Efecto de la Radiación UV C en la Bioacumulación de Mn (II) en bacterias aisladas de las Pampas de la Joya – Arequipa
121
0
0
Texto completo
(2) ASESORA DE TESIS Dra. María Valderrama Valencia.
(3) JURADO DE SUSTENTACIÓN. PRESIDENTE Dr. Ronald D. Navarro Obiedo. SECRETARIO Dra. María del Carmen Eguren Verano. ASESORA DE TESIS Dra. María Valderrama Valencia.
(4) El presente trabajo ha sido realizado en el Laboratorio de Genética de la Escuela Profesional de Biología, de la Universidad Nacional de San Agustín (UNSA), bajo la supervisión de la Dra. María Rosario Elsa Valderrama Valencia, dicha investigación ha sido financiado por la Universidad Nacional de San Agustín luego de haber sido seleccionado en la Convocatoria EO53-2016-02 lanzada por la Universidad Nacional de San Agustín de Arequipa y el Consejo Nacional de Ciencia, Tecnología e Innovación Tecnológica – CONCYTEC y el Fondo Nacional de Desarrollo Científico, Tecnológico y de Innovación Tecnológica – FONDECYT al suscribir el Convenio Marco de Cooperación Interinstitucional..
(5) DEDICATORIA:. A mis padres, José Luis y Felipa Domitila. A mis tíos, Pedro y Valeriana. A mis primos, Noé y Nancy. A mi enamorada Magaly..
(6) AGRADECIMIENTOS:. Gracias a Dios, a mis padres José Luis y Felipa Domitila, a mis tíos Pedro Arapa y Valeriana Mamani, y a mi enamorada Magaly con quienes he vivido y me apoyaron y alentaron para culminar satisfactoriamente mis estudios; al equipo de.
(7)
(8) ÍNDICE. RESUMEN...........................................................................................................................................10 ABSTRACT .........................................................................................................................................11 INTRODUCCIÓN ..............................................................................................................................12 OBJETIVOS........................................................................................................................................15 OBJETIVO GENERAL ................................................................................................................15 OBJETIVO ESPECÍFICOS ..........................................................................................................15 CAPÍTULO I .......................................................................................................................................16 MARCO TEÓRICO ...........................................................................................................................16 1.1. EXTREMÓFILOS Y AMBIENTES EXTREMOS 16 1.2. CATEGORÍAS DE EXTREMÓFILOS Y EXTREMOTROFOS 17 1.3. DISTRIBUCIÓN DE EXTREMÓFILOS Y EXTREMOTROFOS 18 1.4. DIVERSIDAD MICROBIANA DE AMBIENTES EXTREMOS 19 1.5. EL DESIERTO DE LAS PLAMPAS DE LA JOYA ....................................................................20 1.5.1. Temperatura: .......................................................................................................................21 1.5.2. Humedad: ............................................................................................................................21 1.5.3. La radiación solar y dirección del viento ............................................................................22 1.6. RADIACIÓN ULTRAVIOLETA (UVR)......................................................................................23 1.6.1. La luz ..................................................................................................................................23 1.6.2. Radiación UV-C..................................................................................................................24 1.6.3. Inactivación cinética ...........................................................................................................26 1.6.4. Daño celular por la Radiación Ultravioleta .........................................................................27 1.7. MICROORGANISMOS EXTREMÓFILOS RESISTENTES A LA RADIACIÓN 28 1.8. MECANISMOS DE DEFENSA CONTRA LA RADIACIÓN ULTRAVIOLETA 30 1.8.1. Proteínas de reparación del DNA dañado ...........................................................................30 1.8.2. Multiplicidad, alineación y estructura del genoma .............................................................32 1.9. IDENTIFICACIÓN DE MICROORGANISMOS 33 1.9.1. Identificación morfológica ..................................................................................................33 1.9.2. Identificación molecular .....................................................................................................33 1.9.2.1. Características del ARNr 16S ..........................................................................................33 1.9.2.2. Análisis bioinformatico de secuencias .............................................................................34 1.9.3. Árboles filogenéticos y cladogramas ..................................................................................35 1.10. EL MANGANESO Y LA RESISTENCIA A LA RADIACIÓN: 36 1.10.1. Complejos de manganeso..................................................................................................37 1.10.2. Bioproducción de Manganeso ........................................................................................... 40 i.
(9) 1.10.3. Medición de la producción de manganeso: ....................................................................... 41 1.11. EFECTO DE LA RADIACIÓN Y EL SER HUMANO 43 1.11.1. La radiación ultravioleta y el melanoma maligno cutáneo ................................................ 43 1.12. IMPLICANCIAS BIOTECNOLÓGICAS DE MICROORGANIMSO EXTREMÓFILOS RESISTENTES A LA RADIACIÓN 46 1.12.1. Biomedicina: ..................................................................................................................... 46 1.12.2. Implicancias terapéuticas .................................................................................................. 48 CAPITULO II ..................................................................................................................................... 51 MATERIALES Y MÉTODOS .......................................................................................................... 51 2.1. LUGAR DE EJECUCIÓN: 51 2.2. METODOLOGÍA: 52 2.2.1. Zona de muestreo ................................................................................................................ 52 2.2.2. Toma de muestras: .............................................................................................................. 52 2.2.3. Aislamiento de microrganismos nativos del Desierto de las Pampas de la Joya ................. 54 2.2.4. Caracterización de las colonias aisladas.............................................................................. 54 2.2.4.1. Caracterización morfológica ................................................................................... 54 2.2.4.1.1. Caracterización morfológica macroscópica .......................................................... 54 2.2.4.1.2. Caracterización morfológica microscópica ....................................................... 54 2.2.4.2. Caracterización bioquímica:.................................................................................... 55 2.2.5. Selección de bacterias más viables a radiación UV-C ........................................................ 55 2 .2.5.1. Acondicionamiento de la Cámara de luz UV-C ........................................................ 55 2.2.5.2. Identificación de las bacterias más viables .............................................................. 56 2.2.5.3. Curva de supervivencia a radiación UV-C de las bacterias más viables ..................... 57 2.2.6. Caracterización molecular de las cepas más viables: .................................................. 58 2.2.6.1. Cultivo en medio líquido de las cepas más viables ............................................... 58 2.2.6.2 Extracción de ADN de las cepas más viables ......................................................... 58 2.2.6.3. Evaluación del ADN extraído de las cepas extremófilas ....................................... 59 2.2.6.4. Amplificación del segmento genético ADNr 16S .................................................. 60 2.2.6.5. Condiciones para Reacción en Cadena de la polimerasa (PCR) ............................... 60 2.2.6.6. Visualización de los productos amplificados por la PCR ....................................... 60 2.2.6.7. Secuenciación del segmento genético ADNr 16s de las bacterias más viables ...... 61 2.2.6.8. Análisis de homología y construcción del cladograma ......................................... 61 2.2.7. Medición de la concentración de Mn (II) ...............................................................61 2.2.7.1. Curva de calibración ............................................................................................ 61 2.2.7.2. Extracción de Manganeso (Mn II) a partir de las bacterias más viables a UV-C ..... 63 2.2.7.3. Determinación de Mn (II) ..............................................................................63 ii.
(10) CAPÍTULO III .................................................................................................................................... 64 RESULTADOS ................................................................................................................................... 64 3.2. Aislamiento de las bacterias nativas del Desierto de las Pampas de la Joya 64 3.3. Caracterización de las colonias aisladas 65 3.3.1. Caracterización morfológica ............................................................................................... 65 3.3.2. Caracterización bioquímica................................................................................................. 69 3.4. Identificación de las bacterias más viables 70 3.5. Curva de supervivencia a la radiación de las dos cepas más viables ..................................... 72 3.6. Caracterización molecular de las cepas más viables a radiación UV-C: ................................ 72 3.6.1. Extracción del ADN de las cepas bacterianas extremófilas más viables ....................... 72 3.6.2. Amplificación del segmento genético ADNr 16S ......................................................... 73 3.6.3. Secuenciación de los segmentos genéticos ADNr 16S ................................................ 74 3.6.4. Análisis de homología de las secuencias del segmento genético ADNr 16S en el banco de datos. ....................................................................................................................76 3.6.5. Construción del cladograma ...................................................................................... 78 3.7. Medida de la concentración intracelular de Mn (II) ............................................................... 79 CAPÍTULO IV .................................................................................................................................... 80 DISCUSIÓN ........................................................................................................................................ 80 CONCLUSIONES............................................................................................................................... 86. ii i.
(11) ÍNDICE DE TABLAS. Tabla 1. Implicancias terapeuticas de microorganismos extremófilos aislados resistentes a radiacion UV. (Fuente: Shick y Dunlap, 2002) .................................................................................................... 48 Tabla 2. Códificación de las muestras, ubicación y características de los puntos muestreados de las Pampas del Desierto de la Joya. ............................................................................................................ 53 Tabla 3. Soluciones preparadas para la curva de calibración de Manganeso (II) ................................. 62 Tabla 4. Código y número de morfotipos de aislados axénicos obtenidos por puntos de suelo muestreado del Desierto de las Pampas de la Joya. .............................................................................. 65 Tabla 5. Caracterización morfológica de las cepas bacterianas del Desierto de las Pampas de la Joya. .............................................................................................................................................................. 66 Tabla 6. Caracterizacion de la Prueba de Gram para los 40 aislados axénicos. ................................... 68 Tabla 7. Caracterización de la reacción de la prueba de catalasa de las 40 cepas aisladas axénicas. .. 69 Tabla 8. Pureza y cantidad de ADN extraído de las cepas JR1_1 y JR1_10 aisladas del Desierto de las Pampas de la Joya. ................................................................................................................................ 73 Tabla 9. Resumen de la comparación, por homología, de las secuecnias del gen ADNr 16S de las cepas bacterianas JR1_01 y JR1_10 con las secuencias depositadas en bases de datos. ................................ 76. iv.
(12) ÍNDICE DE FIGURAS. Fig. 1. Esquema del origen de los diferentes tipos de radiación y sus efectos sobre extremófilos. (Fuente: Lopez-Malo y Palou, 2005) .................................................................................................................. 24 Fig. 2. Dimerización fotoquímica de timina inducida por UV (Lopez-Malo y Palou, 2005) ............... 25 Fig. 3. Poder germicida de las diferentes longitudes de onda. (Fuente: Bolton, 2001) ......................... 26 Fig. 4. Las curvas de supervivencia durante los tratamientos típicos UVC. (Fuente: Shama, 1999) ... 27 Fig. 5. Micrografía electrónica de transmisión de D. radiodurans La escala es de 0,5 µm. (Fuente: Horikoshi et al., 2011) .......................................................................................................................... 30 Fig. 6. Mapas de Fluorescencia de Rayos X de la distribución cualitativa y de los gradientes de manganeso en D. radiodurans. La abundancia aproximada de Mn se observa líneas de colores; la línea verde muestra una concentración de 3 mM; la línea azul oscuro, 2 mM; la línea malva, 1 mM y la linea azul claro, 0,3 mM para el Fe (rojo, 0,5 mM). Escala de barra 0.5 mm. (Fuente: Daly et al., 2007) ... 37 Fig. 7. Inmunoensayo de transferencia de Western de grupos carbonilo unidos a proteínas en extractos celulares (Fuente: Daly et al., 2004) ..................................................................................................... 39 Fig. 8. Red de protección la radiación en D. radiodurans, incluidos los genes y antioxidantes que evitan el daño celular (Luan et al., 2014) ........................................................................................................ 40 Fig. 9. Diagrama esquemático de un espectrofotómetro convencional (Fuente: Wiegand et al., 2017). .............................................................................................................................................................. 43 Fig. 10. Respuestas moleculares y cancerígenos de los melanocitos a la radiación UV (Fuente: Matsumura y Ananthaswamy, 2004) .................................................................................................... 45 Fig. 11. Estructura química de escitonemina. (Fuente: Fleming y Castenholz, 2008) .......................... 49 Fig. 12. Tipos de suelo muestreado en el Desierto de las Pampas de la Joya A. Primer tipo de tierra (JR1) (color rojo, la más abundante), B. Tierra blanca (JB) (suelo superficial), C. Tierra ploma arenosa (JP), D. Tierra rojo-rosada (JR2). ......................................................................................................... 52 Fig. 13. Vista satelital de las Pampas de la Joya. Los círculos en color indican los cuatro puntos de muestreo. Los triángulos indican el grado de aridez de los suelos reportados por Valdivia et al. (2012) .................................................................................................................................................... 53 Fig. 14. Cámara de radiación UV-C. En la figura se muestra las dimensiones de la cámara y la distancia entre la lámpara y las muestras tratadas ................................................................................. 56 Fig. 15. Aislamiento de microrganismos crecidos en tres medios de cultivo (MA. R2A y LB) con tres repeticiones de cada suelo de las Pampas de la Joya. ........................................................................... 65 Fig. 16. Células bacterianas Gram positivas. Objetivo x100. ............................................................... 67 Fig. 17. Células bacterianas Gram negativas. Objetivo x100. .............................................................. 68 Fig. 18. Fotografia que muestra la prueba de la catalasa en placa. La formación de burbujas de oxígeno, indica la presencia de la enzima catalasa (catalasa positiva) .................................................. 70 Fig. 19. Factores de supervivencia de los microorganismos axénicos aislados de las Pampas del Desierto de la Joya. ............................................................................................................................... 71 Fig. 20. Curvas de supervivencia a la radiación UV-C de los aislados bacterianos JR1-01 y JR1-10. 72 Fig. 21. Electroforesis en gel de agarosa de los fragmentos de ADN y ADNr 16S. En el carril M se muestra el marcador de pb (UltraRanger 1kb DNA Ladder), en el carril 1 y 2 se muestran el ADN de las cepas JR1_10 y JR1_1, respectivamente, el carril 4 y 5 muestran los fragmentos amplificados y el carril 3 y 6 son los controles negativos del ADN y ADNr 16S, respectivamente.............................. 73 Fig. 22. Alineamiento parcial de la secuencia del gen ADNr 16S de la cepa bacteriana JR1_01 y JR1_10 provenientes de las Pampas de la Joya..................................................................................... 74 v.
(13) Fig. 23. Electrofenograma parcial de las secuencias nucleotídicas del ADNr 16S de la cepa bacteriana JR1_10 .................................................................................................................................................. 75 Fig. 24. Electrofenograma parcial de las secuencias nucleotídicas del gen ADNr 16S de la cepa bacteriana JR1_01 ................................................................................................................................. 75 Fig. 25. Alineamiento parcial de la secuencia del gen ADNr 16S de la cepa bacteriana JR1_01 proveniente de las Pampas de la Joya, con las secuencias depositas en la base de datos NCBI ............ 77 Fig. 26. Alineamiento parcial de la secuencia del gen ADNr 16S de la cepa bacteriana JR1_10 proveniente de las Pampas de la Joya, con las secuencias depositas en la base de datos NCBI ............ 77 Fig. 27. Cladograma construida en base las secuencias del segmento genético ADNr 16S de las cepas bacterianas JR1_10 y JR1_01 aisladas de las Pampas de la Joya. ........................................................ 78 Fig. 28. Efecto de la radiación UV-C sobre la concentración intracelular de manganeso (mM) en cepas de R. kroppenstedtii y M. radiotolerans aislasdas a partir de suelos de las Pampas de la Joya. Los resultados corresponden a promedios de tres repeticiones. La barra de color verde representa a los niveles de Mn en las cepas control y las demás barras de color representan los niveles de Mn de bacterias tratadas con diferentes dosis de radiación UV-C. .................................................................. 79. vi.
(14) ABREVIATURAS. TAE. : Tris ácido acético – EDTA.. EDTA. : Ácido etilendiaminotetraacético.. TE. : Tris – EDTA.. ADN. : Ácido desoxiribonucleico.. ARN. : Ácido ribonucleico. UFC. : Unidades formadoras de colonia. pH. : Potencial de hidrogeniones. PCR. : Polymerase Chain Reaction. Km. : Kilómetros. °C. : Grados centígrados. ENOS. : El Niño-Oscilación del Sur. W/m2. : Watts por metro cuadrado. m/s2. : metro por segundo al cuadrado. UV. : Ultravioleta. UVR. : Radiación ultravioleta. UV-C. : Ultravioleta C. UV-A. : Ultravioleta A. UV-B. : Ultravioleta B. nm. : nanómetros. nM. : nanomolar. mM. : milimolar vii.
(15) ml. : mililitros. mg. : miligramo. Gy. : Gray (unidad radiación). Gy / h. : Gray por hora. J/m2. : Joule por metro cuadrado. RI. : Radiación ionizante. µm. : micrómetros. µl. : microlitros. µM. : micromolar. TEM. : Microscopia Electronica de Transición. XRF. : Microscopia de fluorescencia de Rayos X. EPR. : Resonancia Electrónica Paramagnética. Mn. : manganeso. GNRPS. : Grupo no Ribosomal Péptido Sintasa. LB. : Luria Bertani. MA. : Agar marino. R2A. : Agar R2A. NTP. : Nota Técnica de Prevención. SDS. : Dodecilsulfato Sódico. CTAB. : Bromuro de hexadeciltrimetilamonio o Bromuro de cetiltrimetilamonio. NaCl. : Cloruro de sodio. PCI. : Fenol, Cloroformo y Alcohol isoamílico. rpm. : Revoluciones por minuto viii.
(16) Abs 280. : Absorbancia a 280 nm de longitud de onda. DNAr 16S. : Ácido desoxirribonucleico ribosomal 16S. OD. : Densidad óptica. HNO3. : Ácido nítrico. H2O2. : Peróxido de hidrógeno. KIO4. : Peryodato de potasio. HCl. : Ácido clorhídrico. pb. : Pares de bases. LD10. : Dosis letal (mata al 90% de la población bacteriana). G+C. : Guanina más citosina. Mn / Fe. : Relación entre el hierro y el manganeso. ix.
(17) RESUMEN. En el presente estudio se evaluó el efecto de la radiación UV-C en la bioacumulación de Mn (II) intracelular, como un bioprotector contra el daño oxidativo de la radiación ultravioleta en bacterias extremófilas, que fueron aisladas del desierto de las Pampas de la Joya. La metodología de la investigación incluye el aislamiento de bacterias en 4 puntos diferentes del Desierto de las Pampas de la Joya, elegidas de acuerdo a sus características macroscópicas. Los aislamientos, la caracterización morfológica y la caracterización molecular, fueron hechos en el Departamento de Biologia - UNSA. Las cepas aisladas, fueron sometidas a una dosis inicial de 300 J/m2 para evaluar su viabilidad y seleccionar a las cepas mas resistentes a radición UV-C, para posteriormente someterlas a dosis fraccionadas de 125, 250, 375, 500, 1000 y 1500 J/m2 de radicion UV-C, y medir su concentración intracelular de Mn (II). Las cepas mas resistentes fueron identificadas molecularmente. Los resultados mostraron dos cepas bacterianas altamente resistentes a radiación UV-C, las cuales según la caracterización morfológica y molecular fueron identificadas como Rhodococcus kroppenstedtii y Methylobacterium radiotolerans, esta ultima cepa soporta hasta una dosis de 1000 J/m2 y logra incrementrar su concentración intracelular de Mn (II) hasta 3,9 mM.. Palabras clave: Extremófilos, Mn (II), radiación UV-C, Desierto de la Joya.. 10.
(18) ABSTRACT. In the present study, the effect of UV-C radiation on the bioaccumulation of intracellular Mn (II) was evaluated, as a bioprotector against the oxidative damage of ultraviolet radiation in extremophilic bacteria, which were isolated from the Pampas Desierto de la Joya. The methodology of the investigation includes the isolation of bacteria in 4 different points of the Desert of the Pampas de la Joya, chosen according to their macroscopic characteristics. The isolations, the morphological characterization and the molecular characterization were made in the Department of Biology - UNSA. The isolated strains were submitted to an initial dose of 300 J/m2 to evaluate their viability and select the strains more resistant to UV-C radiation, to subsequently submit them to fractionated doses of 125, 250, 375, 500, 1000 and 1500 J/m2 UV-C radicion, and measure its intracellular concentration of Mn (II). The most resistant strains were identified molecularly. The results showed two bacterial strains highly resistant to UV-C radiation, which according to the morphological and molecular characterization were identified as Rhodococcus kroppenstedtii and Methylobacterium radiotolerans, the latter strain supports up to a dose of 1000 J/m2 and manages to increase its intracellular concentration of Mn (II) up to 3.9 mM.. Keywords: Extremophiles, Mn (II), UV-C radiation, Desierto de la Joya. 11.
(19) INTRODUCCIÓN. Una problemática grave para los seres vivos es la radiación ultravioleta (RUV), que llega a la superficie terrestre, en su mayor parte es la UV-A (315-400 nm), y pequeñas cantidades de UV-B (280-315 nm) (Björn, 2008), los cuales pueden liberar energía más que suficiente para romper enlaces químicos moleculares (Hall y Giaccia, 2012). Por su parte la radiación UV-C, de forma natural se encuentra presente fuera del planeta Tierra, pero que desde un inicio no siempre fue así, ya que esta radiación era la más predominante en la Tierra primitiva (Paulino-Lima et al., 2016). La capa de ozono es una barrera protectora, que evita el paso de la radiación UV-C, protegiéndonos de sus efectos letales. Sin embargo, durante los últimos años esta capa se está deteriorando. De acuerdo con el informe de la Agencia Espacial Civil de Ecuador (EXA) la concentración de ozono en la franja ecuatorial ha disminuido considerablemente, la densidad normal de la capa en esta zona es de 280 – 300 DU (unidades Dobson) sin embargo en la actualidad la densidad de la capa de ozono ha descendido a menos de 225 DU, por lo cual los niveles de radiación solar en Ecuador, Colombia y Perú, han aumentado de manera alarmante (EXA, 2008). Según McKenzie (2016), el Perú es el país con mayor nivel de radiación UV en el mundo. La ciudad de Arequipa, según SENAMHI, registró un índice de radiación con un nivel de 12.
(20) riesgo extremo. Sin duda, es un problema muy grave, ya que la radiación de tipo UV-B y UV-C (200-280 nm) es absorbido directamente por el ADN, dando lugar a una alteración física en su estructura molecular y ocasionando cáncer (Paulino-Lima et al., 2016). Anualmente en el mundo se producen entre dos y tres millones de casos de cáncer de piel no melánico y aproximadamente 132 000 casos de cáncer de piel melánico (OMS, 2003), debido a la fuerte absorción de longitudes de onda inferiores a 320 nm por parte del ADN (McKenzie, 2016.). La exposición de los sistemas biológicos a la radiación genera estrés oxidativo y daño celular, lo cual exige a los seres vivos desarrollan mecanismos de protección (Görner H., 1994; Slade y Radman, 2011). Algunos microorganismos utilizan estrategias como la formación de esporas, la síntesis de pigmentos absorbentes de UV o la acumulación de ciertos compuestos intracelulares para hacer frente a estos daños (Rettberg et al., 2003). Los desiertos, que son ecosistemas que presentan una intensa exposición a radiación ultravioleta (UV), cambios de temperatura y una alta variabilidad bioquímica y geoquímica, representan un escenario interesante para el estudio de la diversidad de microrganismos adaptados a estos ambientes extremos, en donde la vida resulta ser casi imposible. La resistencia a la radiación en microorganismos también se ha correlacionado con la resistencia a la desecación (Mattimore y Battista, 1996), siendo esta última una característica típica de los desiertos (Drees et al., 2006), que generalmente son regiones áridas y semiáridas, que cubren más de un tercio de la Tierra (Cable et al., 2008). Es por ello que en los últimos años los estudios en estos ecosistemas se han incrementado, encontrándose diversas comunidades microbianas de extremófilos, como en el desierto de Sonora, donde se encontraron bacterias, cianobacterias, proteobacterias, actinobacteria, acidobacteria, hasta algunas Archaea. En el desierto de Atacama se han encontrado diferentes géneros: Arthrobacter, Asticcacaulis, Bacillus, Bradyrhizobium, Brevibacillus, Cellulomonas,. Chroococcidiopsis,. Cyanidium,. Deinococcus,. Geodermatophilus,. Gloecapsopsis, Hymenobacter, Kocuria, Mesorhizobium y Sphingomonas (Azua-Bustos et al., 2014(1); Azua-Bustos et al., 2014(2)). Las Pampas de la Joya, es uno de los desiertos más antiguas de la Tierra, con condiciones extremas que ha perdurado entre 15 y 25 millones de años (Dunai et al., 2005; Evenstar et al., 2005), convirtiéndolo en un lugar propicio para el aislamiento de microrganismos resistentes a la radiación alta de UV-C. Los microorganismos han sobrevivido desde la formación de la tierra, expuestos a una variación constante de radiación UV, gracias a sus características morfológicas, genéticas 13.
(21) y fisiológicas (Culotta y Daly, 2013). La presencia de enzimas reparadoras como las fotoliasas, ha sido utilizado como una maquinaria para la reparación del ADN dañado después de la exposición a la radiación (Krisko y Radman, 2013). Otra respuesta ha sido la presencia de complejos catiónicos como el Mn (II), que son metabolitos intracelulares que pueden proporcionar protección celular frente al aumento de la concentración de radicales libres debido a una sobrexposición a radiación (Culotta y Daly, 2013) Aunque altas dosis de UV son perjudiciales para la vida, algunos microorganismos, llamados extremófilos, se han adaptado a estas condiciones y son capaces de sobrevivir expuestos a altas cantidades de radiación UV. La selección de bacterias resistentes a la radiación UV-C, podría ayudar a entender la defensa y reparación molecular. En este estudio se pretende aislar bacterias que resistan radiación UV-C y que sean capaces de acumular Mn (II) intracelular, para comprender el papel fundamental de este catión en la recuperación de células expuestas a la radiación, como una primera línea de defensa contra este daño.. 14.
(22) OBJETIVOS. OBJETIVO GENERAL. Evaluar el efecto de la radición UV-C, en la bioacumulación de Mn (II) en bacterias aisladas de las Pampas de la Joya – Arequipa.. OBJETIVO ESPECÍFICOS. Aislar cepas nativas del Desierto de las Pampas de la Joya, caracterizar morfológica y bioquímicamente las cepas nativas aisladas. Determinar la viabilidad bacteriana frente a la radiación UV-C. Identificar molecularmente las bacterias más viables a la radición UV-C. Evaluar la concentración intracelular de Mn (II) como bioprotector intracelular contra la radiación alta UV-C.. 15.
(23) CAPÍTULO I. MARCO TEÓRICO. 1.1. EXTREMÓFILOS Y AMBIENTES EXTREMOS. Los extremófilos son organismos que están adaptados para crecer de manera óptima en los rangos extremos de la vida, prosperan en condiciones que, desde un punto de vista humano, son claramente hostiles (Horikoshi et al., 2010). MacElroy (1974) fue el primero en acuñar el término ''extremófilos'' en un artículo titulado “Some comments on the evolution of extremophiles”, pero definiciones como extremo y extremófilos son, por supuesto antropocéntricas. Existe una diversidad mucho mayor de organismos conocidos que pueden tolerar condiciones extremas y crecer, pero no necesariamente de manera óptima en hábitats extremos; estos organismos se les define como extremotrofos (Mueller et al., 2005). Un extremófilo es un organismo que se desarrolla en condiciones extremas. El término extremófilo se utiliza con más frecuencia para referirse a organismos que son unicelulares y procariotas, debido a que muchos extremófilos son miembros del dominio arquea; sin embargo, esto es una engañosa concepción, debido a que muchos organismos pertenecientes a las bacterias y eucarias no son extremófilos o solo 16.
(24) presentan ciertas etapas en algunos ciclos de vida extremotróficas. Además, no todos los extremófilos son unicelulares (Kelley et al., 2005).. 1.2. CATEGORÍAS DE EXTREMÓFILOS Y EXTREMOTROFOS. Los extremófilos se desarrollan bajo condiciones que podrían matar a la mayoría de otras criaturas y muchos no podrían sobrevivir en los ambientes considerados globalmente normales. Los ambientes extremos incluyen aquellos con temperaturas muy elevadas (55-121 °C) o temperaturas bajas (de -2 a -20 °C), alta salinidad (NaCl2 - 5M) y alta alcalinidad (pH arriba de 8) o alta acidez (pH menor de 4) (NavarroGonzales et al., 2003). La literatura contiene varios términos que se utilizan para describir a los extremófilos y extremotrofos, y existen sub-definiciones para los organismos que presentan moderada, extremo, hiper-extremo, y/o extremófilos obligados (Horikoshi et al., 2010): . Acidófilo: Un organismo con un pH óptimo para el crecimiento en, o por debajo de 3-4.. . Alcalófilo: Un organismo con un crecimiento óptimo a valores de pH por encima de 10.. . Endólito: Un organismo que vive en el interior de las rocas.. . Halófilos: Un organismo que requiere al menos 1 M de sal para el crecimiento.. . Hipertermófilo: Un organismo que tiene un crecimiento óptimo desde los 80 °C a más altas temperaturas.. . Hipólito: Un organismo que vive en el interior de las rocas en desiertos fríos.. . Metalotolerantes: Organismos capaces de tolerar altos niveles de metales pesados, tales como cobre, cadmio, arsénico y zinc.. . Oligotróficas: Un organismo capaz de crecer en hábitats con niveles muy bajos de nutrientes.. . Barófilo: Un organismo que vive de manera óptima a presiones hidrostáticas de 40 MPa o superior.. . Psicrófilo: Un organismo que tiene una temperatura óptima de crecimiento de 10 ºC o inferior, y una temperatura máxima de 20 °C. 17.
(25) . Radioresistentes: Organismos resistentes a altos niveles de radiación ionizante.. . Termófilos: Organismos que pueden prosperar a temperaturas entre 60 °C y 85 °C.. . Toxitolerantes: organismos capaces de soportar altos niveles de agentes dañinos, tales como disolventes orgánicos.. . Xérofilo: un organismo de crecimiento con poca presencia de agua y resistente a la alta desecación.. Los fenómenos de poliextremofilos y poliextremotrofos, se refiere a organismos adaptados a más de dos condiciones extremas, han recibido relativamente poco estudio detallado (Horikoshi et al., 2010).. 1.3. DISTRIBUCIÓN DE EXTREMÓFILOS Y EXTREMOTROFOS Los ambientes extremos en la Tierra han surgido y seguirán surgiendo como una consecuencia de la actividad tectónica de placas, la naturaleza dinámica de la criósfera, y la formación de cuencas endorreícas y otros fenómenos geotérmicos tales como sistemas de respiraderos hidrotérmicos marinos. Estos ambientes últimos mencionados, están distribuidos a nivel mundial y constan de fumarolas negras y chimeneas de carbonato. Tales manifestaciones tectónicas producen diversamente calor extremo, pH, gases disueltos y metales (Kelley et al., 2005). Los ambientes extremos han evolucionado como resultado de los procesos naturales en escalas de tiempo geológicas (Navarro-Gonzales et al., 2003). Sin embargo, en tiempos más recientes, las condiciones extremas se han impuesto por causa del ser humano, por ejemplo, la salinidad del suelo se incrementó como resultado de perforaciones de pozos para agua de riego, la contaminación radiactiva por plantas de energía nuclear, la contaminación industrial y el uso de agroquímicos por los químicos xenobióticos. Así la totalidad de estos ambientes 'extremos' globales y locales, naturales y antropogénicos tiene proporcionado un notable abanico de oportunidades para la evolución de los organismos (Kelley et al., 2005).. 18.
(26) 1.4. DIVERSIDAD MICROBIANA DE AMBIENTES EXTREMOS En general, la diversidad filogenética de los extremófilos es alta y muy compleja de estudiar. Algunos órdenes o géneros contienen solo extremófilos, mientras que otras órdenes o géneros contienen tanto extremófilos como no extremófilos. Curiosamente, los extremófilos adaptados a la misma condición extrema pueden estar ampliamente dispersos en el árbol filogenético de la vida. Este es el caso de diferentes psicrófilos o barófilos, cuyos miembros se pueden encontrar dispersos en los tres dominios de la vida. También hay grupos de organismos pertenecientes a la misma familia filogenética que se han adaptado a condiciones extremas o moderadamente extremas (Horikoshi et al., 2010). El descubrimiento de microorganismos que habitan en ambientes con temperaturas extremas, pH extremos, altas presiones barométricas y alta salinidad, ha despertado el interés desde el punto de vista biotecnológico debido a las características de estos microorganismos ya que sus biomoléculas son necesariamente resistentes a las condiciones agresivas de su entorno. Esto desemboca en intensos trabajos para intentar comprender los mecanismos íntimos de resistencia, pero también para estudiarlos en la perspectiva del desarrollo de aplicaciones industriales entre las que la PCR (Polymerase Chain Reaction) constituye el más destacado ejemplo. Además, la presencia de microorganismos en ambientes extremos hace especular que la vida podría existir en algunos de los ambientes extremos encontrados en el espacio extraterrestre (Schmidt et al., 2009). La mayoría de las enzimas usadas por la industria hasta la fecha se originan de organismos mesófilos y, a pesar de sus muchas ventajas, el uso de estas enzimas está restringido debido a su estabilidad limitada en los extremos de la temperatura, del pH y de la fuerza iónica. Por otra parte, los extremófilos son una fuente potente de enzimas, que demuestran alta estabilidad bajo condiciones extremas (Navarro-González et al., 2003). Los extremófilos han despertado el interés de varias industrias, debido a sus enzimas, catalizadores biológicos que aceleran las reacciones químicas de la célula, como en el caso de las arqueas extremófilas les permiten colonizar los ambientes más inhóspitos. Las enzimas habituales tienen ciertas limitaciones, sin embargo las extremoenzimas, empiezan a operar justo en el punto donde las habituales dejan de funcionar (Schmidt et al., 2009). Existen numerosos procesos industriales que requieren el uso de estos 19.
(27) aceleradores químicos, tal es el caso de las enzimas que participan en la producción de edulcorantes, papel, síntesis de detergentes, elaboración de alimentos como pan y vino, tratamiento de residuos, extracción de petróleo, obtención de biochips para la identificación de personas y el diagnóstico de enfermedades (Battistuzzi et al., 2008).. 1.5. EL DESIERTO DE LAS PLAMPAS DE LA JOYA. La región hiperárida del desierto de Atacama se extiende desde el norte de Chile hasta el sur de Perú a lo largo de una franja de casi 1200 km y hasta 180 km de anchura. Se considera uno de los desiertos subtropicales mas antigua de la tierra, donde se establecieron condiciones hiperáridas desde el Mioceno. Por otro lado, las Pampas de La Joya, que son parte del desierto de Atacama, presentan una altitud media de 1200 metros sobre el nivel del mar, representa la extensión más septentrional de Perú del desierto de Atacama, aunque no se ha establecido el calendario de sus condiciones muy secas (Newman y Banfield, 2002). Las Pampas de La Joya, se encuentran entre los 15° y 17° de latitud sur, aproximadamente 1000 kilómetros al norte del desierto de Yungay (Chile). Recientemente, se ha informado que esta región de la Joya, presenta suelos con algunas propiedades que parecen estar presentes en los suelos de Marte, como los niveles muy bajos de materia orgánica, una matriz altamente oxidante, y la presencia de minerales ricos en hierro y sales evaporíticos (Valdivia-Silva et al., 2009, ValdiviaSilva y Ortega, 2011). Geológicamente, las Pampas de La Joya puede ser definido como una amplia unidad geomorfológica que se caracteriza por una llanura levantada principalmente por granitos precámbricos limitados en el noreste y suroeste por las estribaciones de los Andes y la Cordillera de la Costa, respectivamente, con una elevación promedio de 1200 metros de altitud, y que se divide en varios sectores por los ríos Sihuas y Vitor. Curiosamente, estas zonas parecen ser más jóvenes en comparación con Yungay (Chile), debido a que las Pampas de La Joya no muestran grandes cantidades de sales exóticas (Michalski, et al., 2004). Una de las razones por las que este desierto presenta condiciones hiperáridas, es debido a la presencia de la Cordillera de la Costa, actuando como una barrera topográfica que condensa la mayor humedad en el lado de sotavento del viento que viene, desde el océano Pacífico. Estos factores físicos sugieren un 20.
(28) nuevo escenario con el fin de evaluar las condiciones ambientales en el comienzo de la formación de depósitos en los suelos hiperáridos (Valdivia-Silva et al., 2009). Es importante destacar que los datos climatológicos, demuestran características muy duras, en donde el desarrollo de la vida se hace casi imposible en esta zona, lo que lo llevo a ser catalogado como un análogo de Marte (McKay et al.2003). La meteorología y climatología que el desierto de las Pampas de la Joya puede presentar es: 1.5.1. Temperatura: El régimen de temperatura en las Pampas de La Joya es templado, llegando a una máxima de 39,9°C y una mínima de -4.5°C. La temperatura media del medio ambiente es de 18,9°C, lo que demuestra que esta región es de aproximadamente 2,5°C más caliente en comparación con la media obtenida para la región de Yungay (Chile), que era 16,5°C de temperatura (McKay et al., 2003). La temperatura media de las rocas tomadas durante los años de trabajo (2004 - 2008) de Valdivia-Silva et al. (2012) fue de 25,2°C (6 ° C más que el medio ambiente), que es significativamente diferente de la media de la temperatura del aire (Mann-Whitney U-test, p <0,05). Los meses identificados como los más fríos en las Pampas de La Joya fueron mayo, junio y julio, que coincide con la temporada de invierno y la entrada de la niebla, siendo de los tres el mes de junio el de menor temperatura (Valdivia-Silva et al., 2012). 1.5.2. Humedad: Los datos obtenidos sobre la humedad durante los años 2004 al 2008, fue escasa, durante este periodo de evaluación (4 años) se obtuvieron valores inferiores a uno, excepto para septiembre y octubre de 2006, donde se produjeron precipitaciones fuertes relacionadas con el fenómeno de El Niño - Oscilación Sur (ENOS). Hubo datos positivos, especialmente durante las noches de los meses de junio, julio y agosto, que coinciden con la temperatura fría y la presencia de densa niebla nocturna. También es interesante observar que las precipitaciones inferiores a 1 mm no generaron una respuesta en el sensor de rocío superior a '1'. Frente a la ausencia de precipitaciones o con precipitaciones inferiores a 1,1 mm, la humedad del suelo se mantuvo cercana al 30%, por lo 21.
(29) cual podemos decir que los suelos son extremadamente áridos (McKay et al.2003; Davis et al., 2010). Friedmann y Ocampo-Friedmann (1977) sugirieron que la principal fuente de agua para los microorganismos hipolíticos como las cianobacterias, en un medio ambiente hiperárido, como lo es el desierto del Negev es la humedad de condensación. Por las observaciones hechas en las Pampas de La Joya, la humedad de condensación no parecen ser la principal fuente de humedad, al igual que en el caso de Yungay (McKay et al., 2003). Sin embargo, es bien sabido que las moléculas de agua son atrapadas en las grietas y poros de cierto tipo de rocas, sales y minerales, confiriéndoles una humedad relativa “a largo plazo” en el subsuelo, pudiendo de esta manera permitir la presencia de microorganismos (Wierzchos et al., 2006; Davila et al., 2008). Por otra parte, una evaluación preliminar de la temperatura de las rocas en el desierto mostró que el cuarzo tenía características de amortiguación de temperatura, incluso en las horas en que las rocas tenían temperaturas superiores a 50 °C, lo que permitió un mayor tiempo de condensación de agua y evitó una evaporación más rápida (Valdivia-Silva et al., 2012). En los minerales de cuarzo colonizados, los niveles de radiación para la fotosíntesis alcanzaron valores entre 3,2 y 0,4% debajo de las rocas, lo que corrobora que este tipo de rocas son excelentes. hábitats. para. los. microorganismos. autótrofos. como. las. cianobacterias. Teniendo en cuenta que sólo 1 o 2 de cada 100 rocas de cuarzos son colonizadas, los factores que modifican este hecho todavía siguen siendo poco conocidos, pero podrían explicarse por los cambios de humedad relativa y microclimas (Davila et al., 2008). 1.5.3. La radiación solar y dirección del viento: La radiación solar anual media registrada en las Pampas de la Joya fue de 408 Wm-2, con un máximo de 1060 Wm-2 al mediodía en los días más secos entre los meses de septiembre y abril, y ligeramente más bajos durante los días más húmedos entre mayo y agosto. La radiación en la superficie es alta y calienta las rocas hasta 20 °C más que el aire, lo que se confirmó por la coincidencia de la temperatura máxima de la roca con la radiación solar máxima. Es importante destacar que el promedio anual de la radiación solar en las Pampas de La Joya 22.
(30) fue es mayor que el valor anual que se encuentra en Yungay (336 Wm-2) (Mckay et al. 2003; Valdivia-Silva et al., 2012). La dirección del viento para estos vientos cálidos y secos fue desde el noroeste hacia el sureste con oscilación entre 180° y 260°. Las velocidades del viento eran entre 5 m/s y 20 m/s, y principalmente por la mañana y por la tarde. Los vientos pueden llegar a ser un importante mecanismo de transporte de sales y minerales, y tenderían a acumular material en los lados este y sureste de las llanuras del desierto (Valdivia-Silva et al., 2012).. 1.6. RADIACIÓN ULTRAVIOLETA (UVR). La radiación es energía en forma de ondas electromagnéticas (por ejemplo, rayos gamma, rayos X, radiación UV, ondas de radio, etc.) que causa el daño oxidativo a las biomoléculas vitales, tales como proteínas, ADN, ARN, y enzimas. La luz solar es responsable de causar cambios en la estructura molecular del ADN mediante la formación de dímeros entre las hebras de moléculas de ADN. Por esta razón la radiación UV se ha relacionado con muchos efectos nocivos en los seres humanos, incluyendo la supresión inmune, dermatitis, envejecimiento prematuro y, en casos extremos, cáncer de piel (Agar et al., 2004). 1.6.1. La luz: La luz es radiación electromagnética (energía radiante) que se desplaza en forma de longitudes de onda. Esta energía viaja en trayectorias en línea recta y en todas las direcciones desde la fuente emisora, que por lo general es una lámpara. El espectro electromagnético se considera continuo; parte del espectro es la anchura de banda estrecha de longitudes de onda, que son visibles para el ojo humano. Estas longitudes de onda se combinan en grupos conocidos como rayos infrarrojos, rayos ultravioletas, rayos X, rayos gamma, y rayos cósmicos (Lopez-Malo y Palou, 2005). La luz puede ser considerada como una corriente de energía. La cantidad de energía transportada por una corriente de luz es un factor de su frecuencia y longitud de onda. Cuando la luz de un nivel de energía es absorbida por la materia, se produce un cambio químico o físico. Los átomos y las moléculas, 23.
(31) sin embargo, sólo absorben las longitudes de onda que proporcionan la cantidad correcta de energía para cambiar su estado (Fig. 1). Típicamente, la longitud de onda de la luz varía de 100 a 400 nm (Giese, 1992). Que a su vez se subdivide en UV onda larga (UVA) entre 315 y 400 nm, UV de onda media (UVB) 280315 nm, y UV de onda corta (UVC) entre 200 y 280 nm (Bolton, 2001).. Fig. 1. Esquema del origen de los diferentes tipos de radiación y sus efectos sobre extremófilos. (Fuente: Lopez-Malo y Palou, 2005). 1.6.2. Radiación UV-C La luz ultravioleta C (UVC) es de onda corta y se conoce por su poder germicida, porque la mayoría de los microorganismos absorben la luz ultravioleta a una longitud de onda de 254 nm, que es suficiente para causar un desplazamiento físico de los electrones y ruptura de los enlaces en el ácido desoxirribonucleico (ADN) (Lopez-Malo y Palou, 2005). La alta absorción de UVC (254 nm) de ADN se asocia con la capacidad de las bases de pirimidina y purina para absorber la luz en esta longitud de onda, induciendo la formación de dímeros unidos covalentemente entre bases adyacentes (Lopez-Malo y Palou, 2005; Adams y Moss, 1995) (Fig. 2).. 24.
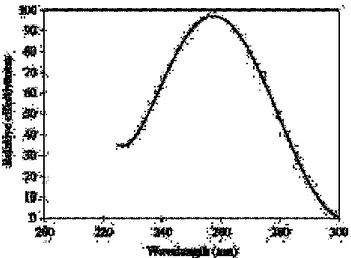
(32) Fig. 2. Dimerización fotoquímica de timina inducida por UV (Lopez-Malo y Palou, 2005). De las cuatro bases constituyentes de ADN, la adenina, citosina, guanina, y timina, el último se somete a una reacción fotoquímica (Figura 2). Si dos timinas base se encuentran adyacentes entre sí, la absorción de un fotón de UV por una de las bases forma un enlace químico que resulta en un dímero (Bolton, 2001). Estos dímeros de timina inhiben la correcta replicación del ADN durante la reproducción de la célula. El mecanismo fundamental de la desinfección con UV, como un método físico de control microbiano, es la dimerización, la fotoquímica de pares de timina, ya que si se forman suficientes pares, el ADN puede ser no replicado (Bolton, 2001). La resistencia de los microorganismos a los tratamientos de radiación UVC, está determinada en gran medida por su capacidad para reparar el daño del ADN (Sastry et al., 2000). Asi mismo se informa, que el pico de absorción de radicion de los cromosomas esta dentro de un rango de 260-265 nm de longitud de onda, que corresponde muy estrechamente a la emisión de longitud de onda de la radiación UVC (Fig. 3), es por ello que las lámparas de baja presión de mercurio (253,7 nm) son utilizados para la inactivacion microbiana (Bolton, 2001). La resistencia de los microorganismos a los tratamientos UVC se determina en gran parte por su capacidad para reparar el daño del ADN causado por la radiación UV. En general, la resistencia a la irradiación UV sigue el patrón Gram-negativos <Gram-positivos <levadura <esporas bacterianas <moldes<virus (Lopez-Malo y Palou, 2005). 25.
(33) Fig. 3. Poder germicida de las diferentes longitudes de onda. (Fuente: Bolton, 2001). 1.6.3. Inactivación cinética La inactivación microbiana por métodos físicos tales como el tratamiento por radiación UVC se suele representar como una curva de supervivencia o una curva de dosis-respuesta. Para los tratamientos de radiación UV, la curva de dosis-respuesta presenta la fracción de la población microbiana original que sobrevive como una función de la dosis UV. Shama (1999) indica que las curvas de supervivencia en los tratamientos UV podrían seguir una cinética de primer orden de inactivación (Fig. 6). La mayoría de las curvas de supervivencia, sin embargo, incluyen una meseta inicial correspondiente a las dosis bajas de UV; de este modo la forma de la curva para la inactivación microbiana por luz UV es sigmoide (Sastry et al., 2000). La meseta inicial es debida a una fase de lesión del microorganismo en respuesta a la exposición UV. Después de la meseta inicial, la cantidad máxima de lesión ha sido superada; por lo tanto, mínima exposición UV adicional sería letal para los microorganismos, y los números de sobrevivientes pueden declinar rápidamente. El final de la curva puede presentar una fase de tailing debido a la resistencia de UV de los microorganismos (Fig. 4). Los estudios relativos a la exposición de radiación UV requerida para obtener una reducción de varios microorganismos usando luz UV de 254 nm, sugieren que las reducciones de registro están relacionadas con la exposición radiación UV (Lopez-Malo y Palou, 2005).. 26.
(34) Fig. 4. Las curvas de supervivencia durante los tratamientos típicos UVC. (Fuente: Shama, 1999). 1.6.4. Daño celular por la Radiación Ultravioleta Los modelos clásicos de la toxicidad de la radiación se basan en la suposición tácita de que la radiación produce daños de manera indiscriminada a macromoléculas celulares, como las proteínas individuales en una célula o por el daño del ADN (Daly, 2009). Se ha propuesto recientemente que las bacterias naturalmente sensibles mueren por la radiación, debido principalmente a la oxidación de proteínas, por lo que los microrganismos resistentes presentan complejos de manganeso que protegen a las enzimas de reparación de ADN y permiten la supervivencia de las células bacterianas (Daly, 2009; Daly et al., 2010). Dale (1940), fue el primero en argumentar que la radiación daña a las proteínas, basándose en los hallazgos de enzimas en solución acuosa, que fueron onactivadas por pequeñas dosis de rayos X (menos de 50 Gy). La posibilidad de que la resistencia a la radiación pueda aumentar, fue apoyada por estudios que mostraron que la radiosensibilidad de una enzima no es una entidad fija, sino una variable, donde la inactivación se podría evitar mediante la adición de nucleótidos, azúcares, aminoácidos, y una variedad de otros compuestos orgánicos. La radiobiología estableció que la principal vía de lesión causada por radiación, era por especies moleculares reactivas derivadas de la ionización del agua, el cual genera radicales hidroxilo (HO●), superóxido (O2) y peróxido de hidrógeno (H2O2) como productos principales de ionización (von Sonntag, 1987). 27.
(35) En la década de 1930, la acción letal de la radiación solar había sido atribuido principalmente a los daños directos (Scott, 1937). La idea de Dale, sobre el daño de las proteínas producto de la toxicidad celular, fue suplantada en 1960 por el tema de la radiobiología dogmática ''la muerte por daño del ADN''. Se ha venido investigando sobre los modelos de toxicidad por radiación ya casi 5 décadas de investigación del género Deinococcus, y como resultado, numerosas investigadores llegaron a la conclusión de que el daño al ADN y los lípidos es un proceso secundario, y que las proteínas son objetivos iniciales más probables de daño de la radiación celular y deben colocarse a la parte superior en la jerarquía de las lesiones inducidas por la radiación responsable de la muerte celular (Daly et al., 2007; Krisko y Radman, 2010). Ciertamente, el ADN es un objetivo fundamental en todas las células irradiadas (Scott, 1937), pero su supervivencia depende en última instancia de la capacidad de las proteínas para reparar el daño (Daly et al., 2010). Se demostró, ya desde 1973 (Blok y Loman, 1973), mediante las curvas de supervivencia de bacterias sensibles a radiación, que solo era necesario unos pocos eventos para producir su inactivación, ya que la supervivencia de las células depende en última instancia de la reparación de su genoma (Daly et al., 2007; Daly, 2009).. 1.7. MICROORGANISMOS RADIACIÓN. EXTREMÓFILOS. RESISTENTES. A. LA. Diferentes microorganismos han desarrollado formas de sobrevivir bajo niveles altos de radiación. Dentro de la especie más estudiada destaca Deinococcus radiodurans que es capaz de resistir los efectos supraletales de la radiación ionizante (RI) y radiación ultravioleta (RUV) (> 1.000 J/m2). Las cianobacterias endolíticas son reportadas con una gran capacidad de protegerse de los efectos nocivos de la radiación UV (Rastogi et al., 2010), así como otros microorganismos, como Rhodanobacter sp. y Desulfuromonas ferrireducens que pueden sobrevivir en presencia de altos niveles de radionucleidos (Green et al.2012). La capacidad de los organismos para sobrevivir a altos niveles de radiación se ha relacionado con sus mecanismos de reparación eficiente del ADN y su capacidad de producir productos metabólicos primarios y 28.
(36) secundarios (Singh y Gabani, 2011). Los metabolitos, pigmentos, y enzimas que producen estos microorganismos pueden ser inducidas o activados por técnicas biotecnológicas, para producir fármacos, medicamentos contra el cáncer, así como antibióticos y productos agrícolas de importancia comercial (Kumar et al., 2010). Los microorganismos que prosperan bajo radiación extrema se denominan extermófilos radioresistentes (Gabani et al. 2012). Ellos se han encontrado en nichos ambientales amplios tales como elevaciones altas (sierras) y campos abiertos donde los niveles de radiación UV son altos. El agotamiento continuo de la capa de ozono ha influido en gran medida la cantidad de radiación UV en la biosfera de la Tierra. Además, el amplio uso de elementos radiactivos y compuestos para producir energía, en la medicina, la investigación, y la industria ha producido desechos radiactivos en el medio ambiente. Los accidentes nucleares como el desastre nuclear de Fukushima Daiichi en 2011 y la catástrofe de Chernobyl en 1986 también han provocado un aumento de los radionucleidos y los radioisótopos en el medio ambiente. Otras formas de radiación encontrados en el medio ambiente incluyen radiación gamma y rayos X, que se sabe que son perjudiciales para los seres humanos. (Pryakhin et al., 2012) Por el momento, el grupo más resistente a la radiación de los organismos descubiertos hasta ahora pertenece a la familia bacteriana Deinococcaceae (Fig. 5), que muestra típicamente cerca del 100% de supervivencia luego de los efectos agudos de exposiciones a 12.000 Gy o 1,000 J/m2 (254 nm) ultravioleta (UV) y puede crecer en condiciones extremadamente duras de radiación (60 Gy / h) (Daly 2000). El miembro más caracterizado de este grupo de extremófilos es Deinococcus.. 29.
(37) Fig. 5. Micrografía electrónica de transmisión de D. radiodurans La escala es de 0,5 µm. (Fuente: Horikoshi et al., 2011).. 1.8. MECANISMOS DE DEFENSA CONTRA LA RADIACIÓN ULTRAVIOLETA. 1.8.1. Proteínas de reparación del DNA dañado Es de gran interés entender los cambios en la genómica, proteómica y perfiles metabólicos (metabolómica) de los organismos resistentes a la radiación para comprender cómo sobreviven (Singh, 2006; Singh et al., 2011). La inducción del gen uvrA en D. radiodurans revela la expresión de proteínas, tales como UvrABC, cuyas funciones son la reparación del ADN y la supervivencia del organismo en presencia de radiación UV. Además, se identificaron proteínas adicionales como recA, recD, recF, recG, recO de, mutS, mutL, RuvB, etc. (Singh y Gabani, 2011). Gabani et al. (2012) revelaron que los microorganismos, Cellulosimicrobium cellulans (UVP1) y Bacillus pumilus (UVP4), cultivados bajo radiación, sufrían una sobre-expresión diferencial de muchas proteínas y metabolitos que no se han identificado aún. En Bacillus sp. HKG 112 gamma resistente, se encontraron dos proteínas, la flagelina de 38 kDa y la proteína S-layer de 86,5 kDa, quienes tuvieron una sobre expresión significativa después de la exposición a la radiación (Gupta et al., 2011).. 30.
(38) Además, el estudio de las proteínas frente a la radiación, uno de los eventos más comunes es el aumento de los niveles de expresión por inducción de estrés a dicha radiación, como la proteina DR1199 en D. radiodurans que pueden estar implicados en la desintoxicación de la célula de ROS (Leulliot et al., 2009), en Chlamydomonas sp. ICE-L hubo un aumento de los niveles de la proteína Hsp70 de hasta tres veces en presencia de la radiación UVB (Liu et al., 2010). Otro análisis comparativo de la proteómica de D. radiodurans y D. deserti reveló que la proteína HU fue la más abundante, y que esta sería una histona de unión del ADN, confiriéndole cierta resistencia en las rupturas del ADN (de Groot et al., 2005). Liedert et al. (2010) informaron que en D. geothermalis, había 34 proteínas abundantes, que hasta ese entonces no tenían ninguna función conocida, y quizás podrían relacionarse con la tolerancia al estrés causada por radiación del organismo. La presencia de las enzimas de reparación del ADN altamente eficientes en D. radiodurans, permiten reparar cientos de rupturas en el ADN. Una de las enzimas más importantes, vendría a ser la ADN Polimerasa de la familia X, cuya importancia fue demostrada luego de eliminar la enzima por mutación, observándose que hubo una disminución en la tasa de reparación de ruptura de ADN bicatenario y también un aumento en la sensibilidad a radiación luego de eliminar esta enzima por mutación (Leulliot et al., 2009). Así mismo, existen microorganismos con rutas o vías metabólicas que intervienen en la reparación del daño (Nath y Bharathi, 2011), como Dictyostelium discoideum, que es capaz de sobrevivir a dosis extremadamente altas de radiación debido a la presencia de la vía Fanconi Anemia (FA). Zhang et al. (2009) revelaron que en D. discoideum la interrupción de la nucleasa Xpf resultó ser para la célula, una razón de hipersensibilidad extrema a la radiación. También se reveló que la nucleasa Xpf funcionaba con la vía FA. Otro estudio encontró que la proteína Dclre1 era responsable de la reparación de ruptura de las hebras de ADN (Hsu et al.2011). Por otro lado, Muller-Taubenberger et al. (2011) informaron que la presencia de la histona Dot1 en el ADN contribuiría a la resistencia de rayos UV en D. discoidium.. 31.
(39) 1.8.2. Multiplicidad, alineación y estructura del genoma Hasta la década de 1970, las bacterias se las consideraban generalmente como organismos haploides con sólo una copia de su genoma en células en reposo. Esta posibilidad podría haber explicado la gran sensibilidad a la radiación de la mayoría de las bacterias y su ruptura de doble cadena del inglés “Double Strand Dreak” (DSB), que en un genoma bacteriano habría sido letal. Sin embargo, estudios posteriores revelaron que las bacterias resistentes y sensibles son multigenómicas. Por ejemplo, D. radiodurans y E. coli tienen cuatro a ocho genomas haploides por célula durante su crecimiento logarítmico (Hansen, 1978; Akerlund et al., 1995), pero sólo D. radiodurans es resistente. Otros ejemplos notables son Micrococcus luteus, Micrococcus sodonensis y Azotobacter vinelandii, que contienen al menos diez genomas haploides por célula, pero son sensibles a radiación (Minton, 1996). En un modelo de reparación temprana, la alineación de D. radiodurans con cuatro a ocho genomas por célula se tomó como punto de partida para la reparación de DSB (Minton y Daly, 1995). Este modelo reveló dos predicciones principales: en primer lugar, la recombinación dependiente de fragmentos homólogos procedentes de ubicaciones genómicas ampliamente separadas debía de mostrar un fuerte efecto sobre la irradiación. En segundo lugar, la Microscopía Electrónica de Transmisión (TEM) del ADN cromosómica en D. radiodurans reveló la existencia de estructuras que unen a los cromosomas. Ambas predicciones fueron probadas y los resultados moleculares mostraron altos. niveles. de. recombinación. entre. fragmentos. homólogos. independientementes de su origen genómico, y que no presentan estructuras de conexión basado en TEM (Daly y Minton, 1997). Otro modelo propuso que los altos niveles de condensación cromosómica facilitaban la reparación mediante la proximidad entre DSB, y que el manganeso promueve la condensación de sus nucleótidos. Este modelo también se descartó, ya que no mostró nucleoides condensados, sin embargo, las células permanecieron extremadamente resistentes, y que células crecidas en medios empobrecidos de manganeso mostraron nucleoides condensados y células sensibles a radiación (Daly et al., 2004). Por lo tanto, la forma estructural de sus nucleoides no juega un papel decisivo en la radioresistencia. 32.
(40) Además, debemos mencionar, que D. radiodurans contiene numerosas, pero pequeñas repeticiones nucleares (SNRs) y cuadruplos de secuencias (Makarova et al., 2001); ambos tipos de secuencias podrían contribuir potencialmente a la estructura del genoma, sin embargo, estas secuencias no fueron identificados en los genomas de D. geothermalis o D. deserti (Makarova y Daly, 2010). En resumen, hay características raras basadas en la estructura, la secuencia o multiplicidad del genoma y aun no se puede establecer una base molecular inequívoca de radioresistencia.. 1.9. IDENTIFICACIÓN DE MICROORGANISMOS. 1.9.1. Identificación morfológica: La identificación microbiana puede realizarce a través del estudio de características morfológicas, respuestas a tinción y resultados de pruebas bioquímicas aplicadas a los microorganimos (Gamazo et al., 2005) Las bacterias sembradas en un medio sólido, al dividirse, se mantienen relativamente fijas en su posición formando colonias, pudiendo variar entre una y otra en algunos aspectos; como en tamaño, forma, superficie, consitencia, pigmentación, y olor, que son consideradas como las características mas relevantes, por lo que, el estudio de la morfología de las colonias es obligatorio para iniciar la identificación de las bacterias (Bergey et al., 2012) Para la identificación morfológica y taxonómica de las bacterias se deben tomar en cuenta el tamaño, forma y determinados detalles que caracterizan a los principales grupos de bacteria, aportando bases escenciales para su estudio sistemático e identificación (Vargas-Flores y Kuno-Vargas, 2015).. 1.9.2. Identificación molecular: 1.9.2.1. Características del ARNr 16S El DNAr 16s es un polirribonuleótido codificado por el gen rrs también denominado RNA ribosomal 16s. A partir de esta secuencia genética se puede 33.
(41) obtener información filogenética y taxonómica. En los eucariotas es el DNAr 18s, una macromólecula equivalente, dado que los DNAr 16s y DNAr 18s proceden de las subunidades pequeñas de los ribosomas (Rodicio y Mendoza, 2004). La comparación de las secuencias del DNAr 16s es un herramienta poderosa para deducir la filogenia, evolución y la relación entre bacterias, arqueobacterias y organismos eucariotas, ya que el análisis de estas secuencias, puede revelar la presencia de una o varias secuencias caraterísticas que se denominan oligonucleótidos base, se trata de secuencias específicas cortas que aparecen en todos (o en la mayor parte de) los miembros de un determinado grupo filogético, y nunca (o solo raramente) están presentes en otros grupos, incluidos los más próximos (Rodicio y Mendoza, 2004; Tabssum et al., 2018). La identficacion basada en la secuencia del gen que codifica el DNAr 16s en los laboratorios microbiológicos se centra en cepas cuya identificación por métodos fenotípicos resulta imposible, como es el caso de bacterias no cultivables, por sus requerimientos nutricionales complejos o su lento crecimiento. Sin embargo, la taxonomía bacteriana basada en la secuencia del ADN debe incluir observaciones fenotípicas decisivas, para un enfoque equilibrado (Tabssum et al., 2018) 1.9.2.2. Análisis bioinformatico de secuencias Se define a la bioinformática como un campo de la ciencia en la cual confluyen varias disciplinas tales como la computación y la tecnología de la información (Wang et al., 2009). Las secuencias obtenidas, son alineadas y comparadas con secuencias ya depositadas en las bases de datos disponibles, como el Genbank de la National Center for Biotechnology Infrmation (NCBI) de los Estados Unidos y el European. Molecular. Biology. Laboratoy. (EMBL). en. el. Europen. Bioinformatics Institute (Aguilar-Ramírez, 2002), y se pueden utilizar herramientas como Basic Local Alignment Search Tool (BLAST) (Kent, 2002) y ClustalW (Chenna et al., 2003) para los respectivos análisis bioinformáticos. 34.
(42) BLAST es un programa informatico de alineamiento de secuencias de tipo local, ya sea del ADN o de proteínas, el programa es capaz de comparar una secuencia problema (también denominada como secuencia query) contra una gran cantidad de secuencias que se encentran en uan base de datos, el algoritmo de BLAST encuentra las secuencias de la base de datos mas parecido a la secuencia problema, normalmente es utlizado para encontrar secuencias homólogas (Wang et al., 2009). 1.9.3. Árboles filogenéticos y cladogramas: El análisis filogenético es importante estudiarlo ya que permite estudiar las relaciones evolutivas entre especies y la clasificación de estos organismos en familia, género o especie, además de ello es importante por que nos ayuda a entender el proceso de evolución adaptativa a nivel molecular. Existen tres métodos comúnmente utilizados en estudios de sistemática filogenética: I) la Cladística, usando el principio de Máxima Parsimonia; II) Maximum Likelihood y la III) Inferencia Bayesiana. En el método de la Máxima Parsimonia se usa un mínimo de presunciones a priori sobre los caracteres utilizados como fuente de información —se asume que cualquier carácter heredable es una homología potencial. Entonces, todos los caracteres son tratados de igual manera, es decir con la misma influencia al momento de inferir los árboles filogenéticos debido a que no se puede (o no se quiere) identificar homoplasias a priori. En la Máxima Parsimonia, el árbol filogenético que se prefiere es el que implica la mínima cantidad de cambios evolutivos (“pasos” evolutivos) que se requieren para explicar una determinada matriz de caracteres (Kent, W. J. 2002). Maximum Likelihood y la Inferencia Bayesiana son métodos estadísticos basados en modelos de evolución molecular, donde se toma en cuenta conocimiento a priori acerca de los caracteres, especialmente cuando son caracteres moleculares (frecuentemente secuencias de nucleótidos de ADN). El método Maximum Likelihood estima la probabilidad de qué tan bien la matriz de caracteres es explicada por los árboles filogenéticos, mientras que la Inferencia Bayesiana estima la probabilidad de qué tan bien los árboles filogenéticos son explicados por los datos (la matriz de caracteres) (Huelsenbeck et al. 2001, Brooks et al. 2007). 35.
Figure



+7




Documento similar
Por lo tanto, el objetivo del presente estudio fue evaluar el efecto de la irradiación UV-C sobre los cambios en el contenido de fenoles y flavonoides totales, β-caroteno,
Resumen: Se estudió el efecto de la radiación UV-C sobre la flora nativa y la capacidad antioxidante de la mezcla para té compuesto por toronjil ( Melissa officinalis ), ortiga
Determinar el efecto del uso combinado de la radiación UV-C y atmósfera modificada sobre el contenido de los compuestos antioxidantes: fenoles totales y
Con relación a este último punto, en el presente trabajo se planteó como objetivo estudiar el efecto de la radiación UV-B artificial sobre la síntesis de pigmentos
La radiación UV-C es un tecnología emergente que puede ser muy útil para mejorar la calidad nutricional y microbiológica de los frutos, sin embargo es importante estudiar el efecto
