30 Cinética de la expresión del plásmido miostatina-1 en células de fibroblastos de ratón (3T3-L1) desde 1 hora hasta 24 horas después de la transfección. La inyección de dsiRNA-mstn-1 in vivo redujo los niveles de expresión de mstn-1 en los músculos del hongo juvenil, generando hipertrofia de las fibras musculares.
Características estructurales del tejido muscular en peces …
Representación de la estructura del tejido muscular en Lutjanus guttatus (tomado de Torres-Velarde et al., 2016). Tipos de tejido muscular: músculo rojo (lento) con alta actividad mitocondrial, músculo intermedio con actividad mitocondrial moderada y músculo blanco (rápido) con baja actividad mitocondrial (tomado de Torres-Velarde et al., 2016).
Origen del sistema muscular durante la etapa del desarrollo
Durante la segmentación, se ha descrito otra subpoblación de MPC, denominada precursores del músculo no adaxial, localizada en los somitas laterales posteriores que dan lugar a fibras musculares rápidas y constituyen la mayoría de las células del somita (Hirsinger et al., 2004; Currie e Ingham). 2001; Devoto et al., 1996; Ekker et al., 1992). Sin embargo, aunque tanto las fibras musculares lentas como las rápidas se derivan de progenitores comunes, sus destinos celulares están determinados a medida que las células migran hacia la extremidad (Hirsinger et al., 2004; Kardon et al., 2002; Hughes y Blau, 1992).

Crecimiento muscular post-embrionario
Mientras que el esclerotoma aparece a cierta distancia de la notocorda, lo que se refleja en la reducción de los derivados somitas. Este mecanismo representa la principal y, en algunas familias, la única forma de aumentar las fibras musculares después de la eclosión, por lo que la formación de miotubos musculares lentos se produce durante toda la vida.
Genes miogénicos en peces
Sólo cuando el tejido muscular está dañado se reanuda la formación de miotubos para reparar las fibras dañadas (Johnston, 2006; Fernandes et al., 2005). Secuencia de la expresión de genes miogénicos (MRF) durante el desarrollo del tejido del músculo esquelético (tomado de Torres-Velarde et al., 2016).
Miostatina
13 disminuyen a las 3 y 20 hpe (horas después de la eclosión), respectivamente; Finalmente, tanto mstn-1 como mstn-2 aumentan alrededor del período de metamorfosis, a los 21 dpe (días después de la eclosión), un período en el que mstn-1 es más abundante (De Santis et al., 2012). Por este motivo, se ha propuesto que la función del gen de la miostatina en peces no es exclusiva del crecimiento muscular, sino que también podría participar en otros procesos fisiológicos (Funkenstein et al., 2009).
Silenciamiento génico post transcripcional
Mecanismo del RNAi
Justificación
Hipótesis
Objetivo general
Objetivos particulares
AISLAR Y CARACTERIZAR LOS TRANSCRITOS COMPLETOS DE
Metodología
- Recolección de organismos…
- Aislamiento y caracterización in silico de mstn-1 y mstn-2
- Extracción de RNA y síntesisde cDNA…
- Cuantificación de expresión génica por PCR tiempo real (qPCR)…. 24
- Análisis estadístico.…
La calidad de las secuencias se evaluó utilizando Samtools 0.1.19 y finalmente la predicción de transcripciones de codificación se realizó con TransDecoder y Blast (Escalante-Rojas et al., 2018). Los cálculos de expresión génica se realizaron según el método comparativo ΔCT (Livak y Schmittgen, 2001), para el cual se utilizó el gen de referencia β-actina, según análisis de estabilidad realizados previamente (Torres-Velarde et al., 2015).

Resultados
- Aislamiento de genes miogénicos y atrogenes.…
- Análisis in silico del transcrito de miostatina-1.…
- Análisis in silico del transcrito de miostatina-2…
Siguiendo las condiciones y protocolos informados anteriormente para el análisis de la expresión de mstn-1 en L. Los niveles de expresión de mstn-2 son muy bajos durante el desarrollo embrionario y larvario de L.

Discusión
Alineamiento de las secuencias de nucleótidos de las regiones 5'UTR de mstn-1 en diferentes especies de teleósteos, el alineamiento finaliza en el codón de inicio (ATG). Alineamiento de las secuencias de nucleótidos de las regiones 3'UTR de mstn-1 en diferentes especies de teleósteos, el alineamiento inicia desde el codón de parada de la proteína mstn-1. Alineamiento de las secuencias de nucleótidos de las regiones 5'UTR de mstn-2 en diferentes especies de teleósteos, el alineamiento finaliza en el codón de inicio (ATG).

SOBRE-EXPRESAR LA miostatina-1 DE L. guttatus EN UNA LÍNEA
Silenciamiento por RNAi del gen de miostatina in vitro…
Con el descubrimiento del gen de la miostatina, se han desarrollado varias estrategias moleculares para manipular su expresión génica con el objetivo de mejorar el crecimiento en animales reproductores (Funkenstein et al., 2009). Ambos experimentos indicaron que la eficiencia del bloqueo depende de la secuencia de shRNA utilizada, ya que el rango de silenciamiento es muy amplio, oscilando entre el 30 y el 90% (Tripathi et al., 2012; Stewart et al., 2008). Se ha informado que la unión de un ARNip al promotor mstn promueve el silenciamiento transcripcional y que su silenciamiento induce cambios epigenéticos, provocando la remodelación del promotor; incluso la combinación de los dos mecanismos silenciadores (PTGS y TGS) produce mejores resultados (Roberts et al., 2012).
Metodología
- Diseño y construcción del plásmido de expresión de miostatina-1…
- Transfección del plásmido de miostatina-1 y proteína verde
- PCR punto final
- Expresión de GFP y miostatina-1 en células de fibroblastos de ratón
- Diseño y generación de los dsiRNAs y shRNAS para silenciar la
- Ensayos de co-transfección para lograr el silenciamiento de la
- Cultivo celular
- Cuantificación por PCR tiempo real
- Western Blot
62 La transfección del plásmido de expresión de miostatina-1 también se realizó con el polímero XFect (Clontech), de acuerdo con las instrucciones del fabricante. Finalmente, la expresión de miostatina se confirmó mediante PCR de punto final con los cebadores Lg-qPCR-F2 y Lg-qPCR-R2 (Tabla 6). 65 shRNA y 21 h después de la transfección inicial, se transfectó el vector de expresión de miostatina-1 pargo y las células se recolectaron después de 96 h para analizar los niveles de transcripción de mstn-1 mediante qPCR.

Resultados
- Diseño y construcción del plásmido de expresión de miostatina-1…
- Transfección del plásmido de miostatina-1 y proteína verde
- Expresión de GFP y miostatina-1 en células de fibroblastos de ratón
- Obtención de dsRNAs y shRNAS para silenciar la miostatina-1 de
- Ensayos de co-transfección para lograr el silenciamiento de la
PCR de punto final con cebadores Células Fish-Mstn-F2/R2 GH3 transfectadas con el plásmido de expresión Pargo mstn-1 (1, 2). Células GH3 transfectadas con plásmido de expresión GFP 4 h (A) y 24 h (B) después de la transfección con polímero Xfect. Después de 21 horas, se transfectó el plásmido de expresión mstn-1 y 3 horas más tarde se recogieron las células.

Discusión
Los resultados de este experimento fueron interesantes, ya que los niveles de mRNA de mstn-1 lograron disminuir significativamente con las dos secuencias de shRNA utilizadas, mientras que dsiRNA1025 fue la que dio mejores resultados y dsi920 no funcionó correctamente. Dado que los shRNA presentaron mejores resultados de silenciamiento en este sistema con células de mamíferos, se eligieron para evaluar la duración del efecto, determinando la expresión de mstn-1 cada 24 h a 96 h después de la transfección de los shRNA (sh915 y sh1047). , esta última vez fue eliminada del análisis debido a la observación de muerte celular. 79 aumentaron significativamente los niveles de ARNm de mstn-1 a partir de las 24 h posteriores a la transfección, manteniendo niveles bajos hasta las 72 h posteriores a la transfección.
SILENCIAR LA miostatina-1 EN CULTIVO EX VIVO DE TEJIDO
Glucocorticoides y miostatina…
La familia Foxo de factores de transcripción (1, 3 y 4) se activan directamente en presencia de GC, regulan la expresión de murf1 y atrogina-1 (Bodine y Furlow, 2015) e inhiben la síntesis de proteínas al inhibir la proteína mtor (Raptor). el cual inhibe el complejo mTOR1 (TORC1), sin afectar a TORC2 (Morris et al., 2015). Durante el proceso de atrofia muscular inducida por GC en mamíferos, se observó un aumento en los niveles de expresión de mstn, lo que se asoció con la existencia de elementos sensibles a GC en el promotor mstn de mamíferos (Schakman et al., 2008; Gilson et al., 2007 ; Ma et al., 2003; Ma et al., 2001). De manera similar, existen reportes donde mstn activa las ligasas murf1 y atrogina-1 (McFarlane et al., 2006), además de reducir los niveles de mtor, un elemento esencial en la síntesis de proteínas a través de la vía de señalización Akt/TORC1. /p70S6K (Trendelenburg et al., 2009).
Metodología
- Cultivo ex vivo
- Cinética de expresión de cultivo muscular ex vivo
- Silenciamiento de mstn-1 ensayo preliminar
- Exposición a dexametasona
- Inducción con DEX y silenciamiento de mstn-1 de L. guttatus
- Extracción de RNA y análisis de expresión génica por tiempo real
- Análisis estadístico
Las células se recolectaron 72 h después de la transfección para el análisis de la expresión génica mediante qPCR. Los cálculos de expresión génica se realizaron según el método ΔCT comparativo (Livak y Schmittgen, 2001), para el cual se utilizó la media geométrica de dos genes de referencia (β-actina y GAPDH). Secuencias de oligos utilizados para el análisis de la expresión génica en cultivos ex vivo Tm del cebador (°C) Secuencia del tamaño del producto 5'-3'.

Resultados
- Cinética de expresión de cultivo ex vivo
- Silenciamiento de mstn-1 ex vivo (ensayo preliminar)
- Dexametasona eleva los niveles de expresión de miostatina-1
- Silenciamiento de mstn-1 mediante RNAi en tejido muscular ex vivo
- La exposición a DEX regula la expresión de genes miogénicos y
- El silenciamiento de mstn-1 regula la expresión de factores de
La expresión de mstn-1 aumentó en cultivos ex vivo de tejido muscular expuesto a DEX en las dos dosis evaluadas (50 nM, 100 nM), manteniéndose niveles elevados desde las 48 horas de exposición hasta las 72 horas sin diferencias significativas entre ambas dosis (Figura 45). ). Efecto de DEX sobre los niveles de expresión de genes miogénicos durante el silenciamiento de mstn-1 por RNAi. Efecto de la dexametasona sobre los niveles de expresión atrogénica durante el silenciamiento de mstn-1 por ARNi.

Discusión
Se observaron resultados similares en ratones tratados con DEX y RU486 (antagonista de sGC), que bloquearon la activación de mstn y redujeron la atrofia muscular inducida por DEX (Ma et al., 2003). Según los autores, el silenciamiento de mstn-1 durante nuestros experimentos aumenta la expresión de los mismos genes miogénicos, lo que confirma la participación de Mstn-1 en la inhibición de la miogénesis (Yang et al., 2007; Langley et al., 2002). El silenciamiento de mstn-1 nos permitió evaluar su papel en el proceso catabólico del tejido muscular, ya que mstn-1 parece promover directamente la degradación de proteínas a través del complejo foxo3b/Smad3, seguida de la activación de la expresión de murf1; sin.

SILENCIAR LA miostatina-1 IN VIVO EN L. guttatus PARA
Estrategias de administración de siRNAs
Los liposomas son estructuras esféricas con una capa bilipídica y un núcleo acuoso donde se suspenden las moléculas de ARN; Existen modificaciones de estas estructuras donde se agregan lípidos catiónicos para promover la entrada del ARN a través de la membrana celular (Chakraborty et al., 2017). Algunos métodos para introducir ARNip, miARN o shARN en las células son inyecciones directas, electroporación, baños de inmersión y encapsulación de alimentos (Chakraborty et al., 2017; Sen y Ghatak, 2015; De Vos et al., 2014). La base de la técnica es la encapsulación de moléculas de ARNip con polímeros no tóxicos para protegerlas del proceso de digestión así como de las células del sistema inmunológico hasta llegar al tejido diana (Elzoghby et al., 2013; Xu). et al., 2012).
Silenciamiento in vivo de miostatina-1
106 La entrega de moléculas de ARN in vivo no es una tarea sencilla. Además de seleccionar el vehículo y la vía de administración correcta, es importante considerar otros aspectos para que se consiga el efecto biológico esperado, como determinar las concentraciones y dosis adecuadas, y evaluar los posibles efectos secundarios, tanto del vehículo como del las moléculas de ARN, especialmente porque la investigación sobre la formulación y estabilidad de vehículos utilizando materiales a escala nanométrica aún es escasa (Huang et al., 2017). Después de siete semanas de administración, no se observaron diferencias en el peso de los organismos tratados en comparación con los controles, pero sí una disminución en los niveles del transcrito mstn (Terova et al., 2013).
Metodología
- Cultivo de peces
- Diseño experimental
- Inyección de dsiRNA mstn-1
- Extracción de RNA y síntesis de cDNA
- Cuantificación de expresión génica por qPCR
- Análisis histológicos
- Análisis estadísticos
Se aplicó un ANOVA de dos vías para determinar el efecto del tratamiento (dsiRNA, scRNA, JetPei y Salina) y el tiempo posterior a la inyección (48 horas, 5 y 60 días) sobre los niveles de expresión de mstn-1, con múltiples comparaciones ejecutadas (Fisher) . en caso de encontrar diferencias significativas (P<0,05). En el análisis histológico se realizó la prueba de Kruskal-Wallis y comparaciones múltiples de Dunn (P<0,05) para comparar el área de los haces musculares; mientras que para analizar las diferencias en el número de haces de músculos por campo se aplicó un ANOVA unidireccional y comparaciones múltiples con Fisher (P < 0,05). Para el análisis de ganancia de peso y talla se utilizó ANOVA unidireccional y comparaciones múltiples de Fisher (P < 0,05); mientras que la correlación entre mstn-1, peso total, longitud total, aumento de peso y altura se evaluó mediante el coeficiente de Spearman (P <0,05).
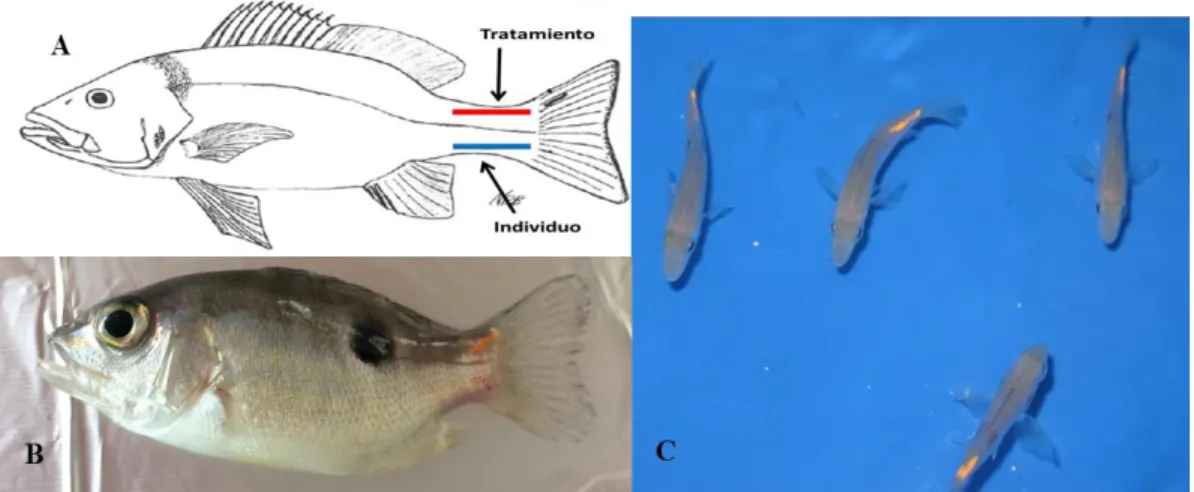
Resultados
- Silenciamiento de mstn-1 in vivo en tejido muscular de L.guttatus…
- Análisis histológicos
- Evaluación del crecimiento peso-talla
- Relación mstn-1, peso y talla
Aumento de peso (a) y aumento de altura (b) durante el experimento de silenciamiento in vivo en los períodos 0-5, 5-30 y 30-60 ppp y durante todo el experimento (0-60 ppp); dsiRNA corresponde a secuencias que bloquean mstn-1 inyectado con JetPei; Los scRNA corresponden a secuencias de control inyectadas con JetPei;. JetPei corresponde al control del polímero utilizado como vehículo, y la solución salina corresponde al control inyectado con solución salina sola. Peso y tamaño final a 60 dpi, equivalente a 145 dpe; dsiRNA corresponde a secuencias que bloquean mstn-1 inyectado con JetPei; Los scRNA corresponden a secuencias de control inyectadas con JetPei; JetPei corresponde al control del polímero utilizado como vehículo, y la solución salina corresponde al control inyectado con solución salina sola.

Discusión
Ambas secuencias seleccionadas (shRNA y dsiRNA) lograron bloquear el transcrito de mstn-1 72 h después de la transfección en cultivos ex vivo. Los resultados anteriores nos llevaron a elegir la secuencia de dsiRNA (dsi1025) para realizar el silenciamiento de mstn-1 in vivo. La inyección local de dsiRNA en tejido muscular redujo significativamente el mRNA de mstn-1 durante 120 h después de la administración, generando un fenotipo de hipertrofia muscular.

Claves de acceso a gen bank para secuencias utilizadas en análisis
Factor de condición e Índice hepatosomático
No se encontraron diferencias entre los tratamientos en términos de HI de los peces a 48 hpi (figura) y 60 dpi (figura). Sin embargo, a 5 ppp, los peces inyectados con el vehículo JetPei aumentaron significativamente su HI (Figura 64), lo que indica cambios en el metabolismo y las reservas de energía de los peces (Schmitt y Dethloff, 200).

Protocolos
Retire los filetes de músculo con una hoja de bisturí esterilizada y coloque los trozos en placas de Petri de 10 cm con medio de cultivo (2% de antibióticos). Mantenga la placa de Petri cubierta cuando esté fuera de la campana de cultivo. En una campana de cultivo, transfiera el tejido muscular a otra placa de Petri con medio de cultivo limpio.
Productos derivados de la tesis
Keywords: Muscle atrophy; Protein degradation, Muscle development, Fish, Ex vivo culture Underlined: Presenting author [email protected]. Successfully received: submission Mechanisms of stress-related muscle atrophy in fish: an ex vivo approach to Mechanisms of Development. Title: Mechanisms of stress-related muscle atrophy in fish: an ex vivo approach Journal: Mechanisms of Development.






