Modelos de Regulación Genética en Procariontes Basados en Redes de Petri Edición Única
148
0
0
Texto completo
(2) Modelos de Regulación Genética en Procariontes Basados en Redes de Petri Presentada por:. José Carlos Rebón Gutiérrez Aprobada por:. Dr. Edgar Dantán González Profesor Investigador del Centro de Investigación en Biotecnología, UAEM Asesor de Tesis. Dr. Jorge Luis Folch Mallol Profesor Investigador del Centro de Investigación en Biotecnología, UAEM Sinodal. Dr. Juan Frausto Solís Profesor Investigador del Departamento de Computación, ITESM Campus Cuernavaca Sinodal. Dr. Fernando Ramos Quintana Director de Investigación y del Programa de Graduados en Informática y Computación, ITESM Campus Cuernavaca Asesor de Tesis ii.
(3) A mis padres y a mis hermanos. A Carmen.. iii.
(4) AGRADECIMIENTOS Hay tantas personas y razones para agradecer, pero antes está Dios. A mis padres y a mis hermanos: no tengo palabras; simplemente, gracias por todo… A Carmen Cordero, por creer en mí. Gracias por siempre estar ahí, por el cariño, por la paciencia, y por muchas cosas más que no podría terminar de decir. A los de siempre: Sergio Méndez, Ileana Mondragón, Adrián Cortés, Miguel Ángel Peláez y Daniel Martínez... no es necesario mencionar por qué... A los de ahora: Lenin García, Abel Arredondo, Ricardo Domínguez, Carlos Figueroa, Luis Gerardo Díaz, Erika Collado, María Guadalupe Díaz, Sandra López y Moisés Quiroz... tampoco lo es... A Ismael Sánchez, pues sin su ayuda no habría logrado llegar hasta aquí. Al Dr. Fernando Ramos, por su invaluable guía, que permitió la culminación de este trabajo. Al Dr. Edgar Dantán, por su apreciable asesoría, no sólo en la revisión de la tesis. A los doctores Juan Frausto y Jordi Folch, por sus valiosas aportaciones como revisores y sinodales. Al Instituto Tecnológico y de Estudios Superiores de Monterrey, Campus Cuernavaca, por todas las facilidades económicas proporcionadas durante mis estudios. A Pedro Nájera, gran amigo, maestro y consejero. A María González, Rocío Cerecero, Jorge Álvarez, Juan Álvarez, Dalila Jiménez, Víctor Hugo Zárate y a todos mis profesores. Quiero finalmente agradecer y felicitar a mis compañeros del posgrado, por el apoyo durante las clases y proyectos, especialmente a Rogelio Herrerías, César Arronte y Quetzali Campos.. iv.
(5) RESUMEN El trabajo de tesis presentado trata sobre la aplicación de modelos formales computacionales basados en redes de Petri a mecanismos de regulación genética en sistemas biológicos. Los sistemas estudiados son procariontes: el operón lactosa de la bacteria Escherichia coli, y el bacteriófago lambda que afecta a la misma bacteria. La complejidad de estos sistemas se relaciona con el número de elementos y de interacciones que participan en sus mecanismos regulatorios. La complejidad de estos sistemas tiene un efecto inmediato en el entendimiento de su funcionamiento y de su comportamiento dinámico en tiempos determinados. Por funcionamiento se entiende conocer qué genes se expresan hasta qué grado, y en qué tiempos. El comportamiento dinámico implica la naturaleza cambiante del sistema debido a las múltiples interacciones entre proteínas y moléculas que logran la expresión genética. La complejidad también se refiere a los diversos componentes interrelacionados que tienen problemas de dimensionalidad y acoplamiento, lo que provoca dificultad en el entendimiento intuitivo del mecanismo de regulación. Simular sistemas de regulación genética mediante modelos formales es necesario para poder describir sin ambigüedad su estructura y su dinámica. El objetivo es analizar la expresividad de las redes de Petri para modelar mecanismos regulatorios, tratando sus problemas de comportamiento dinámico y complejidad. Se evaluará su desempeño a través de simulaciones que permitan observar su expresividad. Por expresividad, se busca que el modelo sea capaz de representar las características dinámicas de los sistemas, y en varios niveles de abstracción, y también se busca poder describir sistemas grandes y complejos, a partir de herramientas propias de los modelos. Para este trabajo de tesis se han seleccionado las redes de Petri por su capacidad para modelar las características dinámicas de los procesos, capacidad que se aprovechará para modelar el dinamismo de los sistemas de regulación genética. Algunos autores han desarrollado extensiones de las redes de Petri para simular los procesos discretos y continuos de algunos sistemas de regulación. La contribución principal en este trabajo se centra en el desarrollo de un modelo a varios niveles de abstracción que permite facilitar el entendimiento del funcionamiento y comportamiento de los sistemas biológicos estudiados. Las redes de Petri coloreadas resultaron suficientemente expresivas para el objetivo buscado. Al final, se obtiene un modelo discreto de cada uno de los sistemas biológicos estudiados, el cual representa adecuadamente su dinámica, proporcionando un análisis a varios niveles de abstracción, facilitando el entendimiento del funcionamiento y comportamiento de los mecanismos de regulación estudiados. v.
(6) CONTENIDO Resumen .....................................................................................................................................................................v Índice de Figuras ......................................................................................................................................................viii Índice de Cuadros y Tablas .......................................................................................................................................x 1. Introducción ...........................................................................................................................................................1 1.1 Antecedentes..................................................................................................................................................2 1.1.1 Células procariotas y células eucariotas ..............................................................................................2 1.1.2 Regulación genética...............................................................................................................................3 1.1.3 La bioinformática ....................................................................................................................................3 1.1.4 Modelos y simulaciones.........................................................................................................................4 1.2 Planteamiento del problema ..........................................................................................................................6 1.3 Objetivos .........................................................................................................................................................7 1.3.1 Objetivo general .....................................................................................................................................7 1.3.2 Objetivos particulares.............................................................................................................................7 1.4 Alcances y limitaciones ..................................................................................................................................8 1.5 Metodología de desarrollo .............................................................................................................................9 2. La Célula y Regulación de la Expresión Genética ............................................................................................10 2.1 Características de las células eucariotas....................................................................................................12 2.2 Características de las células procariotas ..................................................................................................14 2.3 Estructura genética ......................................................................................................................................16 2.3.1 Las proteínas........................................................................................................................................17 2.4 El dogma central de la biología molecular..................................................................................................18 2.4.1 Transcripción ........................................................................................................................................19 2.4.2 Traducción ............................................................................................................................................20 2.4.3 El código genético ................................................................................................................................21 2.4.4 Promotores, operadores, represores, operones, exones, intrones...................................................22 2.4.5 Polimerasas ..........................................................................................................................................23 2.5 Regulación de la expresión genética ..........................................................................................................25 2.5.1 Regulación en procariotas ...................................................................................................................27 3. Redes de Petri .....................................................................................................................................................31 3.1 Redes de Petri blanco y negro ....................................................................................................................31 3.2 Redes de Petri coloreadas...........................................................................................................................34 3.2.1 Sintaxis y semántica en el uso de las redes.......................................................................................35. vi.
(7) 3.3 Concepto del tiempo en las redes de Petri.................................................................................................44 4. Trabajos Relacionados Relevantes....................................................................................................................46 4.1 Redes de Petri en regulación genética .......................................................................................................47 5. Representaciones en Redes de Petri de Sistemas Regulatorios Bacterianos................................................49 5.1 Modelo del mecanismo de retroalimentación del fago lambda con redes de Petri..................................49 5.1.1 El bacteriófago lambda ........................................................................................................................49 5.1.2 Redes de Petri híbridas .......................................................................................................................54 5.1.3 Efecto de las concentraciones de las proteínas CI y Cro..................................................................57 5.1.4 Mecanismo de retroalimentación de CI y Cro ....................................................................................59 5.1.5 Representación de la etapa temprana de expresión genética del fago lambda en HPN ................60 5.2 Modelo del mecanismo regulatorio del operón lac con redes de Petri .....................................................64 5.2.1 El operón lac.........................................................................................................................................64 5.2.2 Redes de Petri híbridas funcionales ...................................................................................................67 5.2.3 El switch de control de la transcripción...............................................................................................70 5.2.4 Regulación positiva ..............................................................................................................................71 5.2.5 Regulación negativa.............................................................................................................................72 5.2.6 Hidrólisis de lactosa en glucosa y galactosa ......................................................................................74 6. Desarrollo del Modelo, Simulación y Análisis de Resultados...........................................................................77 6.1 El operón lac a varios niveles de abstracción ............................................................................................77 6.1.1 Simulación del modelo obtenido del operón lac con redes de Petri coloreadas..............................87 6.1.2 Análisis de resultados ..........................................................................................................................90 6.1.2.1 Nivel bajo de abstracción.............................................................................................................90 6.1.2.2 Nivel medio de abstracción .........................................................................................................95 6.1.2.3 Nivel alto de abstracción..............................................................................................................99 6.2 El bacteriófago lambda a varios niveles de abstracción..........................................................................103 6.2.1 Simulación del modelo obtenido del fago lambda con redes de Petri coloreadas.........................109 6.2.2 Análisis de resultados ........................................................................................................................110 6.2.2.1 Nivel bajo de abstracción...........................................................................................................111 6.2.2.2 Nivel medio de abstracción .......................................................................................................115 6.2.2.3 Nivel alto de abstracción............................................................................................................120 6.3 Resumen del análisis de resultados..........................................................................................................124 7. Conclusión .........................................................................................................................................................128 7.1 Trabajo a futuro ..........................................................................................................................................130 8. Referencias Bibliográficas ................................................................................................................................132. vii.
(8) ÍNDICE DE FIGURAS Figura 1.1: Abstracción de una situación que lleva a la construcción de un modelo.. 5. Figura 2.1: Esquema de una célula ecuariota.. 12. Figura 2.2: Esquema de una célula procariota.. 14. Figura 2.3: Estructura de la doble hélice del ADN.. 16. Figura 2.4: Dogma central de la biología molecular.. 19. Figura 2.5: Esquema de la transcripción.. 20. Figura 2.6: Esquema de la traducción.. 21. Figura 2.7: Código genético estándar.. 22. Figura 2.8: Niveles en que ocurre la regulación de la expresión genética.. 26. Figura 2.9: Operón triptófano, ejemplo de operón represible.. 28. Figura 2.10: Operón lac, ejemplo de operón inducible.. 29. Figura 3.1: Red de Petri blanco y negro sencilla.. 32. Figura 3.2: CPN que describe el sistema de asignación de recursos en su marca inicial M0.. 36. Figura 3.3: Transición T2 de la Fig. 3.2.. 39. Figura 3.4: Transición T1 de la Fig. 3.2.. 41. Figura 3.5: Ejemplo de una red de Petri Coloreada.. 42. Figura 5.1: Ilustración del fago lambda con medidas aproximadas.. 49. Figura 5.2: Mapa genético del fago λ.. 50. Figura 5.3: Ciclo de vida del fago λ.. 51. Figura 5.4: Operón de dos genes para ejemplificar una HPN.. 56. Figura 5.5: Representación en HPN del operador OR de la Tabla 5.1.. 58. Figura 5.6: Mecanismo de retroalimentación de CI y Cro.. 59. Figura 5.7: Etapa temprana de expresión genética del fago lambda en HPN.. 61. Figura 5.8: Efecto de la presencia de lactosa en la transcripción del operón lac.. 65. Figura 5.9: Efecto de la ausencia de lactosa en la transcripción del operón lac.. 66. Figura 5.10: Transcripción reprimida y unión del complejo AMPc-CAP con el operón lac.. 67. Figura 5.11: Regulación positiva por AMPc del operón lac.. 67. Figura 5.12: Notaciones gráficas de lugares y transiciones en HPNs, HFPNs, y HDNs.. 68. Figura 5.13: Representación en HFPN del switch de control de la transcripción del operón lac.. 70. Figura 5.14: Representación en HFPN de la regulación positiva en el operón lac.. 71. Figura 5.15: Representación en HFPN de la regulación negativa del operón lac.. 73. Figura 5.16: Representación en HFPN de los mecanismos regulatorios del operón lac.. 75. viii.
(9) Figura 6.1: Estructura y lazos regulatorios del operón lac.. 78. Figura 6.2: Diagrama de bloques del operón lac al más alto nivel de abstracción.. 79. Figura 6.3: Red de Petri al más alto nivel de abstracción del operón lactosa.. 80. Figura 6.4: Red de Petri desarrollada para modelar el switch de transcripción.. 81. Figura 6.5: Red de Petri desarrollada para modelar la regulación negativa.. 82. Figura 6.6: Red de Petri desarrollada para modelar la regulación positiva.. 83. Figura 6.7: Red de Petri desarrollada para modelar la transcripción de genes estructurales.. 83. Figura 6.8: Red de Petri desarrollada para modelar con los módulos integrados.. 84. Figura 6.9: HFPN para el operón lactosa sobrepuesta al esquema biológico.. 86. Figura 6.10: Red de Petri coloreada desarrollada para modelar el comportamiento de los mecanismos regulatorios del operón lac en CPN Tools en la marca inicial.. 87. Figura 6.11: Imagen de las declaraciones para la CPN de la Fig. 6.10.. 88. Figura 6.12: Despliegue en pantalla del Caso 1.. 90. Figura 6.13: Despliegue en pantalla del Caso 2.. 91. Figura 6.14: Despliegue en pantalla del Caso 3.. 92. Figura 6.15: Despliegue en pantalla del Caso 4.. 93. Figura 6.16: Resumen ilustrado de los cuatro posibles casos.. 94. Figura 6.17: Despliegue en pantalla del estado inicial a un nivel medio de abstracción.. 95. Figura 6.18: Despliegue en pantalla del Caso 1 a un nivel medio de abstracción.. 96. Figura 6.19: Despliegue en pantalla del Caso 2 a un nivel medio de abstracción.. 97. Figura 6.20: Despliegue en pantalla del Caso 3 a un nivel medio de abstracción.. 98. Figura 6.21: Despliegue en pantalla del Caso 4 a un nivel medio de abstracción.. 99. Figura 6.22: Despliegue en pantalla del estado inicial del modelo abstracto del operón lac.. 100. Figura 6.23: Estado final de la transcripción de los genes en el modelo abstracto.. 101. Figura 6.24: Estado final de los casos en que no hay transcripción de genes en el modelo abstracto.. 102. Figura 6.25: Imagen simplificada de los lazos regulatorios del fago λ.. 103. Figura 6.26: Diagrama de bloques del fago λ, al más alto nivel de abstracción.. 104. Figura 6.27: Red de Petri del fago λ al más alto nivel de abstracción.. 104. Figura 6.28: Red de Petri desarrollada para modelar la regulación de Cro y la lisis.. 105. Figura 6.29: Red de Petri desarrollada para modelar la regulación de CI y la lisogenia.. 106. Figura 6.30: Red de Petri desarrollada para modelar bacteriófago λ.. 107. Figura 6.31: HFPN para el bacteriófago lambda sobrepuesta al esquema biológico.. 108. Figura 6.32: Red de Petri desarrollada para modelar el comportamiento del fago lambda en la marca inicial.. 109. ix.
(10) Figura 6.33: Estado final de la red desarrollada para modelar el mecanismo regulatorio del fago lambda en el caso en que no hay anti-terminador ni producción de moléculas que inducen las vías.. 111. Figura 6.34: Estado de la red desarrollada que provoca el crecimiento lítico.. 112. Figura 6.35: Estado final de la red desarrollada en lisis.. 113. Figura 6.36: Estado de la red desarrollada que provoca el crecimiento lisogénico.. 114. Figura 6.37: Estado final de la red desarrollada en lisogenia.. 115. Figura 6.38: Despliegue de la red desarrollada a un nivel medio de abstracción en la marca inicial.. 116. Figura 6.39: Estado de la red que provoca el crecimiento lítico a un nivel medio de abstracción.. 117. Figura 6.40: Estado final de la red para la lisis a un nivel medio de abstracción.. 118. Figura 6.41: Estado de la red que provoca el crecimiento lisogénico a un nivel medio de abstracción.. 119. Figura 6.42: Estado final de la red para la lisogenia a un nivel medio de abstracción.. 120. Figura 6.43: Despliegue en pantalla de la red desarrollada a un alto nivel de abstracción.. 121. Figura 6.44: Estado final de la red para la lisis a un alto nivel de abstracción.. 122. Figura 6.45: Estado final de la red para la lisogenia a un alto nivel de abstracción.. 123. Figura 6.46: Diagrama en forma de pirámide que explica los niveles de abstracción.. 125. Figura 6.47: Diagrama que muestra la interacción entre elementos.. 126. ÍNDICE DE CUADROS Y TABLAS Cuadro 3.1: Descripción de declaraciones en CPN ML.. 37. Tabla 5.1: Relaciones entre concentraciones de CI, Cro, luz UV y los promotores PRM y PR y el operador OR.. 57. Tabla 6.1: Resultado de transcripción de los genes dependiendo del estado final de la red de Petri.. 90. x.
(11) Figura 6.33: Estado final de la red desarrollada para modelar el mecanismo regulatorio del fago lambda en el caso en que no hay anti-terminador ni producción de moléculas que inducen las vías.. 111. Figura 6.34: Estado de la red desarrollada que provoca el crecimiento lítico.. 112. Figura 6.35: Estado final de la red desarrollada en lisis.. 113. Figura 6.36: Estado de la red desarrollada que provoca el crecimiento lisogénico.. 114. Figura 6.37: Estado final de la red desarrollada en lisogenia.. 115. Figura 6.38: Despliegue de la red desarrollada a un nivel medio de abstracción en la marca inicial.. 116. Figura 6.39: Estado de la red que provoca el crecimiento lítico a un nivel medio de abstracción.. 117. Figura 6.40: Estado final de la red para la lisis a un nivel medio de abstracción.. 118. Figura 6.41: Estado de la red que provoca el crecimiento lisogénico a un nivel medio de abstracción.. 119. Figura 6.42: Estado final de la red para la lisogenia a un nivel medio de abstracción.. 120. Figura 6.43: Despliegue en pantalla de la red desarrollada a un alto nivel de abstracción.. 121. Figura 6.44: Estado final de la red para la lisis a un alto nivel de abstracción.. 122. Figura 6.45: Estado final de la red para la lisogenia a un alto nivel de abstracción.. 123. Figura 6.46: Diagrama en forma de pirámide que explica los niveles de abstracción.. 125. Figura 6.47: Diagrama que muestra la interacción entre elementos.. 126. ÍNDICE DE CUADROS Y TABLAS Cuadro 3.1: Descripción de declaraciones en CPN ML.. 37. Tabla 5.1: Relaciones entre concentraciones de CI, Cro, luz UV y los promotores PRM y PR y el operador OR.. 57. Tabla 6.1: Resultado de transcripción de los genes dependiendo del estado final de la red de Petri.. 90. x.
(12) 1. INTRODUCCIÓN Entender la actividad de los sistemas regulatorios genéticos es un reto, ya que es necesario comprender su funcionamiento y su comportamiento dinámico, en el cual intervienen múltiples interacciones. Se debe describir sin ambigüedad su estructura, para lo cual es necesaria la aplicación de modelos matemáticos y/o computacionales. La expresividad de estos modelos ayuda no sólo a la comprensión, sino también al estudio de mecanismos más complejos. Como ya se mencionará más adelante en el documento, modelar y simular sistemas de regulación genética mediante modelos formales es necesario porque se puede describir sin ambigüedad a los procesos involucrados (además, mediante técnicas gráficas, se podría lograr la visualización del comportamiento dinámico). Este trabajo de tesis trata acerca del uso de modelos formales matemáticos-computacionales expresivos para el análisis de sistemas de regulación genética.. Concretamente, se analizan modelos de redes de Petri aplicados al estudio de. mecanismos de regulación genética en sistemas biológicos, en particular, el operón lac y el fago λ. El trabajo se centra en la hipótesis de que una herramienta computacional, como las redes de Petri, cumple con el concepto de expresividad definido en los objetivos, para ayudar a la mejor comprensión de los sistemas de regulación genética. Conocida la complejidad de este tipo de sistemas biológicos, es necesario contar con herramientas que faciliten su estudio. Para poder llevar a cabo el análisis, el resto del trabajo se estructura como sigue. En este capítulo, se introducen brevemente algunos conceptos para dar contexto a este trabajo de tesis, se plantea el problema de investigación, se establecen los objetivos y los alcances del trabajo, y se diseña la metodología a desarrollar. En el Capítulo 2 se describen a detalle los fundamentos biológicos del trabajo, con énfasis en las células procariotas, el dogma central de la biología molecular, y los principios de la regulación de la expresión genética. En el Capítulo 3 se discuten los conceptos, propiedades y sintaxis de las redes de Petri y, específicamente, de las redes de Petri coloreadas. En el Capítulo 4 se mencionan, y se hace una breve descripción de los trabajos existentes en los cuales se aplican las redes de Petri para el estudio de mecanismos de regulación genética. El Capítulo 5 trata exclusivamente de los dos trabajos principales sobre los que se centra este estudio. En el Capítulo 6 se encuentra la descripción del modelo desarrollado, las simulaciones y el análisis de los resultados obtenidos. En el Capítulo 7 se concluye el trabajo, y se mencionan posibles trabajos a futuro sobre la misma línea. Al final están las referencias que sustentan la tesis. 1.
(13) 1.1 Antecedentes. 1.1.1 Células procariotas y células eucariotas. Los organismos tienen un conjunto en común de genes y proteínas porque comparten un origen en común. Se puede entender que la unidad fundamental de los procesos y otras características de los seres vivos se deban a la herencia desde un ancestro en común que ya poseía los mecanismos moleculares, desde un punto de vista evolutivo [HARTL 2001]. A través de la evolución, y de las similitudes encontradas en la secuencia de nucleótidos en moléculas de ARN de los ribosomas, se han distinguido tres grandes reinos de organismos: el Reino Bacteria, el Reino Archaea, y el Reino Eukarya. El Reino Bacteria comprende a la mayoría de las bacterias, y las cianobacterias (anteriormente denominadas algas verdes o azules). Las células de estos organismos carecen de un núcleo rodeado por una membrana y de mitocondria, están envueltas por una pared celular, y se dividen por fisión binaria. El Reino Archaea comprende microorganismos cuyas células carecen de membranas internas. Aunque sus mecanismos de replicación genética son parecidos a los de los organismos del Reino Eukarya, su metabolismo es muy parecido al de las bacterias. El Reino Eukarya incluye a todos los organismos cuyas células contienen una red elaborada de membranas internas, un núcleo rodeado por una membrana y mitocondrias (plantas, animales, hongos, y muchos organismos unicelulares como amibas y protozoarios). Su ADN está organizado en cromosomas verdaderos, y la división celular ocurre por el proceso de mitosis. Los miembros de los reinos Bacteria y Archaea se unen en el gran grupo de los procariontes. Se dice que sus células son procariotas ya que carecen de núcleo. Literalmente, procariota significa “antes (de la evolución) del núcleo”. Es evidente que los miembros del Reino Eukarya son eucariontes, o lo que es lo mismo, sus células son eucariotas, que significa “núcleo bien formado”. En esta investigación, se trabajará únicamente con organismos cuyas células sean procariotas, como es el caso de las bacterias. Concretamente, una labor profunda sobre algunos de los sistemas regulatorios de la bacteria Escherichia coli.. 2.
(14) 1.1.2 Regulación genética. La regulación genética (o regulación de la expresión genética) se refiere a los procesos que afectan y controlan la acción del gen a nivel de traducción o transcripción. En éstos se regulan los productos funcionales de un gen (ya sea una molécula de ARN o una proteína) [REGULATION 2007]. La regulación genética le da el control a la célula sobre su estructura y función, además de ser la base de la versatilidad y adaptabilidad de cualquier organismo. La expresión genética puede ser regulada en varias etapas, entre las que se encuentran: la modificación química y estructural del ADN, la transcripción, la traducción, el transporte de ARN, la degradación del ARNm, y modificaciones pos-transcripción y traducción. El control de la expresión genética es regulado de una forma altamente organizada para asegurar que genes específicos sean expresados en los tiempos y niveles apropiados en respuesta a varios estímulos genéticos y ambientales. En las células eucariotas, la expresión genética es controlada en múltiples niveles, desde el agrupamiento de la maquinaria elemental para la transcripción de promotores específicos, hasta el procesamiento y maduración del ARN transcrito. Interrupciones de estos eventos en los seres humanos contribuye a muchas enfermedades, como el cáncer, síndromes metabólicos, y desórdenes en el desarrollo [LEWIN 2004].. 1.1.3 La bioinformática. Dentro de las ciencias computacionales, por los tópicos y conceptos manejados y definidos, esta tesis podría incluirse dentro el dominio de la bioinformática. De acuerdo con el Centro Nacional para la Información Biotecnológica (National Center for Biotechnology Information, 2001) [EL PORTAL 2007], “la bioinformática es un campo de la ciencia en el cual confluyen varias disciplinas tales como la biología, la computación, y la tecnología de la información. El fin de este campo es facilitar el descubrimiento de nuevas ideas biológicas así como crear perspectivas globales a partir de las cuales se puedan discernir principios unificadores en biología”. La bioinformática es una disciplina científica emergente que utiliza la tecnología de la información para organizar, analizar y distribuir información biológica con la finalidad de responder preguntas complejas en biología. Es un área de investigación multidisciplinaria que se puede definir como la interfase entre dos ciencias, la biología y. 3.
(15) la computación, impulsada por el descubrimiento del genoma humano, y la expectativa de que la investigación genómica puede mejorar drásticamente la condición y calidad de vida humana. Como otra definición, bioinformática es la aplicación de computadoras y métodos informáticos en el análisis de datos experimentales y simulación de los sistemas biológicos [BIOINFORMATICS 1998]. Las principales aplicaciones de la bioinformática son la simulación, la minería de datos, y el análisis de datos obtenidos del estudio de las moléculas relevantes para la vida, así como el diseño y desarrollo de herramientas tales como bases de datos. Como ya se ha mencionado, la bioinformática se basa en dos grandes áreas de la ciencia: las ciencias biológicas y las ciencias computacionales, razón por la cual existen dos grandes líneas de investigación. En la primera, las ciencias computacionales utilizan modelos de las biológicas. En la segunda, las ciencias biológicas utilizan modelos y herramientas de las ciencias computacionales.. 1.1.4 Modelos y simulaciones. En esta tesis se utilizarán modelos para representar sistemas biológicos. Por esta razón, conviene conocer de antemano la definición de modelo. Un modelo es una representación de la realidad, que se simplifica de alguna manera para poder estudiarla y manipularla [GARCÍA PELAYO 1994]. Dicho modelo debe ser lo suficientemente complejo para responder a las preguntas e inquietudes que lo produjeron, pero no tanto que no se pueda entender. Un evento se puede considerar como una ocurrencia que cambia el estado del sistema. Existen varios tipos de modelos: los matemáticos, descriptivos, estadísticos, de entrada-salida, de eventos discretos, de eventos continuos, o combinados [BANKS 1998]. Un modelo de eventos discretos intenta representar a los componentes de un sistema y sus interacciones de forma que los objetivos se puedan cumplir.. La mayoría de los modelos. matemáticos, estadísticos y de entrada-salida representan explícitamente las entradas y salidas de un sistema, pero sus componentes internos se representan por medio de relaciones matemáticas o estadísticas. Un ejemplo es el modelo matemático de la física: fuerza = masa x aceleración,. 4.
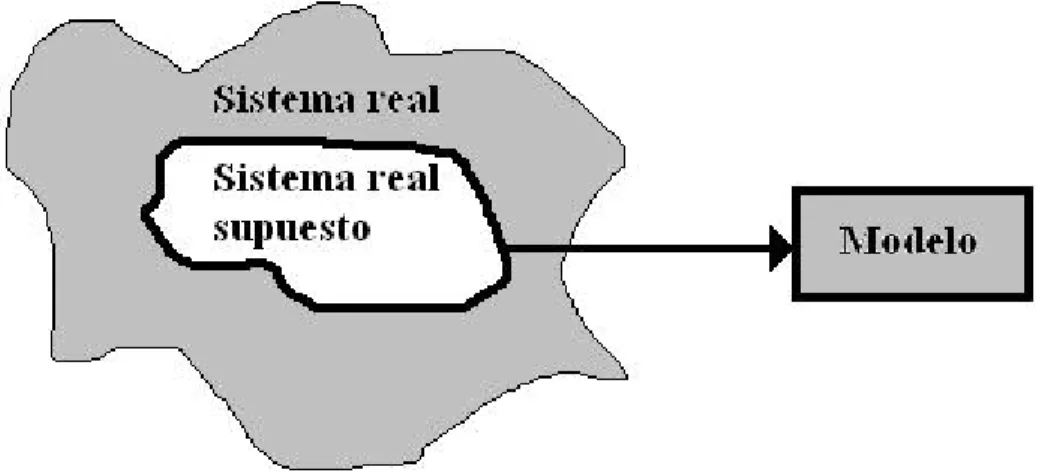
(16) que se basa en la teoría. Un modelo de eventos discretos incluye una representación detallada de los componentes internos reales del sistema. Además, la mayoría de los modelos de eventos discretos son dinámicos, lo que implica que el paso del tiempo juega un papel crucial. Por otro lado, la mayoría de los modelos matemáticos y estadísticos son estáticos debido a que representan al sistema en un punto fijo en el tiempo.. Figura 1.1: Niveles de abstracción de una situación real que lleva a la construcción de un modelo.. En la figura 1.1 se aprecian los niveles de abstracción de una situación de la vida real que nos llevan a la construcción de un modelo. Un modelo se puede entender como una abstracción de una suposición de un sistema real. Un sistema real supuesto es una abstracción de la situación real que se obtiene al concentrarnos en la identificación de los factores que controlan el comportamiento del sistema real. El modelo es una abstracción del sistema real supuesto que identifica las relaciones pertinentes del sistema [TAHA 1995]. Por otra parte, simulación es la imitación de la operación de un sistema o proceso del mundo real a través del tiempo. Es una metodología necesaria para la solución de problemas reales. Se usa para descubrir y analizar el comportamiento de un sistema, realizar preguntas del estilo “qué pasa si...”, y también como ayuda en el diseño de sistemas reales. Tanto sistemas existentes como conceptuales se pueden modelar con simulación [BANKS 1998]. Uno de los conceptos implicado en la simulación es el de modelo, que ha sido abordado previamente en esta sección.. 5.
(17) 1.2 Planteamiento del problema. La dificultad de los sistemas regulatorios se relaciona con el entendimiento de su funcionamiento y su comportamiento dinámico debido a las múltiples interacciones y a la necesidad de predecir fenómenos intrínsecos. Por funcionamiento se entiende conocer qué genes se expresan hasta qué grado, y en qué tiempos. El comportamiento dinámico implica las múltiples interacciones entre ADN, ARN, proteínas y otras moléculas que logran la expresión genética. En términos del problema biológico, complejidad se refiere a la dificultad que presentan las redes regulatorias al involucrar diversos componentes interconectados que contienen problemas de dimensionalidad y acoplamiento, lo que complica el entendimiento intuitivo de su dinámica. Modelar y simular sistemas de regulación genética mediante modelos formales es necesario porque se puede describir sin ambigüedad su estructura.. Además, mediante técnicas. probabilísticas, se podría lograr una predicción de su comportamiento, por ejemplo, el orden cronológico de la expresión de determinados genes, y a partir de qué momento se inicia dicha expresión dadas ciertas condiciones. Para este trabajo de tesis se han seleccionado las redes de Petri por su capacidad para modelar las características dinámicas de los procesos, misma que se aprovechará para modelar el dinamismo de los sistemas regulatorios genéticos. Por estas razones, la pregunta de investigación que se ha planteado es la siguiente: ¿las redes de Petri, como modelo formal computacional, poseen características expresivas para simplificar el entendimiento del funcionamiento y comportamiento de un sistema regulatorio? El término expresividad está planteado como una serie de características que se buscan en el modelo, de forma que la manifestación de éstas pueda saltar a la vista. Con base en una definición de diccionario [GARCÍA PELAYO 1994], la definición propuesta en esta tesis para expresividad es la siguiente: “capacidad de un modelo para manifestar con claridad una serie de características”. Una definición más completa, donde se incluyen las características con que deberá contar el modelo, se presenta en la siguiente sección.. 6.
(18) 1.3 Objetivos. 1.3.1 Objetivo general. Analizar la expresividad de las redes de Petri para modelar sistemas de regulación genética. Para lo anterior, se evaluará el desempeño de los modelos a través de simulaciones que permitan observar la expresividad de los mismos. Los puntos a evaluar son las características que, del concepto de expresividad planteado, deben tener los modelos. Las propiedades de expresividad que el modelo propuesto en este trabajo debe mostrar son las siguientes: •. Capacidad para representar las características dinámicas de los sistemas regulatorios.. •. Capacidad para describir sistemas regulatorios grandes y complejos.. 1.3.2 Objetivos particulares. Una vez claro el objetivo general de la tesis, los siguientes quedan como objetivos específicos: •. Efectuar un mapeo entre la sintaxis y la semántica de las redes de Petri coloreadas, con los elementos de los mecanismos regulatorios estudiados para poder simularlos con esta herramienta.. •. Realizar un modelo basado en redes de Petri coloreadas donde se muestre de forma computacional el funcionamiento de los modelos desarrollados.. •. Evaluar y verificar la estructura y funcionamiento de los modelos desarrollados con expertos en el área de la regulación genética.. 7.
(19) 1.4 Alcances y limitaciones. Los sistemas regulatorios que se estudiaron en este trabajo son los del operón lac y el bacteriófago lambda, modelados con redes de Petri. Para esto, se seleccionaron las redes de Petri coloreadas (CPN), que serán introducidas y descritas posteriormente. Además, se hace un análisis de las redes de Petri híbridas, que fueron utilizadas como herramienta para modelar redes regulatorias por otros autores, y que a su vez motivaron la realización de este trabajo. Se hizo una simulación con un software para desarrollar redes de Petri coloreadas de los sistemas regulatorios seleccionados mencionados anteriormente, buscando observar su expresividad en términos de los conceptos definidos en la Sección 1.3. Además se hizo un análisis de los resultados obtenidos, y se obtuvieron conclusiones. En este punto, dadas las características de la herramienta utilizada (redes de Petri coloreadas), se realizó un modelo discreto, o “discretizado”, tanto del operón lac como del fago lambda. Con esto se espera poder apreciar claramente los estados de activación e inhibición propiciados por las moléculas participantes en cada caso, en cada modelo. En el Capítulo 5 se encuentran las características y el funcionamiento de los sistemas procariontes mencionados, y en el Capítulo 6 se explica el porqué de la discretización mencionada. La principal limitación radica en que las características de la herramienta a utilizar (i.e., CPN Tools v. 124 [CPNTOOLS 2007]), no permiten representar los factores continuos de los sistemas biológicos. Para tratar este problema, se trabajó con modelos discretos. Incluso, la estructura de los modelos fue evaluada y verificada por expertos en biotecnología, con amplio conocimiento en los sistemas biológicos estudiados. Por otra parte, el problema de la predicción del comportamiento de elementos genéticos no será tratado en este trabajo ya que el software seleccionado para simular modelos de redes de Petri coloreadas no cuenta con aspectos de predicción que pudieran hacer posible este tipo de inferencias.. 8.
(20) 1.5 Metodología de desarrollo Para tratar de entender sistemas de regulación genética, ha habido intentos de simular adaptaciones de algunas redes específicas, principalmente relacionadas con bacterias. Los resultados de algunos de estos intentos han sido satisfactorios. Sin embargo, es necesario evaluar qué tan expresivo puede ser un modelo formal de uno de estos sistemas. Para poder enfrentar los problemas que esto conlleva, en este trabajo se seguirá la metodología de desarrollo siguiente: •. Estudiar y comprender el fenómeno de la regulación genética.. •. Estudiar los modelos existentes aplicados al operón lac y el bacteriófago lambda, concretamente, aquellos relacionados con redes de Petri.. •. Caracterizar al fenómeno de regulación en los organismos, destacando las variables relacionadas con la expresividad que se necesitan para el modelado.. •. Realizar un estudio de los trabajos relacionados relevantes a esta tesis, trabajos donde se han usado modelos matemáticos en regulación genética, y concretamente, redes de Petri.. •. Ilustrar el uso de las redes de Petri coloreadas para los modelos del operón lac y el fago lambda a través de simulaciones computacionales.. •. Analizar los resultados y obtener conclusiones sobre la expresividad del modelo propuesto basado en redes de Petri para modelar redes regulatorias a varios niveles de abstracción.. •. Evaluar y comprobar el funcionamiento de los modelos con expertos en biotecnología con amplio conocimiento en los sistemas estudiados (operón lac y fago lambda).. 9.
(21) 2. LA CÉLULA Y REGULACIÓN DE LA EXPRESIÓN GENÉTICA No se sabe con certeza cuándo aparecieron por primera vez las células en la Tierra [KARP 2005]. Sin embargo, a partir de la invención del microscopio electrónico, se inició el estudio de una gran variedad de células. Así se encontró que las hay de dos tipos básicos: las procariotas y las eucariotas. Se diferencian principalmente por su tamaño y tipos de estructuras internas. Existen muchas diferencias básicas entre los dos tipos de células, así como muchas similitudes. Las propiedades que comparten reflejan que las células eucariotas casi con certeza evolucionaron a partir de ancestros procariontes [KARP 2005]. A causa de su ancestro común, ambos tipos de células poseen un lenguaje genético idéntico, un grupo común de vías metabólicas y muchas propiedades estructurales comunes. Por ejemplo, los dos tipos celulares están limitados por membranas plasmáticas de estructura semejante que sirve como una barrera de permeabilidad selectiva entre el medio interior y el exterior. Aunque las paredes celulares de procariotas y eucariotas pueden tener funciones semejantes, su composición química es muy diferente. En su interior, las células eucariotas son mucho más complejas que las procariotas en estructura y función. Esto es puede observar en las figuras 2.1 y 2.2. A pesar de esta diferencia estructural, ambas contienen una región que posee el material genético de la célula rodeado por citoplasma. El material genético de la célula procariota está presente en un nucleoide: una región celular no bien definida, sin membrana, que diferencia al ADN del citoplasma circundante. En contraste, las células eucariotas tienen un núcleo: una región separada por una estructura membranosa compleja llamada envoltura nuclear. Como ya se ha indicado, esta diferencia en la estructura nuclear es la base de los términos procariota y eucariota. Las células procariotas contienen cantidades de ADN relativamente pequeñas: el contenido de ADN en una bacteria se encuentra entre 600,000 y 8’000,000 pares de bases (pb), lo cual es suficiente para codificar cientos o hasta miles de proteínas. Las células eucariotas contienen mucha más información genética. Como ejemplo, un organismo eucarionte relativamente simple, como la levadura, tiene cerca de 12 millones de pb, que codifican alrededor de 6200 proteínas [KARP 2005]. Las células procariotas y eucariotas poseen cromosomas que contienen ADN. Las eucariotas muestran un número determinado de cromosomas, cada uno de los cuales cuenta con una sola molécula lineal de ADN. En contraste, casi todos los procariontes que se han estudiado contienen un cromosoma circular único. 10.
(22) El citoplasma de los dos tipos de células también es muy diferente. El de una eucariota tiene una gran diversidad de estructuras. Incluso los eucariontes más simples son más complejos estructuralmente que una bacteria promedio. Las células eucariotas tienen una disposición de organelos limitados por membrana. Entre éstos están: las mitocondrias, donde la energía química está disponible para alimentar las actividades celulares; un retículo endoplásmico, donde se elaboran muchas de las proteínas y lípidos de la célula; un aparato de Golgi, donde los materiales se clasifican, modifican y transportan a destinos celulares específicos; y diferentas vesículas membranosas de tamaños diversos. Las células vegetales incluyen cloroplastos en su citoplasma, que es donde se realiza la fotosíntesis. Las membranas y vesículas trabajan en el transporte de sustancias de una parte a otra de la célula, y entre su interior y el ambiente. En contraste, el citoplasma de las células procariotas está libre en esencia de estructuras membranosas. Las células eucariotas también contienen estructuras sin membrana.. Los túbulos y. filamentos del citoesqueleto son de este tipo, y participan en la contractilidad, movimiento y soporte celular. El citoesqueleto de las procariotas, que hasta hace poco se pensaba inexistente [KARP 2005], es mucho más simple funcional y estructuralmente. Ambos tipos de células tienen ribosomas, que son partículas no membranosas donde se elaboran las proteínas de la célula. Aunque los ribosomas de las procariotas son más pequeños y con menos componentes, el ensamblado de proteínas ocurre de manera similar en los dos tipos de células. Este mecanismo se explica en la siguiente sección. Existen más diferencias relevantes. Las eucariotas se dividen por un proceso complejo de mitosis, en el cual los cromosomas duplicados se condensan en estructuras compactas separadas por un sistema que contiene microtúbulos. Este aparato conocido como huso mitótico, permite a cada célula hija recibir un ordenamiento equivalente de material genético. En las procariotas no hay compactación de cromosomas ni huso mitótico. El ADN se duplica y las dos copias se separan de manera sencilla y precisa por el desarrollo de una membrana celular entre las dos. La mayor parte de los organismos procariontes corresponde a organismos asexuales, ya que sólo contienen una copia de su único cromosoma y carecen de procesos comparables a la meiosis (que es el proceso durante el cual el número de cromosomas se reduce de tal forma que las células formadas contienen sólo un miembro de cada par de cromosomas homólogos, es decir, cromosomas en los que cada uno lleva una de dos copias del material genético portado por la célula), formación de gametos o fecundación verdadera [KARP 2005]. 11.
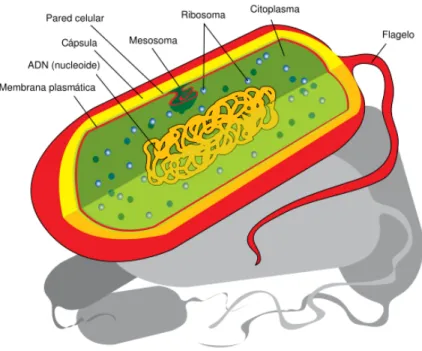
(23) Las células eucariotas poseen diversos mecanismos de locomoción compleja, mientras que los mecanismos de las procariotas son relativamente sencillos. Aquí, el movimiento se lleva a cabo mediante un filamento delgado de proteína llamada flagelo, el cual sobresale de una célula y es capaz de girar. La rotación del flagelo ejerce presión contra el líquido circundante y da lugar a que la célula se impulse a través del medio. Algunas células germinales también poseen flagelos, pero son más complejas que las procariotas, y el movimiento lo genera un mecanismo diferente [KARP 2005]. En los párrafos anteriores se mencionan algunas diferencias entre las células eucariotas y procariotas. A continuación se establecen las características principales de cada tipo de célula, a manera de resumen.. 2.1 Características de las células eucariotas. Figura 2.1: Esquema de una célula ecuariota, donde se aprecian los principales organelos que la conforman.. La fig. 2.1 muestra un esquema de una célula eucariota animal común. A continuación se da una lista de las características principales de las eucariotas que no se encuentran en procariotas. Esta lista básicamente resume las cualidades de este tipo de célula que se explicaron en la sección previa. Las características son: 12.
(24) •. La célula se divide, a grandes rasgos, en núcleo y citoplasma, separado por una envoltura nuclear membranosa que contiene estructuras complejas de poros.. •. Los cromosomas son complejos y están compuestos por ADN y proteínas relacionadas capaces de compactarse en estructuras mitóticas.. •. Los organelos citoplásmicos son membranosos y complejos. Entre ellos están el retículo endoplásmico, y el aparato de Golgi, entre otros.. •. Hay organelos citoplásmicos especializados para respiración aerobia (las mitocondrias) y, en el caso de las plantas, para fotosíntesis (los cloroplastos).. •. Tienen un sistema complejo de citoesqueleto, que incluye microfilamentos, filamentos intermedios, y microtúbulos.. •. Los flagelos, cuando existen, son complejos.. •. Son capaces de ingerir materiales líquidos y sólidos, y atraparlos dentro de vesículas membranosas plasmáticas por los procesos de endocitosis (mecanismo para captar líquidos y sólidos dentro de la célula) y fagocitosis (proceso mediante el cual las células captan materiales en partículas).. •. Las células vegetales, en concreto, las plantas, tienen paredes celulares que contienen celulosa.. •. La división celular utiliza un huso mitótico que contiene microtúbulos para separar cromosomas.. •. Hay presencia de dos copias de genes por célula (diploidía), cada gen proviene de cada uno de los dos padres.. •. Hay presencia de enzimas diferentes para sintetizar ARN (las polimerasas de ARN, que serán abordadas más adelante en el capítulo, véase el apartado 2.4.5).. •. La reproducción sexual requiere meiosis y fecundación.. •. La base de la expresión genética es el dogma central, que es la vía por la cual el ADN se codifica en proteínas a través de moléculas intermediarias de ARN.. •. Éstas deben ser maduradas, de forma que las secciones con información relevante (exones) sean separadas de las secciones que no codificarán (intrones) para formar parte de la molécula de ARN.. •. Un segmento de la secuencia llamado promotor le indica a ciertas enzimas llamadas polimerasas cuál es el gen que debe ser transcrito, y posteriormente, traducido. 13.
(25) 2.2 Características de las células procariotas. Figura 2.2: Esquema de una célula procariota, con sus principales componentes.. Las características comunes a eucariotas y procariotas son las siguientes: •. La membrana plasmática es de estructura similar en ambos casos.. •. La información genética está codificada en el ADN mediante códigos genéticos idénticos.. •. Los mecanismos de trascripción y traducción de la información genética son similares.. •. Los ribosomas son semejantes, como lo es la elaboración de proteínas.. •. Existen algunas vías metabólicas que comparten ambos tipos de células (como la glicólisis y el ciclo de ciertos ácidos [KARP 2005]).. •. La conservación de la energía química en ATP (adenosín trifosfato) se hace mediante un aparato similar, localizado en la membrana plasmática en las procariotas y el la membrana mitocondrial en la eucariotas.. •. Los mecanismos de fotosíntesis son parecidos entre las cianobacterias y las plantas verdes.. •. Al igual que en el caso de las eucariotas, la expresión genética se lleva a cabo con base en el dogma central.. 14.
(26) Ahora, algunas características propias de las procariotas son las siguientes: •. El ADN se encuentra sin acompañamiento proteico, formado en una sola cadena circular.. •. La división celular ocurre por fisión binaria.. •. No hay organelos reconocibles en el citoplasma, además de algunos agregados celulares menores, como mesosomas, que son pliegues espirales hacia el interior de la célula en la membrana, compuestos de lípidos y proteínas, que funcionan como sitios de inicio para la división celular.. •. En algunos casos existe una especie de compartimiento acuoso que rodea al citoplasma, separado por la pared celular y la membrana plasmática.. •. Se alimentan por absorción, a través de la pared celular.. •. Los organismos procariontes son unicelulares. Su metabolismo es muy variado, y muchos soportan condiciones ambientales que les imprimen tensión, o estrés a través de factores como temperatura o acidez.. •. Las estructuras de los mecanismos regulatorios son diferentes. Los procariontes agrupan sus mecanismos regulatorios en secuencias de genes controladas por un operador y un represor llamadas operones.. •. La polimerasa de ARN reconoce una secuencia a la cual se une para iniciar la transcripción de los genes estructurales.. •. Posteriormente, el transcrito maduro será traducido a cadenas de aminoácidos que formarán las proteínas requeridas para el funcionamiento del organismo.. A pesar de las diferencias, la estructura genética es idéntica en procariotas y eucariotas.. 15.
(27) 2.3 Estructura genética. Se demostró que el ADN es el material genético a través de una serie de experimentos [HARTL 2001], pero a principios del siglo XX aún había algunas preguntas sin responder. En la estructura de Watson-Crick se propuso al ADN como dos largas cadenas de subunidades, cada una enroscada alrededor de la otra para formar una doble hélice. La doble hélice sigue el sentido de una rosca a derechas vista desde abajo. Las subunidades de cada hebra son los nucleótidos, cada uno de los cuales contiene uno de cuatro constituyentes químicos, llamados bases, adjuntos a una molécula fosforilada de cinco azúcares llamada desoxirribosa. Las cuatro bases del ADN son: adenina (A), guanina (G), timina (T), citosina (C).. En cualquier posición de las hebras del ADN, el. apareamiento es entre la A y la T, y entre la G y la C. Es decir, A–T , y G–C se dicen complementarias, significando que cada base a lo largo de la hebra de ADN se encuentra apareada con otra base en la hebra opuesta.. Figura 2.3: Estructura de la doble hélice del ADN. Ilustra las cuatro bases nitrogenadas y el esqueleto.. En la doble hélice del ADN, (Fig. 2.3), cada hebra de ADN tiene una polaridad que está determinada por la dirección en la que los nucleótidos están apuntando. A la “cabeza” se le 16.
(28) denomina el extremo 3’, y a la “cola” se le denomina el extremo 5’. En la doble hélice del ADN, las hebras apareadas están orientadas en direcciones contrarias: el extremo 5’ de una hebra está alineada con el extremo 3’ de la otra hebra. Los fundamentos bioquímicos de esta polaridad y el razonamiento para la orientación contraria de las hebras en la doble hélice sobrepasan los límites de este trabajo. La forma en que funciona la estructura del ADN obedece a los siguientes puntos: la secuencia de bases en el ADN se puede copiar usando por separado las dos hebras complementarias como plantilla para la creación de una nueva hebra con una secuencia complementaria de bases. El ADN puede contener información genética codificada en la secuencia de bases.. Cambios en la. información genética (o mutaciones) pueden resultar de errores en el copiado donde se altera la secuencia de bases.. 2.3.1 Las proteínas. Lo que se crea del código complejo de ADN son proteínas, una clase de macromoléculas que lleva a cabo la mayoría de las actividades en la célula. Las células están hechas casi en su totalidad de proteínas. Las proteínas estructurales dan a la célula rigidez y movilidad, las proteínas que forman poros en la membrana controlan el tráfico de otras moléculas hacia el interior o el exterior de la célula, y las proteínas receptoras regulan la actividad celular en respuesta a señales moleculares del medio exterior, o de otras células. Las proteínas también son responsables de la mayoría de las actividades metabólicas de las células. Son esenciales para la síntesis y simplificación de moléculas orgánicas y para regular la energía química necesaria en la actividad celular. Las enzimas, que también se demostró que son proteínas, son los catalizadores que aceleran las reacciones bioquímicas en la célula [HARTL 2001]. La forma en que los genes codifican en proteínas se resume en el dogma central.. 17.
(29) 2.4 El dogma central de la biología molecular. El ADN codifica proteínas indirectamente a través de los procesos de transcripción y traducción. La ruta indirecta de transferencia de información. se conoce como el dogma central de la biología molecular [HARTL 2001]. El término dogma se entiende como “conjunto de creencias”. Se le denomina de esta forma porque en sus inicios surgió como una teoría, misma que ha sido confirmada experimentalmente desde entonces. A pesar de esto, el término persiste. El dogma central es el principio fundamental de la genética porque resume cómo la información genética en el ADN se expresa en la secuencia de aminoácidos de una cadena de polipéptidos. El concepto principal en el dogma central es que el ADN no se codifica en proteínas directamente, sino que lo hace a través de moléculas intermediarias de ácido ribonucleico (ARN). La estructura de esta molécula es similar a la del ADN. Las diferencias consisten en el azúcar (el ARN contiene ribosa, mientras que el ADN desoxiribosa), el ARN es una cadena sencilla (y no doble como el ADN), y finalmente, el ARN contiene a la base uracilo (U), en lugar de la timina (T) del ADN. Tres tipos de ARN toman parte en la síntesis de proteínas: •. Una molécula de ARN mensajero (ARNm) lleva la información genética del ADN y se usa como plantilla para la síntesis de polipéptidos. Hay una proporción alta de nucleótidos que se codifican como aminoácidos.. •. Varios tipos de ARN ribosomal (ARNr) junto con algunas proteínas, constituyen las partículas llamadas ribosomas, que es donde la síntesis de polipéptidos ocurre.. •. Un conjunto de moléculas de ARN de transferencia ARNt), cada una de las cuales lleva un aminoácido en particular y una región de reconocimiento de tres bases que se aparearán con un grupo de tres bases adyacentes en el ARNm. El aminoácido de cada una de las subunidades de ARNt que participa en la traducción, se agrega al final de la cadena de polipéptidos en crecimiento.. 18.
(30) Figura 2.4: Dogma central de la biología molecular (modificado de [HARTL 2001]).. La fig. 2.4 ilustra el dogma central. Como resumen, la secuencia de nucleótidos en un gen especifica la secuencia de nucleótidos en una molécula de ARN mensajero; a su vez, la secuencia de nucleótidos en el ARN mensajero especifica la secuencia de aminoácidos en la cadena de polipéptidos.. 2.4.1 Transcripción. El proceso de sintetizar una hebra de ARN de una molécula de ADN se llama transcripción. La secuencia de bases en el ARN es complementaria a la plantilla de ADN, excepto que la U (en lugar de la T) se aparea con la A. Cada hebra de ARN tiene polaridad (un extremo 5’ y un extremo 3’) y, como en la síntesis de ADN, los nucleótidos se van agregando al extremo 3’ de la hebra creciente de ARN. Así, el extremo 5’ de la hebra de ARN se sintetiza primero, y la transcripción ocurre a lo largo de la molécula de ADN en la dirección 3’ a 5’. Cada gen incluye secuencias de nucleótidos que indican el principio y el fin de la transcripción. Para cada gen, la longitud de la transcripción de ARN 19.
(31) es más pequeña que la longitud del ADN en el cromosoma [HARTL 2001]. La fig. 2.5 muestra un esquema del proceso de transcripción en el dogma central.. Figura 2.5: Esquema de la transcripción. La doble hélice se extiende para que la ARN polimerasa pueda iniciar la transcripción de una molécula de ARN a partir de la cadena molde. Después de la transcripción, la molécula de ADN vuelve a su estado original.. 2.4.2 Traducción. La síntesis de un polipéptido bajo la dirección de una molécula de ARN se llama traducción. A pesar de que la secuencia de bases en el ARNm codifica para la secuencia de aminoácidos, las moléculas que de hecho hacen la traducción son las de ARNt. La molécula de ARNm se traduce en grupos de tres bases llamados codones. Para cada codón en el ARNm que especifica un aminoácido, hay una molécula de ARNt que contiene el grupo complementario de tres bases 20.
(32) adyacentes que se aparean con el codón. El aminoácido correcto se agrega al otro extremo del ARNt. El ARNm se lee codón por codón. Cada codón que especifica un aminoácido se aparea con un grupo complementario de tres bases en una molécula de ARNt simple. Un extremo del ARNt se agrega al aminoácido correcto, de forma que el aminoácido se sume a la línea. La cadena de polipéptidos crece a ritmo de un aminoácido a cada paso hasta que se encuentra con cualquiera de los tres codones que especifican el alto. En este punto, termina la síntesis de la cadena de aminoácidos y la cadena polipéptida se libera del ribosoma.. Figura 2.6: Esquema de la traducción. Para cada codón en el ARNm que especifica un aminoácido, hay una molécula de ARNt que contiene el grupo complementario de tres bases adyacentes que se aparean con el codón. El aminoácido correcto se agrega al otro extremo del ARNt.. 2.4.3 El código genético. La tabla completa que muestra la decodificación de los codones en el ARNm en aminoácidos se conoce como código genético. La tabla se muestra en la fig. 2.7. Para cualquier codón, la columna de la izquierda corresponde al primer nucleótido en el codón (leído desde el extremo 5’), la fila de la parte superior corresponde al segundo nucleótido, y la columna de la derecha corresponde al tercer nucleótido.. 21.
(33) Figura 2.7: Código genético estándar. El codón está dado por el cuerpo de la tabla, junto con el aminoácido que especifica. La columna de la izquierda corresponde al primer nucleótido en el codón (leído desde el extremo 5’), la fila de la parte superior corresponde al segundo nucleótido, y la columna de la derecha corresponde al tercer nucleótido. Cada aminoácido está designado por su nombre completo.. 2.4.4 Promotores, operadores, represores, operones, exones, intrones. Existen otros tipos de moléculas y proteínas importantes que aparecen en un sistema regulatorio. Un promotor es una secuencia de ADN a la cual se une la ARN polimerasa para iniciar la transcripción. Un operador es un segmento regulatorio en el ADN que interactúa con una proteína represora específica para controlar la transcripción de los genes estructurales adyacentes. Un represor es una proteína que se une específicamente a una secuencia regulatoria adyacente a un gen (concretamente, el operador), y bloquea la transcripción del gen. Un operón es un conjunto de genes estructurales adyacentes regulados por un operador y un represor, que regula la expresión genética en procariontes. Un poco más adelante se hablará de las polimerasas. Las secuencias de ADN tienen también denominación, de acuerdo con su función al final del proceso de transcripción. Un exón es cualquier región de ADN que se transcribe a la molécula final de ARNm, en lugar de ser removida, y contiene la información para producir la proteína codificada 22.
(34) en el gen. Cada exón codifica un fragmento específico de la proteína completa. Principalmente en las células eucariotas, los exones están separados por largas regiones de ADN llamadas intrones. Un intrón es una secuencia de ADN en un gen que no codifica, que se trascribe, pero que es posteriormente extirpada al formar la molécula de ARNm madura.. 2.4.5 Polimerasas Una polimerasa es una enzima que funciona como catalizador de un enlace covalente de nucleótidos. Dos ejemplos son: la ADN polimerasa y la ARN polimerasa. La ADN polimerasa es una enzima que cataliza la síntesis de ADN a partir de desoxirribonucleótidos y de una molécula de ADN que sirve como plantilla para ser copiada. Esta enzima interviene en el proceso de replicación (o duplicado de una molécula de ADN) como agente catalizador, acelerando los procesos. Mientras que en las procariotas hay cinco tipos de ADN polimerasas, en las eucariotas se conocen quince tipos de la enzima. La reacción en cadena de la polimerasa (PCR, polymerase chain reaction), es una técnica de replicación molecular cuyo objetivo es generar un gran número de copias de un fragmento de ADN particular partiendo de un mínimo. La ADN polimerasa se usa en esta técnica por sus propiedades para replicar hebras de ADN. La enzima opera de la siguiente manera: en cada replicación, la ADN polimerasa y otras enzimas participantes sintetizan dos nuevas hebras de ADN complementarias respecto a las dos originales. Durante el proceso, la ADN polimerasa reconoce un nucleótido no apareado de la cadena original y la combina con un nucleótido libre que tiene la base complementaria correcta. Luego, la enzima cataliza la formación de nuevos enlaces covalentes que ligan el fosfato del nucleótido libre entrante con el azúcar del nucleótido agregado en la cadena en crecimiento [VÁZQUEZ-CONTRERAS 2003a]. Por otro lado, la ARN polimerasa es una enzima capaz de polimerizar los ribonucleótidos para sintetizar ARN a partir de una secuencia de ADN que sirve como plantilla.. Además de la. polimerización de los ribonucleótidos, la ARN polimerasa tiene otras funciones, como localizar y unirse a promotores de la molécula de ARN, desenrollar parcialmente la molécula de ADN que sirve como plantilla, y terminar la cadena. La ARN polimerasa cataliza consecutivamente la elongación de la hebra de ARN, al mismo tiempo que enrolla y desenrolla la doble cadena de ADN, y termina la transcripción. En las células procariotas, la misma enzima cataliza los tres diferentes tipos de ARN, 23.
(35) mientras que en las eucariotas, se tiene un tipo de ARN polimerasa para cada tipo de ARN. La reacción química que cataliza la ARN polimerasa consiste en la unión de ribonucleótidos adenosín trifosfato (ATP), uracilo trifosfato (UTP), guanina trifosfato (GTP), y citosina trifosfato (CTP), liberándose los grupos fosfato [VÁZQUEZ-CONTRERAS 2003b]. Acerca de los tres diferentes tipos de ARN, y sus funciones, ya se ha hablado en secciones anteriores.. 24.
Figure
![Figura 2.8: Niveles en que ocurre la regulación de la expresión genética: a nivel transcripcional, a nivel del procesamiento y a nivel traduccional [KARP 2004]](https://thumb-us.123doks.com/thumbv2/123dok_es/3206287.581377/37.918.250.668.482.948/figura-niveles-regulación-expresión-genética-transcripcional-procesamiento-traduccional.webp)
![Figura 3.2: CPN que describe el sistema de asignación de recursos en su marca inicial M 0 [JENSEN 1997]](https://thumb-us.123doks.com/thumbv2/123dok_es/3206287.581377/47.918.141.779.103.726/figura-describe-sistema-asignación-recursos-marca-inicial-jensen.webp)
+7
![Figura 3.3: Transición T2 de la fig. 3.2 con arcos y expresiones de arco correspondientes [JENSEN 1997]](https://thumb-us.123doks.com/thumbv2/123dok_es/3206287.581377/50.918.201.720.292.600/figura-transición-fig-arcos-expresiones-arco-correspondientes-jensen.webp)
![Figura 3.4: Transición T1 de la fig. 3.2 con sus arcos, expresiones de arco y guardia correspondiente [JENSEN 1997]](https://thumb-us.123doks.com/thumbv2/123dok_es/3206287.581377/52.918.176.749.102.529/figura-transición-arcos-expresiones-arco-guardia-correspondiente-jensen.webp)
![Figura 5.7: Etapa temprana de expresión genética del fago lambda en HPN [MATSUNO 2000]](https://thumb-us.123doks.com/thumbv2/123dok_es/3206287.581377/72.918.193.705.115.845/figura-etapa-temprana-expresión-genética-fago-lambda-matsuno.webp)
![Figura 5.15: Representación en HFPN de la regulación negativa del operón lac [DOI 2004]](https://thumb-us.123doks.com/thumbv2/123dok_es/3206287.581377/84.918.134.778.234.727/figura-representación-hfpn-regulación-negativa-operón-lac-doi.webp)
![Figura 5.16: Representación en HFPN de los mecanismos de transcripción, traducción e hidrólisis de lactosa en el operón lac [DOI 2004]](https://thumb-us.123doks.com/thumbv2/123dok_es/3206287.581377/86.918.132.776.119.883/figura-representación-mecanismos-transcripción-traducción-hidrólisis-lactosa-operón.webp)

Documento similar
"No porque las dos, que vinieron de Valencia, no merecieran ese favor, pues eran entrambas de tan grande espíritu […] La razón porque no vió Coronas para ellas, sería
Cedulario se inicia a mediados del siglo XVIL, por sus propias cédulas puede advertirse que no estaba totalmente conquistada la Nueva Gali- cia, ya que a fines del siglo xvn y en
que hasta que llegue el tiempo en que su regia planta ; | pise el hispano suelo... que hasta que el
por unidad de tiempo (throughput) en estado estacionario de las transiciones.. de una red de Petri
Missing estimates for total domestic participant spend were estimated using a similar approach of that used to calculate missing international estimates, with average shares applied
The part I assessment is coordinated involving all MSCs and led by the RMS who prepares a draft assessment report, sends the request for information (RFI) with considerations,
Con todo, los objetivos de este estudio son comprobar qué estándares de masculinidad hegemónica reproducen los youtubers españoles más relevantes y descubrir en qué medida
Esta información resulta útil para el diseñador de la interfaz, que puede decidir modificar el modelo CUI (en el caso del proceso “1” de la Figura 3), o bien modelos de más
