El impacto de la Ley de Bioterrorismo en la producción y procesamiento de alimentos: historia, regulaciones y plan de seguridad. NUEVAS PERSPECTIVAS SOBRE LA SEGURIDAD ALIMENTARIA 14 los genes y el ADN encuentran cada vez más aplicaciones en la industria alimentaria. En los últimos años, los ciudadanos de la UE se han preocupado por la seguridad de los alimentos que consumen.
La extracción de fungicidas de la matriz de la uva utiliza la mezcla diclorometano/acetona (75/25, v/v). NUEVAS PERSPECTIVAS SOBRE SEGURIDAD ALIMENTARIA 46 Finalmente, el método propuesto se ha aplicado al análisis de muestras de uva blanca para vinificación de la D.O. NUEVAS PERSPECTIVAS SOBRE LA SEGURIDAD ALIMENTARIA 100 Los valores de equivalencia tóxica (TEQ) siguieron esencialmente el mismo patrón de distribución que los niveles de los otros congéneres de PCB (Atuma, S. S., et. al., 1998).
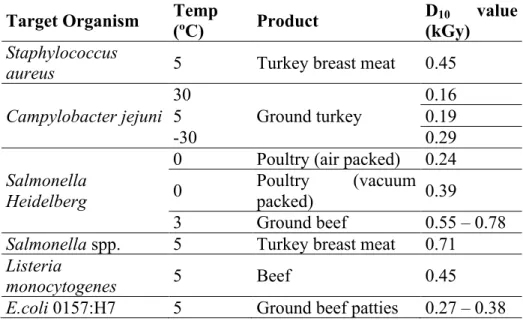
NUEVAS PERSPECTIVAS SOBRE SEGURIDAD ALIMENTARIA 109 Capítulo 4 Microbiología de la irradiación de alimentos Microbiología de la irradiación de alimentos.

Las bacteriocinas y la conservación de los alimentos
Son muchas las aplicaciones desarrolladas de las bacteriocinas en los alimentos incorporadas ya sea mediante la fermentación del producto por las bacterias iniciales que son, a su vez, productoras (producción in situ) -opción sólo viable cuando la flora láctica es mayoritaria-, o añadiendo la bacteriocina. directamente en los alimentos (Schillinger et. al., 1996). Destaca la aplicación en productos lácteos, especialmente en quesos para la prevención de Listeria monocytogenes (Davies et. al., 1997); en productos fermentados como alternativa a la adición de nitritos (Taylor et. al., 1984) y en productos pesqueros como alternativa a los ácidos benzoico y ascórbico en el almacenamiento refrigerado de artemia (Einarsson y Lauzon, 1995).
La producción de probióticos y bacteriocinas
Además, en el caso de aplicaciones en alimentación animal, debido a la naturaleza de las fuentes de nitrógeno utilizadas (principalmente peptonas cárnicas), no se pueden incorporar junto con los probiontes en el pienso, lo que supone un proceso previo de separación de la cosecha microbiana que lo hace más caro de obtener. De hecho, existen precedentes del uso de aguas residuales del procesamiento de mejillones y suero como materia prima para procesos microbianos para producir metabolitos de interés. Finalmente, se presentan dos aplicaciones de las bacteriocinas y la producción de biomasa: desarrollo de envases activos y sustitución de antibióticos en la alimentación animal.
Cultivos preliminares sobre SL y EPM
Cultivo en suero de leche
Por el contrario, en el caso de Pc 1.02, tanto el crecimiento como la producción de bacteriocina fueron aproximadamente 6 veces mayores que los obtenidos en SLD. NUEVAS PERSPECTIVAS SOBRE SEGURIDAD ALIMENTARIA 151 4: Efecto conjunto de las concentraciones de tampón, nitrógeno, fósforo y azúcar sobre la producción de bacteriocinas en medios residuales. En consecuencia, se desconocen las posibles interacciones y efecto conjunto de los nutrientes sobre la producción.

Cultivos en suero de leche diluido (SLD)
Además, el aumento de la variable P, contrariamente a su efecto sobre la síntesis de nisina, también tuvo un efecto negativo específico significativo sobre la producción de pediocina, lo que por otro lado indica la naturaleza secundaria del metabolito. En el caso de la nisina, el aumento de la variable T redujo la respuesta, en mayor medida a valores bajos de N, aunque apenas afectó a la producción de biomasa, lo que indica que su influencia en la síntesis de nisina se debe específicamente a la magnitud del ΔpH que crea en el cultura. Los resultados mostrados en la Figura 6 muestran que el efecto de la zona de amortiguamiento sobre la biomasa fue prácticamente insignificante, mientras que las diferencias en la producción de las tierras bajas se multiplicaron por un factor de tres.
Cultivos en efluentes del procesado de mejillón (medio 0,5 MH) Los resultados obtenidos de los cultivos sobre EPM se muestran en
Por el contrario, en el caso de la pediocina (Figura 5), la acción de la glicina y la variable C tuvo poca importancia para la respuesta, probablemente porque la lactosa y la glicina no son buenas fuentes de carbono y nitrógeno para Pediococcus. El aumento de la concentración del tampón provocó una disminución exponencial en los títulos de antibióticos que resonó con la disminución del tamaño del ΔpH. En el caso de la pediocina (Figura 8), la glicina tampoco fue una buena fuente de nitrógeno.

Cultivos en efluentes del procesado de mejillón (medio 0,5 MH) También sobre este medio y para ambas bacteriocinas, los resultados
Los resultados que condujeron a las superficies de respuesta que se muestran en las Figuras 9 y 10 muestran en ambos casos dos tipos básicos de comportamiento de producción de biomasa y bacteriocina dependiendo de la naturaleza de la fuente de nitrógeno utilizada. NUEVAS PERSPECTIVAS DE SEGURIDAD ALIMENTARIA 161 aminoácidos de las fuentes de N complejo, parece razonable pensar que los efectos positivos se deben a su alto contenido en minerales, vitaminas o los aminoácidos serina, cisterna y treonina, que se encuentran en niveles altos. proporción y se denominan precursores, específicos para la síntesis de nisina sin afectar el rendimiento final de las células (De Vuyst, 1995). En los apartados anteriores se demostró que el crecimiento de Lactococcus lactis y Pediococcus acidilactici y la producción de sus bacteriocinas depende de la concentración y naturaleza de las fuentes de nitrógeno, carbono y fósforo, así como del gradiente de pH creado en el cultivo.

Producción de nisina sobre 0,5 MH alimentado con glucosa
Esta intensificación de la actividad metabólica estuvo acompañada de un fuerte consumo de fuentes de nitrógeno y fósforo, mientras que la ingesta de glucosa, contrariamente a lo esperado, disminuyó respecto a la fase anterior. NUEVAS PERSPECTIVAS SOBRE SEGURIDAD ALIMENTARIA 170 Por otro lado, dado que la concentración de glucosa se mantuvo en un nivel alrededor de 5 g/L durante toda la incubación, es razonable creer que existe una limitación del flujo de carbono a partir de la segunda mitad de la incubación. fase de transición. Para verificar esta hipótesis, se realizaron balances de carbono durante el cultivo basándose en la estequiometría de los productos de la fermentación heteroláctica y teniendo en cuenta el consumo de glucosa implicado en el crecimiento y mantenimiento celular.

Producción de nisina sobre medio 0,5 MH alimentado con medio 10 MH
Sin embargo, a excepción del fósforo, cuyo consumo fue lineal al iniciar la fase de heterofermentación, el resto de nutrientes presentaron diferentes tasas de consumo durante los últimos 2/3 de la incubación. Así, el consumo de nitrógeno se ralentiza progresivamente, probablemente porque por el efecto combinado del consumo y el muestreo, aumenta la proporción relativa de la mencionada fracción proteica en el medio de cultivo, de difícil metabolización (Pastrana, 1991). Por otro lado, la absorción de glucosa se acelera en mitad de la fermentación, que corresponde al período de máxima actividad metabólica (en términos de biomasa y producción de heterometabolitos), para disminuir ligeramente hacia el final de la fermentación, cuando el crecimiento se vuelve lineal.

Producción de pediocina sobre medio 0,5 MH alimentado con glucosa
NUEVAS PERSPECTIVAS SOBRE SEGURIDAD ALIMENTARIA 175 biomasa y pediocina, y el inicio de la síntesis de metabolitos de la fase heterofermentativa tampoco fue simultáneo. Antes de las 88 horas, la producción de pediocina y el consumo de fósforo eran prácticamente lineales. Con el agotamiento del fósforo, la producción de ácido láctico se detuvo y la producción de biomasa y pediocina se desaceleró, mientras que el etanol apareció como un nuevo producto heteroláctico.

Producción de pediocina sobre medio 0,5 MH alimentado con medio 10 MH
AT, Pr, PT y NT: azúcares totales, proteínas, fósforo total y nitrógeno total aportados (Δ), consumidos (Ο) y restantes en el medio ambiente. NUEVAS PERSPECTIVAS SOBRE SEGURIDAD ALIMENTARIA 179 Sin embargo, incluso en este último caso, la fenomenología del cultivo puede explicarse en los mismos términos metabólicos, cinéticos y de equilibrio material que en el primer experimento. De hecho, aplicando el razonamiento utilizado para el balance de carbono y el consumo de nitrógeno (Figura 20), se puede observar que en todos los casos se utilizan proteínas, excepto como fuente de nitrógeno, como fuente de carbono, contabilizando así los déficits en el molar de carbono. balance.

Producción de bacteriocinas sobre suero de leche
NUEVAS PERSPECTIVAS SOBRE SEGURIDAD ALIMENTARIA 180 la aparición de un metabolismo heteroláctico en el caso de la nisina (Figura 21). Lact, Gluc, Pr, PT y NT: lactosa, glucosa, proteínas, fósforo total y nitrógeno total suplementados (◊), consumidos (€) y restantes (Ο) en el medio. En el caso de la producción de biomasa probiótica y bacteriocinas, se han realizado pocos aportes en esta área, y esto se debe en gran medida a la dificultad para identificar las variables relevantes en el sistema del que dependen ambas producciones.

Un modelo para la producción de nisina sobre medio 0.5 MH
Por un lado, la producción de biomasa depende del aporte de nitrógeno, que en el caso del Lc 1,04 actúa como nutriente limitante. Por otro lado, la producción de nisina se ve afectada negativamente por valores de pH en el cultivo por debajo del rango de 4-4,5. NUEVAS PERSPECTIVAS SOBRE SEGURIDAD ALIMENTARIA 187 7.2: Un modelo para la producción de pediocina en medio 0,5 MH.
Un modelo para la producción de bacteriocinas sobre suero de leche Sobre este medio los modelos utilizados para describir la producción
Para construir un modelo para la producción de pediocina, se asumió, al igual que para la nisina, que el sistema parecía estar regulado de manera “primaria” por el fósforo que afecta la tasa de crecimiento y por valores de pH superiores a 4, 5 que ralentizan su síntesis (figuras 15 y 17). La aplicación de ambos modelos a los resultados experimentales de biomasa y pediocina obtenidos en medio 0.5 MH (Figuras 15 y 19) también fue satisfactoria (Figuras 26 A y B), confirmando la naturaleza primaria de este antibiótico (b< En el caso de la nisina (Figura 28), llama la atención el hecho, similar a la solución de Nisaplin, de que los extractos de diferentes medios de cultivo fueron menos sensibles a la papaína que a las otras enzimas. Superficies de respuesta que muestran el efecto de la temperatura y el pH sobre las estabilidades de nisaplina, nisina y pediocina producidas por Lc 1.04 y Pc 1.02. Los resultados (Figura 30) de las isotermas de nisina para diversas temperaturas y materiales muestran un comportamiento diferente. Uno de los problemas de proporcionar bacterias probióticas a los animales es garantizar que se absorban en una cantidad que garantice un efecto beneficioso. Así, el pienso elaborado con preparados de las dos especies probióticas (Lc 1.02 y Pc 1.04) se almacenó en las condiciones habituales de almacenamiento, evaluándose semanalmente la viabilidad de las bacterias lácticas mediante la determinación del número de unidades formadoras de células. colonias (UFC). Como se puede observar (Figura 32), tanto Lc 1.04 como Pc 1.02 mostraron una alta viabilidad en el tiempo, superando en ambos casos el 75% de la población inicial. NUEVAS PERSPECTIVAS SOBRE SEGURIDAD ALIMENTARIA 200 se consideraron comunes en lechones y en ningún caso superaron niveles de significancia. Sin embargo, cabe resaltar que para todos los tiempos de muestreo, el lote tratado con el antibiótico es el que logró una reducción más intensa en comparación con el control y el lote tratado con Lc 1.04 (Figura 33, parte superior). Sin embargo, los resultados obtenidos parecen confirmar que la presencia de Pc 1.04 en el pienso provoca cambios en el desarrollo de recuentos de coliformes similares a los del antibiótico (Figura 33, abajo).
Estabilidad frente a las proteasas

Aplicación de probióticos en la alimentación de lechones
Efecto de la alimentación con probióticos sobre la flora intestinal de lechones