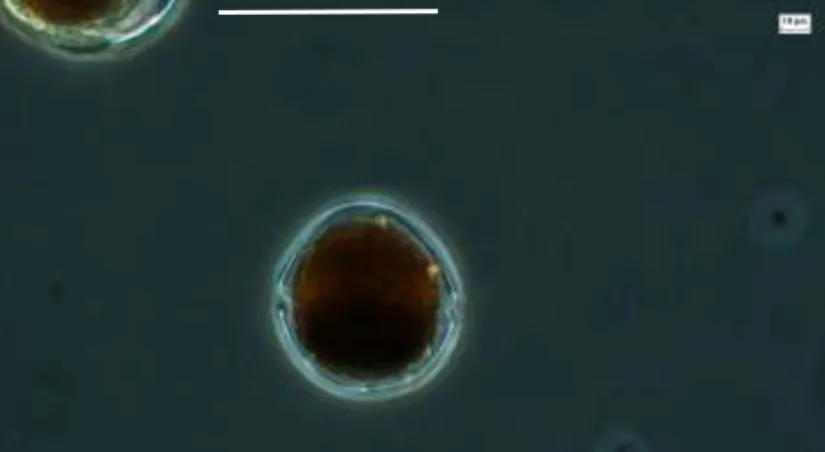
INTRODUCCIÓN
Acuicultura e infecciones bacterianas
Por otro lado, el rápido crecimiento e intensificación de los sistemas de acuicultura marina ha afectado la calidad y seguridad de los organismos, aumentando la incidencia de infecciones bacterianas en criaderos, engordes y estanques de engorde, provocando diversas enfermedades en peces, moluscos y crustáceos. , provocando muertes masivas y pérdidas económicas importantes (Falaise et al., 2016; Santos y Ramos, 2018; EIA, 2009). Una solución obvia a este problema fue la administración de antibióticos en forma de fármacos metafilácticos y profilácticos (Carvalho et al., 2012).
Características generales de las bacterias patógenas marinas
- Vibrio spp
- Vibrio parahaemolyticus
Las bacterias patógenas marinas se pueden encontrar suspendidas en el agua, asociadas con partículas, en los sedimentos del fondo marino o en los cuerpos o células de organismos marinos (Belkin y Colwell, 2005). Vibrio es una de las principales causas de enfermedades bacterianas en los organismos acuáticos, se clasifican como oportunistas, cuando los huéspedes tienen el sistema inmunológico comprometido, también debido al estrés provocado por la intensificación de los sistemas marinos.
Las microalgas como fuente de compuestos antibacterianos
- Pigmentos fotosintéticos
- Clorofilas
- Carotenoides
- Lípidos
- Ácidos grasos
Incluso entre los lípidos no polares, los ácidos grasos libres se encuentran en cantidades menores (Mimouni et al., 2018). Según el número de dobles enlaces, los ácidos grasos se dividen en saturados (SAFA), monoinsaturados (MUFA con un doble enlace) y poliinsaturados (PUFA con más de 2 dobles enlaces) (Wang et al., 2017).

Lípidos y pigmentos en diatomeas y dinoflagelados
- Nanofrustulum shiloi (J.J. Lee, Reimer & McEnery) Round, Hallsteinsen & Paasche,
- Odontella aurita (Lyngbye) C. Agardh 1832
- Alexandrium margalefii (Balech, 1994)
- Gymnodinium catenatum (Graham, 1943)
Comparación de ácidos grasos (% de ácidos grasos totales) que se identificaron de la diatomea Odontella aurita en el presente trabajo y en informes anteriores. Comparación de ácidos grasos (% de ácidos grasos totales) que fueron identificados de la diatomea Nanofrustulum shiloi, del presente trabajo y de informes anteriores.

ANTECEDENTES
Compuestos bioactivos antibacterianos de diatomeas y dinoflagelados
Los dinoflagelados y las diatomeas son microalgas que producen una variedad de pigmentos y productores de ácidos grasos que normalmente comprenden del 15 al 25% de su biomasa seca (Mansour et al., 1999; Yi et al., 2017). En el caso de los dinoflagelados, ofrecen una amplia variedad de ácidos grasos y pigmentos, como ácidos grasos altamente insaturados de cadena larga (Mansour et al., 1999; Leblond et al., 2003). Este compuesto mostró una actividad limitada contra las bacterias pero una fuerte actividad antifúngica (Sharma et al., 1968).
Lípidos y ácidos grasos como antibacterianos
No se observó actividad antibacteriana contra la bacteria E. 2016), trabajaron con 21 diatomeas y 7 dinoflagelados y obtuvieron extractos de acetona y realizaron pruebas antibacterianas contra bacterias Gram positivas y negativas. Walter y Mahesh (2000) probaron extractos de 11 diatomeas marinas cultivadas hasta el inicio de la fase estacionaria y utilizaron diferentes disolventes para obtener extractos que probaron contra 2 bacterias Gram positivas y 11 Gram negativas. Desbois y otros (2008) también probaron extractos de la diatomea Phaeodactylum tricornutum contra bacterias patógenas del medio marino que son perjudiciales para los seres humanos.
Pigmentos fotosintéticos como antibacterianos
Compuestos antibacterianos utilizados contra V. parahaemolyticus
En el caso de Thanigaivel et al. 2014) probaron extractos etanólicos de la macroalga Chaetomorpha antennina, contra aislados de Vibrio de cultivos de camarón. En ambos tipos de ensayos mostraron efectos bactericidas, los cuales dependieron de la microalga utilizada, y en el caso de un extracto hidrófilo se encontró un efecto inhibidor, al contar las células viables, para C.
JUSTIFICACIÓN
Peso seco total, orgánico e inorgánico y porcentaje de cenizas de las diatomeas Odontella aurita y Nanofrustulum shiloi y los dinoflagelados Alexandrium margalefii y Gymnodinium catenatum. El porcentaje de pigmentos totales con base en el peso seco de las diatomeas Odontella aurita y Nanofrustulum shiloi y los dinoflagelados Alexandrium margalefii y Gymnodinium catenatum. El porcentaje de lípidos totales con base en el peso seco de las diatomeas Odontella aurita y Nanofrustulum shiloi y los dinoflagelados Alexandrium margalefii y Gymnodinium catenatum.
Composición total de ácidos grasos en relación al peso seco de las diatomeas Odontella aurita y Nanofrustulum shiloi, y los dinoflagelados Alexandrium margalefii y Gymnodinium catenatum. Porcentajes de inhibición de extractos de pigmentos totales por el método de la resazurina de las diatomeas Odontella aurita y Nanofrustulum shiloi, y de los dinoflagelados Alexandrium margalefii y Gymnodinium catenatum. Porcentajes de inhibición de extractos de lípidos totales por el método de la resazurina de las diatomeas Odontella aurita y Nanofrustulum shiloi, y de los dinoflagelados Alexandrium margalefii y Gymnodinium catenatum.
Comparación de los ácidos grasos (% de ácidos grasos totales) identificados en el dinoflagelado Gymnodinium catenatum a partir del trabajo actual e informes anteriores.

HIPÓTESIS
OBJETIVOS
Objetivo general
Objetivos particulares
Se calcula la pendiente (m) de la ecuación (Y= mx + b), la cual corresponde al factor de corrección utilizado para la cuantificación de cada uno de los ácidos grasos de las muestras. En el caso de los ácidos grasos, su acción depende de la composición de la pared celular, y este efecto inhibidor aumenta en los ácidos grasos insaturados, con el número de insaturados (Thormar, 2011). Comparación de los ácidos grasos (% de ácidos grasos totales) identificados a partir del dinoflagelado Alexandrium margalefii en el presente trabajo y en informes anteriores.
MATERIAL Y MÉTODOS
Material biológico y condiciones de cultivo
- Microalgas
- Bacterias
Trabajamos con la diatomea Nanofrustulum shiloi (CIB-24) recolectada en octubre de 2009 en Balandra Bay, B.C.S. Se ampliaron a matraces de 2 litros y se cosecharon en fase exponencial tardía, 3 semanas (1 x 106 células mL-1). Luego se cortó en medio TSA con NaCl al 2% y se incubó a la misma temperatura.
Recuento celular de las microalgas
La cepa se criopreservaó en glicerol y se reactivó en medio TSB con NaCl al 2% mediante incubación durante 24 horas a 30°C con agitación.
Peso seco total, orgánico e inorgánico de las microalgas
Posteriormente se colocaron en sobres de papel de aluminio y se quemaron a 490°C en la mufla TERLAB (modelo TE-M12D) durante 6 horas. Luego se colocaron en el desecador durante 1,5 h y se pesaron en una balanza analítica marca OHAUS Pioneer (modelo PA 114). El proceso se repitió hasta que se aseguró que se lograba un peso constante de los filtros.
Extracción de pigmentos totales
Los extractos de pigmentos combinados se diluyeron con acetona de calidad HPLC al 100% hasta que las lecturas de absorbancia en diversas longitudes de onda estuvieron dentro del rango de absorbancia inferior a 1. Las lecturas de absorbancia se tomaron en un espectrofotómetro marca HACH (modelo DR5000). ), mediante un barrido espectral. de 400 a 700 nm. A partir de las lecturas de absorbancia obtenidas se calcularon las concentraciones de clorofila a, c y caroteno total.
Cuantificación de pigmentos por HPLC-DAD
A partir de los lípidos totales obtenidos previamente, la extracción de los ácidos grasos se realizó mediante la reacción de metanólisis (derivatización) agregando 2.5 mL de la mezcla HCl:CH3OH (5:95) (Sato y Murata, 1988). Asimismo, en bacterias gramnegativas puede existir la presencia de ácido teicoico, que inhibe la acción de los ácidos grasos (Kohler et al., 2009). El ácido linoleico (18:2ω-6) ha mostrado un efecto inhibidor del crecimiento en bacterias grampositivas, pero no en bacterias gramnegativas (Thormar, 2011); El ácido linolénico (18:3ω-3) también ha sido uno de los que mayor propiedad antibacteriana tiene (Churchward et al., 2018).
Cuantificación de lípidos totales por métodos espectrofotométricos
Cuantificación de ácidos grasos por GC-FID
La mezcla anterior se lavó añadiendo 1,5 ml de hexano de calidad HPLC y se agitó nuevamente. La identificación se realizó comparando el tiempo de retención (TR) de la mezcla de estándares comerciales con el de las muestras de microalgas. Para cuantificar los ácidos grasos de las muestras se realiza una regresión lineal de las curvas de calibración de cada uno de los ácidos grasos comerciales.
Extracción de lípidos para la evaluación de antibacterianos
La curva de calibración fue preinyectada con una mezcla comercial de ácidos grasos metilados (Sigma T6543, ácido tricosanoico).
Extracción de pigmentos para la evaluación de antibacterianos
La evaluación de la actividad antibacteriana se realizó mediante la determinación de la concentración mínima inhibitoria (CMI) en microplacas de 96 pocillos para cada uno de los extractos de diferentes microalgas. En esta investigación, el caroteno principal fue la diadinoxantina, que según se informó correspondía al 20 al 30% del total de carotenoides en los dinoflagelados (Demers et al., 1991). Sólo se cuantificaron dos IAM, 18:1ω-9 con 3,4%, un porcentaje bajo de lípidos totales como en esta investigación.
Curvas de crecimiento bacteriano
Evaluación de concentración mínima inhibitoria por el método de resazurina
A cada pocillo se le añadió un volumen de 150 µL de los extractos a evaluar por cuadruplicado, se utilizaron diferentes concentraciones (Cuadro 3), iniciando con una concentración en el caso de lípidos totales de 490 µg EPA/DHA y en pigmentos. Como control negativo se utilizó una microplaca con el mismo orden de carga de los extractos, sin inóculo bacteriano. Pasado el tiempo de incubación utilizamos como indicador la resazurina, que en un ambiente sin crecimiento celular cambia de morado a azul (se oxida), cuando ocurre el crecimiento se vuelve rosado (reducido) debido a la respiración aeróbica de las bacterias.
Análisis estadísticos
RESULTADOS
- Peso seco total, orgánico e inorgánico de las diatomeas y dinoflagelados
- Pigmentos totales por espectrofotometría
- Cuantificación e identificación de pigmentos totales por HPLC-DAD
- Cuantificación lípidos totales por espectrofotometría
- Cuantificación de ácidos grasos por GC-FID
- Curvas de crecimiento bacteriano
- Evaluación de concentración mínima inhibitoria por el método de resazurina
En todas las microalgas se identificó clorofila a, fue el pigmento encontrado en mayor concentración. El ácido graso saturado encontrado en mayor concentración en las cuatro microalgas fue el ácido palmítico (16:0). El AGP en mayor porcentaje en el caso de las diatomeas fue el ácido linoleico (18:2ω-6), el cual no se encontró en A.

DISCUSIÓN
Pigmentos en O. aurita, N. shiloi, G. catenatum y A. margalefii
Lípidos totales en O. aurita, N. shiloi, G. catenatum y A. margalefii
Ácidos grasos en O. aurita, N. shiloi, G. catenatum y A. margalefii
También informaron 4 ácidos grasos insaturados, 18:4ω-3 al 0,66%; este ácido graso no se encontró en la caracterización del perfil de ácidos grasos que realizamos. Reportaron valores de EPA (6,79%) y DHA (0,47%), que también son similares a los reportados en el perfil realizado en la presente investigación (ANEXO B, Tabla B2). La proporción de EPA que informaron un 17% fue significativamente mayor que la reportada en el presente trabajo. 1991), a partir de cultivos que realizaron perfiles de ácidos grasos del dinoflagelado G. catenatum, calcularon una cantidad total de AGS de 36,1%, similar a la presente investigación.
Propiedades antibacterianas de pigmentos y ácidos grasos
Otro posible modo de acción de los ácidos grasos, en el caso de que ingresen a la célula, es inhibir la biosíntesis de ácidos grasos en las bacterias, este es un proceso que no ocurre con los compuestos antibacterianos existentes y por lo tanto es una posibilidad mínima. de resistencia (Churchward et al., 2018). Por otro lado, las bacterias Gram positivas, al tener múltiples capas de peptidoglicano, proporcionan una porosidad que permite que los agentes antibacterianos penetren en la pared celular (Alves et al., 2020), lo que podría explicar que en las bacterias Gram positivas S. Toxinas paralizantes en moluscos bivalvos durante una proliferación de Gymnodinium catenatum Graham en la Bahía de La Paz, México.
Bioprospección de la diatomea Nanofrustulum shiloi (Bacillariophyceae), nativa del noroeste de Baja California Sur, México, para su uso como alimento alternativo en sistemas acuícolas. Aislamiento, purificación y mantenimiento de cepas de microalgas. eds) Métodos y herramientas analíticas en la evaluación de la biomasa de microalgas, 2ª ed.

CONCLUSIONES
LITERATURA CITADA
Isolation and structural characterization of two antibacterial free fatty acids from the marine diatom Phaeodactylum tricornutum. Transcriptome analysis of the effect of polyunsaturated fatty acids against Vibrio vulnificus infection in Oreochromis niloticus. Fatty acid profile and temperature in the cultured marine diatom Odontella aurita. eds) Microbiology of Waterborne Diseases, Elsevier, pp.
ANEXOS