POLYAMINES CONTRIBUTION TO THE IMPROVEMENT OF CROP PLANTS 1
TOLERANCE TO ABIOTIC STRESS 2
Menéndez1-3, Ana Bernardina#; Rodriguez1, Andrés Alberto; Maiale1, Santiago 3
Javier; Rodriguez Kessler 4, Margarita; Jimenez Bremont2, Juan Francisco and 4
Ruiz1, Oscar Adolfo 5
1 IIB-INTECh (CONICET-UNSAM). Chascomús. Buenos Aires. Argentina.
6
2 Division de Biologia Molecular. Instituto Potosino de Investigacion Cientifica y 7
Tecnologica (IPICYT). San Luis Potosi, S.L.P. México.
8
3 Department of Biodiversity and Experimental Biology, Faculty of Sciences, 9
University of Buenos Aires (DBBE, FCEN, UBA).
10
4 Facultad de Ciencias. Universidad Autónoma de San Luis Potosí(UASLP).
11
San Luis Potosí, S.L.P. México.
12
# Corresponding author 13
14
ABSTRACT 15
Plant development and productivity are negatively regulated by environmental 16
stresses. The loss of productivity is triggered by a series of morphological, 17
physiological, biochemical and molecular stress-induced changes. The 18
development of diverse strategies to obtain stress-tolerant plants are currently 19
one of the most active fields in plant research, which is expected to help 20
preventing the dramatic reduction in crop yields due to global changing effects.
21
Therefore, the identification of stress-regulatory genes and signaling molecules 22
involved in the process of stress tolerance should allow the development of 23
novel strategies to obtain tolerant plants.
24
Polyamines (PAs) are polycationic compounds with a recognized role in plant 25
growth and development, as well as in abiotic and biotic stress responses. In 26
this chapter, we review and discuss the information concerning the 27
modifications in polyamines levels in response to drought, salinity and cold 28
stresses, focusing on crop species.
29
The comparison of common and specific responses in different crop plants 30
suggests the view that polyamines actively participate in stress signaling 31
through an intricate metabolic network. However, the precise mechanism(s) of 32
action by which PAs could protect crop plants from challenging environmental 33
conditions remains unclear.
34 35
INTRODUCTION 36
Polyamines (PAs) are aliphatic biogenic amines present in most Prokaryotes 37
and all Eukaryotic organisms (Takahashi and Kakehi, 2010; Fuell et al., 2010).
38
These small molecules are essential for life. At physiological pH, PAs are found 39
as protonated, positively charged molecules containing two (diamine), three 40
(triamine) or four (tetraamine) amine groups, what favors their electrostatic 41
interaction with several macromolecules such as nucleic acids, proteins and 42
lipids (Igarashi and Kashiwagi, 2000; Childs et al., 2003). The polycationic 43
nature of PAs is one of the most important properties linking these natural 44
compounds to several cellular and physiological processes, and new 45
connections between PAs and other molecules, revealing new insights into the 46
PA biological role are being continually discovered.
47
At the cellular level, PAs participate in diverse fundamental processes such as 48
transcription, translation, DNA replication, chromatin condensation, cell 49
signaling, cell division and differentiation, senescence and cell death. In 50
addition, diverse roles in membrane stabilization, ion channel regulation, cation- 51
anion balance, modulation of enzyme activities, and protein modification have 52
been also described (Childs et al., 2003; Shabala et al., 2007; Handa and 53
Mattoo, 2010).
54
In plants, PAs are present in micromolar (10 μM) up to millimolar 55
concentrations (Galston and Sawhney, 1990). The most common PAs are 56
spermidine (Spd; NH2(CH2)3NH(CH2)4NH2), spermine (Spm;
57
NH2(CH2)3NH(CH2)4NH(CH2)3NH2) and their obligate precursor putrescine (Put;
58
NH2(CH2)4NH2). Spd is structurally an unsymmetrical molecule that can be 59
aminopropylated at each end, forming either Spm or thermospermine (tSpm) 60
(Knott et al., 2007). In plants, PAs distribution differs among tissues and 61
developmental stages, being Put and Spd more abundant than Spm and tSpm 62
(Naka et al., 2010). Also, tSpm seems to be present in all plants, while Spm 63
appears to be restricted to flowering plants (Fuell et al., 2010).
64
Besides Put, Spd and Spm, less common PAs have been described in plants 65
such as cadaverine (Cad), norspermidine, norspermine, homocaldopentamine, 66
homocaldohexamine, 1,3-diaminopropane and 4-aminobutylcadaverine, among 67
others (Kuehn et al., 1990; Fujihara et al., 1995; Kuznetsov et al., 2007).
68
Although their concentrations in plants are much higher than those of 69
phytohormones, plant PAs are considered as growth regulators, since they play 70
fundamental roles in a wide range of growth, differentiation and morphogenetic 71
processes during the course of plant ontogeny. Roles in embryogenesis, seed 72
germination, rhizogenesis, organogenesis, floral initiation and development, as 73
well as in vascular development, leaf senescence, fruit development and 74
ripening have been described for these molecules (Slocum, 1991; Kakkar et al., 75
2000; Kakkar y Sawhney, 2002; Pang et al., 2007). Lately, a great deal of 76
attention has been paid to the protective effect of PAs during plant response to 77
biotic and abiotic stresses (Liu et al., 2007; Gill and Tuteja, 2010; Vera-Sirera et 78
al., 2010).
79
Polyamine biosynthesis 80
Intracellular PA concentration is tightly regulated through their biosynthesis and 81
catabolism, and modulated by cellular transport and conjugation with other 82
organic molecules such as hydroxycinnamic acids and proteins (Bagni and 83
Tassoni, 2001; Edreva et al., 2007; Fincato et al., 2011). Polyamines can be 84
found as conjugated forms, eg: covalently attached to compounds of low 85
molecular weight, (typically hydroxycinnamic, p-coumaric, caffeic and ferulic 86
acids) and high molecular weight molecules (proteins or cell wall polymers).
87
Enzymes such as putrescine-cafeoil-CoA transferase are responsible for the 88
formation of hydroxycinnamic acid conjugates (Martin-Tanguy, 1997), phenolics 89
compounds that are related to the flowering process and the plant response to 90
pathogen attack (Flores and Martin-Tanguy, 1991; Martin-Tanguy, 1997). On 91
other hand, transglutaminases channel the conjugation of PAs to the γ- 92
carboxamide group of endo-glutamic residues of proteins, especially in the 93
chloroplast, where this activity is stimulated by light (Del Duca et al. 1995;
94
Dondini et al., 2003). In addition, the compartmentalization of enzymes involved 95
in PA metabolism suggests a spatio-specific regulation of these important 96
amines (Borrel et al., 1995; Kamada-Nobusada et al., 2008; Fincato et al., 97
2011).
98
The first step in PA biosynthesis is the diamine Put formation. In plants and 99
some bacteria, this process occurs by decarboxylation of arginine via arginine 100
decarboxylase (ADC; EC 4.1.1.19) in a pathway involving agmatine and N- 101
carbamoylputrescine as intermediates, and the corresponding enzymes 102
agmatine iminohydrolase (EC 3.5.3.12) and N-carbamoylputrescine 103
amidohydrolase (EC 3.5.1.53) (Figure 1). The ADC pathway for Put 104
biosynthesis in plants appears to be derived from endosymbiotic gene transfer 105
between the cyanobacterium precursor of chloroplasts and the eukaryotic 106
nucleus (Illingworth et al., 2003). In animals, fungi, and also in most plants, Put 107
is synthesized directly from ornithine via the cytosolic ornithine decarboxylase 108
(ODC; EC 4.1.1.17). Evolutionary compartmentalization of Put biosynthesis in 109
chloroplasts is accomplished by ADC signal sequences that import this enzyme 110
into the plastid (Borell et al., 1995; Illingworth et al., 2003). Both ODC and ADC 111
enzymes use pyridoxal 5’-phosphate as cofactor.
112
The higher PAs, Spd and Spm are synthesized from Put through the successive 113
activities of Spd synthase (SPDS; EC 2.5.1.16) and Spm synthase (SPMS; EC 114
2.5.1.22) through the addition of aminopropyl groups. In addition, tSpm is also 115
synthesized from Spd (an asymmetric molecule that allows the formation of two 116
isomers, Spm or tSpm, respectively), trough the activity of a thermospermine 117
synthase (tSPMS; Knott et al., 2007). The aminopropyl moiety is derived from 118
methionine, which is first converted into S-adenosylmethionine (SAM) and then 119
decarboxylated via S-adenosylmethionine decarboxylase (SAMDC; EC 120
4.1.1.50). SAMDC is considered the mayor regulatory enzyme involved in 121
higher PA biosynthesis and plays an essential role in modulating ethylene 122
production in plants, since the precursor of ethylene 1-aminocyclopropane-1- 123
carboxylic acid is also derived from SAM (Bagni and Tassoni, 2001). However, 124
Del Duca et al., (1995) and Tassoni et al. (2000) provided data showing the 125
occurrence of a back-conversion pathway: Spd added to Helianthus tuberosus 126
chloroplasts and Arabidopsis plants, respectively, was converted to Put 127
Recently, polyamine oxidase enzymes involved in back-conversion pathway in 128
Arabidopsis have been identified (Moschou et al., 2008).
129
130
Subcellular localization and transport 131
Polyamines are present in all cell compartments and may be specially detected 132
in actively growing tissues where cell division or elongation takes place.
133
Cytochemical, immunochemical, autoradiographic and subcellular fractions 134
studies suggest that the largest PAs reservoirs in plants are the cell wall and the 135
vacuole (Bagni and Pistocchi, 1991, Mariani et al. 1989; Slocum, 1991). In 136
addition, PAs have been found in the cytoplasm, nucleus, plasma membrane, 137
mitochondria and chloroplasts. In the latter compartment, PAs are associated 138
with components of the electron transport chain, by both electrostatic and 139
covalent interactions (Bagni and Pistocchi 1991; Kotzabasis et al., 1993, 140
Torrigiani et al. 1986; Votyakova et al., 1999). The information regarding sub- 141
cellular localizations of enzymes involved in plant polyamine metabolism in 142
plants is scarce. Immunocytochemical and bioinformatics studies indicate that 143
ADC is mainly present in the chloroplast and to a lesser extent in the nucleus 144
(Borrel et al., 1995, Bortolotti et al., 2004, Illingworth et al., 2003). Inhibitor 145
binding and fractionation studies suggest that ODC is located in the cytoplasm 146
and nucleus (Slocum, 1991). In plants of A. thaliana, ODC activity (but no 147
homologous gene) was reported in plastid membrane (Tassoni et al., 2003). In 148
contrast, SPDS and SAMDC activities are generally located in the cytoplasm 149
(Slocum, 1991), whereas there is no information on SPMS. As mentioned 150
above, Cu+2- and flavin oxidases occur predominantly in the apoplast, although 151
it has also been suggested a cytoplasmic and vacuolar localization (Cervelli et 152
al. 2004; Cona et al., 2003).
153
Using cell cultures, protoplasts and petals as models, it has been shown that 154
the transport of aliphatic amines through the plasma membrane of plant cells is 155
bidirectional, saturable, energy dependent and under hormonal control, at least 156
for auxins and cytokinins (Bagni and Pistocchi, 1991). It has also been 157
demonstrated that the transport of Put in maize roots is non-competitively 158
inhibited by inorganic cations (Ca+2, Mg+2) and Spm. On the other hand, the 159
existence of at least two transport systems, one for diamines and another for 160
polyamines has been put forward (Di Tomaso et al., 1992, Hart et al., 1992). It 161
was also proposed that the interaction between polyamines and membranes 162
would arbitrate important cellular events, such as receptor-mediated signal 163
transmition. In E. coli, several periplasmic proteins that bind polyamines are 164
known such as PotD and PotF, which are part of two transmembrane transport 165
systems (pPT104 and pPT79) and bind Spd and Put, respectively (Sugiyama et 166
al. 1996; Vassylyev et al., 1998). In vascular plants like zucchini (Cucurbita 167
pepo) and maize (Zea mays), plasma membrane proteins that specifically bind 168
Spd have been identified, purified and analyzed (Tassoni et al., 1996; Tassoni 169
et al., 2002). Despite being slightly mobile cations, due to their strong 170
interaction with cell wall components, the distant translocation of PAs through 171
xylematic and phloematic conducts has been demonstrated (Antognoni et al.
172
1998; Bagni and Pistocchi, 1991; Caffaro et al., 1994).
173
174
Polyamine catabolism 175
Polyamines may be deaminated by oxidation, which constitutes the main PA 176
catabolic pathway (Federico and Angelini, 1991). Recent data on PA oxidation 177
in plants has led to propose the several possible functions that this pathway 178
could fulfill (reviewed by Kusano et al., 2008). In plants PAs catabolism 179
proceeds via Cu2+-oxidases (Fig. 2; diaminoxidase: DAO, EC 1.4.3.6) and 180
flavin-oxidases (poliaminoxidase: PAO, EC 1.4.3.4), present in the apoplastic 181
and peroxisomal compartments (Medda et al., 1995; Sebela et al., 2001). In the 182
apoplast, Put, Spm, and Spd are oxidized to 1,3-diaminopropane, H2O2, and the 183
corresponding aldehyde, while in the peroxisome, Spm is converted to Put, via 184
the intermediate Spd (Rea et al., 2004; Cona et al., 2006; Moschou et al., 185
2008). The Cu+2- amino oxidase oxidizes the primary amine of diamines and 186
polyamines, with the concomitant production of H2O2, NH4+
and the 187
corresponding aldehyde, while the flavin oxidase enzyme oxidizes the 188
secondary amino of Spd and Spm, producing H2O2, 1, 3-diaminopropane and 189
the corresponding amino aldehyde. Also, both mono- and dicotyledonous 190
species may catabolize Put to γ-aminobutyric acid (GABA), an important 191
modulator of several physiological proccessess (Bouchereau et al., 1999).
192
Aside from their participation in the catabolism and their contribution to cellular 193
homeostasis, amine oxidases take part in important physiological events 194
through their reaction products, mainly H2O2. On one hand, they are related to 195
lignin biosynthesis and crosslinking reactions of the cell wall, occurring during 196
xylem maturation and plant cell elongation (Cona et al. 2003; Federico and 197
Angelini, 1991; Laurenzi et al. 1999; Laurenzi et al. 2001; Moller and 198
McPherson, 1998) as well as to the cell wall strengthening that take place 199
during the infection by plant pathogens (Cona et al., 2006). On the other hand, 200
they regulate cell PAs level in plants subjected to stress conditions (Bagni and 201
Tassoni, 2001; Cona et al., 2006). For example, a steep decline of Spd and 202
Spm observed in rice (Oryza sativa) plants, 6 days after imposition of drought 203
stress was assigned to PAO amino oxidation (Capell et al., 2004). It has been 204
also reported that ethylene promotes DAOs and PAOs activities, which could be 205
related to reductions in PAs contents observed in several crop species (Li et al., 206
2004).
207
208
Involvement of PAs on crop plant response to drought, salt and cold 209
stresses 210
Drought stress 211
Polyamines are thought to play protective roles during water deficit/drought 212
stress. These molecules may act as osmolytes and bind non-covalently to the 213
negatively charged groups of membrane phospholipids, thus contributing to the 214
stabilization of membrane conformation (Hanzawa et al., 2000). They also 215
regulate pH alterations due to osmotic stress, by reversing H+-ATPase and H+- 216
PPase inactivity. Table 1 shows main results obtained in diverse studies on 217
polyamine metabolism in several crop species cultivated under water deficit- 218
related conditions. In barley (Hordeum vulgare), a close relationship between 219
PAs metabolism and leaf turgor was found, when leaf turgor was maintained at 220
control level, Put and Spm accumulated, whereas the levels of both PAs sharply 221
diminished when leaf turgor was lost (Turner and Stewart, 1986). Moreover, 222
Flores and Galston (1984) postulated that turgor maintenance is a requirement 223
for PAs accumulation during drought stress.
224
Put accumulation and ADC activation occur under unfavorable conditions 225
(Bouchereau et al., 1999). However, it has been often observed that drought 226
stress induces the selective accumulation of Put in drought-sensitive plant 227
species, whereas in drought-tolerant ones, PAs metabolism shifts towards Spd 228
and Spm biosynthesis. When wheat (Triticum aestivum) plants were water 229
stressed with polyethylene-glycol (PEG 6000), increased levels of free-Spd and 230
free-Spm levels in leaves of a drought-tolerant cultivar were observed, whereas 231
free-Put titer was built up in a drought-sensitive cultivar of that crop (Liu et al., 232
2004). Yang et al. (2007) tested whether rice polyamines were involved in 233
drought tolerance. Six rice cultivars differing in drought tolerance were 234
subjected to well watered and water-stressed treatments during their 235
reproductive period. Water stress increased ADC, SAMDC, and SPDS activities 236
in rice leaves, in consonance with rises observed in Put, Spd, and Spm levels.
237
The augmented contents of the higher PAs (Spd and Spm) under water stress 238
were significantly correlated with drought tolerance. The authors concluded that 239
incremented levels, higher PAs and insoluble-conjugated Put, as well as early 240
accumulation of free PAs during drought is a desirable physiological trait for rice 241
during its adaptation to this stress. Basu et al. (2010) compared differential 242
biochemical responses of the salt-sensitive (IR-29), salt-tolerant (Pokkali) and 243
aromatic (Pusa Basmati) rice varieties during PEG-induced dehydration stress.
244
They found that drought resistant cultivars had higher free Spd and Spm in the 245
leaves than drought-susceptible ones during the whole period of water 246
withholding. Moreover, stressed Pokkali rice plants appeared to suffer lesser 247
damages, in parallel with the maximum accumulation level of the higher PAs, 248
Spd and Spm.
249
The effect of water stress has also been studied in several non-gramineous 250
species. In 15-day-old chickpea (Cicer arietinum) seedlings, increment of Spd 251
and Spm levels were observed after exposure to osmotic stress created by 252
PEG 6000 during 4 days (Nayyar and Chander, 2004). Vetiver grass (Vetiveria 253
zizanioides), is considered to have future potential as a source of bio-fuel and 254
cellulosic ethanol (Paul et al., 2008). Zhou and Yu (2010) studied the variations 255
of PA contents in plants of this species when stressed with 20, 40 and 60%
256
PEG solutions during 6 days. Their results showed that under osmotic stress 257
free and conjugated Put decreased, whereas free and conjugated Spd and Spm 258
amounts increase. Lei (2008) used Populus przewalskii as a tree model species 259
to investigate the acclimation and adaptation to drought stress, in particular the 260
ROS damaging effects and their scavenging systems. P. przewalskii plants 261
subjected to water withholding treatment showed reduced biomass, shoot 262
height and basal diameter. Drought stress also increased Put and Spd, while 263
little change was observed in the Spm level (Lei, 2008). Cacao (Theobroma 264
cacao) plants subjected to 10 days of drought treatment showed augmented 265
Put, Spd and Spm in leaves (Bae et al., 2008). Also, a correlation was found 266
between enhanced expression of TcODC and TcSAMDC genes with changes in 267
leaf water potential. These expressions were preceded by induction at 7 days of 268
TcADC and TcSAMDC genes in roots. In leaves of this species TcSPDS and 269
TcSPMS genes were not responsive to drought, but the expressions of these 270
genes were slightly up-regulated in stressed roots. Authors speculated that 271
since PAs are associated with root development, it is possible that the induction 272
of PA biosynthesis genes in roots were involved in shifting the root architecture 273
of cacao plants in response to stress, as it was suggested in the subantartic 274
cruciferous species Pringlea antiscorbutica (Hummel et al., 2002). PAs 275
involvement in the development and ripening of fruit emerges from variations 276
observed in their levels in several fruit crops (Serrano et al. 1995; Ponappa and 277
Miller 1996; Geny et al. 1999, Shiozaki et al. 2000). Antolín et al. (2008) 278
investigated how the balances of PAs were affected by drought deficit irrigation 279
of grapevine (Vitis vinifera). The PAs level was analyzed at distinct stages of 280
berry ripening: onset of veraison, middle veraison and harvest. Their data 281
showed that at the onset of veraison, berry PAs concentrations were higher in 282
deficit irrigation treatments than in control grapevines, although those 283
differences disappeared during ripening. Toumi et al. (2010) reported that total 284
PAs were significantly lower in control tolerant grapevines and higher in control 285
sensitive genotypes, and these titers respectively increased and decreased 286
after drought treatment. Water withholding treatments applied to pepper 287
(Capsicum annuum L.) seedlings during one week resulted in elevated levels of 288
Cad and Put in leaves (Sziderics et al., 2010), whereas PA concentration was 289
reduced in roots. Authors suggested that PAs might be involved in the stress 290
protection of pepper leaves rather than in the osmotic adaptation to drought. On 291
other hand, because proline may be synthesized from ornithine via ornithine 292
aminotransferase (Delauney and Verma 1993), thus competing for the substrate 293
(ornithine) with the PA biosynthetic pathway (Theiss et al. 2002), reduced levels 294
of Put, Spd and Spm could be a consequence of a preferential proline synthesis 295
in roots.
296
Up-and down-regulation of genes involved in PAs metabolism provoked by 297
drought stress has been studied in several crops. In a large-scale study on 298
changes in transcript abundance (Öztürk et al., 2002), drought-induced 299
transcripts of two ADC genes were detected in leaves and roots of barley plants 300
subjected to water deficit. By means of a microarray analysis, decreased of the 301
SAMDC2 transcript (TM00041253, Tian et al. 2004) was observed in the 302
reproductive organs of maize (Zea mays L.), at an early stage of water deficit 303
(Zhuang et al., 2008). With the aim of identifying drought-responsive genes and 304
compounds in potato, Evers et al. (2010) analyzed transcriptomic and targeted 305
metabolite of two potato clones (Solanum tuberosum L.) of the Andean cultivar 306
group, Sullu and SS2613. These clones presented different drought-tolerance 307
phenotypes, as exposed to a continuously increasing drought stress in a field 308
trial. Upon drought, genes encoding for PAs biosynthesis ADC and SAMDC 309
enzymes were upregulated in both clones. In grape seedlings grown in vitro, 310
VvADC and VvSPMS inductions were observed within one week, after 350 mM 311
mannitol treatment (Liu et al., 2011).
312
Exogenous PAs addition to plants has early called the attention of several 313
researchers since the observed growth promotion effect resembled those of 314
phytohormones (Rastogi and Davies, 1991). The binding of Spd and Spm to 315
proteins or to nucleic acids protects them from degradation and provide them 316
with a higher conformational stability under stress conditions. The exogenous 317
application of Spd to osmotically stressed oat (Avena sativa) plants stabilized 318
the structure of thylakoid proteins Dl and D2, cytochromes and Rubisco 319
(Tiburcio et al., 1994; Besford et al. 1993). When treated with Spd, water- 320
stressed cucumber (Cucumis sativus L.) seedlings showed enhanced guaiacol 321
peroxidase activity and a reduction of superoxide dismutase and catalase 322
activities compared to untreated, water-stressed ones (Kubis, 2008). The author 323
suggested that PAs are able to moderate the activity of scavenging system 324
enzymes and influence the oxidative stress intensity. Likewise, exogenously 325
applied PAs increased drought tolerance of rice by improving leaf water status, 326
photosynthesis and membrane properties (Farooq, et al., 2009). Recently, in 327
vitro Citrus plants pre-treated with Spm showed improved tolerance to 328
dehydration stress through less water loss and lower electrolyte leakage (Shi et 329
al., 2010). Pretreatment with Spm led to higher endogenous PAs content and 330
the activation of antioxidant enzymes. Authors assigned the reduced water loss 331
to increased stomatal closure. On other hand, they attributed the lower 332
electrolyte leakage to inhibition of lipid peroxidation and biomembranes 333
stabilization due to diminution of ROS levels. The addition of Spm led to 334
drought-stressed Pinus strobus seedlings to sustain higher photosynthesis and 335
lower transpiration rates (Islam et al. 2003).
336
There is an amount of evidence supporting the influence of PAs on membrane- 337
associated enzymes activities (Srivastava and Rajbabu, 1983; Reggiani et al., 338
1992). Reggiani et al. (1992) reported that plasma membrane H+-ATPase from 339
rice coleoptiles is activated by PAs. Under osmotic stress, less membrane 340
peroxidation, greater H+-ATPase activity and reduced senescence were 341
registered in honey brew (Cucumis melo L.) supplemented with exogenous Spd 342
or Spm, compared with the corresponding control without PAs (Lester, 2000).
343
Treatment with PEG brought about significantly higher increments of 344
conjugated-Spd and-Spm contents, and H+-ATPase activity in root plasma 345
membranes of a drought-tolerant than those found in a drought-sensitive wheat 346
cultivar (Liu et al., 2004). In addition, exogenously added Spd alleviated osmotic 347
stress injury in drought-sensitive seedlings, in parallel with a huge enhancement 348
in the root plasma membrane H+-ATPase activity. Later, it was shown that 349
treatment with methylglyoxayl-bis (guanylhydrazone) (MGBG), an inhibitor of 350
SAMDC, aggravated PEG injury to drought-tolerant seedlings, with a 351
concomitant reduction of the root PM-H+-ATPase activity (Liu et al., 2005).
352
These results pointed out a possible involvement of these PAs in PM-H+- 353
ATPase activity and water stress tolerance of wheat seedlings.
354
The over-expression of genes encoding enzymes that mediate in diverse 355
pathways of PAs anabolism has become a promising approach for obtaining 356
transgenic plants with higher drought stress tolerance. The introduction of a 357
human SAMDC gene under the control of a constitutive promoter (CaMV35S) in 358
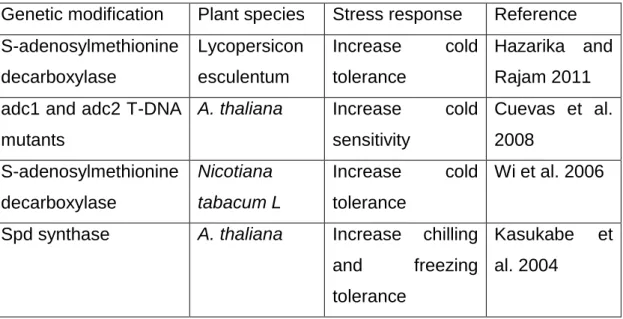
tobacco (Nicotiana tabacum var. xanthi) led to increased conjugated Spd and 359
Put titers and improved drought tolerance (PEG 20,000), as well as to other 360
abiotic and biotic stresses (Waie and Rajam, 2003). Sweet potato (Ipomoea 361
batatas- cv. Kokei 14) plants transformed with the Cucurbita ficifolia-derived 362
SPDS gene (FSPDS1), doubled their Spd content and produced higher storage 363
tissue biomass, compared with the wild-type (Kasukabe et al., 2006). In 364
addition, transgenic plants were more tolerant to paraquat (a powerful oxidative 365
stress inducer) than the wild-type, suggesting that the observed improved 366
tolerance may be in part due to enhanced oxidative stress tolerance.
367
Previously, Capell et al., (2004) generated transgenic rice plants expressing the 368
Datura stramonium ADC gene and evaluated their response to drought stress.
369
They observed that wild-type plants responded to the onset of drought stress by 370
increasing endogenous Put levels, but not those of Spd and Spm. In contrast, 371
transgenic plants expressing D. stramonium ADC showed improved drought 372
tolerance, in parallel with much higher levels of Put, what led to increased Spd 373
and Spm synthesis.
374
Prabhavathi and Rajam (2007), introduced in eggplants (Solanum melongena) 375
the ADC gene under the control of the constitutive promoter CaMV35S.
376
Transgenic seedlings of this crop showed enhanced PAs level due to the 377
augmented ADC activity, and also higher DAO activity. PAs-accumulating 378
transgenic eggplants exhibited an augmented tolerance level to drought 379
imposed through 7.5 and 10% PEG-20,000, among other abiotic and biotic 380
stresses. Several lines of a transgenic European pear (Pyrus communis L.
381
‘Ballad’) over-expressing the gene encoding for the apple SPDS (MdSPDS1) 382
were created by Agrobacterium-mediated transformation and tested for 383
tolerance to osmotic stress (300 mM mannitol, Wen et al., 2008). The 384
transgenic line having the highest Spd accumulation and expression level of 385
MdSPDS1, showed the strongest tolerance to this stress. Ten days after 386
mannitol treatment, a slight decrease in Put, and significant enhancements of 387
Spd (33%) and Spm titers, and (Spd + Spm)/Put ratio were observed in the 388
transgenic line, compared with the wild type. Later, He et al. (2008) showed that 389
the transgenic line expressing the MdSPDS1 contained superior antioxidant 390
enzyme activities, and lower levels of malondialdehyde and H2O2 than the wild- 391
type, suggesting that transgenic plants are more tolerant. In order to dissect the 392
roles of Put from the higher PAs (Spd and Spm), Peremarti et al. (2009) 393
generated transgenic rice plants constitutively expressing an heterologous 394
SAMDC gene from D. stramonium, so that the levels of higher PAs were 395
increased without affecting Put levels. These transgenic lines were able to 396
recover from drought stress in comparison to control plants. Hazarika and 397
Rajam (2011) generated transgenic tomato (Lycopersicon esculentum Mill.) 398
plants with the human SAMDC gene, and evaluated the transgenic plants for 399
tolerance to drought, among other stresses. Transgenic plants presented higher 400
PAs levels and improved tolerance against drought, with respect to 401
untransformed, control plants. In turn, transcription factors may influence PAs- 402
mediated adaptation to a variety of abiotic stresses (Chen et al. 2002). It was 403
shown that over-expression of CaPF1 (a C. annuum pathogen and freezing 404
tolerance-related protein) in transgenic tissue of eastern white pine (Pinus 405
strobus L.), prevented the decrease of PAs and resulted in a dramatic increase 406
in tolerance to drought, freezing and salt stress (Tang et al., 2007). These 407
authors suggested that CaPF1 may influence, by a so far unknown mechanism 408
on PA biosynthesis, enhancing stress tolerance in pine plants expressing the 409
transgene.
410
Abscisic acid (ABA) is recognized as a major plant hormone during drought 411
stress, since it inhibits growth and stomatal opening. Upon water deficit, both 412
ABA biosynthesis in roots and its transport to the leaves are enhanced, leading 413
to its accumulation in guard cells. In the stomata, ABA induces the release of 414
water and loss of turgor of guard cells, provoking the closure of the stomata 415
pore (Anderson et al., 1994; Allan et al., 1994). It is known that different 416
Populus species and ecotypes may differ in their stomatal responsiveness to 417
ABA (Chen et al. 2002; Yin et al. 2004; Zhang et al.). Chen et al. (2002) 418
reported that the drought-induced decline of PAs concentrations in a sensitive 419
Populus species was accompanied by leaf shedding, whereas the tolerant 420
species maintained higher PAs levels and did not shed its leaves. The same 421
authors showed that in water-stressed Poplar, xylem ABA reduces PA contents, 422
and hypothesized that this fact might intensify the sensitivity of the leaf to 423
ethylene, thus accelerating defoliation. Bae et al. (2008) reported that in cacao 424
(T. cacao), the induction by ABA (100 mM solution applied to soil) of all 5 ESTs 425
associated with PA biosynthesis (TcODC, TcADC, TcSAMDC, TcSPMS and 426
TcSPDS) was low, similarly to what has been observed in rice (Li and Chen, 427
2000), where even a fall in SAMDC1 was registered 12 h after rice plants were 428
treated with ABA. The decline in transcript levels was assigned to changes in 429
mRNA stability (Li and Chen, 2000). The gene induction patterns triggered by 430
ABA in cacao leaves and roots disagree with those observed in other plant 431
species. ADC2 was highly induced by ABA (50 mM) in Arabidopsis (Pérez- 432
Amador et al., 2002). Also in Arabidopsis, Alcázar et al. (2006) observed that 433
ADC2, SPMS, and SPDS1 were highly induced by drought and greatly reduced 434
by this stress in ABA insensitive mutants. ABA triggered significant alterations in 435
the PA catabolic pathway of grapevine leaf, but at the same time, it also 436
induced the activity of biosynthetic enzymes ADC, ODC, and SAMDC (mainly in 437
the tolerant genotype), justifying the interplay between PA anabolism and ABA 438
signaling pathways in grapevine (Toumi et al., 2010). This induction, which took 439
place within 1 h post-treatment, resulted in different enzyme induction patterns 440
in the tolerant and sensitive genotypes, with the sensitive genotype responding 441
later and less profoundly. On other hand, PAs oxidation concerned PAOs in the 442
tolerant genotype and DAOs in the sensitive genotype were evaluated. On the 443
base of this information it was proposed the following model: PA biosynthesis is 444
higher in the tolerant than in sensitive grapevine genotypes; in both genotypes, 445
PAs follow the exodus route and are catabolized in the apoplast by AOs, 446
producing H2O2; in the case of high intracellular PA titers/PA anabolism, 447
tolerance is enhanced via induction of additional defensive genes/responses; in 448
the case where PA titers/PA anabolism is low, H2O2 enhances programmed cell 449
death.
450
It has been shown that nitric oxide (NO)-treated plants have increased tolerance 451
to drought (Garcia-Mata and Lamattina 2001, 2002). Arasimowicz-Jelonek et al.
452
(2009) demonstrated the occurrence of a functional cross-talk between PAs and 453
NO in cucumber leaves under drought stress. Although exogenous PAs (1 mM 454
Put, Spd and Spm) did not affect NO production in well-watered cucumber 455
seedlings, their treatment with Spm and Spd, prior to water deficit imposition, 456
induced early and higher NO levels in leaves of drought-stressed cucumber 457
plants, with respect to the control and Put treated ones.
458
Recently, Alcázar et al. (2010) discussed advances in the cross talk between 459
PAs and ABA, integrating them with other abiotic stress-related metabolic 460
routes such as reactive oxygen species (ROS) signaling, generation of NO, 461
modulation of ion channel activities and Ca2+ homeostasis.
462
463
Saline stress 464
First reports on the induction of plant PAs metabolism by salt stress, as well as 465
their possible alleviating roles on plant salt tolerance can be traced back to the 466
eighties (20th Century). Many authors have reported that PAs accumulation is 467
the immediate response observed in different crop plants species after 468
exposure to saline conditions (Erdei et al., 1996; Chattopadhayay et al., 2002;
469
Ghosh et al., 2011). Most significant changes in polyamines level upon 470
salinization appear to be those of Spm, according to data reported in rice 471
(Maiale et al., 2004), maize (Jiménez-Bremont et al., 2007; Rodríguez et al., 472
2009) and wheat (Reggiani et al., 1994; El-Shintinawy, 2000). Thus, under 473
salinity, the pool of Put would be directed to Spd and finally to Spm synthesis 474
(Groppa and Benavides, 2008). In rice, (Krishnamurthy y Bhagwat, 1989; Roy 475
et al., 2005; Roychoudhury et al., 2008) wheat (El-bassiouny and Bekheta, 476
2005) and barley (Liu et al., 2006), the build up of the Spm level has been 477
described as an indicator of salt tolerance, whereas Put accumulation has been 478
associated with salt sensitivity. In bean, two cultivars with contrasting tolerance 479
to salt and drought stress, Pinto Villa (tolerant) and Canario 60 (sensitive) 480
showed differential PAs accumulation in leaves. In particular, Spm was 481
accumulated in the tolerant cultivar at 150 and 400 mM NaCl, while a decrease 482
in PA content was evident in the sensitive cultivar (Hernández-Lucero et al., 483
2008). Roy et al. (2005) clearly demonstrated that deficiencies of salt-sensitive 484
rice cultivars, due to high Na+ accumulation or salinity stress-induced K+ loss, 485
could be overcome by exogenously supplied Spd, necessary to Spm synthesis.
486
In general, plants respond to abiotic stress by increasing ADC activity 487
(Bouchereau et al., 1999). Roy and Wu (2001) reported that under salinity, rice 488
plants transformed with a gene encoding an oat ADC increased the PAs level 489
and plant biomass as a consequence of a higher ADC activity. Chattopadhyay 490
et al. (1997) reported that ADC transcripts and activity increased in rice cultivars 491
as early as 1 h after the stress treatment was imposed, followed by a sharp 492
decrease after prolonged salt treatment, in the case of salt-sensitive cultivar.
493
Roy and Wu (2005) transformed rice plants with a Tritordeum SAMDC gene 494
and observed a three to four-fold rise in Spd and Spm levels in transformed 495
plants under NaCl-derived stress. Li and Chen (2000) reported that the 496
expression of the SAMDC1 gene in rice seedlings was dramatically induced by 497
salinity. The transcript levels of SAMDC1 in two rice varieties differing in salt 498
tolerance were found to be higher in the salt-tolerant than in the salt-sensitive 499
variety. Although there are reports in which some PA biosynthetic genes such 500
as ADC and SAMDC transcripts were barely affected by NaCl treatment 501
(Jiménez-Bremont et al., 2007), there is evidence of up-regulation of other PA 502
biosynthetic genes under salinity: ZmSPMS1 in maize (Rodríguez-Kessler et 503
al., 2006; 2010; Rodríguez-Kessler and Jimenez-Bremont, 2008), OsSPDS in 504
rice (Imai et al., 2004), and ZmSPDS1 in maize (Jiménez-Bremont et al., 2007).
505
Although the mechanisms that govern PA metabolism-mediated salt tolerance 506
remain unclear, some reports have shed light in the last years. Mansour and Al- 507
Mutawa (1999) reported that Spd or Spm but not Put alleviates the cellular 508
alterations in wheat roots under saline stress, possibly by plasma membrane 509
protection. Accordingly, Spm and Spd significantly prevented the leakage of 510
electrolytes and amino acids from roots and shoots of rice subjected to salinity 511
(Chattopadhayay et al., 2002). Saline stress-induced elevation of PA levels may 512
represent an adaptive mechanism in which the uptake of Na+ and leakage of K+ 513
from mesophyll cells are reduced (Pang et al., 2007). Pre-treatment with PAs 514
prevented salt-induced K+ leakage in mature root zone of hydroponically grown 515
maize, apparently by effects on cell membrane transporters in a highly specific 516
way (Pandolfi et al., 2010). Shabala et al., (2007) showed that PA treatment 517
substantially reduced the NaCl-induced K+ efflux from the pea leaf mesophyll, 518
most likely by blocking the non-selective cation channels. Zhao and Qin (2004) 519
reported that exogenous PAs application could maintain tonoplast integrity and 520
function in barley seedlings under saline conditions. Legocka and Kluk (2005) 521
reported higher levels of PAs bound to microsomal membranes in Lupinus 522
luteus seedlings in salinity, and proposed that PAs most likely stabilized 523
microsomal membrane surfaces, protecting them against NaCl stress damage.
524
Many authors suggested that, under salinity and other environmental adverse 525
conditions, PAs act as antioxidants though their precise role is still a matter of 526
debate (Groppa and Benavides, 2008). Under NaCl-induced stress, a higher 527
level of lipid peroxidation was observed in the salt sensitive, relative to the salt- 528
tolerant wheat (El-bassiouny and Bekheta, 2005) and rice (Roychoudhury et al., 529
2008) cultivars, which showed augmented Spd and Spm levels, not observable 530
in the salt sensitive cultivars.
531
Xing et al., (2007) analyzed the effects of treatments with different NaCl 532
concentrations, with or without aminoguanidine (AG, a specific DAO inhibitor) 533
on endogenous PAs, GABA content and DAO activity in soybean roots (Glycine 534
max L. Merr., cultivar Suxie-1). Results showed significant Put, Cad and Spd 535
decreases with increasing salt concentrations, assignable to the promotion of 536
DAO activity by salinity and consequent stimulation of PA degradation. In 537
parallel, GABA accumulation raised with growing NaCl concentrations, strongly 538
suggesting its origin in PA degradation.
539
ROS are necessary in many plant developmental processes (Foreman et al., 540
2003; Demidchik and Maathuis, 2007), such as in the elongation zone of maize 541
leaves during leaf extension (Rodríguez et al., 2002). In maize, the salt-induced 542
decrease of extracellular ROS contributes to the reduction of leaf elongation 543
(Rodríguez et al., 2004). In turn, the diminution of the extracellular ROS has 544
been attributed to the inhibitory effect of NaCl on the NADPH oxidase complex 545
(Rodríguez et al., 2007). Rodríguez et al., (2009) reported that under saline 546
stress extracellular ROS registered in the elongation zone of maize leaves are 547
produced principally by PAO, contributing partially to counteract the growth- 548
inhibiting effect caused by salinity. In turn, this same phenomenon was 549
described in soybean hypocotyls grown under NaCl stress (Campestre et al., 550
2011). In soybean, saline stress increased Spm and Cad levels and copper 551
amine-oxidase (CuAO) activity. Treatment with the CuAO inhibitor showed a 552
significant reduction of ROS in the elongation zone, and plants grown in Cad- 553
amended culture medium showed longer hypocotyls in saline conditions 554
(relative to the unamended treatment), an effect that was abolished by the 555
CuAO inhibitor. Since H2O2 functions as a signal molecule activating many of 556
the plant responses deployed to cope with stress, it is believed that its 557
generation from PAs oxidation (along with PA depletion) might orchestrate, at 558
least partially, plant adaptation to these conditions (Moschou et al., 2008;
559
Rodríguez et al., 2009).
560
561
Cold stress 562
In temperate climates, plants species have acquired a certain degree of cold 563
tolerance, depending of the genetic background, cold hardness and exposure 564
time (Janska et. al 2010). Plant physiologists use the term freezing to mean 565
temperatures below 0°C, chilling for temperatures between 0°C and the 566
minimum temperature necessary for growth, and temperatures between that 567
minimum and the optimum is denominated suboptimal temperature for growth.
568
Such a difference in stress terminology is not trivial, since the physiological 569
response of a plant species may be different in each case. Crop species such 570
as rice, maize and soybean are exceptionally subjected to freezing periods, 571
more regularly they endure chilling and the most common situation is 572
suboptimal growth temperature. Whereas freezing kills these plant species, 573
chilling and suboptimal temperature constitute an important constrain to 574
productivity. In the last two cases, damage levels depend on the magnitude of 575
temperature diminution and the exposure time. In contrast, wheat, barley and 576
oat crops are normally grown under freezing, chilling and suboptimal growth 577
temperatures, being freezing tolerated to some extent by these species.
578
There are two different strategies to overcome low temperature stress:
579
avoidance and tolerance. In terms of crop production, avoidance may be 580
determined by the sowing period, growth cycle and agronomic management, 581
but tolerance is a genetic feature, peculiar to each cultivar, which constitutes a 582
major tool for crop production management in areas characterized by low 583
temperatures.
584
Cultivar response to low temperature stress involves important biochemical and 585
molecular changes. Essentially, plants increase the production of protective 586
compounds that affect cell lipid composition, thus participating in membrane 587
stabilization and maintaining plasma membrane functionality (Janska et. al 588
2010). Biochemical changes also include the synthesis of cryoprotectant 589
molecules as soluble sugars (saccharose, raffinose, stachyose, trehalose), 590
sugar alcohols (sorbitol, ribitol, inositol) and low-molecular weight nitrogenous 591
compounds (proline and glycine betaine). Symplastic and apoplastic soluble 592
sugars directly contribute to membrane stabilization (Livingston et al. 2006).
593
Also, compounds such as tripeptidthiol, glutathione, ascorbic acid (vitamin C) 594
and a-tocopherol (vitamin E) are important for their antioxidant activity (Chen 595
and Li, 2002). PAs are also involved in the stress response to low temperatures.
596
Different modifications at the transcriptional and metabolic levels have been 597
reported, mainly in A. thaliana. Currently, attempts are being made to 598
manipulate PAs metabolism genes in order to obtain plants tolerant to low 599
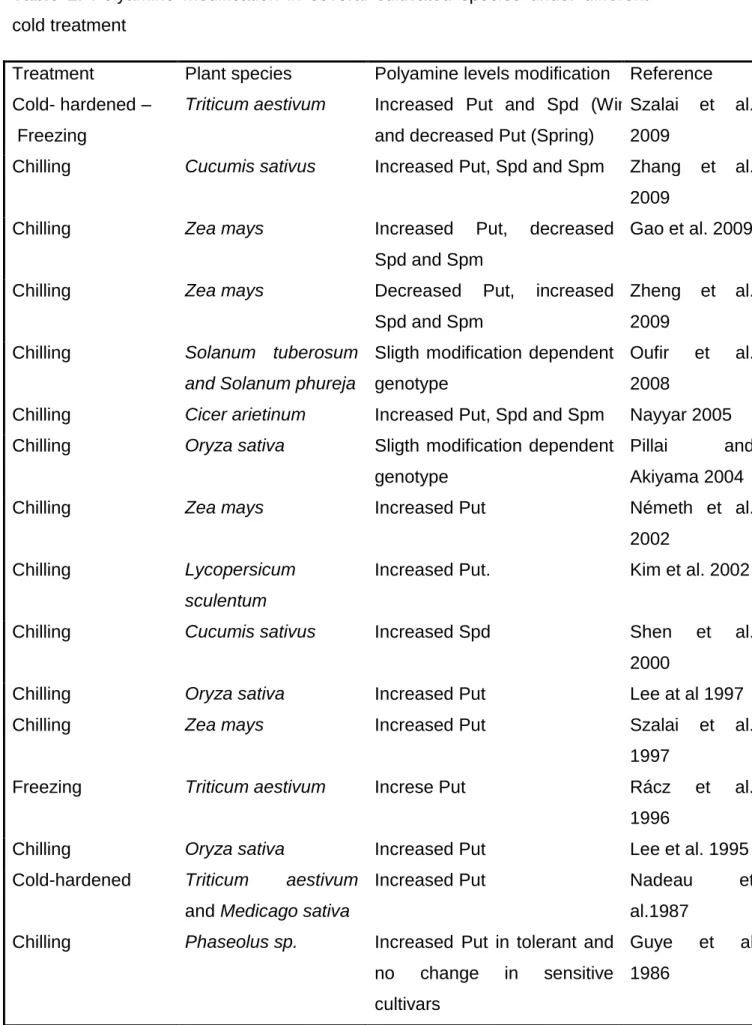
temperature stress. Table 2 shows variations in PAs levels during the response 600
to cold temperature in several crop species.
601
In five bean (Phaseolus sp.) cultivars differing in chilling response, Guye et al.
602
(1986) found that prior to chilling treatment; PA levels did not appear to be 603
correlated with chill-tolerance, since levels in non-chilled controls were highest 604
in cultivars of medium chill-sensitivity. These authors also found that the Put 605
levels were increased in tolerant cultivars, whereas no changes were observed 606
in sensitive ones. They concluded that it is the change in Put titer rather its 607
absolute level what appears to be correlated with chill-tolerance (Guye et al., 608
1986). In two wheat cultivars with slightly difference in response to cold 609
tolerance, Nadeau et al. (1987) found a 6 to-9-fold increased Put level during 610
cold acclimation, whereas a smaller raise was observed in the Spd content and 611
conversely, Spm decreased. These authors also found an increase in Put level 612
of alfalfa (Medicago sativa) under cold stress. ADC activity declined in cold 613
treated plants, whereas no major variations were observed in ODC activity 614
levels, suggesting that ADC is the main enzyme responsible for the 615
incremented Put levels under the cold-hardening condition. In turn, it was 616
described that DAO activity varied in parallel with Put content in many plants 617
species. In a short term freezing stress experiment in wheat, a notable Put and 618
agmatine accumulation was observed when plants were subjected to -2°C 619
during 6 h (Rácz et al., 1997). The synthesis of agmatine indicated that Put 620
accumulation was mediated by ADC. Again, no major variations were observed 621
in ODC activity levels, reinforcing the idea that ADC is the main enzyme 622
responsible to increase in plant Put levels during cold hardening. In experiments 623
carried out with chilled (5°C) maize; Szalai et al. (1997) found a continuous 624
raise in the Put level, which was more pronounced under the light condition in 625
comparison to darkness. One day after chilling, Spd also increased in light, 626
whereas it decreased in darkness. After the second day, chilling provoked a 50 627
and 80% fall in the Spd content in light and dark, respectively, compared with 628
the unstressed control. Likewise, experiments performed on winter and spring 629
wheat grown under low and normal light conditions showed that changes in PAs 630
contents were markedly light dependent (Szalai et al., 2009).
631
Using seedlings of two inbreeded maize lines differing in cold sensitivity, it was 632
found that Put concentrations escalated after chilling stress in mesocotyl and 633
coleoptile, but the root Put concentration remained unchanged (Gao et al.
634
2002). Inversely, Spd and Spm concentrations decreased after chilling stress in 635
the three mentioned seedling organs. On the other hand, the electrolyte leakage 636
in cold stressed tissues was lower in the tolerant than in the sensitive cultivar, 637
whereas the level of this parameter was lower in the coleoptile than in the 638
mesocotyl, in both cultivars. Stepwise regression analysis of these data showed 639
that chilling injury in roots was generally correlated with Spd concentration while 640
in the mesocotyl, injury was mainly correlated with Put and Spd concentrations.
641
Zheng et al. (2009) found that Put was reduced, but Spd and Spm were 642
increased during chilling stress (5°C, 48 h) in maize. However, Spd and Spm 643
contents were higher in the tolerant than in the sensitive cultivar. The values of 644
(Spd + Spm)/Put ratios were negatively correlated with malondiahldehyde 645
contents, whereas treatment with MGBG resulted in raised malondialdehyde 646
content and reduction of germination percentage in both maize cultivars.
647
Lee et al. (1997) observed that PAs contents as well as ADC and SAMDC 648
activities were increased in the stress tolerant cultivar Tainung 67. Furthermore, 649
ABA was increased under chilling conditions in according to Put levels whereas 650
treatments with Put inhibitors provoked enhanced sensitivity to chilling while 651
pretreatments with ABA improved tolerance to cold stress., In this sense, it has 652
been reported a correlation between ABA and Put with chilling stress tolerance 653
in 11 rice cultivars (Lee et al. 1995).
654
On other hand, Pillai and Akiyama (2004) found that the rice OsSAMDC gene 655
was induced in the tolerant cultivar Yukihikari but not in the sensitive TKM9 one.
656
Spd levels increased in shoots of Yukihikari and it was not altered in TKM9, 657
whereas Put and Spm remained unchanged in both cultivars. OsSAMDC was 658
also induced by Ethephon in both cultivars, but this gene was not responsive to 659
salt, drought, submergence, mannitol or ABA.
660
In cucumber plants, chilling induced a notable Spd raise in a tolerant cultivar, 661
but not in a sensitive cucumber cultivar (Shen et al. 2000). Also, Put synthesis 662
during the rewarming period in the tolerant cultivar, but there was no change in 663
the sensitive one. In contrast, Zhang et al. (2009) reported increased PAs 664
content in tolerant cucumber plants and a slight Put increase in sensitive ones.
665
Such apparent incongruence might be attributed to the different chilling 666
conditions employed: 3°C in the first case versus 15/ 8°C in the second. Shen et 667
al. (2000) reported that augmentations in Put and Spd were preceded by 668
enhancements in ADC and SAMDC activities. Pretreatment of sensitive plants 669
with Spd prevented chill-induced increments in leaf H2O2 contents and 670
nicotinamide adenine dinucleotide phosphate (NADPH)-oxidase activity, 671
alleviating chilling injury. On the other hand, the application of MGBG to a 672