Efecto de diferentes fuentes de lípidos en dietas de vacas lecheras sobre la expresión de genes relacionados con la síntesis y secreción de grasa láctea de células somáticas en leche
73
0
0
Texto completo
(2) AGRADECIMIENTOS. Este estudio fue realizado en parte gracias a FONDECYT 1170400 (Fondo Nacional de Desarrollo Científico y Tecnológico, Chile) y la Vicerrectoría de Investigación of Pontificia Universidad Católica de Chile (Proyecto Puente P1608). Quisiera agradecer a la Fundación AgroUC y al laboratorio de alimentos del INTA de la Universidad de Chile por permitirnos utilizar sus dependencias para llevar a cabo el estudio realizado. Así como también a Hans A. Yoldi Harding (Comercial e Industrial Soho S.A.), quien nos donó el aceite de oliva utilizado en el presente estudio. A los docentes e investigadores Einar Vargas Bello, María Sol Morales y Jaime Romero, por compartir sus conocimientos y ser guías en este proceso, que en más de una ocasión se volvió más complejo de lo esperado. También agradecer a todos aquellos que nos ayudaron durante las semanas que duró el estudio en Pirque, a Nicolás Vera, Jonathan Becerra, Don Carlos, Don Luis, Don Juan y Roberto por la paciencia y ayudarnos a buscar a las vacas cuando se nos olvidaba cerrar alguna puerta y terminaban en los otros corrales. A quienes nos acompañaron en los muestreos, ayudando a que fueran días menos cansadores y con más risas Lucas Arze, Ricardo Gebauer, José María Godoy, Daniela Piña, y Stefanie Vyhmeister. Sobre todo, a Nathaly Cancino y Pietro Sciarresi por ayudarme cada vez que corría en círculos por no entender lo que hacíamos y en qué no estábamos metiendo. Gracias por dedica su tiempo para enseñarme y tener momentos de discusión que llevaron a que este trabajo finalmente pudiera ver la luz, gracias por ir juntos aprendiendo sobre la marcha. Y, por último, y más importante de todos, a mi familia que desde un principio me permitió continuar estudiando y apoyarme en este largo proceso. ¡Gracias!. 2.
(3) INDICE ABREVIACIONES ____________________________________________________ 5 ABSTRACT _________________________________________________________ 6 INTRODUCCIÓN _____________________________________________________ 7 REVISIÓN BIBLIOGRÁFICA ____________________________________________ 9 Definición de leche __________________________________________________ 9 Composición de la leche bovina ________________________________________ 9 Estructura de la glándula mamaria ______________________________________ 9 Síntesis y secreción de ácidos grasos lácteos en rumiantes __________________ 10 Producción y consumo de aceite de oliva ________________________________ 16 Efecto del aceite de oliva en la producción animal y composición láctea ________ 16 Producción de aceite de palma ________________________________________ 17 Efecto del aceite de palma en producción animal __________________________ 18 Estudios relacionados con la expresión de genes en la glándula mamaria _______ 18 Métodos para aislar las células somáticas de la glándula mamaria ____________ 20 HIPOTESIS ________________________________________________________ 22 OBJETIVO GENERAL ________________________________________________ 22 OBJETIVOS ESPECÍFICOS ___________________________________________ 22 MATERIALES Y MÉTODOS ___________________________________________ 23 1.. Animales y tratamientos ________________________________________ 23. 2.. Obtención de muestras _________________________________________ 24. 2.1.. Muestras de alimento __________________________________________ 24 2.1.1. Análisis químico __________________________________________ 24 2.1.2. Determinación del perfil de ácidos grasos de dietas y aceites ________ 25. 2.2.. Muestras de leche ____________________________________________ 26 2.2.1. Determinación del perfil de ácidos grasos de la leche ______________ 26 2.2.2. Aislamiento de células somáticas _____________________________ 27 3.
(4) 3.. Genes analizados ______________________________________________ 27. 3.1.. Preparación de ADN complementario (ADNc) _______________________ 29. 3.2.. Condiciones quantitative Polymerase Chain Reaction (qPCR) ___________ 29. 3.3 Evaluación de los amplicones ______________________________________ 30 3.3.1. Evaluación del tamaño de los amplicones _______________________ 30 3.3.2 Evaluación de secuencia de amplicones ________________________ 31 3.4 Evaluación expresión relativa mRNA ________________________________ 31 4.. Análisis estadístico _____________________________________________ 33. RESULTADOS Y DISCUSIÓN__________________________________________ 34 1.. Producción y composición láctea ___________________________________ 34. 2.. Perfil de ácidos grasos en la leche _________________________________ 36. 3.. Genes involucrados en el metabolismo de ácidos grasos ________________ 39. 3.1 Efectos del grado de insaturación de la suplementación lipídica en la expresión relativa de los genes analizados con relación al grupo Control ________________ 40 3.2 Evolución de la expresión relativa de los genes involucrados con el metabolismo lipídico en glándula mamaria con relación al tiempo ________________________ 47 4.. Limitantes y factores a considerar para la interpretación de los resultados ___ 50. CONCLUSIONES ___________________________________________________ 53 RESUMEN _________________________________________________________ 54 BIBLIOGRAFÍA _____________________________________________________ 55 ANEXOS __________________________________________________________ 66. 4.
(5) ABREVIACIONES ACACA: Acetyl-coenzyme A carboxylase Alpha ACSL1: Acyl-CoA synthetase long-chain family member 1 ACSS2: Acyl-CoA synthetase short-chain family member 2 PLIN2: Adipose differentiation related protein (adipophilin) AGPAT6: 1-acylglycerol-3 phosphate O- acyltransferase 6 BTN1A1: Butyrophilin, subfamily 1, member A 1 CD36: thrombospondin receptor DGAT1: Diacylglycerol acyltransferase 2 DGAT2: Diacylglycerol acyltransferase 2 FABP3: Fatty acid-binding protein 3 FABP4: Fatty acid-binding protein 4 FADS1: Fatty acid desaturase 1 (delta-5 desaturase) FASN: Fatty acid synthase GPAM: Glycerol-3-phosphate acyltransferase, mitochondrial LPIN1: Lipin 1 LPL: Lipoprotein lipase SCAP: SREBP cleavage activating protein SCD: Stearoyl-CoA desaturase (delta-9-desaturase) SLC27A: Soluble carrier protein 27 A very low-density lipoprotein receptor SREBF1: Sterol regulatory element-binding transcription factor 1 XDH: Xanthine dehydrogenase. 5.
(6) Efecto de diferentes fuentes de lípidos en dietas de vacas lecheras sobre la expresión de genes relacionados con la síntesis y secreción de grasa láctea de células somáticas.. ABSTRACT Various genes participate regulating the content of milk fat, from the uptake of fatty acids (AG) from the bloodstream to the secretion of the lipid droplet to the lumen of alveolus. The objective was to determine the expression of genes related to the metabolism and secretion of milk fat in the mammary gland in cows supplemented with AG of varying degrees of saturation. For 63 days 15 multiparous cows (189 ± 28 days in lactation), were divided into 3 groups according to body condition: Control without supplementation, olive oil (OO) and hydrogenated palm oil (HVO; both 30 g of oil / kg of MS). Every 21 days, the AG profile of the milk was analyzed by gas chromatography, and the relative expression of 17 candidate genes was studied using qPCR. There were no differences in food intake, weight or body condition. OO milk production was 9.7% higher and showed a decrease in medium chain AGs and an increase of 18 carbon AGs. Supplementing with OO produced an increase in the expression of PLIN2 and THRSP, while HVO generated a downward regulation of the latter during day 63 as well as a decrease in FABP3 and FABP4. In conclusion, supplementing with AG affects the activation of AG, intracellular transport, fat globule formation and a transcription factor (SREBF1). The latter is suppressed by the action of AGPI and is regulated upwards by the action of AGS, therefore, HVO would have a greater lipogenic effect than OO.. Key words: gene expression, milk fatty acids, ruminants, somatic cells.. 6.
(7) INTRODUCCIÓN La leche es un alimento de un alto valor nutricional, compuesto principalmente por agua, proteína, grasa, azúcares, minerales, entre otros. Dentro de estos, la grasa no sólo tiene importancia a nivel nutricional, sino que también otorga propiedades organolépticas, físicas y contribuye con características para el procesamiento de la leche y productos lácteos (Chilliard y Ferlay, 2004). A pesar de esto, la grasa láctea ha sido un punto a considerar por los profesionales de la salud debido a que se relacionó durante mucho tiempo con el aumento en el riesgo de aterosclerosis, enfermedades cardiovasculares e hipertensión arterial (Ulbricht y Southgate, 1991), es por esto que se han realizado variados estudios enfocados en mejorar el perfil de ácidos grasos, de manera de aumentar sus beneficios nutricionales. La composición de la grasa láctea puede variar según la genética animal, etapa de lactancia y la alimentación (Kliem y Shingfield, 2016). Siendo esta última importante debido a que la grasa tiene sus orígenes a partir de la alimentación, de la actividad del microbiota presente en el rumen y de reservas corporales (McGuire y Bauman, 2002), en el caso de ácidos grasos de cadena larga, y aquellos ácidos grasos de cadena corta y media se sintetizan en la glándula mamaria en un proceso conocido como síntesis de novo, en cuanto al ácido palmítico este tiene su origen en ambas vías metabólicas (Leroux et al., 2016). Diversos genes participan a lo largo de estas vías permitiendo la regulación de sus procesos, entre los que se encuentran aquellos asociados con la captación de aquellos ácidos grasos provenientes del torrente sanguíneo (LPL), transporte intracelular (ACSSL1, FABP3), síntesis y desaturación de ácidos grasos (ACACA, FSN, SCD), formación de la gota lipídica (PLIN2, BTN1A1) y secreción de ésta (XDH; Leroux et al., 2016; Bionaz y Loor, 2008a) Durante la última década se han realizado estudios de genómica funcional basados en qRT-PCR describiendo los efectos de las dietas suplementadas con ácidos grasos insaturados, principalmente oleico y linoleico, sobre diversos genes que se relacionan con el metabolismo de lípidos en la glándula mamaria (Harvatine y Bauman, 2006; Bionaz y Loor, 2008a, Kadegowda et al., 2009). Para poder realizar este tipo de estudios, existen diversos métodos para aislar las células somáticas, tales como biopsia, a partir de la cual se puede realizar microdisección con láser, o la obtención de células. 7.
(8) directamente de la leche mediante la ordeña, de la cual se puede aislar glóbulos de grasa, aislamiento de células mediante anticuerpos, y, en el caso de este estudio, aislamiento de células epiteliales. Este último tiene la ventaja de ser un método poco invasivo, que puede ser repetido a lo largo del tiempo y que es tan representativo como la biopsia del tejido mamario (Boutinaud, 2008). El conocimiento de los fenómenos que ocurren en la glándula mamaria podría contribuir al desarrollo de nuevas tecnologías en el manejo y crianza de los bovinos de leche. Si bien son diversos los estudios que evalúan la expresión génica en células epiteliales de la glándula mamaria, son escasos aquellos que utilizan como fuente de suplementación aceite de oliva, y estos se han enfocado principalmente en la suplementación en dietas de cabras y en los cambios que se producen a nivel de la leche o quesos elaborados (Chiofalo et al., 2004; Gómez-Cortes et al., 2008). Debido a lo anterior es que este trabajo se tiene por objetivo determinar la expresión de los genes relacionados con el metabolismo y secreción de la grasa láctea en la glándula mamaria de vacas alimentadas con fuentes de ácidos grasos de distintos grados de saturación, por lo cual se evaluó el efecto de adicionar en la dieta un 3% de aceite de oliva y aceite hidrogenada de palma en la dieta.. 8.
(9) REVISIÓN BIBLIOGRÁFICA Definición de leche La leche es la secreción mamaria normal de animales lecheros obtenida mediante uno o más ordeños sin ningún tipo de adición o extracción, destinada al consumo en forma de leche líquida o a elaboración ulterior (CODEX STAN 206-1999). Desde un punto de vista fisiológico, la leche es una emulsión de grasa y agua donde se encuentran disueltos carbohidratos, proteínas, minerales y vitaminas, los cuales son producidos o transportados a través de la glándula mamaria (Strucken et al., 2015). Composición de la leche bovina La leche bovina está compuesta aproximadamente por 87% agua, 4,6% lactosa, 3,4% proteínas, 4,2% grasa, 0,8% minerales y 0,1% vitaminas. Estos valores pueden ir modificándose según diversos factores tales como la raza, alimentación, manejos productivos, etapa de lactancia, estación del año (Mansson, 2008) y ciclo estral (McDonald et al., 1979; Toledo-Alvarado et al., 2018). Estructura de la glándula mamaria La glándula mamaria es un órgano exocrino, una glándula sudorípara o cutánea modificada, característico de la clase Mammalia, utilizado para alimentar a sus crías con el producto de su secreción. Es un órgano complejo formado por múltiples tipos de células incluyendo células mioepiteliales, estroma y células del sistema inmune (Cánovas et al., 2014). El tejido secretor de la ubre son las estructuras denominadas alveolos, los cuales se encuentran rodeados por células mioepiteliales que intervienen durante el proceso de la eyección de la leche. A su vez, los alveolos son irrigados por arteriolas que son ramificaciones de la arteria mamaria, la cual deriva de una rama de la arteria pudenda externa. En cuanto al sistema nervioso, la glándula mamaria es inervada por nervios que derivan de los nervios inguinales y del plexo mesentérico posterior del sistema simpático. Las glándulas mamarias carecen de fibras parasimpáticas (Urroz, 1991). Durante la lactancia la producción de leche es regulada principalmente por la lactosa que se encuentra en el alveolo, debido a que influye en la presión osmótica entre la 9.
(10) sangre y el alveolo, y así la cantidad de agua extraída hacia el interior del alveolo (Zhao y Keating, 2007). Ciertas sustancias como los minerales y vitaminas pasan a través de la membrana celular directamente desde la sangre mediante proteínas de transporte, al interior del alveolo el aparato de Golgi tiene un rol importante en el transporte intracelular y secreción del calcio, ya que forma vesículas que lo transportan asociado a caseínas (Neville y Watters, 1983; Figura 1).. Figura 1: Proceso síntesis de leche a nivel alveolar (Fuente: adaptado parcialmente de Wattiaux, 1996). Síntesis y secreción de ácidos grasos lácteos en rumiantes La grasa láctea está compuesta por un 98% de triacilglicerol, 0,2% de AG no esterificados (AGNEs) y esteres de retinol, y un 0,02% de monoacilglicerol y diacilglicerol. La leche contiene más de 400 tipos de AG de los cuales, los más abundantes son aquellos cuya conformación es de 4 a 18 carbonos (Lock y Bauman, 2004; Shingfield et al., 2012). Estos AG pueden ser incorporados en la leche de dos formas, a través de la captación de AGNEs y de trialcilglicerol provenientes de la sangre arterial y de la síntesis de novo en la glándula mamaria, que representan entre un 45 y 60% del total de ácidos grasos de la leche (Shingfield et al., 2012; McGuire y Bauman, 2002). 10.
(11) Como resultado de la síntesis de novo se obtiene el total de los AG de 4 a 12 carbonos, el 95% de ácido mirístico (C14:0) y aproximadamente el 50% del ácido palmítico (C16:0), mientras que los AG que contienen más de 18 carbonos en sus cadenas son originados a partir de la absorción de AG desde intestino delgado y de las reservas de tejido adiposo (Shingfield et al., 2012). El proceso conocido como síntesis de novo, ocurre a partir de los AG acético y butírico, así como también de cuerpos cetónicos, principalmente acetoacetato y β-hidroxibutirato (Chilliard et al., 2001; Palmquist, 2006), los cuales se obtienen principalmente de la digestión ruminal de la fibra dietaria (Lock y Bauman, 2004). Se estima que el 60% del β-hidroxibutirato se utiliza en dicho proceso (Bionaz y Loor, 2008a). Estos componentes mediante quilomicrones y lipoproteínas de muy baja densidad (VLDL por su sigla en inglés) son transportados hacia la glándula mamaria vía sanguínea, donde son captados por las enzimas lipoproteína lipasa (LPL; Palmquist y Jenkins, 1980), y el receptor de lipoproteínas de muy baja densidad (VLDLR), esta última, actúa sobre aquellos quilomicrones y VLDL que contienen en su estructura la molécula Apo-B48 (Bionaz y Loor, 2008a). Un porcentaje menor de AG ingresa a la célula mediante difusión pasiva, ya que los principales 2 de ingreso son mediante traslocasas de ácidos grasos y mediados por proteínas como es el caso del receptor FAT/CD36 el cual es una glicoproteína que transporta ácidos grasos de cadena larga hacia el interior de diferentes tejidos, principalmente tejido adiposo (Bionaz y Loor, 2008a). Una vez al interior de la célula, para que los AG puedan utilizarse en la síntesis de triacilglicerol y en la síntesis de novo previamente deben ser activados, lo cual ocurre vía ACSL1 (Acil-CoA sintetasa cadena larga miembro de la familia 1), esto se refiere a que los AG de cadena larga son esterificados con un CoA, mientras que la activación de AG de cadena corta es mediante la acción de la Acil-CoA sintetasa cadena corta miembro de la familia 2, ACSS2, el primero tiene una mayor afinidad con acetato, mientras que el segundo tiene más afinidad con propionato (Bionaz y Loor, 2008a). Luego, gracias a la acción de la enzima acetil-CoA carboxilasa alfa, ACACA, se producen ácidos grasos de cadena corta y palmitato a partir de acetato, el cual es un punto crítico en la síntesis de novo, por su parte la enzima FASN (sintasa de ácidos grasos) utiliza acetil-CoA y butirilCoA para producir estos AG (Bionaz y Loor, 2008a). Las células de la glándula mamaria sólo tienen la capacidad de elongar las cadenas de AG a partir de la condensación de. 11.
(12) acetil-CoA hasta los 16 carbonos, debido a la ausencia de enzimas que permitan una mayor elongación de las cadenas (Chilliard et al., 2001). Por su parte, aquellos AG de cadena larga, como se mencionó anteriormente, se obtienen a partir de lípidos de origen alimentario, los cuales sufren dos procesos a nivel del rumen, la lipólisis, que es la liberación de los ácidos grasos de los ésteres presentes en los lípidos de los alimentos, y la biohidrogenación, lo que corresponde a la saturación de los dobles enlaces de los ácidos grasos (Martínez et al., 2010). Este último proceso ocurre por la acción reductora que tiene el rumen, gracias a la importante presencia de protozoos y bacterias como Butyrivibrio fibrisolvens (Kim et al., 2000). Hay que considerar que el rumen de una vaca lechera contiene 40 a 50 litros de fluido ruminal, el cual a su vez contiene 1010 a 1011 bacterias y 105 a 106 protozoos por mililitro de fluido (Lock y Bauman, 2004). Dicha presencia de microorganismos permite que un gran porcentaje de los lípidos que ingresan al rumen sean metabolizados, sobre el 85% de los triglicéridos, fosfolípidos y glicolípidos que están presentes en la dieta pasan por el proceso de lipólisis (también descrito como hidrólisis) y se describe que un 70-95% del ácido linoleico y 85-100% del ácido linolénico son hidrogenados (Beam et al., 2000). Los rumiantes son capaces de obtener ácido linoleico conjugado y ácido vaccénico a partir de la dieta, a pesar de que los granos y el forraje que son utilizados en la alimentación de estos animales no los contienen, debido a la acción de las bacterias ruminales, ambos ácidos grasos corresponden a intermediarios de la hidrogenación del ácido linoléico y linolénico, cuyo producto final es el ácido esteárico (Figura 2; Chin et al., 1994; Lock y Bauman, 2004). Tal como se aprecia en la figura 2, hay dos grupos de bacterias que participan en la biohidrogenación, por un lado, están las bacterias pertenecientes al “Grupo A” que hidrogenan los AGPI obteniendo AG 18:1 trans, cómo es el ácido vaccénico, y luego se encuentran las bacterias del “Grupo B”, las cuales metabolizan estos AG 18:1 trans obteniendo ácido esteárico (Palmquist et al., 2004). El ácido linoleico también puede ser sintetizado a partir del ácido vaccénico por acción de la enzima delta 9 desaturasa (SCD; Griinari et al., 2000).. 12.
(13) Figura 2: Proceso de biohidrogenación del ácido linoleico (Traducido de Bauman y Griinari 2003). Dichos AG de cadena larga, una vez que ingresan a la glándula mamaria, tienen un movimiento intracelular muy lento, por lo que se requieren de transportadores específicos como FABP (proteínas de unión de ácidos grasos), moléculas que tienen diversas funciones asociadas al metabolismo lipídico, ya que participa en el proceso de captación y exportación de AG, regula las concentraciones de sustratos y productos en el citosol, permite el movimiento de vesículas que contienen AG a través del citosol en la célula (Storch y Thumser, 2000), tienen gran afinidad con acil-CoA, en el caso de la glándula mamaria (Bionaz y Loor, 2008a) y también regulan la expresión de genes relacionados con el metabolismo lipídico, ya que actúa en la mantención pasiva de AG en la célula e interactúa de forma directa con receptores de hormonas nucleares (Storch y Thumser, 2000). La familia FABPs está compuesta por 9 miembros, de los cuales las que más se expresan son FABP3 y FABP4, las que, junto con participar como moléculas de transporte, se postula que podrían amortiguar el efecto negativo de los AG activados, previniendo así la inhibición de las enzimas ACACA y SCD (Bionaz y Loor, 2008a). Esta última es de gran importancia debido a que solo un pequeño porcentaje de los AG captados por la glándula mamaria han sido insaturados por acción de microorganismos a nivel ruminal, por lo que genera AGMI al agregar un doble enlace en la posición 9 de los AG de 14, 16 y 18 carbonos (Bionaz y Loor, 2008a), y una manera de estimar su 13.
(14) acción es mediante el índice de desaturación, el cual se determina al calcular la relación entre el AG insaturado cis-9 y la sumatoria entre AG insaturado cis-9 y el AG saturado específico (Morales et al., 2000). El proceso de secreción de la grasa láctea comienza en el retículo endoplásmico rugoso, donde se sintetiza triacilglicerol mediante la acción de enzimas transferasas. Aún no es claro cómo los recién sintetizados AG se van agregando a la gota lipídica, que hasta ese momento tiene un diámetro menor a 0,5 µm de diámetro y que a medida que avanza por el citoplasma se va recubriendo con proteínas y lípidos polares de la membrana reticular. En algunos casos estas gotas lipídicas se van fusionando con otras mientras se van transportando hacia el borde de la célula. Durante la expulsión de la gota lipídica en el borde apical de la célula se van adquiriendo proteínas de membrana y fosfolípidos. Entre las proteínas se encuentran mucinas, xantina deshidrogenasa (XDH), receptor trombospondina (CD36), butirofilina (BTN1A1) y adipofilina (PLIN2; (Bauman et al., 2006). BTN1A1 es una proteína integral presente en la membrana apical de la célula, la cual se une a XDH promoviendo que la gota lipídica se envuelva con la membrana plasmática (Figura 3; Bauman et al., 2006).. 14.
(15) Figura 3: Interrelación entre las vías que regulan la síntesis de grasa láctea en el tejido de la glándula mamaria (Fuente: Bionaz y Loor, 2008a).. 15.
(16) Producción y consumo de aceite de oliva Debido a que Chile tiene un clima mediterráneo gracias a la presencia de la cordillera de los Andes y al océano Pacífico, es que la industria de la producción de olivos ha ido en aumento en los últimos años, alcanzando un total de 25.000 hectáreas, concentrándose las principales plantaciones entre la región de Atacama y el Maule. Durante el año 2017 la producción nacional fue de 20.000 toneladas de aceite de oliva, lo que representa un 0,7% de la producción mundial (Chile Oliva, 2017). El año 2016 a nivel mundial se produjeron un total de 2,54 millones de toneladas de aceite de oliva, un 19,6% menor a la temporada anterior, esto debido a las altas temperaturas registradas en sectores secanos (ChileOliva, 2017). Los principales productores son países de la Comunidad Europea tales como España (42% de la producción mundial), Italia y Grecia, seguidos por Turquía, Marruecos, Siria y Túnez (ChileOliva, 2015). En cuanto al consumo de este tipo de aceite, a nivel mundial durante el período 20162017 se consumieron en total 2,73 millones de toneladas, siendo Italia y España los principales países consumidores, seguidos por EEUU, Turquía y Marruecos (International Olive Oil Council, 2019). En el caso de nuestro país, los datos reportados datan del año 2015, indicando que para ese momento se consumían 6.588 toneladas de aceite, de los cuales sólo un 10% correspondía a aceites importados, esto debido a que se reconoce las características nutritivas y la calidad del aceite nacional (ChileOliva, 2015). Efecto del aceite de oliva en la producción animal y composición láctea Si bien el aceite de oliva como tal en la alimentación del ganado no se ha masificado tanto, si lo ha sido en el caso de la pomaza de oliva, la cual ha siendo ampliamente utilizada debido a que reduce los costos de alimentación y así como también reducen los desechos que se obtienen de la industria del aceite de oliva, lo que corresponde al cuesco, piel y parte de la pulpa de la oliva, así como también aquel aceite residual que no es utilizada para la producción de aceite destinada a consumo humano (Castellani et al., 2017).. 16.
(17) Los principales ácidos grasos del aceite de oliva y de la pomaza de oliva, son los ácidos oleico, palmítico y linoleico, mientras que el ácido esteárico y el linolénico se encuentran en menores proporciones (Castellani et al., 2017). Diversos estudios se han realizado evaluando la suplementación de ácido oleico en la dieta de rumiantes, detectando que si bien no genera grandes cambios en la producción láctea en vacas, si se produce una reducción en el porcentaje de grasa y cambios en el perfil de AG de la leche reduciendo el porcentaje de AG de cadena corta (Selner y Schultz, 1980; Castellani et al., 2017), lo cual también se ha evidenciado en el caso de la leche ovina, en la cual al utilizar pomaza de oliva, junto con los cambios mencionados se generó un incremento en la relación AGPI/AGS, disminución de los índices aterogénico y trombogénico de la leche ovina y aumento el contenido de ácido oleico en la leche (Chiofalo et al., 2004; Vargas-Bello et al., 2013). A pesar de los beneficios que genera la suplementación con aceite de oliva, altas dosis de AGMI en la dieta, tales como el ácido oleico, no se recomienda en rumiantes debido a que inhibe los microorganismos del rumen lo cual puede afectar la producción y composición de la leche (Jenkins, 1993). Producción de aceite de palma Entre los años 1995 y 2015 la producción mundial de aceite de palma ha aumentado de 15,2 a 62,6 millones de toneladas, siendo los principales países productores Indonesia (53%) y Malasia (32%), también se ha visto un aumento en su producción en países de Centro América, Tailandia y África Occidental (European Palm Oil Alliance, 2018). Lo cual se debe en parte a la eficiencia de producción en comparación a otros tipos de aceites, ya que por hectárea cultivada el aceite de palma produce 3,8 toneladas, muy superior al girasol (0,7 ton/ha) o a la soya (0,5 ton/ha; European Palm Oil Alliance, 2018). Las toneladas producidas de aceite de palma representan el 38,7% del total de grasas y aceites que se generan a nivel mundial (World Oil, 2016 obtenido de European Palm Oil Alliance, 2019). Los principales países desde donde se importa este producto a nuestro país son Colombia, Perú y Ecuador, alcanzando un total de 14,4 toneladas importadas durante el año 2016 (FAOSTAT, 2019).. 17.
(18) Efecto del aceite de palma en producción animal Durante la década de los años 90 el aceite de palma fue una alternativa de suplementación muy utilizada debido a que incrementa la densidad energética de la dieta de los rumiantes sin generar efectos negativos en la fermentación ruminal, tal como ocurre con los AGPI (Jenkins, 1993). Este tipo de aceite tiene la característica de presentar sobre un 50% de ácido palmítico, 1,9% ácido esteárico, 12,6% C18:1, 15,3% C18:2 y 1,2% C18:3 (Manso et al.,2009). Según lo indicado por Kupczyński et al. (2012) suplementar a las vacas con aceite de palma no genera cambios en la condición corporal, producción de leche ni en el contenido de sólidos totales. En dicho estudio compararon el perfil de AG con vacas suplementadas con aceite de pescado, y se evidenció que aquellas vacas que recibieron el tratamiento de aceite de palma produjeron leche con un 71,2% de AGS, 25% AG insaturados, un menor contenido de isómeros del ALC (menos del 1%), así como también una disminución en aquellos AGPI ω-3 y ω-6. Estudios relacionados con la expresión de genes en la glándula mamaria Se han realizado variados estudios con relación a la expresión de genes en la glándula mamaria de bovinos, siendo uno de los más citados el realizado por Bionaz y Loor (2008a) el que ayuda a comprender la variación en la expresión relativa de genes a lo largo de la lactancia, desde 15 días previos al parto hasta el día 240 postparto, sin la influencia de la suplementación en la dieta. En ese estudio se puede apreciar que la mayoría de los genes analizados se activan principalmente durante los primeros 60 días de lactancia y que las relaciones que estos genes tienen entre ellos forman una compleja red que permite coordinar las distintas funciones relacionadas con la síntesis y secreción de ácidos grasos hacia la leche. Similares estudios se han realizado en otras especies de la familia Bovidae, el año 2017 en China (Lee et al., 2017), se realizó la caracterización de la expresión de genes en yaks (Bos grunniens), a diferencia de las vacas, los yaks producen un 10% menos de leche con un contenido de grasa mayor, 5-7g /100 g. El comportamiento de los genes es similar al de las vacas lecheras a lo largo de la lactancia, pero aquellos genes coordinados por SREBF1, es decir aquellos encargados de síntesis de novo (ACSS2, ACACA y FABP3) y síntesis de triglicéridos (GPAM, AGPAT6 y LPIN1) se activan en 18.
(19) mayores niveles que en vacas y aquellos coordinados por PPARG, se expresan en menor medida, siendo estos involucrados en la formación de la gota lipídica (PLIN2, XDH y BTN1A1). Ambos estudios, de Bionaz y Loor (2008a) y Lee et al. (2017), permiten comprender cómo es el metabolismo lipídico normal de la glándula mamaria a lo largo de la lactancia de los bovinos, y son una herramienta útil para poder utilizarlos como base al momento de realizar estudios donde se quiera evaluar el efecto de la suplementación de la dieta en el metabolismo enzimático relacionado con la producción y secreción de grasa láctea. Fougère y Bernard (2018) suplementaron la dieta de cabras y vacas con almidón de trigo + aceite de maíz, aceite hidrogenada de palma, y algas marinas, observando que, de los 21 genes estudiados, 14 presentaron diferencias en cuanto a la especie, siendo 5 más abundantes en vacas (FADS3, ACSL1, PPARA. LXRA y PPARG), y 9 fueron más abundantes en cabras (FASN, CD36, FABP3, LPL, GPAM, LPIN1, CSN2, MFGE8, e INSIG1). En cuanto a la dieta, el aceite de maíz y las algas marinas generaron una reducción en la producción y contenido de grasa láctea mientras que el aceite hidrogenado de palma generó un aumento en el contenido de grasa láctea. En términos de metabolismo de los ácidos grasos, se apreció que el primer tratamiento redujo la abundancia de los factores de transcripción PPARG, INSIG1 y SP1, y la suplementación con algas marinas también redujo la abundancia de INSIG1. Al realizar la interacción dieta x especie, se observó que sólo LPL presentó una disminución en la abundancia en cabras alimentadas con algas marinas, y que aumentó cuando eran alimentadas con aceite hidrogenada de palma, en el caso de las vacas no presentó diferencias estadísticas. Previamente se han realizado otros utilizando variadas fuentes de alimentos que generan cambios en la grasa láctea y del metabolismo lipídico a nivel de glándula mamaria, entre los que se encuentran los realizados con vegetales y aceites altos en AGPI, entre los que predominan el aceite de linaza, que presenta un alto contenido de ácido alfa-linolénico (Angulo et al., 2012; Bernard et al., 2005b, Elis et al., 2016), aceite o harinilla de cártamo o safflower en inglés, cuyo ácido graso predominante es el ácido linoléico (Murrieta et al., 2006), aceite de pescado, caracterizado por contener AG de cadena larga sobre 20 carbonos (Carreño et al., 2016., Bernard et al., 2017; Toral et al.,. 19.
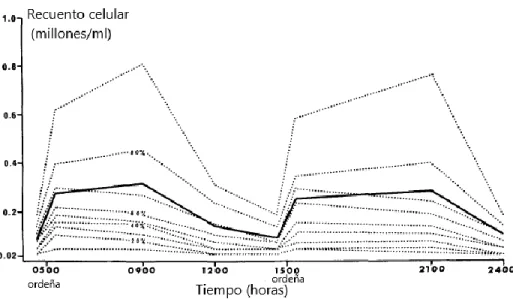
(20) 2016), y dietas que inducen depresión de grasa láctea (Harvatine y Bauman, 2006; Peterson et al., 2016; Piperova et al., 2000; Ticiani et al., 2016). Métodos para aislar las células somáticas de la glándula mamaria Se consideran células somáticas de la leche principalmente a leucocitos (glóbulos blancos) y algunas provenientes del tejido secretorio mamario (células epiteliales), estas últimas son eliminadas y renovadas de forma normal por la glándula mamaria (Cánovas et al., 2014; Boutinaud et al., 2015). A partir de estas células se puede obtener información para el estudio de la expresión de genes que regulan el metabolismo relacionado con la secreción de leche y sus diversos componentes, es por esto que se han desarrollado variados métodos para poder aislar las células somáticas de la glándula mamaria (Boutinaud, 2008). El método clásicamente utilizado para estudiar el tejido mamario es la biopsia, el cual requiere de un procedimiento quirúrgico para poder llevarlo a cabo, es decir, se realiza una incisión, la cual se debe ir rotando en cada muestreo, controlar la hemorragia y evitar posibles infecciones, por lo que no es un procedimiento sencillo y puede generar daños en el tejido (Boutinaud et al., 2015). A partir de tejido mamario congelado obtenido mediante biopsia se puede realizar microdisección con láser, cuya calidad de ARN puede verse afectado dependiendo del tiempo entre que se realiza la biopsia y la muestra es congelada, así como también dependiendo del tiempo que se tarda en cortar en secciones en el micrótomo (Cánovas et al, 2014), además de ser un procedimiento más simple y económico (Toral et al., 2016). Otros métodos se realizan al obtener leche mediante la ordeña, de la cual se pueden aislar células epiteliales, glóbulos de grasa y aislar células mediante anticuerpos. Realizar el aislamiento a partir de las células de la leche tiene la ventaja de ser menos invasivo, permitiendo que se puedan tomar muestras de forma repetitiva en el tiempo, con un menor riesgo que generar infección (Boutinaud, 2008). De acuerdo al estudio realizado por Smith y Shultze (1967) la secreción de células en la leche tiene un comportamiento cíclico durante el día (Figura 4), por lo que el mejor momento para poder realizar la toma de muestras en la leche es 4 horas posterior a la ordeña de la mañana debido a que se encuentra una mayor concentración de células, mientras que inmediatamente previo a la ordeña de la mañana el recuento celular 20.
(21) disminuye drásticamente y posterior a la ordeña el recuento celular entre los cuartos varía ampliamente. La integridad del ARN (RIN) varía considerablemente dependiendo del tipo de método utilizado. Al usar electroforesis tanto la microdisección como el aislamiento de células de la leche presentaron perfiles similares, mientras que los glóbulos de grasa tenían un peso molecular menor debido a la presencia de material citoplasmático, así como también presentó un mayor contenido de genoma bacteriano ya que la membrana del glóbulo tiene gran afinidad con las bacterias al contener fosfolípidos, proteínas y glicofosfolípidos (Cánovas, 2014).. Figura 4: Recuento de células somáticas en leche a lo largo del día (Fuente: traducido de Smith y Schultze, 1967).. 21.
(22) HIPOTESIS Suplementar con distintas fuentes de lípidos, con distintos grados de saturación de ácidos grasos afecta la expresión de genes relacionados con la síntesis y secreción de los ácidos grasos en la glándula mamaria de vacas lecheras, así como también en el perfil de AG de la leche. OBJETIVO GENERAL Determinar los cambios en la expresión de genes relacionados con la síntesis de lípidos presentes en la glándula mamaria al suplementar vacas en lactancia media con dos fuentes de lípidos. OBJETIVOS ESPECÍFICOS 1). Comparar la expresión de genes relacionados con el metabolismo y secreción de. ácidos grasos en la glándula mamaria de vacas lecheras al suplementar la dieta con aceite de oliva y aceite hidrogenada de palma. 2). Determinar el efecto de la suplementación de lípidos sobre los genes. relacionados con el metabolismo y secreción de ácidos grasos a lo largo del tiempo. 3). Relacionar el efecto de la suplementación con ácidos grasos en la dieta en el. perfil de AG de la leche con la expresión de genes relacionados con el metabolismo y secreción de ácidos grasos en la glándula mamaria en vacas lecheras durante la lactancia media.. 22.
(23) MATERIALES Y MÉTODOS 1. Animales y tratamientos Se trabajó con 15 vacas multíparas (hasta 3 partos), preñadas, con 189 ± 28 días en lactancia, una producción promedio de 33,4 ± 8,3 kg de leche al día, las cuales fueron divididas en 3 grupos según su condición corporal (CC), en una escala de 1 a 5, donde 1: emaciada y 5: obesa (Wildman et al., 1982), y en relación con su producción láctea. La CC de los grupos al inicio del estudio fueron en promedio 2,8 ± 0,3; 3,0 ± 0,0 y 2,8 ± 0,3. Este estudio se realizó durante 9 semanas en la estación experimental de Pirque de la Pontificia Universidad Católica de Chile (33°38′28″S, 70°34′27″O), entre los meses de septiembre y diciembre del año 2016. Los animales fueron estabulados en corrales individuales de 2,4 x 6 mt con acceso continuo a agua. En cuanto la alimentación, durante los primeros 7 días se alimentaron sólo con dieta basal, mientras estaban en el proceso de adecuación al sistema de estabulación y luego por 63 días se alimentaron con los respectivos tratamientos. La dieta basal contenía un 65% de forraje (ensilaje de maíz y de alfalfa) y un 35% de concentrado satisfaciendo las necesidades de las vacas de 650 kg en lactancia media consumiendo 26,5 kg de materia seca diaria (Tabla 1; NRC, 2001). Los tratamientos fueron grupo Control (N=5) al cual no se le adicionaron lípidos; grupo OO suplementado con aceite de oliva (N=5; 30 g/kg de materia seca) y un tercer grupo, HVO, el cual se suplementó con aceite hidrogenada de palma (N=5; 30 g/kg de materia seca). El alimento se entregaba una vez al día, posterior a la ordeña de la mañana, mediante el uso de carro forrajero. Los tratamientos eran administrados de forma separada y mezclados manualmente en la ración diaria que recibían las vacas (Anexos 1 y 2). Las vacas eran ordeñadas 3 veces al día (7:00, 15:00 y 21:00 horas) en una sala de ordeño tipo Tandem 2 x 6, con sistema de extracción automático de pezoneras, y equipada con un sistema de gestión de rebaños DelProTM (DeLaval, Suecia). Los procedimientos realizados fueron aprobados por el Comité de Ética de la Pontificia Universidad Católica de Chile.. 23.
(24) Tabla 1 Ingredientes de dietas Control, aceite de oliva (OO) y aceite hidrogenada de palma (HVO) Dieta Control OO HVO Composición (% Materia seca) Soiling de alfalfa 28,9 28,9 28,9 Ensilaje de maíz 27,0 27,0 27,0 Destilado de malta 23,1 23,1 23,1 Maíz grano 8,3 8,3 8,3 Salvado de trigo 6,2 6,2 6,2 Heno de alfalfa 2,6 2,6 2,6 Grano de soya 2,0 2,0 2,0 Harina de canola 1,5 1,5 1,5 a premix vitaminas y minerales 0,4 0,4 0,4 Aceite de oliva 0 3,0 0 Aceite hidrogenado de palma 0 0 3,0 a Contenido por kg: 25,000 mg de P; 80,000 mg de Ca; 25,000 mg de Mg; 1,612 mg de S; 300,000 UI de vitamina A; 50,000 UI de vitamina D3 y 1,600 UI de vitamina E, nd: no detectado.. 2. Obtención de muestras Los muestreos se realizaron cada 21 días (21, 42 y 63 d). En los cuales se realizó la medición de condición corporal y estimación de peso mediante perímetro torácico, obtención de muestras de alimento y leche (anexo 3), de la cual se analizó el perfil de ácidos grasos y se aislaron células somáticas de la leche. La producción diaria de leche era registrada diariamente de manera electrónica, mientras que el recuento de células somáticas (RCS) se obtuvo del control lechero oficial, realizado por la empresa Cooprinsem. 2.1.. Muestras de alimento. 2.1.1. Análisis químico Se obtuvieron muestras de alimento directo del comedero de las vacas y almacenado a -20°C hasta su posterior análisis, realizado en el laboratorio de Nutrición en la Facultad de Agronomía de la Pontificia Universidad Católica de Chile, donde se midieron los distintos componentes mediante procedimientos estándar (AOAC, 2001). La materia seca se obtuvo utilizando 5 g de muestra en una estufa a 105°C por 30 minutos, para la obtención de cenizas se utilizan 2 g de muestra y se colocan en la mufla 24.
(25) a 600°C por 2 horas (AOAC, método oficial 942.05). La proteína cruda se midió mediante el método de Kjeldahl, en el cual se requiere 1 g de muestra a la cual se le realiza digestión en bloque con adición de ácido sulfúrico (H 2SO4), un catalizador de cobre y destilación con ácido bórico (método oficial 984.13). La fibra cruda se obtuvo mediante digestión utilizando una solución con 1,25% de ácido sulfúrico y 1,25% de hidróxido de sodio (método oficial 950.02). El procedimiento de obtención de extracto etéreo requiere de 3 g de muestra de alimento, la cual se coloca inicialmente en la estufa a 105°C por 30 minutos, y luego se coloca en el extractor de solvente Velp Scientifica Ser148, el cual utiliza 60 ml de éter de petróleo (AOAC, 1990; método oficial 920.39). En cuanto a la fibra detergente neutro y fibra ácido detergente, se utiliza el equipo Ankom. Por último, la lignina se obtiene utilizando ácido sulfúrico al 72%, luego de haber medido la fibra ácido detergente. 2.1.2. Determinación del perfil de ácidos grasos de dietas y aceites En el caso de las dietas y los aceites utilizados para cada tratamiento, se utilizó 1 g de materia seca, se realizó la extracción de ácidos grasos mediante el protocolo de Blight y Dyer (1959), en el cual se utilizó una mezcla de cloroformo y metanol en una proporción 2:1 y posteriormente fueron metilados siguiendo el protocolo de Christie (1982). Los ácidos grasos metilados fueron analizados utilizando un cromatógrafo de gases líquidos Shimadzu modelo GC-2010, el cual está equipado con una columna Rtx 100 m x 0,32 mm x 0,2 µm. Las condiciones del equipo, para realizar el análisis fueron: una temperatura inicial de 110°C por 4 min, que luego incrementó a 160°C, con un aumento de 5°C/min, esta temperatura se mantuvo por 10 min, a continuación, aumentó a 225°C a 3°C/min manteniéndose por 10 min y finalmente se incrementó a 240°C a 3°C/min. Las temperaturas del detector de entrada y de ionización fueron de 260°C, la relación de división fue de 15:1 y para la inyección se usó 2µl. El flujo de gas transportador hidrógeno, para el detector, fue de 40 mL/min. El flujo de aire fue de 400 mL/min, y el flujo de gas auxiliar nitrógeno fue de 40 mL/min. Los peaks de los ácidos grasos en el cromatograma se identificaron mediante el uso de un estándar de ésteres metílicos de ácidos grasos (FAME; Supelco 37 Component FAME mix, Bellefonte, PA, USA) y los estándares de referencia para C18:1t11 (ácido vaccénico) y C18:1c9t11 (ácido linoleico conjugado; CLA) (Nu-Chek-Prep Inc., Elysian, MN, USA).. 25.
(26) 2.2.. Muestras de leche. Las muestras de leche se obtuvieron 3-4 horas después de la ordeña de la mañana, de manera manual, con el objetivo de maximizar el recuento de células somáticas de la leche (Smith y Schultze, 1967; Boutinaud y Jammes, 2002). Al momento de la toma de muestra se inspeccionaba visual y físicamente cada cuarto, con el objetivo de detectar aumento de tamaño, cambio de coloración o temperatura. Por su parte, la leche era evaluada mediante el uso de fondo oscuro para comprobar ausencia de mastitis clínica, y además se realizó california mastitis test (CMT) para descartar aquellos cuartos que presentaban mastitis subclínica. Se limpiaron los pezones con agua y jabón de clorhexidina, luego se aplicó pre dipping (solución de povidona yodada) y finalmente RNaseZap (Ambion, Austin, TX, USA), en la punta de los pezones con gasas estériles. Cabe señalar que después de la aplicación de cada solución los pezones fueron secados con toallas de papel. Posterior a la preparación, se recolectaron 400 ml de muestra de leche (100 ml de cada cuarto sano) en frascos de cultivo celular y luego se almacenó a 4°C hasta su posterior procesamiento, inmediatamente después de la obtención de la muestra. 2.2.1. Determinación del perfil de ácidos grasos de la leche Para la obtención del perfil de ácidos grasos de la leche, se utilizó la capa cremosa de la leche que era removida al momento de aislar el pellet de células somáticas. Para este proceso se utilizaron 50 µl de EDTA y se centrifugó a 4°C a 2000 RPM por 15 min, se separó mediante el uso de asas estériles y libres de RNAasa, y se almacenó en tubos eppendorf de 1,5 ml a -10°C hasta su posterior procesamiento, el cual se llevó a cabo en el laboratorio de Ciencias Animales en la Facultad de Agronomía de la Pontificia Universidad Católica de Chile. Para la separación de los ácidos grasos, cada muestra se centrifugó por 30 min a 12000 rpm, sin adicionar solventes, siguiendo el método de Feng et al. (2004), se obtuvo el sobrenadante, y posteriormente se realizó el proceso de transesterificación de acuerdo al protocolo de Christie (1982).. 26.
(27) 2.2.2. Aislamiento de células somáticas El procedimiento utilizado en la extracción de células somáticas fue obtenido de Wickramasinghe et al. (2012) y de Suárez-Vega et al. (2015), considerando sus modificaciones. Una vez que se removió la capa cremosa y el sobrenadante, se lavó el pellet con 10 ml de PBS (pH 7,4) y 0,5 µl de EDTA, se centrifugó a 2000 rpm por 15 minutos y se descartó el sobrenadante. Este procedimiento se realizó las veces necesarias hasta obtener un sobrenadante transparente y libre de materia grasa, el cual generalmente era necesario realizarlo en 3 oportunidades. Luego el pellet de células se resuspendió en 750 µl de Trizol (Invitrogen, Carlsbad, CA, USA), para estabilizar la molécula de ARN y se transfirió a tubos eppendorf de 1,5 ml y almacenándolo en un termo conservador de nitrógeno líquido a -80°C. Las muestras posteriormente se trasladaron al laboratorio de Biotecnología de Alimentos del INTA, donde se realizó la extracción de ARNm. 3. Genes analizados Para este estudio se seleccionaron 3 genes de control interno o de referencia, UXT, EIF3K y GADPH, los cuales fueron utilizados para normalizar los resultados obtenidos en las muestras a partir de los genes de interés; 13 genes candidatos y 4 genes reguladores de la transcripción. Los genes de referencia fueron seleccionados a partir de las publicaciones realizadas por Kadewogda et al. (2009) y Bonnet et al. (2013). Estos genes se relacionan con la síntesis de ácidos grasos y están agrupados según las siguientes funciones: síntesis y desaturación de ácidos grasos (AG); síntesis de triglicéridos; ingreso de AG a la célula; formación de glóbulo de grasa y, activación y transporte intracelular de acetato y AG. Las secuencias forward y reverse de los partidores, indicados en la tabla 2, se obtuvieron desde la base de datos GenBank, las cuales se ingresaron al programa BioEdit para poder determinar si se encontraban dentro de la secuencia del gen seleccionado. Los partidores fueron sintetizados, a partir de estas secuencias, por la compañía IDT (Integrated DNA Technologies; Coralville, IA, USA).. 27.
(28) Tabla 2: Genes involucrados con la síntesis y secreción de AG en la leche bovina Gen. Acceso GenBank. Secuencia Bp Ingreso de ácidos grasos a la célula F. 327 ACACAGCTGAGGACACTTGCC LPL BC119091 101 R. 427 GCCATGGATCACCACAAAGG F. 98 GCCCAGAACAGTGCCATATGA VLDLR AJ609502 103 R. 200 TTTTCACCATCACACCGCC Síntesis y desaturación de ácidos grasos F. 3709 CATCTTGTCCGAAACGTCGAT ACACA AJ132890 101 R. 3809 CCCTTCGAACATACACCTCCA F.6473 ACCTCGTGAAGGCTGTGACTCA FASN NM_001012669 92 R.6564 TGAGTCGAGGCCAAGGTCTGAA F. 809 TCCTGTTGTTGTGCTTCATCC SCD AY241933 101 R. 909 GGCATAACGGAATAAGGTGGC Activación y transporte intracelular de acetato y ácidos grasos F. 1929 GTGGGCTCCTTTGAAGAACTGT ACSL1 BC119914 120 R. 2048 ATAGATGCCTTTGACCTGTTCAAAT F. 1871 GGCGAATGCCTCTACTGCTT ACSS2 BC134532 100 R. 1970 GGCCAATCTTTTCTCTAATCTGCTT F. 458 GAACTCGACTCCCAGCTTGAA FABP3 NM_174313 102 R. 559 AAGCCTACCACAATCATCGAA F: 401 TGGTGCTGGAATGTGTCATGA FABP4 NM_174314 101 R: 501 TGGAGTTCGATGCAAACGTC Síntesis de triglicéridos F. 190 CCACTGGGACCTGAGGTGTC DGAT1 NM_174693 101 R. 290 GCATCACCACACACCAATTCA F: 389 CATGTACACATTCTGCACCGATT DGAT2 BT030532.1 100 R: 488 TGACCTCCTGCCACCTTTCT F. 147 TGGCCACCAGAATAAAGCATG LPIN1 NM_001206156 101 R. 247 GCTGACGCTGGACAACAGG Formación de glóbulos de grasa F. 161 TGGTCTCCTCGGCTTACATCA PLIN2 BC102211 81 R. 241 TCATGCCCTTCTCTGCCATC Regulación de la transcripción F. 1188 CCATGTGCACTTCAAGGAGGA INSIG1 NM_001077909.1 108 R. 1295 ATGTCGATCTTGCGTGTGGAG F. 1188 CCATGTGCACTTCAAGGAGGA SCAP. NM_00101889. SREBF1. NM_001113302. THRSP. AY656814. R. 1295 ATGTCGATCTTGCGTGTGGAG F. 2824 TGTCCACAAAAGCAAATCGC R. 2990 TGTCGACCACCTCTGGCTTC F. 631 CTACCTTCCTCTGAGCACCAGTTC R. 781 ACACACTGACCAGGTGACAGACA. 108 101 151. Ref. Bionaz y Loor, 2008a Bionaz y Loor, 2008a Bionaz y Loor, 2008a Bionaz y Loor, 2008a Bionaz y Loor, 2008a Bionaz y Loor, 2008b Bionaz y Loor, 2008a Bionaz y Loor, 2008b Bionaz y Loor, 2008b Bionaz y Loor, 2008a Bionaz y Loor, 2008a Bionaz y Loor, 2008b Bionaz y Loor, 2008a Harvatine y Bauman, 2006 Harvatine y Bauman, 2006 Bionaz y Loor, 2008a Harvatine y Bauman, 2006. F: Forward; R: Reverse; Bp: Pares de bases.. 28.
(29) 3.1.. Preparación de ADN complementario (ADNc). El protocolo de preparación del ADNc está conformado por 3 pasos; degradación del ADN contaminante, alineamiento de cebadores (o primers) y Transcripción Reversa. Antes de llevar a cabo estos procedimientos fue necesario determinar la concentración de ARN de las muestras de leche, con el objetivo de estandarizar la concentración dentro de un rango de 50 y 150 ng/µl (estipulado por el fabricante). Además, para este protocolo se utilizó una muestra control (sin ARN), para asegurar la ausencia de ADN durante la síntesis de ADNc. El primer paso es la degradación del ADN contaminante, el cual se realizó a través de la incubación con enzima ADNsa y así evitar alteración de los resultados. Para ello se utilizaron 3 µl de agua libre de nucleasas, Buffer de reacción 10X (RQ1 DNase) y ADNsa (RQ1 libre de RNasa). Luego se agregaron 7 µl de templado o ARN (muestra), y 7 µl de agua libre de nucleasas en el caso del control (-) de la reacción. Posteriormente las muestras se incubaron a 37°C por 30 minutos en termociclador para favorecer la acción de la enzima ADNasa. Una vez finalizada la reacción, se adicionó 1 µl de Stop solution, con el propósito de cesar la función de la enzima. El alineamiento de los cebadores busca sintetizar el ADNc a partir de la molécula de ARN. En dicha reacción se utilizaron 6 µl de la muestra tratada con ADNsa y 1 µl de cebador al azar. Para lograr el alineamiento de los cebadores y la síntesis de ADN, las muestras fueron incubadas a 70°C por 5 minutos. Durante el paso de Transcripción Reversa se elaboró una solución compuesta por: 4 µl de agua libre nucleasa; 4 µl de buffer 5X Improm-II; 4,5 µl de MgCl2; 1 µl oligonucleótidos; 0,5 µl de inhibidor de ARNasas (Rnasin) y 1 µl de enzima Transcriptasa reversa (Improm-II). Las muestras se incubaron a 25°C por 5 minutos (Alineamiento), luego a 42°C por 6 minutos (Extensión) y finalmente a 70°C por 15 minutos para inactivar las enzimas. 3.2.. Condiciones quantitative Polymerase Chain Reaction (qPCR). Cada reacción fue llevada a cabo en triplicado y con una solución de reacción compuesta por 200 µl de LightCycler® 480 SYBR Green I Master, concentración 2x (Roche, Basilea, Suiza), 140 µl de agua Roche y 10 µl de cada uno de los partidores. Las amplificaciones fueron realizadas en el equipo AriaMx Real-Time PCR (Modelo G8830A, Agilent 29.
(30) Technologies) y bajo las siguientes condiciones; un primer ciclo (Hot start 1) a 50°C por 2 minutos, seguido de un segundo ciclo (Hot start 2) a 95°C por 10 min. La amplificación constó de 40 ciclos, con denaturación a 95°C por 15 segundos, seguido por alineamiento a 60°C por 1 minuto y finalmente un ciclo (melt) a 95°C por 30 segundos, 65°C por 30 segundos y 95°C por 30 segundos. Se utilizaron dos temperaturas de alineamiento, 60ºC y 62°C (Tabla 3). Tabla 3: Condiciones qPCR para los genes candidatos. Gen ACACA, FADS2, FASN, SCD, LPL, VLDLR, ACSS2, DGAT1, DGAT2, PLIN2, SREBF1 FATP. Primer Programa 10 uM 2 min x 50ºC 40* 15 s x 95ºC (den.) 10 min X 1 min x 60ºC (ann.+ext.) 95ºC. Referencia Bionaz y Loor, 2008b. 10 uM. 40* 15 s x 95ºC (den.) 1 min x 62ºC (ann.+ext.). Bionaz y Loor, 2008a. 40* 15 s x 95ºC (den.) 1 min x 60ºC (ann.+ext.). Loor et al., 2005. 40* 15 s x 95ºC (den.) 1 min x 60ºC (ann.+ext.). Bionaz y Loor, 2008a. 40* 15 s x 95ºC (den.) 1 min x 62ºC (ann.+ext.). Harvatine y Bauman, 2006 Genes de referencia. 2 min x 50ºC 10 min X 95ºC ACSL1 10 uM 2 min x 50ºC 10 min X 95ºC FABP3, 10 uM 2 min x 50ºC FABP4, 10 min X LPIN1 95ºC INSIG1, 10 uM 2 min x 50ºC SCAP, 10 min X THRSP 95ºC GAPDH, 10 uM 2 min x 50ºC UXT, EIF3K 10 min X 95ºC Ann: alineado; Ext: extensión; *: ciclos.. 40* 15 s x 95ºC (den.) 1 min x 60ºC (ann.+ext.). 3.3 Evaluación de los amplicones 3.3.1. Evaluación del tamaño de los amplicones Se evaluó el tamaño de los amplicones sintetizados mediante electroforesis en gel de acrilamida. Para la elaboración del gel se utilizaron 3,25 mL de agua bidestilada, 6,6 mL de acriba-bisacrila, 2,5 mL de TBE (Tris, Borato y EDTA) a una concentración 5X, 65 µL de persulfato de amonio al 25% y 12,5 µL de tetrametiletilendiamina (TEMED). Para 30.
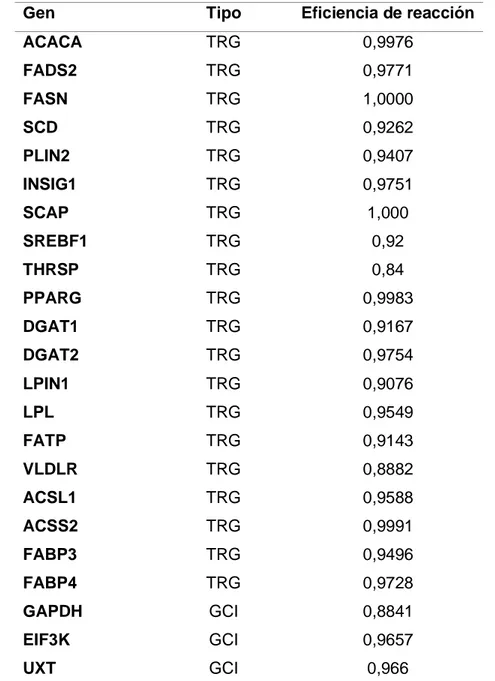
(31) realizar la reacción se utilizó TBE en una concentración de 1X como solución buffer y para la migración de los amplicones se aplicaron 200 volts por 25 min. En cuanto a la tinción y revelado de la reacción, primero se fijó el gel 50 mL de etanol-ácido acético a concentración 1X, se calentó por 10 seg y luego se sumergió en 50 mL de AgNO3 a una concentración 1X y se calentó nuevamente por 10 seg utilizando un horno microondas. Posteriormente se lavó el gel mediante el uso de agua bidestilada. Luego, se sumergió en una solución compuesta por 20 mL de NaOH al 7,5%, 30 mL de agua bidestilada y 0,5 mL de formaldehído al 37%, y se calentó en el horno microondas por 20 seg logrando revelar las bandas de migración. Por último, se lavó nuevamente el gel con agua bidestilada y se fijó usando etanol-ácido acético a una concentración 1X. Se aceptaron en este estudio aquellos amplicones cuyo tamaño obtenido mediante electroforesis fueran similares a los tamaños de los amplicones señalados en la literatura y que se indican como “Bp” en la tabla 2. 3.3.2 Evaluación de secuencia de amplicones Las secuencias de los amplicones sintetizados fueron evaluados por la empresa Macrogen USA (Cambridge, MA, USA), a través de la cual se realizaron cromatogramas de secuenciación los cuales posteriormente fueron analizados mediante el software BLAST (https://blast.ncbi.nlm.nih.gov/Blast.cgi). Este programa contiene una base de datos con la que se compararon los cromatogramas obtenidos y se seleccionaron aquellas secuencias que presentaban similitud con esta base de datos y cuyas secuenciaciones contenían peaks con alta resolución, baja presencia de ruidos y clara identificación de los nucleótidos que conforman las secuencias. 3.4 Evaluación expresión relativa mRNA Para poder determinar la expresión relativa de mRNA se utilizó Pair Wise Fixed Reallocation Randomizantion Test, elaborado por REST (2008), utilizando 2000 iteraciones, es decir, se realizan numerosas reasignaciones aleatorias entre las muestras seleccionadas e indica el número de veces en que la expresión relativa del grupo evaluado al azar el mayor que los datos de la muestra control (Pfaffl et al., 2002). El programa utiliza un modelo matemático que considera las eficiencias del PCR, en este estudio fueron entre 0,84 y 1 (tabla 4), y la desviación del punto de cruce medio (Ct). 31.
(32) entre la muestra y el control, considerando como procedimiento de normalización que la diferencia entre los Cts de la muestra y el control era 0.. Tabla 4: Eficiencia de reacción PCR de genes analizados (TRG) y genes de control interno (GCI) Gen. Tipo. Eficiencia de reacción. ACACA. TRG. 0,9976. FADS2. TRG. 0,9771. FASN. TRG. 1,0000. SCD. TRG. 0,9262. PLIN2. TRG. 0,9407. INSIG1. TRG. 0,9751. SCAP. TRG. 1,000. SREBF1. TRG. 0,92. THRSP. TRG. 0,84. PPARG. TRG. 0,9983. DGAT1. TRG. 0,9167. DGAT2. TRG. 0,9754. LPIN1. TRG. 0,9076. LPL. TRG. 0,9549. FATP. TRG. 0,9143. VLDLR. TRG. 0,8882. ACSL1. TRG. 0,9588. ACSS2. TRG. 0,9991. FABP3. TRG. 0,9496. FABP4. TRG. 0,9728. GAPDH. GCI. 0,8841. EIF3K. GCI. 0,9657. UXT. GCI. 0,966. En este estudio en primera instancia se evaluó la expresión relativa de los genes dentro de cada grupo al día 42 y 63 con relación al día 21, que permitió determinar aquellos genes que se mantenían estables a lo largo del tiempo o que presentaban variación. A. 32.
(33) partir de esto se seleccionaron aquellos que se mantuvieron estables y se calculó su expresión relativa en cada tratamiento en relación con el grupo Control, en cada uno de los periodos (día 21, 42 y 63). Posteriormente para determinar si los tratamientos OO y HVO generaban un efecto similar o distinto en los mismos grupos de genes que en el grupo Control a lo largo del tiempo es que se hizo el cálculo de la expresión relativa a los días 42 y 63 utilizando como control el día 21.. El cálculo utilizado se presenta a continuación, siendo E= eficiencia y su resultado se entiende como la razón de la expresión relativa de los genes indicando las veces que cambió debido al tratamiento ya sea en relación con el día 21 o en relación al grupo Control en cada período: 𝐸𝑥𝑝𝑟𝑒𝑠𝑖ó𝑛 𝑟𝑒𝑙𝑎𝑡𝑖𝑣𝑎:. (𝐸 𝐺𝑒𝑛 𝑏𝑙𝑎𝑛𝑐𝑜)∆Ct gen blanco (día control− día a evaluar) (𝐸 𝐺𝑒𝑛 𝑑𝑒 𝑟𝑒𝑓𝑒𝑟𝑒𝑛𝑐𝑖𝑎)∆𝐶𝑡 𝑔𝑒𝑛 𝑏𝑙𝑎𝑛𝑐𝑜 (𝑑í𝑎 𝑐𝑜𝑛𝑡𝑟𝑜𝑙− 𝑑í𝑎 𝑎 𝑒𝑣𝑎𝑙𝑢𝑎𝑟). 4. Análisis estadístico Se analizó la producción y composición de la leche, en cuanto a los litros producidos, grasa láctea, proteína, y recuento de células totales. El perfil de ácidos grasos de la leche se informa como g/100 g de ácidos grasos totales, a partir de dichos valores se realizó un análisis de varianza (ANOVA), considerando como factor de variación el tipo de suplementación (control, aceite de oliva y aceite hidrogenada de palma) y el tiempo. De existir diferencias entre estos valores se realizó una prueba de diferencia entre medias (Prueba de Tukey). Para determinar significancia estadística se utilizó una probabilidad de P ≤0,05. A partir del perfil de AG de la leche se calcularon los índices aterogénico (IA) y trombogénico (IT). Se utilizaron las fórmulas propuestas por Ulbricht y Southgate (1991), y se presentan a continuación: 𝐼𝐴:. 𝐼𝑇:. C12: 0 + 4 (C14: 0) + C16: 0 (n6 + n3)𝐴𝐺𝑃𝐼 + C18: 0 + ∑ 𝐴𝐺𝑀𝐼. C14: 0 + C16: 0 + C18: 0 (0,5 ∗ C18: 0) + 0.5 ∗ ∑ AGMI + 0.5 ∗ n6 AGPI + 3 ∗ n3 AGPI + ( n3 AGPI/ n6 AGPI). El análisis estadístico se realizó mediante el programa estadístico GenStat versión 12.1 (VSN International Ltd; 2009).. 33.
(34) RESULTADOS Y DISCUSIÓN 1. Producción y composición láctea Los datos productivos y perfil de ácidos grasos han sido reportados previamente (Vargas-Bello-Pérez et al., 2018). Las vacas tuvieron una ingesta promedio diaria de 26,5 Kg de MS sin diferenciarse entre grupos (tabla 5). Tampoco presentaron diferencias en el peso corporal ni en la condición corporal, ya que las dietas fueron formuladas de manera que fueran isocalóricas (ENL=1,6 Mcal Kg-1 de materia seca). El grupo suplementado con aceite de oliva (OO) produjo un 9,7% más de Kg de leche que el grupo Control y aquel suplementado con aceite de palma (HVO), alcanzando un promedio de 34,9 kg de leche diarios. No presentaron diferencias en producción de proteína, pero sí en el contenido de grasa ya que produjeron un 9,28% menos que los demás grupos. El grupo OO presentó una reducción de un 18,14% de AG generados a partir de síntesis de novo (C4:0 a C12:0), presentando 15,61 g/100 g de AG totales, comparados con los 19,07 g y 18,58 g de los grupos Control y HVO. A partir de lo anterior y considerando que no hay cambios en la producción de proteína, se podría indicar que las vacas del grupo OO presentaron síndrome de depresión de grasa láctea (SDGL; Baumgard et al., 2000). Entre los factores de riesgo de SDLG (Jenkins y Harvatine, 2014) se encuentran la adición de ionóforos, un contenido de FDN menor al 25% en la dieta y adición de AG insaturados en la dieta, los primeros dos factores no ocurrieron en este estudio ya que las dietas contenían entre 31 y 33% de FDN, sin embargo, en el caso de OO los AG insaturados superan los 60 g/100 g de AG totales en la dieta (Tabla 6). Estudios realizados con isómeros del ácido linoleico conjugado (ALC) observaron que ALC trans-10, cis-12 reduce la concentración de grasa en la leche de manera exponencial, llegando a una reducción de un 50% a una dosis de infusión de 7,5 g/d -1 (Baumgard et al., 2000; de Veth et al., 2004; Shingfield y Griinari, 2007), mientras que ALC cis-9, trans-11 no genera efecto alguno (Baumgard et al., 2000), y en este estudio, tanto en el grupo OO como HVO hubo un aumento en el contenido de C18:2 cis-9, trans11 en comparación al grupo Control, el cual sería producto de una biohidrogenación incompleta de los AGPI de 18 carbonos presentes en la dieta así como también de una producción endógena debido a la acción de Butirivibrio fibrisolvens, bacterias presentes. 34.
(35) en el rumen, y por el efecto de la enzima SCD, la cual se encuentra en tejido adiposo y la glándula mamaría (Kim et al., 2002), mientras que C18:2 trans-10, cis-12 no fue detectado en la leche y su producción se debería de manera exclusiva a la biohidrogenación ruminal (Griinari et al., 2000). Tabla 5: Desempeño y análisis proximal de la leche de vacas alimentadas con dietas control, aceite de oliva (OO) y aceite hidrogenado de palma (HVO) Dieta OO. Control HVO EE Valor P Producción Ingesta diaria de MS (kg MS/día) 26,5 26,5 26,5 * * b a b Producción de leche, kg/d 31,1 34,9 31,8 3,13 0,04 Grasa 1,02a 0,88b 1,04a 0,12 0,05 Proteína 1,05 0,97 1,08 0,25 0,58 Composición láctea, g/100g Grasa 3,28a 2,83b 3,28a 0,31 0,04 Proteína 3,39 3,16 3,41 0,36 0,29 Recuento células somáticas, × 10 3/ml 358a 145c 254b 82,0 0,02 Peso corporal, kg 662 636 700 79,0 0,23 Condición corporal 2,97 2,77 2,98 0,33 0,34 EE: Error estándar de la media; Valor P representa la probabilidad del efecto del tratamiento. Las medias en la misma fila con diferentes superíndices (a, b, c) son diferentes (P <0,05). (Vargas-Bello-Pérez et al., 2018) En relación al recuento de células somáticas (RCS) el grupo OO presentó un 59,5% menos en comparación a Control y 42,9% menos que HVO, promediando 145.000 células por ml de leche. Esto podría deberse a que la dieta OO, posee un alto contenido de AGPI ω-3 (19,1% de ácido α-linolénico), los cuales tienen efecto antiinflamatorio, debido a que los AGPI como ácido α-linolénico y linoleico una vez absorbidos se pueden convertir en AGPI de cadena muy larga, como. ácido araquidónico (AA),. eicosapentaenoico (EPA) y docosahexaenoico (DHA; Guillou et al., 2010), los que se incorporan en los fosfolípidos de membrana de monocitos, macrófagos y células del endotelio vascular, participando en la cadena de la inflamación (Arterburn et al., 2006; Calder, 2006). Por otro lado, de los AGPI ω-3 derivan mediadores antinflamatorios tales como resolvinas, protectinas y maresinas y también reducen la actividad proinflamatoria de los eicosanoides (Leucotrienos, tromboxanos y prostaglandinas) derivados de AA, producidos a partir de AGPI ω-6 (Serhan y Petasis, 2011).. 35.
(36) Tabla 6: Composición química de dietas Control, aceite de oliva (OO) y aceite hidrogenada de palma (HVO) Dietas OO. HVO. 38,4 14,4 4,6 33,5 19,8 4,2 6,2. 38,9 13,4 7,7 31,1 23,1 4,5 5,0. 38,4 14,3 7,1 33,4 19,4 4,5 6,0. 0,9 0,3 1,1 10,7 5,1 6,4 23,5 1,0 33,8 7,7 9,5. 0,1 0,1 0,2 0,2 1,9 10,1 26,3 32,8 9,0 nd 19,1. nd nd nd 0,1 1,2 58,0 39,8 nd nd 0,4 0,3. Control Composición química (% MS) Materia seca Proteína cruda Extracto etéreo Fibra neutro detergente Fibra ácido detergente Lignina Cenizas Composición de ácidos grasos (g/100g AG) C6:0 C10:0 C12:0 C14:0 C15:0 C16:0 C18 :0 C18:1 cis-9 C18:2 cis-9, cis-12 C18:3 cis-6, cis-9, cis-12 C18:3 cis-9, cis-12, cis-15 Nd: no detectado (Vargas-Bello-Pérez et al., 2018) 2. Perfil de ácidos grasos en la leche. En la tabla 7 se indica la composición de los AG de la leche de cada grupo tratado. En los anexos 5, 6, y 7 se presentan en detalle los perfiles de AG de la leche de los distintos grupos a través del tiempo. Los resultados fueron publicados en Vargas-Bello et al. (2018). No se evidenciaron diferencias en la sumatoria de los AGS, que fue cercana a 80 g /100 g de AG totales. Hubo diferencias significativas en la sumatoria de los AGMI y AGPI siendo el grupo OO donde se presentan en una mayor proporción, con valores de 16,2 y 6,43 g/100 g respectivamente. En el grupo OO se redujo el contenido de C8:0, C11:0 y C12:0, lo que podría explicarse debido a que la suplementación con AG de cadena larga (OO tiene una alta proporción de C18:1 cis-9), afecta negativamente la síntesis de novo en la glándula mamaria por lo cual dichos AG, producidos principalmente mediante esta vía a partir de acetato y 36.
Figure


+2

Outline
Perfil de ácidos grasos en la leche
Efectos del grado de insaturación de la suplementación lipídica en la expresión relativa de los genes analizados con relación al grupo Control
Limitantes y factores por considerar para la interpretación de los resultados Se utilizó en un comienzo el reactivo RNAlater (Sigma-Aldrich, Francia), el cual, de
Documento similar
Products Management Services (PMS) - Implementation of International Organization for Standardization (ISO) standards for the identification of medicinal products (IDMP) in
Products Management Services (PMS) - Implementation of International Organization for Standardization (ISO) standards for the identification of medicinal products (IDMP) in
This section provides guidance with examples on encoding medicinal product packaging information, together with the relationship between Pack Size, Package Item (container)
que hasta que llegue el tiempo en que su regia planta ; | pise el hispano suelo... que hasta que el
6 Para la pervivencia de la tradición clásica y la mitología en la poesía machadiana, véase: Lasso de la Vega, José, “El mito clásico en la literatura española
d) que haya «identidad de órgano» (con identidad de Sala y Sección); e) que haya alteridad, es decir, que las sentencias aportadas sean de persona distinta a la recurrente, e) que
De hecho, este sometimiento periódico al voto, esta decisión periódica de los electores sobre la gestión ha sido uno de los componentes teóricos más interesantes de la
Ciaurriz quien, durante su primer arlo de estancia en Loyola 40 , catalogó sus fondos siguiendo la división previa a la que nos hemos referido; y si esta labor fue de
