12 Avec le début de l'étude du bois mort sont apparus les premiers systèmes de classification de la décomposition, qui sont aujourd'hui largement utilisés. Enquête sur la dynamique des arbres morts en forêt boréale - La structure de la thèse et les liens entre les chapitres.

Résumé
Dans cette étude, nous avons documenté les trajectoires automnales de quatre espèces d'arbres des forêts boréales mixtes et de conifères de l'Amérique de l'Est : le peuplier faux-tremble (Populus tremuloides Michx.), le sapin baumier (Abies balsamea [L.] Mill.), le pin gris (Pinus . banksiana Lam. .) et l'épinette noire (Picea mariana [Mill.] BSP). Le tremble, le pin gris et, dans une moindre mesure, le sapin baumier ont connu des retards considérables avant que les taux de chute ne deviennent importants.
Abstract
Introduction
From a population dynamics perspective, the rate of decline represents both loss in the bottleneck population and recruitment in the stem population. Probability of survival was described with negative exponentials (e.g. Storaunet and Rolstad) or inverse sigmoid functions (e.g. Keen 1929, Garber et al. 2005).
Methods
- Study area
- Data collection
- Tree ring analysis
- Analysis of tree mortality data
- Analysis of snag survival data
- Analysis of degradation pathways
Thus, all data were either right- or left-censored (Gore et al. 1985), which precludes the use of classical analyzes such as logistic regression or survival analysis to construct parsimonious survival relationships. As with other studies on nocturnal dynamics (e.g. Boulanger and Sirois 2006, Aakala et al. 2008), uprooted trees were not included in the analyses, as it is not possible to retrospectively determine whether an uprooted tree was alive or dead before the event. was not dead. .
Results
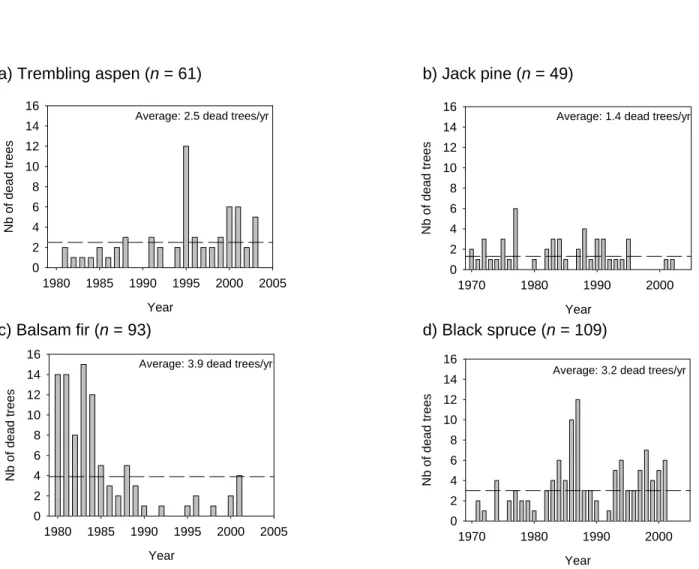
- Tree mortality
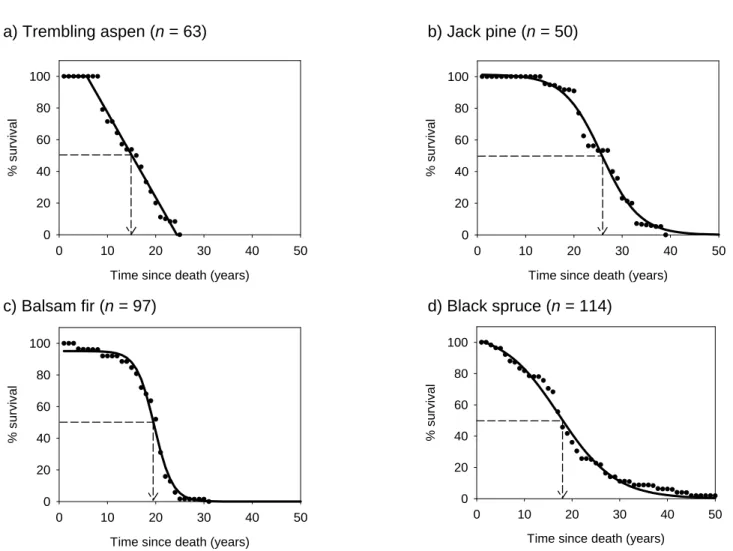
- Snag survival
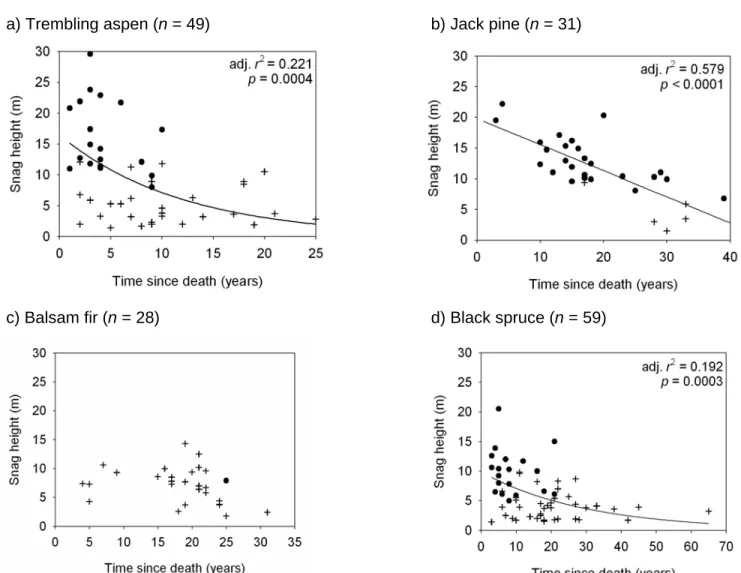
- Degradation pathway
Although balsam fir appeared to be the most brittle species, with only one out of 28 stands unbroken, stands remained fairly tall with most exceeding 5 m 20 years after death (Fig. 1.3c). In black spruce, heels were significantly shorter as time since death increased (Fig. 1.3d), while no transformation in balsam fir allowed both a significant relationship and respect for the normality of residual assumption (Fig. 1.3c).
Discussion
- Tree mortality, snag recruitment and influence of stand dynamics
- Snag survival
- Specific degradation pathway
Thus, most dead stems came from stands where balsam fir dominated forest cover and experienced high mortality before the outbreak (Bergeron et al. 1995). Other authors found negative exponential survival functions, implying a constant decline over time (Everett et al. 1999, Storaunet and Rolstad 2004).
Conclusion
Developing nature-based forestry approaches that incorporate deadwood management necessarily requires baseline data on snag fall rates for long-term planning of deadwood recruitment in managed forest landscapes (DeLong et al. 2008). Wildlife managers will need to consider habitat dynamics and therefore narrow down demography to propose recommendations and guidelines regarding deadwood conservation (e.g. Bull et al. 1997).
Acknowledgements
To effectively incorporate deadwood legacy into forest management strategies, deadwood dynamics must also be studied more thoroughly in managed systems.
Standing deadwood for keystone bird species in the eastern boreal forest: managing for roost dynamics. Coarse woody debris in the southeastern Canadian boreal forest: composition and load variations in relation to stand replacement.
Tables
Based on fire reconstructions by Dansereau (1993, Lake Duparquet Research and Teaching Forest) and Lecomte et al. 56 Table 1.2 Time since death, diameter at breast height (DBH) and comparison between DBH of sticks and logs sampled.
Figures
Dots represent the annual standing/falling cumulative ratio observed (see Methods), while curves represent the general trend.
R ÉSUMÉ
Nous avons mesuré le degré de minéralisation (c.-à-d. perte de densité du bois) de champignons de quatre espèces boréales : le peuplier faux-tremble (Populus tremuloides Michx.), le sapin baumier (Abies balsamea (L.) Mill.), le pin tordu (Pinus banksiana Lamb. ) et l'épinette noire (Picea mariana (Mill.) BSP). Le tremble a montré un taux de minéralisation plus rapide que celui des conifères (k = 0,0351).
A BSTRACT
I NTRODUCTION
Decomposer community composition and interactions between species, which are primarily fungal in boreal ecosystems, can also influence decay rates (Harmon et al. 1986; Edmonds and Eglitis 1989; Boddy 2001). Wood borers are mainly active in recently dead trees in Boreal forest ecosystems (Saint-Germain et al. 2007).
M ETHODS
- Study area
- Field methods
- Sample manipulations
- Tree ring analysis
- Data analysis
In this study, the degree of mineralization is expressed as the loss of wood density and was mainly due to respiration and leaching (Harmon et al. 1986). Despite some concerns about its ability to adequately represent mineralization in some cases (Harmon et al. 2000;
Results
- Mineralization rates
- Factors associated with mineralization rates
Since site conditions in black spruce logs differed from those of other species (see discussion), direct comparisons of mineralization rates may be misleading. Loss in wood density also progressed with time from death to the base of the stand in each species (Fig. 2.1), but mineralization rates were not significantly different from those recorded at breast height in all conifer species.
Discussion
- Mineralization rates of snags
- Modelling mineralization rates of dead wood
- Differential mineralization rates within a snag
- Factors influencing snag mineralization
In the same region, Saint-Germain et al. 2007) found similar results to those obtained in our study. Spruce stands were very open as a result of the successive paludification process (Simard et al. 2007).
Conclusion
So the fungi were more exposed to solar radiation and wind penetration, apparently increasing the drying of the wood. Additionally, black spruce snags were smaller on average than those of other species (Table 2.1), allowing the wood to dry more quickly (Rayner and Boddy 1988).
Acknowledgements
The role of the Douglas-fir beetle and wood borers in the decomposition and release of nutrients from Douglas-fir logs. Red spruce and balsam fir column decay in the White Mountains of New Hampshire.
Tables
Figure
Résumé
La mortalité après un incendie, la persistance des boucles, les schémas de chute des arbres et les variables affectant la dynamique des boucles ont été examinées. La plupart des décès sont survenus dans les deux ans suivant l'incendie, mais le processus de mortalité s'est poursuivi jusqu'à la fin de la période d'étude de dix ans.
Abstract
Introduction
Methods
- Study area
- Data collection
- Data analysis
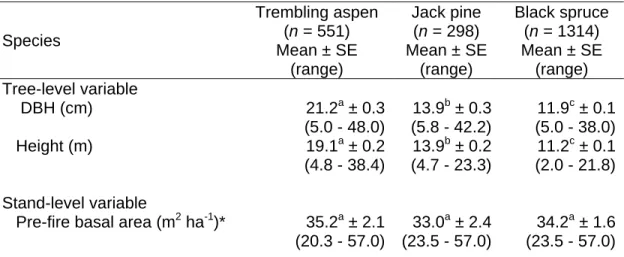
Species, DBH, height, crown length, status (alive or dead) and stem integrity (intact, broken, uprooted) were recorded in 1997, while these measurements were taken at the end of summer for the restored sites in 1998. The landscape level variable was the share of the salvaged area within a radius of 100 m from the center of the permanent plots (SALVG).
Results
- Survival curves of trees after fire
- Persistence curves of snags after fire
- Variables influencing snag persistence
- Tree fall patterns
Specifically, 22.1% of black spruce snags, 44.8% of trembling aspen snags, and 61.9% of jack pine snags were still standing at the end of the observation period. In black spruce (61%), uprooting predominated, while 81% of quaking aspen fell due to breakage at the base.
Discussion
- Survival curves of trees after fire
- Snag persistence after fire
- Factors influencing snag persistence
- Tree fall patterns
In black spruce, reported half-lives ranged from 18 to 40 years in late seral forests (Aakala et al. 2008; Chapter 1), compared to 4.4 years in our study. Burnt standards from Russell et al. 2006) study were salvage recorded using partial cuts and composed of.
Conclusion: Ecological and management implications
Spatial and temporal patterns of tree mortality can also have major implications for various structural habitat attributes for a wide range of species associated with decayed, standing, and fallen deadwood (Drapeau et al. 2009). In ecosystems where nurse logs are important for seedling establishment and development, delayed mortality and recruitment of fallen snags over time may also affect regeneration patterns (Harmon et al. 1986).
Acknowledgements
Fire impacts and crowning in the boreal forest: study of a large fire in western Quebec. In "An analysis of the global boreal forest system." (Eds HH Shugart, R Leemans, GB Bonan) p.
Tables
Moderate fire severity; H: High degree of fire severity; BA: Stand basal area (all species included); SALVG: Proportion of salvage-logged area in a radius of 100 m. Only one stem, which fell after breaking at the base, was previously dead.
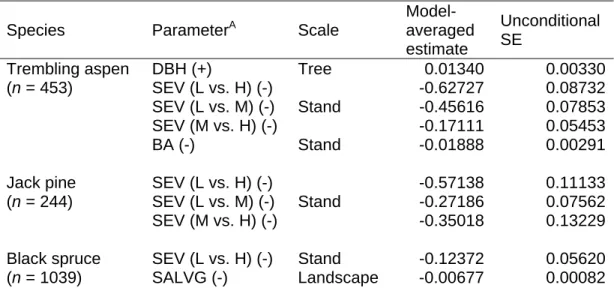
Figures
Different letters indicate significantly different survival curves between stand types as tested using the LIFEREG procedure and the log-rank test (six combinations compared simultaneously). Probability of post-fire barrier persistence over time in three species using the LIFETEST procedure and the log-rank test.
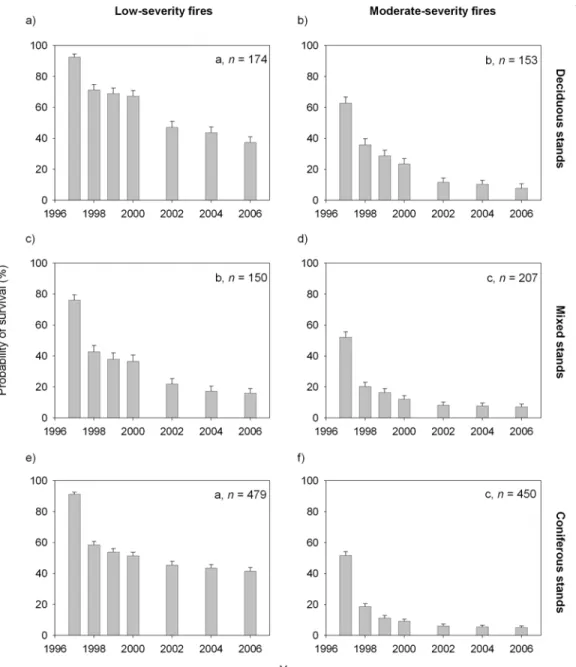
Résumé
Les systèmes de classification pour la répartition des obstructions basés sur des caractéristiques morphologiques sont largement utilisés, mais ont rarement été liés au temps écoulé depuis la mort des arbres ou à la densité du bois. Nous avons également évalué dans quelle mesure un système de classification de la dégradation des obstacles couramment utilisé représente la TDM et la densité du bois.
Abstract
Introduction
Numerous degradation classification systems have been used in North American boreal forests and relationships have been established between the stages of degradation and the age of the problem (i.e., the time elapsed since its death; Campbell and Laroque 2007, Taylor and MacLean 2007, Aakala et al. 2008), or wood density (Saint-Germain et al. 2007). For the eastern boreal forest, Imbeau and Desrochers (2002) developed a system adapted from Bergeron et al.
Methods
- Study area
- Field methods
- Tree ring analysis
- Wood density analysis
- Data analysis
To determine the year of death, each individual tree-ring series generated from all sampled transects was cross-dated with master series produced for balsam fir, pine, and aspen from nearby living trees in the LDRTF (Chapter 1) and from the master chronology , developed for black spruce (Simard et al. 2007). All sections were dried in an oven at 60 °C and weighed to the nearest 0.01 g until the weight was stable for at least 24 h.
Results
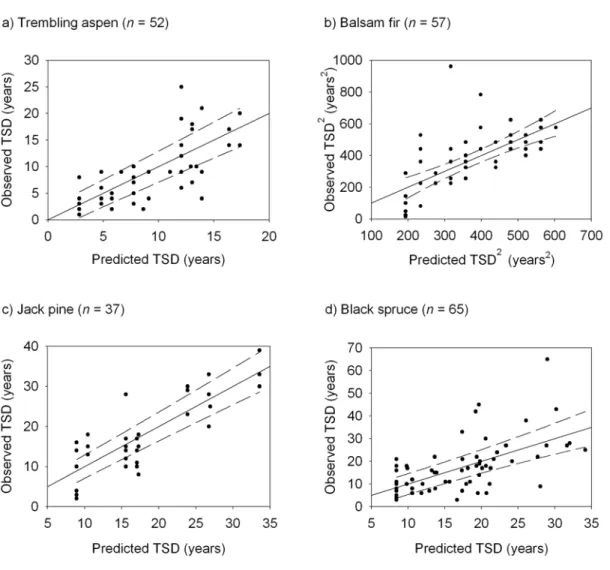
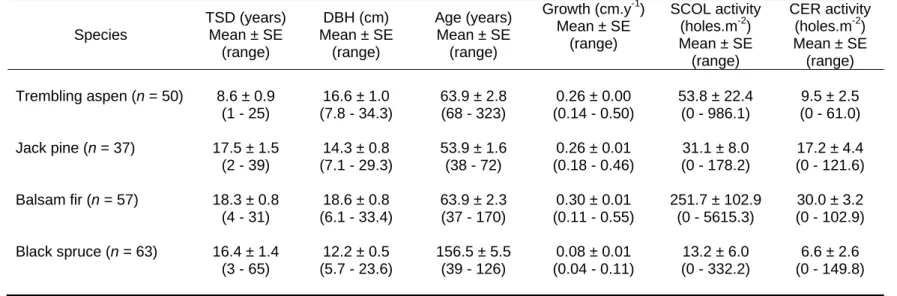
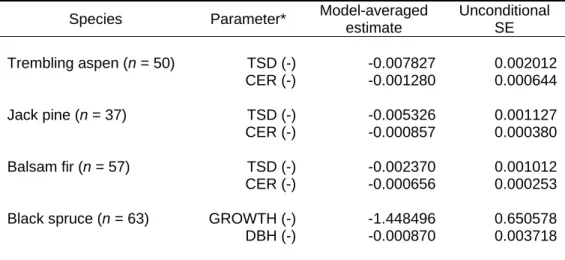
- Morphological attribute predictors of TSD and wood density
- Appropriateness of Imbeau and Desrochers’ degradation classification
- Linkages between degradation classification system, TSD, and wood
When grouping degradation stages to create significantly distinct groups, the number of classes decreased. Three grouped decomposition stages were significantly different in balsam fir (stages and Scots pine (stages whereas only two groups were significantly different in quaking aspen and black spruce p ≤ 0.0001 in all species).
Discussion
- Morphological attribute predictors of TSD and wood density
- Appropriateness of Imbeau and Desrochers’ degradation classification
Examining morphological traits predictive of TSD, Newberry et al. 2004) reported that branch residues (four categories) and fine branch flexibility (three categories) contributed significantly to the prediction of TSD. Wood decomposition, however, has been less frequently studied, especially in the eastern boreal forest of North America (Boulanger and Sirois 2006, Saint-Germain et al. 2007).
Conclusion
This has been documented in terms of decay rates (i.e. the rates at which debris falls to the forest floor, Mäkinen et al. 2006) and wood decay rates (i.e. the rates at which snag wood loses its density; Alban and Pastor 1993, Yatskov et al. 2003, Mäkinen et al. 2006). This may be due to the limitation of the decomposition system to express this process, but also the high variability in TSD and tree density, as well as the low number of observations in certain classes, which may have prevented clear differences from emerging.
Acknowledgements
The inclusion of multiple species in this study enabled us to highlight differences in the response of each species through their degradation. Although common morphological attributes were significant predictors for the two sets of models we developed, the combinations of predictor variables and the rates at which these attributes changed were species-specific.
The relationship between the relative wood density of Picea abies logs and simple classification systems of coarse decayed woody debris. Thick stands of old-growth Pinus sylvestris dominated forests along a gradient of geographic and human influence in boreal Fennoscandia.
Tables
Synthesis of candidate morphological attributes tested in models predicting time since death and wood density along with discrete variable reconfiguration. Summary of multiple linear regression statistics predicting time since death for four boreal tree species.
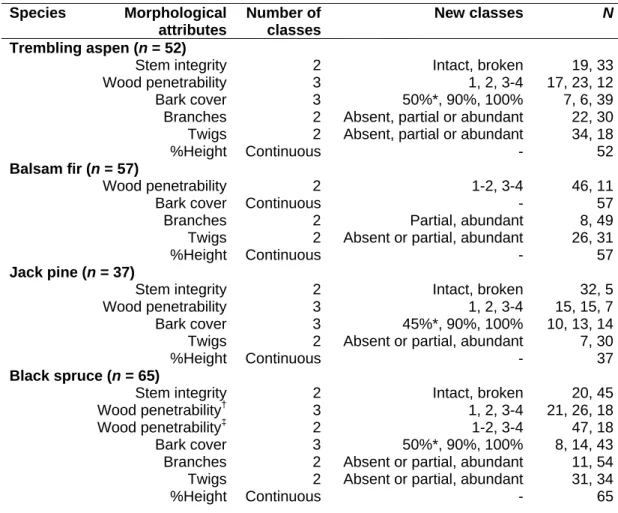
Figures
Dans les écosystèmes où les arbres morts constituent un substrat privilégié pour l'établissement et le développement des semis, la mortalité retardée et la chute des chicots au fil du temps peuvent également influencer les schémas de régénération (Harmon et al. 1986). Boulanger et Sirois 2006 et Bond-Lamberty et Gower 2008 pour l'épinette noire après incendie) et les taux de décomposition des chicots (Saint-Germain et al. 174. peuplier faux-tremble et épinette noire).