on landing gs but not river r outflo ows (Palome era et al., 2007 and references r therein).. 1.4 Echoin ntegratio on acousstic surv veys forr pelagicc fish sttock asseessmentt: the casse of thee Spanish h Meditterranean n survey ys Echointegrration acou ustic surveyys have wid dely been used u for pe elagic fish stock assesssment sincce the late 1970s, afte er it was firrstly proposed by Drage esund and Olsen O (1965 5).Nowadayys echointe egration is the t most ex xtended me ethod for a assessing marine m pelag gic fish stoccks worldw wide (e.g. To oresen et al., a 1998; Dalen D et al., 2003). In ndeed acou ustic surveyys are annually conduccted to esttimate the stocks of p pelagic fishes by manyy internatio onal organissms of fishe eries manag gement, e.g g. Internatiional Counc cil for the Exploration n of the Se ea (ICES) and a the Ge eneral Fish heries Com mmission for the Mediterranean (GFCM). Th he reasons for this arre that aco oustic surve eys allow direct d samp pling at sea a, providing g fishery in ndependentt estimatess of the ab bundance of o the species. Moreovver, they sa ample a larg ge volume of o the wate er column a and allow a wide spatiial coverage e in a relatiively short time span. Echointegrration surveys use sound to dettect organissms that are in the water w colum mn by mea ans of an echosounde e er. It is a methodolog gy that com mbines aco oustic samp pling with pelagic fishing trawlss. Acoustic c sampling provides a an index off fish abun ndance whille pelagic trawls t provide biologic cal informa ation of the e species ass well as in nformation of the com mposition off the comm munity. The e echosoun nder emit pulses p of so ound throug gh the wate er, verticallly from upward to the bottom, and receive the echo oes generate ed by the organisms o e encountered d in the sou und path (F Figure 1.3a). The back kscattered echoes e are received, amplified and a graphiccally plotte ed as echog grams (Figu ure 1.3b). However, the t acquisittion of high h quality da ata was not possible till the intro oduction of digital sig gnal processing into scientific ecchosounderrs by late 1980s 1 and the t establisshment of standard s calibration methods m (Foo ote et al., 1987).. 14.
(33) 1. General IIntroduction n Late autumn Sp panish Mediiterranean acoustic a surveys: histo orical insigh ht In the Spanish Mediiterranean waters aco oustic survveys for pe elagic fish stock assesssment starrted to be conducted c i the 1983 in 3. In 1997 th hese surveyys were ann nually perfo ormed in la ate autumn n (Novembe er-Decembe er), with th he main aim m of obtain ning a recru uitment ind dex for Euro opean ancho ovy (Engrau ulis encrasiccolus) and a an estimatiion of the population p o European of n sardine (SSardina pilcchardus). In n 1997, the surveys Byy that time, echogram ms were recorded r in paper and a any po osterior esstimate im mplied analyysing the in ntegram ma anually on a paper. The installlation in early e 2000s of a scien ntific echossounder with digital signal s proce essing allo owed fasterr and morre accurate e processin ng of the data making it and bioma possiible for the first tim me to estim mate the abundance a ass, not on nly of sardiine and ancchovy but of o the acco ompanying small and medium sized pelagic c fish species: Meditterranean horse macckerel (Tra achurus mediterrane m eus), blue jack mack kerel (Trachurus pictu uratus), rou und sardine ella (Sardin nella aurita a), bogue (B Boops boop ps), Atlanticc mackerel (Scomber scombrus) s and Atlantiic chub macckerel (Sco omber colia as). Their commercial c interest iss lower and d also theiir abundancces tended, but they have also o importance in the ecosystem m. Further, in the la ate 2000s these patte erns might have changed (Iglesia as et al., 2008). 2 Thuss, the initia al objective e was broadened to estimate e the e stock of coastal c pela agic resourrces in an a attempt to move to an n ecosystem m approach.. A hull-mounted splitt-beam SIMRAD EK500 echosound der operate ed from 1990 to 2005 5 and in 2006 2 it wa as substitutted by a SIMRAD S EK K60 echosounder with h the transsducers mo ounted on n a protru uding keel (Figure 1.4). 1 The SIMRAD EK500 E echo osounder op perated at two freque encies, i.e. 38 and 12 20 kHz, whiile SIMRAD EK60 operated at 5 frequencies f s, i.e. 18, 38, 3 70, 120 0 kHz and 200 2 kHz, bu ut in both cases the 38 3 kHz freq quency was used to estimate e th he density and abund dance of pe elagic fish. Regarding the sampliing scheme e, a systema atic samplin ng design cconsisting of o zigzag transects t w applied till 2001. In was n 2002 no acoustic a surrvey was pe erformed ass long as re esources we ere devoted d to study the Prestig ge oil-spill in i Costa da a Morte (Ga alicia, Spain n). In 2003 3 the sampling design n was changed to a systematic sampling design d consiisting of pa arallel equid distant tran nsects perp pendicular to t the bath hymetry in order to ha ave the sam me sampling g intensity in i all the ba athymetric strata. Abundance e and biom mass estima ates of sard dine and anchovy a obttained from m the late autumn Spanish aco oustic surve eys were annually su ubmitted tto the fish heries assesssment gro oups of th he General Fisheries Commissio on for the e Mediterra anean (GFC CM). With the t aim of standardisiing the diffferent acou ustic surveyys performed in European Meditterranean waters w and make them m comparable, anotherr acoustic survey erformed in n summer 2009 2 coincid ding with an nchovy spaw wning. Thiss year startted being pe was the t last when late auttumn acousttic surveys were perfo ormed.. 15.
(34) 1. General IIntroduction n. Fish schools Waterr column n. Sea bottom Figurre 1.3. Schem me of the em mission and reception r of sound by an n echosounde er (left) (mo odified from FAO, 1981) and samp ple echogram m acquired on the 3rd d of Decem mber 2006 in the surro oundings of the Ebro Rive er mouth (rig ght).. el installed at a the Fiigure 1.4. Prrotruding kee hu ull of the R/V R Cornide e de Saaved dra in 20 006 (image by b Joan Miqu uel Batle).. Data a collection Acoustic data was collected on board the rese earch vesse el “Cornide de Saavedra”, 66.7 7 m length (Figure 1.5 5a) during the end of November N a and beginniing of Dece ember in th he context of annual surveys of pelagic fissh stock assessment in n the Span nish Mediterrranean waters (ECOM MED). The su urveys coinccided with the recruittment perio od of ancho ovy and with the spaw wning seaso on for sard dine in the area (Abad d and Girálldez, 1993; Giráldez and Abad, 1995). Data D were continuoussly recorde ed at consttant speed of 10 knotts along parrallel equid distant transects place ed perpendiicular to ba athymetry (Figure 1.5 5B) and using a scienttific split-b beam echossounder, SIM MRAD EK50 00 or EK60, working att 38 kHz (7º x 7º beam m angle, 1 ms pulse le ength, maxiimum ping rate, powe er 2000 W). Acoustic sampling was w perform med from sunrise to sunset covering the en ntire continental shelf between 30 3 and 200 m depth. T The echosou under was calibrated prior to ea ach survey with w a 60 mm m cooper sphere, folllowing stan ndard techniques (Foo ote et al., 1987). Intter-transectt distance was 4 nau utical miless (nm hereafter) were e the contin nental shelff is narrow and 8 nm where w the ccontinental shelf. 16.
(35) 1. General IIntroduction n is wiide (Figure 1.5b). Transects were numbered from 1 to o 128 from m North to South S (Figu ure 1.5b). (a). (b). nide de Figurre 1.5. a) R/V “Corn Saave edra” (imag ge by Joan Miquel Batle e) and b) acoustic sa ampling schem me applied during d the ECOMED E surve eys.. Mid-waterr pelagic fisshing trawlls aimed att obtaining the propo ortion of sp pecies prese ent in the area were performed after sunsset, betwee en 29 and 2 221 m deptth, at 3.5-4 4.5 knots (n nm h-1) forr approxima ately 1 hou ur length. The T net wa as 12 m vertical open ning and 22 mm codend. In our sttudy area, small s pelagics are know wn to aggre egate durin ng the day while disag ggregate an nd ascent through t the e water collumn during the nightt (Fréon et al., 1996). Fish schoo ols are not found f durin ng the nightt (Iglesias et e al., 2003) but dispe erse aggrega ations. Thus, trawls were w carried d out on the ese disperse e fish egations an nd in placess where, du uring the da ay, pelagic fish schoolls were fou und in aggre conccentrations high enoug gh to be trawled t and d trying to o cover all the areas with diffe erent types of echotra aces. The number off pelagic fiishing traw wls made ra anged betw ween 35 and d 71 in the period p 2003 3 to 2009. Fishes of the t speciess found in the t trawl ca atches were e sized to tthe 0.5 cm total lengtth (LT) low wer and, fo or the mosst abundantt species, they were also weighted. Weig ght-length relationship r ps were fittted to an ex xponential model using the root mean square method.. Data a processing g Acoustic data d were integrated d at every Elementarry Distance e Sampling Unit (EDSU) which was w set to 1 nm using g the Miriax x Echoview w software. The amou unt of echo o-integrated d nautical miles was about 1200 0-1300 per year, exce epting for some yearss when the e weather conditions c did not alllow the covverage of tthe entire study area. Echoes off fish schools are consiidered for echointegra e ation while other unwa anted echo oes are exccluded, i.e e. bottom, plankton layers and noise. The result off the echo ointegration n, Nautical Area Scatte ering Coeffficient (NASSC; m2 mn-22) or sA follo owing MacL Lennan et al. a (2002), a measure of o the inten nsity of the e echo refle ected by su urface unit correspond ding to the pelagic co ommunity. It was divided into sp pecies using the comp position of the trawl catches and applying Nakken and a Domassnes metho od for multtiple species (Nakken and a Domasn nes, 1975) but b using backscatteriing cross-se ection by le ength class rather than n the mean backscatte ering cross-ssection, of the form:. 17.
(36) 1. General IIntroduction n. sA i sA. wi bs. w i. (Eq.. 1.1). bs. i. where wi is i the propo ortion of th he species i in the catches (in nu umber of fisshes), and σbs is the backscatteri b ing cross-se ection for each e specie es and lengtth class, instead of th he mean bacckscattering cross-secction used in n Nakken and Dommassnes (1975). The back kscattering cross-sectio on per speccies and len ngth class (σ σbs) is relatted to the target t stren ngth (TS) ass follows:. bs 10. TSS 100 . (Eq. 1.2). where TS is the tarrget strength in decib bels (dB) and a is com mputed with h the form mula TS=20 log l LT + b20 (Simmond ds and MacL Lennan, 200 05) where b20 is empirrically obtained and diiffers amon ng species and a LT is the total length class. A b20 of -72.6 dB was used for sa ardine (S. pilchardus), p , anchovy (E. ( encrasiccolus) and round sardiinella (S. aurita) a (Degnbol et al., a 1985); -68.7 dB for f Atlanticc chub macckerel (Sco omber colia as), Medite erranean horse macckerel (T. mediterrraneus), h horse mac ckerel (Tracchurus tracchurus) and d blue jack k mackerel (Trachurus picturatu us) (Lillo ett al., 1996 6); -67 dB for bogue e (B. boop ps) (Foote, 1980); -84.9 dB Atlantic mac ckerel (Scom mber scom mbrus) (Edwards et al., 1984 4) and -67 7.5 dB for blue wh hiting (Micrromesistiuss poutassou u) (Foote et e al., 198 86). Thus, sAi for ea ach speciess was obtained.. 18.
(37) 1. General IIntroduction n. 1.5 Structu ure and objective o es of thee presentt work The prese ent work iss structured d in two main m section ns, accordiing to the main objecctives. The e first secttion is aimed at estim mating the e random e error (precision) assocciated to the abundance estim mates of anchovy a and sardine in the Sp panish Mediterranean waters in late autum mn while analysing th he structure e of the sp patial distribution of the popula ations. In particular, p the error derived d fro om the sam mpling desig gn will be estimated as it is su upposed to be one off the main contributo ors to random error (ICES, 1998)). The relattionship be etween the spatial disstribution of o the species and the e sampling design (tyype and de ensity) will allow us tto estimate e the preciision of the e abundance e estimatess originated d by the sam mpling desiggn. Data de erived from m acoustic su urveys will serve to this purpose. The second section will w explore e the relatio onship betw ween the ha abitat of sa ardine and anchovy a with environm mental facttors, as well as sardine e eggs. Speccifically, bo ottom depth and sate ellite enviro onmental data d will be b combine ed with datta derived from acou ustic surveyys. Accordin ng to the modelling, m the t potentiial habitat of anchovyy and sardiine through hout the Mediterranean n will be ass well analyysed. The work will include e an Annex x containing g a revision of the pote ential sourc ces of errorr that are present in acoustic surveys and a a quan ntitative analysis of their prese ence and im mportance in the Spa anish Mediterranean acoustic a surrveys, especially referrring to the e derived sa ardine and anchovy a abu undance estimations. Following the presen nt chapter of o General Introductio on (Chapter 1); Chapter 2 inclu udes the an nalysis of th he sample variance v off abundance e estimatess of sardine e and anch hovy by mea ans of transsitive geosttatistics in one dimenssion as well as the analysis ; Chapter 3 is of th he one dim mensional spatial disstribution of o these populations p devo oted to ana alyse the tw wo dimensio onal spatia al distribution of sardiine and anc chovy by means m of intrinsic geo ostatistics; Chapter 4 and 5 willl include p potential ha abitat mode elling of sa ardine and anchovy byy means of Generalise ed Additive Models (GA AMs). Finallly, Chapte er 6 will include a General G Disscussion an nd Chapterr 7 the Ge eneral Concclusions.. 19.
(38)
(39) Section I. Spatial distribution, random error and the sampling scheme.
(40)
(41) Chapter 2. Latitudinal and inter-annual distribution of European anchovy (Engraulis encrasicolus) and sardine (Sardina pilchardus) and sampling uncertainty in the Western Mediterranean.
(42)
(43) 2. Lattitudinal & inter-annual in distribution. Ch hapterr 2. Latitu udinall and interr-annu ual disstributtion of o Eurropean n anch hovy (Engrauulis enccrasicollus) an nd saardine (Sarddina pilcharddus) an nd sam mpling g unceertain nty in the Westeern Meediterrraneaan 2.1 Introdu uction Pelagic eccosystem is one of the e most productive in the world as it repre esents abou ut a 95% of the world w ocea an producttion (Marg galef, 1993 3). In We estern Mediterranean, small pellagic fish and concre etely two species, ssardine (Sa ardina pilch hardus) and d anchovy (Engraulis encrasicolu us), are th he two mo ost importa ant in term ms of landed d biomass and a commercial intere est (Lleonarrt and Mayn nou, 2003). They repre esented bettween 65 and 92 % of the biomasss estimated d by acousttic techniqu ues in the Balearic Se ea coastal pelagic fissh commun nity during the period d 2003 to 2006 (unpublished da ata, Instituto Español de Oceano ografía, IEO O). As small pelagic fiishes, they hold a key role in the e energy tra ansfer betw ween the low wer and the e upper leve els of the trophic t cha ain. The se ensitivity off their recrruitment to o environmental varia ability and changes c (Co ole and McG Glade, 1998 8; Lloret ett al., 2004)) along with h their shorrt life span may trigg ger abrupt increases and decrea ases of the eir populations which may affecct both the fisheriess that sustain and th he structurre and fun nctioning off the ecosyystem (Curry et al., 2000; 2 Shannon et al.,, 2000; Dasskalov, 200 02). Landin ngs of anch hovy and sa ardine decllined from the middle e 1990s an nd 2005 in the contin nental shelff of the Balearic Sea (G Giráldez ett al., 2006a a,b; Palome era et al., 2 2007). More eover, there e is an inccreasing co oncern abo out the posssible over-exploitatio on of anch hovy’s recru uitment sin nce an imp portant fraction of th he fishing effort relie es on immature fishe es (Pertierra a and Lleon nart, 1996). The size at a first matturity of an nchovy occu urs in the area a at 12.5 5 and 12.7 cm for malles and fem males respecctively, whiile the miniimum legall size in the e area is 9 cm c (Girálde ez and Abad d, 1995; Perrtierra and Lleonart, 1996). In Western Mediterra anean anch hovy spawn ns between April and October with w a peak k in summer (June-Aug gust) with sea s surface e temperatu ures between 15º and 22ºC (Palo omera, 198 89, 1992; García and a Palome era, 1996)). During summer water w enricchment occcurs at a lo ocal scale which w is mainly m related to riverr runoff, frrontal syste ems or upw welling of de eep waters favoured by b the prese ence of sub bmarine can nyons (Fontt et al., 198 87; Estrada a, 1996; Arn nau et al., 2004). 2 Anch hovy spawning is assoc ciated to th hese areass of high hydrographi h ic enriching processe es (Palomerra, 1989, 1992; Garccía and Palo omera, 199 96; Olivar et e al., 2001). River outflows o pla ay an impo ortant role in the distrribution pattterns of an nchovy earlyy-life stage es as anchovvy spawns in the proxiimity of rivers even in regions of low rivver discharrge (García and Palom mera, 1996 6). Sardine spawns frrom autum mn to sprin ng with a peak in w winter (Jan nuaryFebruary) (e.g. Palomera and Olivar, 1996; Olivar et al., 2001, 2003 3) when su urface temp perature falls to 20ºC.. For sardin ne spawnerrs, the max ximum abun ndances and d the highe er probabillities of suiitable habitat have been observved in wate er tempera atures below w 18ºC (Oliiver, 1957; Olivar et al., a 2001; Tugores et al., a 2011). T The spawniing of sardiine occurs mainly m in in nshore wate ers with the e highest concentratio on of eggs found f in de epths below w 100 m (Oliivar et al., 2001, 2003 3; Tugores et e al., 2011).. 25.
(44) 2. Latitudin nal & inter-an nnaual distriibution Since the ‘1990s, an n acoustic survey s was annually performed p to estimate e the abun ndance and d biomasss of sardiine and anchovy a along the entire Sp panish Mediterranean continental c l shelf (ECO OMED surveyys). The surrvey was ca arried out in n late m was to estimate e an nchovy’s re ecruitment as well ass the autumn and itss main aim abun ndance and biomass off sardine an nd of the other accomp panying pelagic fish sp pecies as Mediterrane M ean horse mackerel (Trachurus mediterrraneus), ro ound sardiinella (Sard dinella auriita) and bo ogue (Boopss boops). Understandi U ing the spa atial distrib bution of sm mall pelagic fish speccies, its variation in time t as we ell as the p precision of o the estim mates may be helpful in assessing g fisheries managemen m nt and wou uld benefit other fields of knowle edge. Howe ever, published information aboutt the spatia al distributiion of the pelagic p fish stocks pre esent in thiss region is scarce s (Perrtierra and Castellón, 1987; Abad d et al., 1998 a, b; Igllesias et al., 2003; Be ellido et al., 2008) as stated Olivvar et al. (2003). ( Mo oreover, th he need of o incorporation of error e estim mation to stock assesssment esttimates is crucial in order to develop an nd implem ment sustainable mana agement po olicies for the main pe elagic fisherries. A detailed analysis of the error e (syste ematic and d random)) associate ed to echo ointegration n acoustic surveys, an nd especiallly in multi-specific a areas, is no ot an easy task due to t the variety of sourrces of unc certainty in n this kind of surveys,, e.g. transsformation of acousttic intensitty into fissh density, trawl sam mpling, sp pecies migration, bubbles of airr in the su urface of th he water column, c mo ovement of the resea arch vessell, hydrographical cond ditions or vessel avoiidance (e.gg., Aglen, 1994; Simm monds and MacLennan n, 2005). Thus, we fo ocus on the e estimation n of the ovverall samp pling uncerttainty as itt is thoughtt to be one e of the ma ain contribu utors to random errorr in acoustiic surveys (Internation nal Council for the Exp ploration off the Sea – ICES, 1998 8). Transitivve geostatisstics in one e dimension (T1D) (M Matheron, 19 965) enable es an unbia ased estima ation of the sampling variance in spatially autocorrellated data, such as accoustic datta, and it is i applied to the cum mulated abundance byy transect. This meth hod was ussed for the e first time e in fisheries acoustics in the ‘1990s and d was consiidered suittable for monitoring m total quan ntities of stock in regularly sp paced samp pling schem mes (Petitgas, 1993), which are e generally used in accoustic surrveys. Furth hermore, in n our studyy area the morphology m y of the pla atform and the distrib bution of th he transectts makes th he analysiss of one dimensional data a goo od proxy of o the latitu udinal distrribution of the t species. The main objective of the pre esent studyy is to provide an esstimation of the preciision associiated to ab bundance esstimates fo or the two main smalll pelagic sp pecies in We estern Mediterranean Spanish wa aters, ancho ovy and sarrdine derive ed from aco oustic data. The latitu udinal spatial distribution of the ese two spe ecies and th heir variatiion in time are also analysed. a So ome remark ks on accurracy of the estimates are provided as well as a revisio on of the pros p and cons of the T1D T geostatistical apprroach. Atte ention is paid to the po ossible implications in stock assesssment.. 2.2 Materiaals and methods m s Four yearss of acoustic data colle ected in the e context of o annual su urveys of pe elagic fish stocks asse essment in the Spanish h Mediterra anean wate ers (ECOMED D) from 2003 to 2006 6 was used d. The stud dy area co omprises th he continental shelf of the Sp panish Mediterranean Sea S (Figure e 2.1) from south Cape e Cerbère tiill the wetlland Albufe era de Valen ncia, with a total plan nar area of 4,200 4 nm2 (≈ ( 14,406 km2). Morpholog gical featurres of the continental c shelf divid de the stud dy area into o two subareas. The Northern subarea s (NSS), extendss from soutth Cape Ce erbère till Cape Salou canyons (p u and is ch haracterised d by a narrow shelf cutted c by submarine s planar 2 area ~1200 nm m ). The So outhern sub barea (SS), which exttends from m Cape Salo ou to 26.
Figure



Documento similar
These data clearly reveal a complex structure for Spanish CP both in root and embedded clauses: They involve sentences with more than one instance of a complementizer heading
The systematic uncertainties that are considered in this analysis can be divided into two main categories: the uncertainty in the determination of the SM background and the
For a short explanation of why the committee made these recommendations and how they might affect practice, see the rationale and impact section on identifying children and young
The expansionary monetary policy measures have had a negative impact on net interest margins both via the reduction in interest rates and –less powerfully- the flattening of the
Jointly estimate this entry game with several outcome equations (fees/rates, credit limits) for bank accounts, credit cards and lines of credit. Use simulation methods to
In our sample, 2890 deals were issued by less reputable underwriters (i.e. a weighted syndication underwriting reputation share below the share of the 7 th largest underwriter
This work shows differences in public participation in China and Western countries, conceptually and empirically. To do so, the different philosophical and public
The data obtained from the study area provide a broader view of the concentration, distribution, enrichment, sources, and the potential environmental contamina- tion by As in
