Efecte del pH sobre el valor de Tm en diferents versions de Npu DnaE 102 Taula 4.3. Valors de t1/2 en diferents reaccions de tall i empalmament in vitro Npu DnaE C+1S en.
Valors dels Kobs observats en les diferents reaccions de tall i unió in vitro de. Taula que mostra els valors p obtinguts en el test estadístic t-student per veure si hi havia diferències entre la quantitat total de producte tallat i la unió formada.
Codi de tres lletres Codi d’una lletra Aminoàcid
ÍNDEX GENERAL
Expressió, purificació i caracterització de l’estabilitat térmica de la
GB1 com a exteïna amb doble mutació C1S i C+1S 115 4.3 Estudi estructural per RMN del procés associatiu de la inteïna Npu DnaE 117 4.3.1 Construcció, expressió i purificació de variants INIC_sc 118 4.3.2 Anàlisi bioinformàtica i predicció de la grau de plegament 121 4.3.3 Anàlisi d'estructura secundària amb dicroisme circular 124 4.3.4 Anàlisi d'estructura amb ressonància magnètica nuclear 125.
RESUM
Per aconseguir-ho, vam purificar i estudiar el plegament de tres variants de Npu DnaE que tenen diferents longituds del fragment IC. Aquesta reconstitució de l'estructura global ja es nota en els primers 21 residus del fragment IC.
RESUMEN
Para ello, hemos purificado y estudiado el plegamiento de tres variantes de Npu DnaE que tienen diferentes longitudes de fragmento IC. De esta forma, se confirma que el plegamiento global de esta inteína está condicionado por una sinergia en el plegamiento de los fragmentos IN e IC.
SUMMARY
Introducció
- LES INTEÏNES .1 Característiques generals
- Descobriment
- Organització
- Nomenclatura
- EL MECANISME DE TALL I UNIÓ .1 El procés
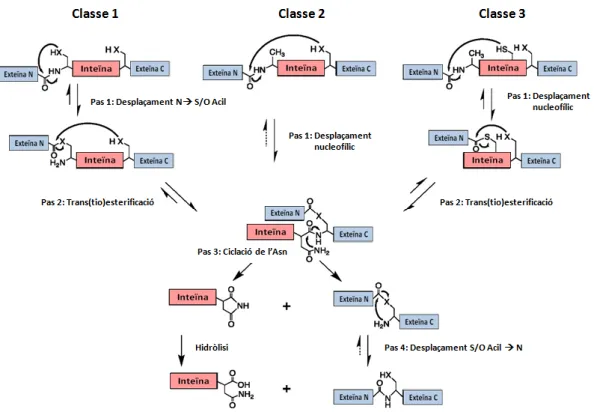
El mecanisme de tall i unió va romandre desconegut durant 6 anys a causa de la velocitat del procés. Al mateix temps, es produeix la ciclització d'un Asn de l'extrem C-terminal de la inteïna i la formació d'un enllaç peptídic que és una característica exclusiva del procés de tall i empalmament de proteïnes (Xu i Perler, 1996).

DESPLAÇAMENT N S/O ACIL
Aquest procés té lloc a través d'un conjunt d'atacs nucleòfils dels residus N- i C-terminals que provoquen atacs als enllaços amida i èster i fins i tot als seus grups amino.
TRANS(TIO)ESTERIFICACIÓ
El pas 1 és el més ràpid de tot el mecanisme de tall i unió i la velocitat de reacció d'aquest pas pot ser fins a 100 vegades més ràpida que tot el procés global (Romanelli et al., 2004). Du i col·laboradors van establir que His B:10 primer actua com a base per desprotonar la cadena lateral de Cys 1 (A:1) i després dona protons al nitrogen amida de l'enllaç peptídic (Du et al., 2009). la reacció a favor
- Classificació
- Alteracions en residus clau de la seqüència de la inteïna
- Inteïnes sense la His 10 de la regió B
- Inteïnes sense la penúltima His
- Inteïnes amb Gln o Asp com a últim residu
- El procés de tall i unió en trans
- APLICACIONS
- Expressió i modificació de proteïnes
- Inteïnes com a mediadores de l’etiquetatge de proteïnes
- Regulació de l’activitat proteica a partir del mecanisme de tall i unió condicionat
- Inteïnes com a biosensors
- Requisits de les inteïnes per ser utilitzades com a eines biotecnològiques
- LA INTEÏNA Npu DnaE
El seu 10 de la regió B (figura 1.3) és essencial per al primer pas del mecanisme de tall i unió. El mecanisme de tall i unió escindirà els fragments de la inteïna i connectarà la proteïna pels seus extrems N i C.

CLSYETEILT VEYGLLPIGK IVEKRIECTV YSVDNNGNIY
Inteïnes més utilitzades en el camp de la biologia molecular per al desenvolupament de diferents tècniques. La taula mostra el nombre d'aminoàcids per a cada fragment d'inteïna i la semivida (t1/2) calculada a partir de la constant de primer ordre (Vila-Perello i Muir, 2010).
TQPVAQWHDR GEQEVFEYCL EDGSLIRATK DHKFMTVDGQ
La inteïna Npu DnaE de Nostoc punctiforme PCC73102 forma part de la família d'inteïnes DnaE descrita en cianobacteris (Caspi et al., 2003) i pertany a la classe d'inteïnes 1 tot i que no té el penúltim His a la regió G. precursor de la subunitat α de l'ADN polimerasa III d'aquest organisme.
MLPIDEIFER ELDLMRVDNL PN
Fins ara, una de les inteïnes que millor compleix aquests requisits és la trans-inteïna Npu DnaE.
MIKIATRKYL GKQNVYDIGV ERDHNFALKN GFIASN
- L’estructura tridimensional
- Cinètica i afinitat d’unió de la inteïna
- Robustesa de la inteïna
- Promiscuïtat de la inteïna
- El procés associatiu dels fragments I N i I C
- Objectius
Tanmateix, és essencial que el primer residu de la inteïna sigui un Cys (Cheriyan et al., 2013). Quan els dos fragments d'inteïnes tenien una càrrega neta negativa o positiva, no es va observar cap activitat.

OBJECTIUS
Material i Mètodes
- MATERIAL BIOLÒGIC .1 Soques bacterianes
- E. coli DH5α
- E. coli BL21(DE3)
- Vectors procariotes
- MÈTODES MICROBIOLÒGICS
- Medis de cultiu
- Antibiòtics
- Manteniment de les soques
- Creixement dels cultius
- TÈCNIQUES DE DNA RECOMBINANT .1 Manipulació del DNA
- Obtenció de DNA plasmídic
- Electroforesi de DNA en gel d’agarosa
- Purificació de fragments de DNA a partir de gels d’agarosa
- Estimació espectrofotomètrica de concentracions de DNA
- Utilització d’enzims que modifiquen el DNA
- Mutagènesi dirigida per oligonucleòtid: Mètode “QuikChange TM ”
- Seqüenciació de DNA
- MÈTODES D’EXPRESSIÓ I PURIFICACIÓ DE PROTEÏNES RECOMBINANTS
- Expressió de la inteïna Npu DnaE, I N – I C , I N -GB1 i I C -GB1
- Expressió de I N , les variants I N I C _sc i de la proteasa TEV
- Purificació de la NpuDnaE i de I N -I C
- Purificació de I N i de les variants I N I C _sc
- Purificació de I N -GB1 i I C -GB1
- Purificació de la proteasa TEV
- ANÀLISI I CARACTERITZACIÓ PROTEICA .1 Determinació de la massa molecular
- Determinació de la quantitat de proteïna
Per a la clonació i l'expressió de la inteïna Npu DnaE descrita en aquest treball, es va utilitzar el vector pRSET/EmGFP d'Invitrogen (EUA). Per a la clonació i expressió de Npu DnaE IN descrit en aquest treball, es va utilitzar el vector pSKDuet01 d'Adgene (EUA). Per a la clonació i expressió de l'IC Npu DnaE descrit en aquest article, es va utilitzar el vector pSKBAD2, també d'Adgene (EUA).
La fracció soluble es fa passar per una columna d'afinitat Ni-NTA, de la qual s'elueix la proteïna d'interès amb un gradient d'imidazol. A continuació, es realitza la digestió de la proteasa TEV per separar la proteïna d'interès de la proteïna solubilitzant. Les cèl·lules de la centrifugació d'un litre de cultiu es tornen a suspendre en 80 ml de Tris-HCl 25 mM, NaCl 150 mM pH 8.
Aquesta vegada, però, la proteïna s'elueix mitjançant cromatografia, que s'ha de recollir. La fracció soluble es dialitza contra el tampó d'equilibri en una columna Ni-NTA, de la qual s'elueix amb un tampó amb una alta concentració d'imidazol.
- Electroforesi discontinua en gel de poliacrilamida
- Western blot
- Determinació de l’estabilitat tèrmica
- Estudi del procés de tall i unió
- Càlcul de paràmetres cinètics
El procés de purificació de la proteasa TEV segueix el mateix protocol que es descriu a la secció 2.4.5. Connecteu la font d'alimentació a una tensió constant de 150-200v i deixeu que la separació avanci fins que el tampó d'aplicació que marca el límit de la separació arribi a l'extrem inferior del gel (aproximadament una hora). Els canvis en l'entorn dels residus aromàtics durant aquest procés de desplegament es poden controlar mitjançant mesures d'absorbància a 280 nm, que permeten determinar la temperatura de fusió intermèdia (Tm).
ΔCp: Capacitat calorífica en Kj/Kmol n: punt de tall pre-transició d: punt de tall post-transició q: tall pre-transició. D'aquest ajust s'obté la temperatura de fusió (Tm), que determina l'estabilitat de la proteïna en relació a la temperatura. El càlcul dels paràmetres cinètics del procés de tall i unió es va realitzar a partir de la quantificació del producte de tall i unió a partir dels gels SDS-PAGE obtinguts.
Per normalitzar l'error de càrrega de la mostra, la intensitat de cada banda d'una columna es va expressar com una fracció de la intensitat total de la columna analitzada. El temps de vida mitjana (t1/2), el temps necessari perquè es formi la meitat del producte total observat, es va calcular a partir del valor de la constant de Kobs de primer ordre:
Anàlisi de l’estructura secundària per dicroisme circular
Es van representar les intensitats en funció del temps i la corba resultant es va ajustar a una equació exponencial de primer ordre mitjançant el programari SigmaPlot (Jandel, Regne Unit). L'absorció desigual de la llum polaritzada circularment (activitat òptica) de proteïnes a la regió ultraviolada llunyana o a la regió amida nm) depèn principalment dels enllaços peptídics. Així, les bandes CD de les regions amida contenen informació sobre els enllaços peptídics i sobre l'estructura secundària de la proteïna.
Es van registrar espectres de dicroisme circular entre 200 i 260 nm (UV llunyà) amb la cambra de mostra purgada contínuament amb nitrogen. La cubeta es va rentar prèviament amb una solució d'àcid nítric al 50% i es va esbandir amb aigua milliQ. Després d'introduir les mesures de la trajectòria de la llum de la cubeta (en cm) i la molaritat de la mostra (M), es mesura l'el·lipticitat molar *θ+ (deg·cm2·dmol-1).
On θobs és l'el·lipticitat mesurada en mdeg, MRW és la massa molecular mitjana per residu (Da), c és la concentració de proteïnes en mg/ml i l és el pas òptic de la cubeta en cm. A partir dels valors d'el·lipticitat mitjans per residu, es va fer una predicció de l'estructura secundària de la proteïna mitjançant els programes K2D3 (Louis-Jeune et al., 2011) i Dicroprot (Deleage i Geourjon, 1993).
ESPECTROSCÒPIA DE RESSONÀNCIA MAGNÈTICA NUCLEAR (NMR)
- Assignació de les variants I N I C _sc
ANÀLISI BIOINFORMÀTICA
ANÀLISI ESTADÍSTICA
Resultats i Discussió
- EXPRESSIÓ, PURIFICACIÓ I CARACTERITZACIÓ DE L’ESTABILITAT TÈRMICA DE LA INTEÏNA Npu DnaE,
- Construcció, expressió i purificació de Npu DnaE formant una única cadena polipeptídica
- Expressió i purificació de les subunitats I N -I C de Npu DnaE associades
- Construcció, expressió i purificació de la subunitat I N de Npu DnaE
- Caracterització de la inteïna Npu DnaE, de les subunitats I N – I C associades i de I N aïllada
- ESTUDI DEL PROCÉS DE TALL I UNIÓ
- Estudi del procés de tall i unió en cis de la Npu DnaE C+1S utilitzant la GFP com a exteïna
- Estudi del procés de tall i unió en trans de la Npu DnaE C+1S utilitzant GB1 com a exteïna
- Estudi del procés de tall i unió en trans de la Npu DnaE utilitzant GB1 com a exteïna amb la doble mutació C1S i C+1S
- ESTUDI ESTRUCTURAL MITJANÇANT RMN DEL PROCÉS ASSOCIATIU DE LA INTEÏNA Npu DnaE
- Construcció, expressió i purificació de les variants I N I C _sc
- Anàlisi bioinformàtica i predicció del grau de plegament
- Anàlisi de l’estructura secundària per dicroisme circular
- Anàlisi de l’estructura per ressonància magnètica nuclear
El mateix es va fer amb el gen de la subunitat IC, fent-ho a l'extrem 5'. Això va ser seguit pel procés de purificació mitjançant HPLC d'intercanvi d'anions de la fracció soluble. La figura 4.4 mostra un esquema de la construcció i com es realitza el mecanisme de tall i unió.
La digestió amb TEV es va realitzar per eliminar la MBP de la subunitat IN abans dels assajos de clivage. Abans de començar el procés de producció i purificació de la subunitat IN de Npu DnaE, es va analitzar el grau d'expressió de diferents clons. Npu DnaE és un intein molt interessant en aplicacions biotecnològiques per la seva eficiència en el procés de tall i unió.
Valors de Kobs observats en diferents reaccions de tall i empalmament de NpuDnaE C+1S in vitro en cis*. El gràfic (a) de la figura 4.20 mostra el desenvolupament en percentatge del disseny del producte de tall i empalme en hores per. Esquema de la construcció realitzada en el vector pBADm41 per facilitar el procés de purificació de les variants INIC_sc.
Els primers espectres que es van realitzar van ser els de la subunitat IN i els de tota la inteïna Npu DnaE.

Cadena 2 Sentit Es manté la relació?
- Discussió General
- DISCUSSIÓ GENERAL
- Conclusions
- Bibliografia
En el cas de Npu DnaE, la majoria de les cadenes β es troben en una orientació antiparal·lela. La relació entre les diferents cadenes β de la inteïna Npu DnaE i si aquesta relació es manté entre les variants INIC_sc123 i INIC_sc131. En aquest cas, queda clar que el Cys a la posició 1 és crucial per al mecanisme de tall i unió a la inteïna Npu DnaE.
Grau d'identitat que presenten les seqüències d'inteina Ssp DnaE (blau) i Npu DnaE (vermell). En el cas de Npu DnaE amb la mutació C+1S, l'energia s'obté mitjançant un augment de la temperatura. En conjunt, en aquests estudis cinètics s'ha pogut comprovar que un Ser en posició 1 o +1 inhibeix l'activitat de la inteïna a temperatura ambient.
A més, la termoestabilitat de les exteïnes unides pot afectar significativament l'activitat de les inteïnes. En el cas de la subunitat IN, Ssp DnaE presenta un fragment de 21 residus que no es troba a Npu DnaE. S'ha observat com els primers residus de la subunitat IC ja contribueixen molt al plegament de la inteïna.
El plegament d'una inteïna està condicionat per com es connecten les seves subunitats.