El uso excesivo de fungicidas para el control de Alternaria spp., ha provocado que este patógeno adquiera resistencia. El objetivo fue evaluar la capacidad antagónica de cepas nativas de Trichoderma contra hongos patógenos Alternaria spp. Trichoderma ejerció actividad antagonista contra el patógeno Alternaria y demostró un efecto estimulante en las plantas de brócoli.
El uso excesivo de fungicidas para el control de Alternaria spp., hizo que este patógeno adquiriera resistencia. 3.4.2..Tratamiento para evaluar la capacidad protectora contra Alternaria en el cultivo de brócoli..19 3.5. 3.5.1.. Diseño para evaluación por absorbancia para el desarrollo micelial de Alternaria spp. Diseño experimental para evaluar la capacidad.
3.7.1..Aislamiento e identificación molecular de Trichoderma spp de cultivares de brócoli Aislamiento e identificación molecular de Alternaria spp de. 4 Aislamiento de cultivos puros de Alternaria spp..30 Figura 5 Identificación de Alternaria spp. por PCR………..30 Figura 6 Amplificación del gen ech42 por PCR en Trichoderma spp..31 Figura.
INTRODUCCIÓN
Problema científico
Por lo tanto, para alimentar a esta creciente población mundial, es necesario aumentar la producción de alimentos en aproximadamente un 70% en la agricultura. El aumento significativo de la producción de alimentos ha ayudado a satisfacer las necesidades de seguridad alimentaria global, pero problemas como el calentamiento global, la contaminación ambiental y la explosión demográfica han empujado a las plantas a diversos tipos de estrés biótico como hongos, bacterias, virus, nematodos, malezas y insectos , provocando una pérdida de rendimiento de hasta el 31-42%.
Hipótesis
Objetivos
- Problema General
- Problema Específico
MARCO TEÓRICO
- Generalidades del Brócoli
- Variedades
- Cultivo de brócoli en el Ecuador
- Principales enfermedades del brócoli
- Alternaria spp
- Control biológico de enfermedades
- Trichoderma spp
- Mecanismos de acción
Los síntomas de la enfermedad de Alternaria aparecen en las hojas, pecíolo, inflorescencia, tallo y semillas (Uroj et al., 2019). Estos compuestos se producen como resultado de la degradación enzimática hidrolítica de células funcionales o vegetales (Ahluwalia et al., 2015). Otras enzimas que mejoran la resistencia en las plantas son las quitinasas y las glucanasas (McIntyre et al., 2014).
También incluyen proteínas relacionadas con la patogénesis (PR) (respuesta SAR) y enzimas que desempeñan un papel en la respuesta de defensa antioxidante (McIntyre et al., 2014). Se ha establecido que las especies de Trichoderma compiten por nutrientes, nichos biológicos o puntos de infección con patógenos en la rizosfera de las plantas (Ahluwalia et al., 2015). Dependiendo de su origen biosintético, estos compuestos se pueden agrupar en peptaibol, policétido y terpeno (Hu et al., 2017).
Diferentes especies de Trichoderma sintetizan una amplia variedad de antibióticos (Reino et al., 2008), por ejemplo T. El gen de la endoquitinasa (ech42), que codifica las endoquitinasas, es producido por los hongos Trichoderma spp. Es responsable del micoparasitismo (Barbara et. al. 2011).
MATERIALES Y MÉTODO
- Localización
- Tipo de investigación
- Material genético
- Tratamientos
Para la obtención de cepas de NINTANGA Trichoderma se colocaron 20 trampas en una parcela de 1,5 hectáreas utilizando la metodología X descrita por Vallejo, (2014). Estas cepas de Trichoderma spp fueron purificadas y serán reinoculadas en medio de cultivo PDA, incubadas durante 8 días a 29 °C para la extracción de ADN (Arbito, 2017). Las cepas de Trichoderma se identificaron mediante PCR utilizando ADNg extraído de micelios de hongos utilizando un kit QIAGEN-Start Protocol (EE. UU.).
Las muestras amplificadas se analizaron mediante electroforesis en gel de agarosa al 1,5 % y se tiñeron con bromuro de etidio (0,5 ug/ml) (Johanson, 1994). Las muestras amplificadas se analizaron mediante electroforesis en gel de agarosa al 1,5% y se tiñeron con bromuro de etidio (0,5 ug/ml) (Carsolio, Gutiérrez, Jiménez, Van y Herrera, 2004). Para obtener el micelio y el sobrenadante, los tubos de ensayo se centrifugaron durante 30 minutos a 3500 revoluciones por minuto, el sobrenadante se filtró en miliporos 0,2 µM en viales estériles y las muestras se almacenaron a -4 °C con ayuda de una jeringa, debidamente etiquetado (Rodríguez & Piñeros, 2017).
Se evaluó el efecto de los metabolitos sobre el crecimiento micelial al 10% y se agregarán 720 µL de medio Sabourand; 90 µL de suspensión micelial y 90 µL de una concentración de metabolitos de Trichoderma spp con actividad proteolítica (Harman, 2006). Se colocaron 200 g de sustrato (maíz) limpio e hidratado en bolsas plásticas y se esterilizaron a 15 libras de presión o 125 ºC y se inoculó un segmento de cepas de Trichoderma y se llevó a una sala de propagación donde la temperatura no debía ser mayor a 28 grados. 6 días. Las dosis descritas en la Tabla 4 se utilizaron con productos de Trichoderma spp con una frecuencia de 15 días.
Se aislaron ocho cepas de Trichoderma con diferentes características morfológicas de la rizosfera de cultivares de brócoli, codificadas por el número de trampas seleccionadas y aisladas. El ADN de las cepas aisladas e identificadas se extrajo mediante la técnica de PCR, con análisis de la región ITS, utilizando los cebadores universales ITS 1 y 4, y para la identificación se utilizó el código de barras de ADN utilizando secuencias estándar de 500 a 800 pb. microorganismos en comparación con secuencias NCBI (Tabla 8). Análisis filogenético mediante la secuenciación de las regiones de los genes ITS1, ITS2 y 5.8S rDNA de ocho ecotipos de Trichoderma agrupados en dos clados principales con soporte bootstrap 96.
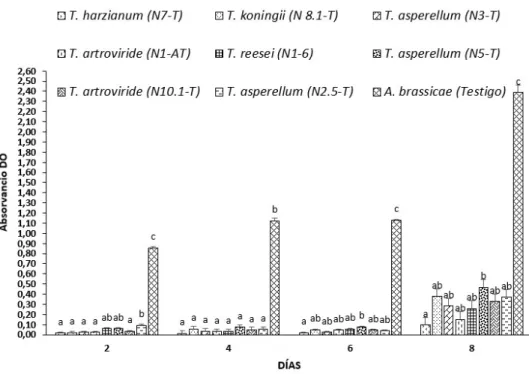
A partir del ADN genómico de los aislados identificados molecularmente se analizó la presencia del gen ech42, que codifica la enzima endoquitinasa responsable de la producción de quitinasa. En condiciones in vitro se evaluó el efecto de los metabolitos celulares de las cepas de Trichoderma identificadas: (T. harzianum, T. koningii, T. asperellum, T. ressei y T. atroviride) sobre el crecimiento micelial de A. El mayor efecto antagónico a los extractos celulares 8 días después de la inoculación con la cepa T.
Los bioproductos se prepararon a partir de aislados de Trichoderma, donde se evaluó el efecto bioestimulador sobre plántulas de brócoli en condiciones de invernadero a las 5 semanas de edad (Figura 9). T2D2 y T1D1 presentaron mayor incidencia de pudrición del pellet en 2.45 y 2.99%, donde las dosis de Trichoderma reducen su efecto antagónico sobre Alternaria en campo.

DISCUSIÓN
- Aislamiento e identificación morfológica y molecular de espe-
- Determinación de la presencia del gen ech42 en Trichoderma spp
- Aislamiento e identificación molecular del patógeno Alternaria spp . 39
- Evaluación de la capacidad protectora de Trichoderma a problemas
Entre los diferentes métodos de secuenciación de nucleótidos, aquellos que se dirigen a la región ITS tienen la mayor probabilidad de identificar con éxito la más amplia gama de hongos (Kruger et al., 2019). Activan la resistencia sistémica inducida (ISR) que es activada por las vías del jasmonato (JA) y del etileno (ET) y producen una gran cantidad de metabolitos como tricoharzanina, ácido 3-indolacético, 2,4-dimetilbenceno (Shoresh et al. , 2019). Expresa los genes LOX2 y PR-1a que están relacionados con la patogénesis y regulados por el ácido salicílico (Contreras et al., 2016).
Coincido con Gómez et al., (2018) quienes obtuvieron una amplificación de 237 pb de longitud para la cepa Trichoderma atroviride mediante el método PCR. Las enzimas quitinasa y glucanasa mejoran la resistencia de las plantas contra patógenos, expresando un gen relacionado con la patogénesis (PR) que juega un papel importante en la respuesta de defensa (Karuppiah et al., 2019). El mecanismo de acción con la antibiosis es donde se forma la mayor cantidad de metabolitos secundarios (Masi et al., 2018).
Las especies de Trichoderma tienen un gran efecto antagónico debido a la gran producción de metabolitos secundarios, los cuales se aíslan del filtrado del medio de cultivo (Hu et al., 2017). Harman et al., (2019) demostraron que la asociación de plántulas de maíz y Trichoderma mejoró el crecimiento tanto de los brotes como de las raíces de las plantas, lo que a su vez condujo a mayores rendimientos. Estos cambios incluyen principalmente la biosíntesis de reguladores del crecimiento, la fotosíntesis, la transpiración y el potencial hídrico (Brotman et al., 2015).
Además, el crecimiento de raíces y brotes en respuesta a la inoculación de Trichoderma da como resultado una mayor absorción de Cu, Na y Zn y otros micronutrientes (Li et al., 2015). Trichoderma es una de las especies competentes en la rizosfera más estudiadas porque su estrategia clave de biocontrol evoluciona en conflicto directo con los patógenos (Juliatti et al., 2019). Este complejo proceso implica eventos secuenciales que inicialmente implican el reconocimiento entre Trichoderma y el hongo patógeno y el enrollamiento alrededor de las hifas (Harman et al., 2014).
Trichoderma atroviride produce 6-pentil-2H-piranona (6-PP), un metabolito volátil que desempeña un papel fundamental en las interacciones Trichoderma-patógeno (El-Hasan et al., 2018). Coincidiendo con Ortuño et al., (2016) donde muestran que con la aplicación de Trichoderma en el cultivo de cáñamo obtuvieron mayores rendimientos debido a que Trichoderma es un gran colonizador de la rizosfera y. Asimismo, Carillo et al., (2020) quienes confirmaron que la cepa T22 provocó un aumento de licopeno en Solanum lycopersicum, un compuesto antioxidante, y aumentó el rendimiento.
CONCLUSIONES
RECOMENDACIONES
Comparative evaluation of two Trichoderma harzianum strains for major secondary metabolite production and antifungal activity. Solubilization of phosphates and micronutrients by the plant growth promoting and biocontrol fungus Trichoderma harzianum Rifai 1295-22. The Nag1 N-acetylglucosaminidase of Trichoderma atroviride is essential for chitinase induction by chitin and of major importance for biocontrol.
Application of Trichoderma harzianum, 6-Pentyl-pyrone and plant biopolymer formulations Modulate plant metabolism and fruit quality of plum tomatoes. The demon Trichoderma harzianum: complex species history resulting in the coexistence of hypothetical biological species, recent agamospecies and multiple relict lineages. Trichoderma harzianum and its metabolite 6-pentyl-alpha-pyrone suppress fusaric acid produced by Fusarium moniliforme.
Pretreatment with Trichoderma harzianum alleviates waterlogging-induced growth changes in tomato seedlings by modulating physiological, biochemical and molecular mechanisms. Molecular identification of Trichoderma spp strains, in vitro growth rate and antagonism against plant pathogenic fungi. First line defense antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defense network.
Co-culture of Trichoderma asperellum GDFS1009 and Bacillus amyloliquefaciens 1841 causes differential gene expression and improvement of wheat growth and biological control activity. Dissolution of phosphate and micronutrients by Trichoderma harzianum and its relationship to tomato plant growth promotion. Modulation of tomato response to Rhizoctonia solani by Trichoderma harzianum and its secondary metabolite harzian acid.
Trichoderma harzianum and Glomus intraradices modify the hormone disruption caused by Fusarium oxysporum infection in melon plants. The use of secondary metabolites extracted from Trichoderma for plant growth promotion in the Andean highlands. Competition for cellulose extraction between Rhizoctonia solani and two Trichoderma isolates in the decomposition of wheat straw.
TasHyd1, a novel hydrophobin gene from the biocontrol agent Trichoderma harzianum, is involved in plant root colonization. Induction of defense responses in cucumber plants (Cucumis sativus L.) by the biocontrol agent Trichoderma harzianum.