XXI Congreso Internacional y XLVI Congreso Nacional de la Sociedad Mexicana de Fitopatología, A. El genoma del bacteriófago ΦXaF-18 asociado a la bacteria fitopatógena Xanthomonas vesicatoria. Certificado por la presentación de póster con el tema: Caracterización molecular del bacteriófago ΦXaf18 asociado a la bacteria fitopatógena Xanthomonas vesicatoria. Reconocimiento por la presentación oral a nivel Estudiante con la asignatura: Genoma del bacteriófago ΦXaf18 asociado a la bacteria fitopatógena Xanthomonas vesicatoria.
Características de los marcos de lectura abiertos (ORF) previstos. genoma del bacteriófago ΦXaF18.………. Comparación de perfiles de restricción in vitro con perfiles. restricción in silico del genoma del bacteriófago ΦXaF18 expuesto a la técnica. Comparación de perfiles de restricción in vitro con perfiles. Restricción in silico del genoma del bacteriófago ΦXaF18 con la enzima BclI……….
INTRODUCCIÓN
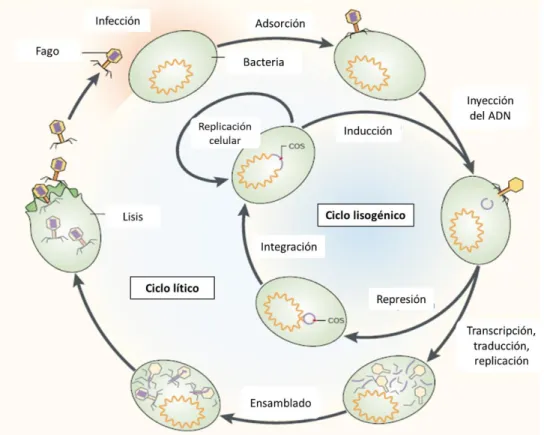
Mientras que el ciclo de vida lisogénico consta de: adsorción, integración del genoma viral al genoma bacteriano y replicación del genoma viral junto con el genoma bacteriano. La abundancia e importancia ecológica de los fagos en la mayoría de los ambientes, acompañada de un conocimiento limitado de su estructura genética, requiere el establecimiento de métodos. Hasta la fecha, obtener una secuencia genómica completa sigue siendo una de las formas más efectivas de profundizar en la biología de un organismo, especialmente para microorganismos cuya biología es difícil de estudiar en el entorno natural o en el laboratorio (Henn et al., 2010).
La facilidad y el bajo costo de la secuenciación de fagos, combinados con el conocimiento de los modelos de fagos, pueden otorgar a la genómica de fagos un papel importante en la genética de poblaciones, en el desarrollo de genomas simples y en el modelado de secuencias de ADN (Brϋssow y Hendrix, 2002). Esta enfermedad está ampliamente distribuida en el mundo (EPPO) y se ve favorecida por las altas temperaturas (25 a 30°C) y la alta humedad ambiental (Mirik y Aysan, 2009; Balogh et al., 2003). El uso de bacteriófagos en la agricultura es una alternativa para combatir enfermedades bacterianas; se han utilizado con éxito para controlar enfermedades de las plantas (Balogh et al., 2003).
ANTECEDENTES
- Agente causal de la mancha bacteriana en tomate y chile
- Tratamiento tradicional de la mancha bacteriana
- Tratamiento de la mancha bacteriana con bacteriófagos
- Diversidad y abundancia de los bacteriófagos
- Clasificación de los bacteriófagos
- Morfología de los bacteriófagos
- Secuenciación de los genomas de bacteriófagos
- Organización genómica de los bacteriófagos
- Interacción bacteria-bacteriófago
- Genómica comparativa
- Metagenómica
- Empaquetamiento del ADN
- Mecanismos genéticos-moleculares para decidir el ciclo lisis-lisogénia…. 22
El análisis de los datos genómicos de los fagos proporciona una imagen clara de la gran diversidad genética de esta población y las similitudes fundamentales. Sin embargo, la frecuencia de los eventos de recombinación puede depender de la distancia filogenética entre los huéspedes (Lima-Méndez et al., 2011), siendo los bacteriófagos más relacionados filogenéticamente los que presentan más homologías. Los bacteriófagos dictan muchos aspectos de la biología bacteriana, afectando muchos aspectos de la fisiología (Clokie et al., 2011), la ecología y la evolución de las bacterias y, por lo tanto, desempeñan funciones importantes en los ecosistemas, p. el ciclo del carbono y
Existe evidencia de transferencia horizontal de genes, por ejemplo, secuencias similares encontradas en fagos muy distantes o genomas de fagos templados en el cromosoma bacteriano (profago) (Hendrix, 2002). Comparando secuencias, es posible comprender estos y otros aspectos de la biología de los bacteriófagos. Al realizar BLAST para identificar homólogos cercanos de la proteína AimR en fagos y profagos, encontraron 112 homólogos, la mayoría pertenecientes al grupo de fagos SPbeta (Erez et al., 2017).

JUSTIFICACIÓN
HIPÓTESIS
OBJETIVOS
Objetivo general
Objetivos específicos
METODOLOGIA
- Bacteriófago/bacteria huésped
- Purificación y multiplicación del bacteriófago ΦXaF18
- Concentración del título del bacteriófago mediante la técnica con
- Conteo de placas de lisis mediante ensayo de doble placa en agar
- Caracterización morfológica del bacteriófago ΦXaF18
- Purificación mediante gradiente de cloruro de cesio (CsCl)
- Tinción y visualización en microscopio electrónico de transmisión
- Caracterización genómica del bacteriófago ΦXaF18
- Extracción de ADN del bacteriófago
- Digestión del ADN para determinar su naturaleza
- Polimorfismo de longitud de fragmentos de restricción (RFLPs) del
- Secuenciación y ensamblado del genoma viral
- Ensayos de PCR
- Análisis bioinformático
- Análisis genético-fenotípico del mecanismo de regulación
- Búsqueda de genes implicados en el mecanismo de regulación
- Búsqueda del péptido arbitrium en el genoma del bacteriófago
- Búsqueda del mecanismo lisis-lisogénia in vivo mediante
Se realizó una centrifugación en gradiente de CsCl para eliminar los residuos contaminantes que impedían la observación en microscopio electrónico de transmisión (TEM) del bacteriófago recuperado de la concentración con PEG8000. La PCR para el bacteriófago se realizó con ocho conjuntos de oligonucleótidos, uno de los cuales corresponde a la secuencia codificante de la subunidad principal de la proteína terminasa (M1, M2, M3, M4, M5, M6, M7 y TER1). Además, se realizó una búsqueda de dominios conservados utilizando los servidores Prosite de Expasy y el Servicio de búsqueda de dominios conservados (CD Search) del NCBI (Sigrist et al., 2010; Marchler-Bauer et al., 2015).
Las búsquedas de ARN de transferencia y ARN ribosómico se realizaron utilizando tRNAscan-SE, RNAmmer y Rfam (Lowe y Eddy, 1997; Lagesen et al., 2007; Griffiths-Jones et al., 2003). Primero, se realizó una alineación de la secuencia de la proteína terminasa de la subunidad principal del bacteriófago ΦXaF18 en Clustal y la distancia de separación de espacios de 5 para ambos. BLASTn se realizó con la secuencia completa del genoma del bacteriófago ΦXaF18 para determinar su similitud con otros bacteriófagos.
Luego de lo cual se realizó un análisis comparativo en el programa Mauve 20150226 (Darling et al., 2004) con el bacteriófago KPhi1. La búsqueda de los genes implicados en el mecanismo de regulación de la lisis-lisogenia, así como la búsqueda del péptido Arbitrium, se realizó en el genoma del bacteriófago ΦXaF18. Por otro lado, se realizó medio acondicionado y posterior evaluación mediante una curva de crecimiento bacteriano para determinar la existencia en el medio de eventuales moléculas liberadas por bacteriófagos que intervienen en la decisión lisogénico-lisogénica del fago ΦXaF18.
El cultivo se centrifuga a 4000 rpm durante 10 minutos a temperatura ambiente, se descarta el sobrenadante y se resuspende el sedimento en un nuevo medio líquido NYG, a partir de esta suspensión se hace un nuevo cultivo, se ajusta la DO600 a 0,1 y se deja incubar. a 28°C y 200 rpm durante 6 horas. Se realizó una curva de crecimiento bacteriano para evaluar la posible presencia de moléculas de señalización en el medio condicionado. El cultivo se centrifugó a 4000 revoluciones por minuto durante 10 minutos a temperatura ambiente, se descartó el sobrenadante y el sedimento se resuspendió en 5 ml de un nuevo medio líquido NYG a 28 °C y 200 revoluciones por minuto durante 6 horas.
Se realizó otra curva de crecimiento de la misma manera, excepto que el bacteriófago se inoculó después de 24 horas y se evaluó hasta 40 horas.

RESULTADOS Y DISCUSIÓN
Purificación y multiplicación del bacteriófago ΦXaF18
- Precipitación de bacteriófagos con Polietilenglicol/NaCl
Determinación de la multiplicidad de infección (MOI) adecuada para la fase exponencial del fago ΦXaF18.
Determinación de la multiplicidad de infección (MOI) adecuada para la
Caracterización morfológica
La Figura 8 muestra las partes del bacteriófago ΦXaF18, mostrando la cápside, el cuello, la cola, la placa basal y la espícula; La presencia de las fibras adhesivas no se pudo observar en ninguna de las micrografías electrónicas, quizás porque eran demasiado delgadas y la resolución no era suficiente, o por otro lado podrían haberse perdido durante el proceso de preparación de la muestra.
Caracterización genómica…
- Determinación de la naturaleza del material genético
- Caracterizacion del ADN del bacteriófago ΦXaF18 por RFLPs
- Secuenciación, ensamblado del genoma del bacteriófago
- Ensayos de PCR para confirmar ensamblado del genoma de
- Diseño de oligonucleótidos
- PCR de la bacteria Xanthomonas vesicatoria
- PCR a partir del genoma del bacteriófago ΦXaF18
- Análisis bioinformático
- Marcos abiertos de lectura (ORF)
- Módulos funcionales
- Predicción de secuencias funcionales
- Empaquetamiento del genoma viral y tipo de extremos
- Análisis filogenético
- Análisis comparativo de genomas de bacteriófagos de
- Genes implicados en el mecanismo de regulación lisis-lisogénia en
- Péptido arbitrium en el genoma del bacteriófago ΦXaF18
- Determinación in vivo del mecanismo de decisión lisis-lisogénia
El bacteriófago ΦXaF18 tiene un genoma de 47.407 pares de bases, lo que lo sitúa dentro de los bacteriófagos de la familia Myoviridae que tienen genomas pequeños. Se realizaron ensayos de PCR como herramienta para la identificación tanto de la bacteria como del bacteriófago. La Figura 18 muestra el genoma del bacteriófago ΦXaF18, en el que se pueden observar los 67 marcos de lectura abiertos y cuatro regiones según la dirección de transcripción.
El genoma del bacteriófago ΦXaF18 se divide en cuatro módulos funcionales: el primer y tercer módulo son estructurales, sin embargo se dividen según el destino final de la proteína, por lo que el primer módulo también tiene el gen que codifica la subunidad terminal mayor de la proteína. como genes que codifican las proteínas estructurales de la cápside; El genoma del bacteriófago ΦXaF18 tiene dos módulos estructurales, uno que contiene genes implicados en la morfogénesis de la cápside y el otro con genes correspondientes a la morfogénesis de la cola; Esta organización de genes estructurales divididos en dos módulos también ha sido informada por Comeau et al. En el primer módulo, junto con las proteínas de la cápside, se encuentra el gen de la subunidad principal de la proteína terminal.
El segundo módulo estructural contiene proteínas de la fibra de la cola, así como proteínas de la placa basal, como informaron Comeau et al. La secuencia de la proteína terminasa de la subunidad principal está altamente conservada, lo que la convierte en una excelente opción para realizar análisis filogenéticos (Kanamaru et al., 2004; Sun et al., 2012). Los ORF 42 y 53 corresponden a proteínas de la fibra de la cola, ya que mostraron similitud con las proteínas de la fibra de la cola del fago OP2 de Xanthomonas (identidad = 77%) para el primer caso y con el fago BcepNY3 de Burkholderia (identidad = 57%) para el segundo caso.
En cuanto a ORF 38, se identificó como una proteína relacionada con la virulencia de la familia E, con una identidad del 99% con la proteína relacionada con la virulencia de la familia E del fago. ORF 56 fue identificada como una lisozima, mostró similitud con una enzima lítica del fago Xanthomonas OP2 (identidad = 0,75%), además, presentó el dominio conservado de la superfamilia de lisozima. ORF 59 corresponde a una proteína resolvasa, que muestra un 70% de identidad con una proteína resolvasa del fago Xanthomonas XPV1, así como un dominio conservado de la superfamilia RusA.
ORF 67 se identificó como una proteína endonucleasa ya que era similar a la superfamilia PD-(D/E)XK de proteínas nucleasas del fago DU_RP_II de Ralstonia (identidad = 0,27%) y representaba un dominio conservado de la proteína Cas4 relacionada en CRISPR. Árbol filogenético construido con la secuencia de aminoácidos de la subunidad principal terminasa del bacteriófago ΦXaF18 y otras secuencias de terminasa de bacteriófagos del orden Caudovirales. Árbol filogenético construido con la secuencia de aminoácidos de la subunidad mayor terminasa del bacteriófago ΦXaF18 y otras secuencias terminasas de bacteriófagos de la familia Myoviridae.

CONCLUSIONES
Bouzar H, Jones JB, Stall RE, Louws F J, Schneider M, Rademaker JL W, de Bruijn F J, and Jackson LE (1999) Multiphasic Analysis of Xanthomonads Causing Bacterial Spot Disease on Tomato and Pepper in the Caribbean and Central America: Evidence for Common genera within and between countries. Elbreki M, Ross RP, Hill C, O'Mahony J, McAuliffe O y Coffey A (2014) Bacteriophages and their derivatives as biotherapeutic agents in disease prevention and treatment. Xanthomonas euvesicatoria, Xanthomonas gardneri, Xanthomonas perforans, Xanthomonas vesicatoria), which causes bacterial spot of tomato and sweet pepper. Gašic K, Kuzmanovic N, Ivanovic M, Prokic A, Ševic M, Obradovic A (2018) Complete Genome of Xanthomonas euvesicatoria Specific Bacteriophage Kphi1, Its Survival and Potential in Pepper Bacterial Spot Control.
Inoue Y, Matsuura T, Ohara T y Azagami K (2006) Genome sequence analysis of OP2, a lytic bacteriophage of Xanthomonas oryzae pv. Iriarte F B, Balogh B, Momol M T, Smith L M, Wilson M y Jones J B (2007) Factors affecting bacteriophage survival on tomato leaf surfaces. Lima-Mendez G, Toussaint A y Leplae R (2011) A modular view of bacteriophage genomic space: identification of host and lifestyle marker modules.
Marco GM, Stall RE (1983) Control of bacterial spot on pepper initiated by strains of Xanthomonas campestris pv. Obradovic A, Jones JB, Momol MT, Balogh By Olson S M (2004) Management of tomato bacteria in the field by foliar applications of bacteriophages and SAR inducers. Rohwer F, Segall A, Steward GF, Seguritan V, Breitbart M, Wolven F y Azam F (2000) The complete genomic sequence of the marine phage Roseophage SIO1 shares homology.
Sadunishvili T, Kvesitadze E, Kvesitadze G (2015) Chapter 3 Xanthomonas vesicatoria-specific virus and its potential to prevent tomato bacterial spot disease. Sun S, Gao S, Kondabagil K, Xiang Y, Rossmann M G y Rao VB (2012) Structure and function of the small terminase component of the DNA packaging machinery in T4-like bacteriophages. Valbuzzi A, Gollnick P, Babitzke P y Yanofsky C (2002) The anti-trp RNA-binding attenuation protein (Anti-TRAP), AT, recognizes the tryptophan-activated RNA-binding domain of the TRAP Regulatory Protein.
Wittebole X, De Roock Sy Opal S M (2014) A historical overview of bacteriophage therapy as an alternative to antibiotics for the treatment of bacterial pathogens.
Artículo enviado a revisión a la revista Microbiology Resource
Secuencia genómica completa
Constancias de participación en congresos
Medios de cultivo y buffers







