UNIVERSIDAD TÉCNICA PARTICULAR DE LOJA
La Universidad Católica de Loja
AREA BIOLÓGICA
TÍTULO DE INGENIERO EN GESTIÓN AMBIENTAL
Ensamblaje de comunidades de termitas (Blattodea: Isoptera) en
dos Bosques Tropicales Amazónicos de Ecuador, Parque Nacional
Yasuní y Parque Nacional Podocarpus
TRABAJO DE TITULACIÓN
AUTORA:
Valladares Romero Cynthia Stefanía
DIRECTOR:
Espinosa Iñiguez Carlos Iván, Ph D.
LOJA-ECUADOR
Esta versión digital, ha sido acreditada bajo la licencia Creative Commons 4.0, CC BY-NY-SA: Reconocimiento-No comercial-Compartir igual; la cual permite copiar, distribuir y comunicar públicamente la obra, mientras se reconozca la autoría original, no se utilice con fines comerciales y se permiten obras derivadas, siempre que mantenga la misma licencia al ser divulgada. http://creativecommons.org/licenses/by-nc-sa/4.0/deed.es
ii
APROBACIÓN DEL DIRECTOR DEL TRABAJO DE TITULACIÓN
Ph D.
Carlos Iván Espinosa Iñiguez
DOCENTE DE LA TITULACIÓN
De mi consideración:
El presente trabajo de titulación “Ensamblaje de comunidades de termitas (Blattodea: Isoptera) en dos Bosques Tropicales Amazónicos de Ecuador, Parque Nacional Yasuní y Parque Nacional Podocarpus”, realizado por Cynthia Stefanía Valladares Romero, ha sido orientado y revisado durante su ejecución, por cuanto se aprueba la presentación del mismo.
Loja, noviembre de 2016
iii
DECLARACIÓN DE AUTORÍA Y CESIÓN DE DERECHOS
“Yo Cynthia Stefanía Valladaes Romero declaro ser autora del presente trabajo de titulación: Ensamblaje de comunidades de termitas (Blattodea: Isoptera) en dos Bosques Tropicales Amazónicos de Ecuador, Parque Nacional Yasuní y Parque Nacional Podocarpus” de la Titulación de Gestión Ambiental , siendo el Ph D Carlos Iván Espinosa Íñiguez director del presente trabajo; y eximo expresamente a la Universidad Técnica Particular de Loja y a sus representantes legales de posibles reclamos o acciones legales. Además certifico que las ideas, conceptos, procedimientos y resultados vertidos en el presente trabajo investigativo, son de mi exclusiva responsabilidad.
Adicionalmente declaro conocer y aceptar la disposición del Art. 88 del Estatuto Orgánico de la Universidad Técnica Particular de Loja que en su parte pertinente textualmente dice: “Forman parte del patrimonio de la Universidad la propiedad intelectual de investigaciones, trabajos científicos o técnicos y tesis de grado o trabajos de titulación que se realicen con el apoyo financiero, académico o institucional (operativo) de la Universidad”
f... .
iv
DEDICATORIA
Cynthia
A la Naturaleza por llenar mi vida de colores
v
AGRADECIMIENTO
A la UTPL, la Titulación de Gestión Ambiental y el Museo de Colecciones Biológicas por los 5 años de formación académica e integral.
A la PhD. Cecilia Dahlsjö, por su motivación, colaboración incondicional y conocimientos compartidos, sin su contribución ésta investigación no podía haber surgido.
Al PhD. Carlos Iván Espinosa, Director de esta tesis, por haberme brindado la oportunidad de trabajar con él, por su guía y paciencia en el desarrollo de este estudio.
Al PhD. David Donoso, por confiar en mí y mi trabajo, con su apoyo pude desenvolverme en ésta rama, llegar a Yasuní y conocer a Cecilia.
A mi padre Ángel Gabriel y al Sr. Milton Pabón, por su compañía y asistencia en la fase de campo.
Al Ing. Diego Domínguez por su ayuda oportuna en mis dudas e inquietudes, valoro y aprecio su amistad.
Al Ministerio del Ambiente de Loja – Zamora y a la Estación Científica Yasuní, por la cooperación pertinente para la realización de éste trabajo.
A mis padres María Esther y Ángel Gabriel, ellos son el motor de mi existencia, su sacrificio y entrega lo valoro infinitamente.
A todos mis amigos, por las experiencias compartidas y alegrarme la vida con sus ocurrencias.
A la naturaleza y termitas por hacer emocionante cada día de mi investigación. Oportuna
vi
ÍNDICE DE CONTENIDOS
APROBACIÓN DEL DIRECTOR DEL TRABAJO DE TITULACIÓN ... II DECLARACIÓN DE AUTORÍA Y CESIÓN DE DERECHOS ... III DEDICATORIA ... IV AGRADECIMIENTO ... V ÍNDICE DE CONTENIDOS ... VI
RESUMEN ... 1
ABSTRAT ... 2
INTRODUCCIÓN ... 3
OBJETIVOS. ... 5
OBJETIVO GENERAL: ... 5
OBJETIVOS ESPECÍFICOS: ... 5
CAPÍTULO I ... 6
MARCO TEÓRICO ... 6
1.1Termitas – Blattodea (Isoptera) ... 7
1.1.1 Generalidades. ... 7
1.1.2 Colonia. ... 8
1.1.3 Reproducción. ... 8
1.1.4 Castas. ... 8
1.1.5 Nidos y Montículos (Estructuras). ... 9
1.1.6 Distribución. ... 9
1.1.7 Abundancia... 10
1.1.8 Diversidad. ... 10
1.2 Importancia Ecológica... 11
1.2.1 Ingenieros del Ecosistema. ... 12
1.2.2 Grupos de Alimentación... 12
1.3 Variables Microambientales. ... 13
1.3.1 Temperatura ... 13
1.3.2 Precipitaciones. ... 14
1.3.3 Altitud ... 14
1.3.4 Microhábitats ... 14
CAPITULO II ... 15
MATERIALES Y MÉTODOS ... 15
2.1 Área de estudio. ... 16
2.1.1 Parque Nacional Yasuní. ... 16
vii
2.2 Muestreo. ... 18
2.2.1 Muestreo de termitas. ... 18
2.2.2 Variables Microambientales ... 19
2.3 Fase de Laboratorio ... 20
2.3.1 Identificación ... 20
2.3.2 Hojarasca y Madera muerta. ... 20
2.4 Análisis de datos. ... 20
2.4.1 Diversidad, Riqueza y Abundancia. ... 20
2.4.2 Variables Ambientales. ... 21
CAPITULO III ... 22
RESULTADOS Y DISCUSIÓN ... 22
3.1 Resultados ... 23
3.1.1 Registro Taxonómico. ... 23
3.1.2 Riqueza y Abundancia de termitas en dos Bosques Amazónicos. ... 25
3.1.3 Variables ambientales. ... 30
3.2 Discusión. ... 31
CONCLUSIONES ... 34
RECOMENDACIONES ... 35
REFERENCIAS BIBLIOGRÁFICAS ... 36
viii
ÍNDICE DE TABLAS Y FIGURAS
Tabla 1. Clasificación de los grupos de alimentación de termitas. ... 13
Tabla 2. Datos de zonas de estudio- Parque Nacional Yasuní y Parque Nacional Podocarpus ... 18
Tabla 3. Registro de especies de termitas en Parque Nacional Yasuní (PNY) y Parque Nacional Podocarpus (PNP) ... 23
Tabla 4. Riqueza y Abundancia relativa (ocurrencias) de especies registradas en los dos Bosques Amazónicos ... 26
Tabla 5. Estimadores no paramétricos de riqueza en los dos tipos de bosque. ... 28
Tabla 6. Coeficiente de Pearson (Correlación) entre variables microclimáticas y abundancia relativa en el Parque Nacional Yasuní y Parque Nacional Podocarpus. .. 30
Figura 1. Áreas Protegidas de estudio. Parque Nacional Yasuní y Parque Nacional Podocarpus ... 16
Figura 2. Zona de estudio - Parque Nacional Yasuní, Bosque Siempreverde de Tierras bajas ... 17
Figura 3. Zona de estudio- Parque Nacional Podocarpus- Sector Bombuscaro ... 17
Figura 4. Método estandarizado según Jones & Eggleton (2000) ... 19
Figura 5. Abundancia relativa en los transectos de los dos sitios de estudio. ... 26
Figura 6. Curvas de rarefacción de especies. ... 27
Figura 7. Microhábitats ocupados por termitas en el Parque Nacional Podocarpus (PNP) y Parque Nacional Yasuní (PNY), en proporción al número total de encuentros. ... 28
Figura 8. Grupos funcionales de termitas en el Parque Nacional Yasuní y Parque Nacional Podocarpus, en proporción al número de ocurrencias. ... 29
1
RESUMEN
Las termitas alcanzan altos niveles de diversidad, abundancia y biomasa, no obstante el conocimiento de la fauna de termitas en los bosques tropicales de Ecuador es incompleto debido a la falta de estudios. En la presente investigación se realizó el estudio del ensamblaje de comunidades de termitas (Blattodea: Isoptera) en dos Bosques Tropicales Amazónicos de Ecuador, en el Parque Nacional Yasuní y Parque Nacional Podocarpus. Las termitas fueron muestreadas y colectadas mediante el protocolo rápido de estimación de termitas, mediante transectos de 100 x 2 metros, divididos en 20 secciones. En los dos Bosques Amazónicos se registró una riqueza de 23 géneros y 69 especies de termitas, el Parque Nacional Yasuní registra el 81,15% de la riqueza total con 56 especies y el Parque Nacional Podocarpus el 34,78 % con 24 especies, compartiéndose 11 especies entre los dos bosques. Finalmente se analizó la respuesta del ensamblaje de termitas a las variables microclimáticas, dando como resultado una relación no significativa, a excepción de la variable “Madera seca y hojarasca” que mostró una relación positiva en la abundancia de termitas en el Parque Nacional Podocarpus.
2
ABSTRAT
Termites reach high levels of diversity, abundance and biomass, yet knowledge of termite fauna in tropical forests in Ecuador is incomplete due to lack of studies. In the present investigation the study of the assemblage of termite communities (Blattodea: Isoptera) in two Tropical Rainforests of Ecuador, in the Yasuní National Park and Podocarpus National Park was carried out. Termites were sampled and collected using the rapid termite estimation protocol, using transects of 100 x 2 meters, divided into 20 sections. In the two Amazon Forests there was a wealth of 23 genera and 69 species of termites, Yasuní National Park recorded 81.15% of the total richness with 56 species and Podocarpus National Park 34.78% with 24 species, sharing 11 species between the two forests. Finally, the response of the termite assemblage to the microclimatic variables was analyzed, resulting in a non-significant relationship, except for the variable "Dry wood and leaf litter", which showed a positive relation in the termite abundance in the Podocarpus National Park.
3
INTRODUCCIÓN
El Ecuador con una extensión de 256.370 km2, es catalogado como uno de los países más ricos en diversidad biológica debido a tres factores: su ubicación en el Neotrópico, la presencia de la cordillera de los Andes y la influencia de las corrientes marinas. (Mittermejer et al., 1997). Estos factores nos confieren una considerable biodiversidad a nivel genético, de especies y ecosistemas; constituyendo un tesoro biológico que es necesario preservar y conservar.
Dentro del contexto de biodiversidad, los invertebrados en Ecuador constituyen un grupo muy amplio, no se sabe con exactitud cuántas y cuáles son las especies que se encuentran en el país. Los diversos estudios de este grupo están en su mayoría enfocados a los que poseen importancia económica, siendo muy pocos los estudios que destacan aspectos de diversidad o endemismo (MAE, 2010). Entre la variedad de invertebrados los insectos comprenden el grupo de animales más diverso y numeroso del reino animal, aunque su número varía según las fuentes consultadas (Adller & Foottit, 2009; Blas & Del Hoyo, 2013).
Considerando que los insectos componen una parte muy importante de la biodiversidad (Wilson, 1992; Morrone et al., 1999) se prevé que los insectos sociales que corresponden al orden Blattodea, infraorden: Isoptera o comúnmente llamadas termitas, alcanzan altos niveles de diversidad, abundancia y biomasa (Eggleton et al., 1996, 1999; Bignell & Eggleton, 2000; Dahlsjö et al., 2014a). Este grupo de insectos se constituyen en organismos esenciales por el rol en la dinámica de los ecosistemas, principalmente tropicales (Nickle & Collins, 1992). Uno de estos roles es la descomposición de material vegetal muerto, que va desde la madera no humificada a material humicado (Donovan et al., 2001).
4 En los siguientes capítulos se encontrará una breve descripción de las generalidades, diversidad, abundancia e importancia ecológica. Conjuntamente con los métodos empleados para la obtención y análisis de datos de ensamblaje de la comunidad de termitas, así como la relación con las variables microclimáticas. Concluyendo se presenta los resultados obtenidos y se los confronta con investigaciones afines.
5
OBJETIVOS.
Objetivo General:
Evaluar el ensamblaje de comunidades de termitas (Blattodea: Isoptera) en dos Bosques Tropicales Amazónicos de Ecuador, Parque Nacional Yasuní y Parque Nacional Podocarpus.
Objetivos Específicos:
Examinar la diversidad, riqueza y abundancia de una comunidad de termitas en dos Bosques Tropicales Amazónicos de Ecuador, Parque Nacional Yasuní y Parque Nacional Podocarpus.
6
CAPÍTULO I
7
1.1 Termitas – Blattodea (Isoptera)
1.1.1 Generalidades.
Las termitas son insectos sociales que corresponden al orden Blattodea, infraorden: Isoptera. A veces reciben el nombre de “hormigas blancas” debido a las semejanzas en su organización social, sin embargo estos organismos no están relacionados con las hormigas sino con las cucarachas (McGavin, 2002).
Tienen un cuerpo blando, con un tamaño de 3 -10 mm de largo, (Artigas, 1994; Camousseight, 1999; Ramirez & Lanfranco, 2001). Poseen antenas multisegmentadas, mandíbulas en el aparato bucal, ojos ausentes o degenerados y alas presentes solo en reproductores (Gillot, 2005; Sermeño-Chicas, 2008). Tienen por lo general colores claros que varían de acuerdo al alimento consumido, debido a que su aparato digestivo suele reflejarse a través del cuerpo (Cammousseight, 1999; Ebeling, 2000;). Además las termitas producen hormonas y feromonas que les permite reconocerse entre miembros de la misma colonia, comunicarse y advertir de algún tipo de invasor, permitiendo una mejor organización y mantenimiento de la sociedad (Ramirez & Lanfranco, 2001).Su alimentación varía desde suelo, tejidos vegetales, hojarasca y madera muerta en todas sus etapas de descomposición. Por lo tanto, algunas termitas tienen efectos específicos en la fertilidad y estructura del suelo (Lee &Wood, 1971; Darlignton et al., 1997), siendo los insectos descomponedores de materia orgánica más dominantes en las regiones tropicales y subtropicales (Bignell & Eggleton, 2000; Dahlsjö et al., 2014a).
Las termitas se clasifican en dos grupos:
8
1.1.2 Colonia.
Las termitas viven en colonias sociales permanentes (McGavin, 2002), las colonias varían desde unos cientos a 6 millones de individuos (Lavelle & Spain, 2001). Por lo general, la colonia posee una parte inanimada y una animada. La parte animada corresponde a los individuos que habitan dentro de la colonia y la parte inanimada a las estructuras construidas por los habitantes de la colonia (Eggleton, 2011; Bignell et al., 2011a). La colonia está formada de una pareja de reproductores, del 0 – 25% son soldados y el resto son obreras (Davies et al., 2003a; Scheffrahn, 2008; Sermeño-Chicas, 2008).
Bignell et al., (2011a) mencionan que dentro de las funciones de la colonia se consideran las siguientes:
- Reproducción y dispersión (Reina, rey, alados)
- Construcción, alimentación y cuidado del nido ( obreras) - Defensa (soldados)
- Protección y fortificación de nidos (obreras)
1.1.3 Reproducción.
Las termitas se caracterizan por presentar un ciclo de vida con metaformosis incompleta, poseen las fases de huevo, ninfa y adulto (Camousseight, 1999; Ramirez & Lanfranco, 2001). Los reproductores se unen con sus parejas, se produce el cortejo, ocurre la autonomía (corte de alas), seguidamente la pareja construye el copulario, se produce la cópula y los primeros huevos, originando un nuevo nido y una nueva sociedad (Camousseight, 1999; Union Sercices, 2000; Ramirez &Lanfranco, 2001).
1.1.4 Castas.
9
1.1.4.1 Reproductores (Reinas, Rey, Alados).
La reina es el único individuo que pone huevos en la colonia y el rey es su compañero con el que la reina se aparea regularmente (Korb, 2008; Eggleton, 2011; Bignell et al., 2011a). Los alados son reproductores que abandonan el nido en enjambre con el fin de dispersarse e iniciar nuevas colonias (Eggleton, 2011; Bignell et al., 2011a)
1.1.4.2 Soldados.
Los soldados son individuos estériles, algunos poseen defensas químicas (Prestwich, 1984; Eggleton, 2011; Bignell et al., 2011a), su principal función es defender a la colonia en particular a la reina y el rey (Eggleton, 2011; Bignell et al., 2011).
1.1.4.3 Obreras.
Las obreras nunca dejan el nido, excepto cuando buscan alimento. Son el pilar fundamental de la colonia y sus tareas se enfocan en la alimentación, cuidado y construcción-reparación de colonias (Eggleton, 2011; Bignell et al., 2011a)
1.1.5 Nidos y Montículos (Estructuras).
Los nidos y montículos son estructuras elaboradas por las colonias de termitas, cumplen funciones de protección contra factores ambientales, mantienen estable el clima interno y lo fortifican contra los depredadores (Eggleton, 2011; Bignell et al., 2011a). Las colonias de termitas viven dentro de los nidos y si el nido está cubierto o hecho con tierra y sobresale por encima del suelo se convierte en un montículo. Sin embargo, no todos los nidos son visibles debido a que muchas colonias se encuentran bajo el suelo, especialmente en los bosques húmedos tropicales (Eggleton et al., 1996).
1.1.6 Distribución.
10 2000).No obstante, se ha reportado la presencia de éstos organismos y poblaciones densas de montículos del género Cubitermes en altitudes elevadas entre los 3000 y 4000 m snm. Aun así, no hay investigaciones cuantitativas que respalden estás observaciones (Darlington, 1985; Eggleton, 2000; Donovan et al., 2002). Sin embargo Gathorne-Hardy et al., (2001) establecen que las diferencias continentales y su relación con los tipos de ecosistemas, determina que la distribución de termitas a nivel altitudinal puede variar.
1.1.7 Abundancia
Las termitas abundan particularmente en bosques lluviosos tropicales de tierras bajas (Bignell & Eggleton, 2000; Dahlsjö et al., 2014b). Pueden constituir el 10% de toda la biomasa animal en los trópicos y el 95% de la biomasa de los insectos del suelo (Wilson, 1992; Watt et al., 1997; Bignell & Eggleton, 2000). Pese a ello su abundancia disminuye con el aumento de la altitud (Eggleton, 2000).
En muchos ecosistemas tropicales sus nidos y montículos dominan el paisaje, y pueden superar a casi todos los grupos de insectos en número de individuos o en biomasa (Bandeira & Vasconcellos, 1999). Dentro de los bosques húmedos alcanzan densidades que pueden superar los 8000 individuos / m2 (Fittkau & Klinge, 1973;
Martius, 1994; Bignell & Eggleton, 2000). Eggleton et al., (1996) manifiestan que la biomasa y abundancia de termitas en bosques tropicales pueden exceder los 100g/m2 y 10,000 individuos/m2 respectivamente. No obstante, las estimaciones de abundancia dependen de los protocolos de monitoreo.
1.1.8 Diversidad.
1.1.8.1 Diversidad Global.
11 de los bosques neotropicales de lluvia con 55 géneros y luego las selvas tropicales de Asía en el Norte de Borneo con 44 géneros (Eggleton, 2011; Bignell et al., 2011a).
1.1.8.2 Diversidad en el Neotrópico.
La diversidad de termitas en el Neotrópico es muy importante, siendo la segunda región zoogeográfica en número de especies (Constantino, 1998). A pesar de los hábitos crípticos y subterráneos de muchas especies, el conocimiento de la taxonomía y biología de algunos grupos de termitas es incipiente y se encuentra incompleto (Costa, 1997; Roisin et al., 1996). En la Amazonia el conocimiento de la fauna de termitas es pequeño, en comparación con las selvas tropicales del Viejo Mundo (Martius, 1994; Ackerman et al., 2009). Mientras que para el Neotrópico comprende el 42% de todas las especies de termitas conocidas con cerca de 55 géneros registrados y 543 especies, para Sudamérica se reconocen 400 especies de termitas (Constantino, 1998).
1.1.8.3 Diversidad en Ecuador.
En Ecuador, el único estudio sobre fauna de termitas en ambientes naturales ha sido realizado por Bahder et al., (2009) registrando una diversidad de 77 morfo-especies, de las cuales 18 especies no pudieron ser identificadas. Actualmente la falta de conocimientos sobre la fauna de termitas está relacionada con la falta de inventarios, concretamente en las tierras bajas del Este y en los bosques nublados de la Cordillera de los Andes (Bahder et al., 2009)
1.2 Importancia Ecológica.
12 Si bien son más reconocidas como destructoras de madera y de material celulósico, estos organismos son muy favorecedores en la dinámica de los ecosistemas, particularmente tropicales (Nickle & Collins, 1992; Sermeño-Chicas, 2008). Cumplen una función importante como consumidores primarios de celulosa y descomponedores de materia orgánica, madera y mantillo (Takuya & Masahiko, 2001; Vargas et al., 2005), gracias al ciclaje de la materia orgánica efectuado por acción de las termitas existe un eficiente retorno de nutrientes a la vegetación, dotando de estructura y fertilidad al suelo (Wood & Sands, 1978; Constantino & Acioli, 2006; Ackerman et al., 2009; Abadía et al., 2013). Bignell & Eggleton (2000) mencionan que hasta el 50% de la descomposición de los residuos orgánicos de origen vegetal en los bosques tropicales pueden ser dotados por las termitas.
1.2.1 Ingenieros del Ecosistema.
Los ingenieros del ecosistema son organismos que directa o indirectamente modifican la disponibilidad de los recursos a otras especies, ocasionando variaciones en el estado físico del medio biótico y abiótico (Jones et al., 1994; 1997). Dentro de los ecosistemas tropicales las termitas cumplen un papel clave como "ingenieros del ecosistema" (Jones et al., 1994; Lavelle et al., 1997) y son probablemente los organismos más importantes en calidad de agentes de la bioturbación y ciclaje de nutrientes del suelo (Bignell, 2006; Jouquet et al., 2011; Dahlsjö et al., 2014a). Las termitas mientras se alimentan oxigenan el suelo y aumentan su porosidad, de esta manera el agua se filtra con más facilidad y se amplía el hábitat disponible para otros organismos (Union Europea, 2010). La dinámica resultante de sus actividades de alimentación y construcción en el suelo cumplen un papel regulador insustituible, compensando el empobrecimiento de la superficie edáfica atribuida por la erosión mecánica y lixiviación (Miklós, 1998; De Franca Alves, 2009).
1.2.2 Grupos de Alimentación.
13 comportamiento alimenticio por ello es importante identificar los grupos de alimentación y clasificarlos con el fin de entender la ecología y procesos en que participan (Donovan et al., 2001), (Tabla 1),
Tabla 1. Clasificación de los grupos de alimentación de termitas.
Grupos de Alimentación
Descripción
Grupo I Termitas inferiores (Familias Kalotermitidae y Rhinotermitidae). La mayoría se alimenta de madera, en particular vegetal vivo o muerto.
Grupo II Termitas de la Familia Termitidae. Se alimentan desde madera, hierbas, hojarasca y micro epifitos.
Grupo III
Termitas de la familia Termitidae. Se consideran “consumidores de suelos ricos en materia orgánica”, se alimentan del suelo con alto contenido de
materia orgánica y madera descompuesta que ha perdido su estructura.
Grupo IV
Termitas de la familia Termitidae. Se consideran los “verdaderos consumidores de suelo”, se alimentan en suelo mineral con bajo
contenido de materia orgánica .
Fuente: Donovan et al., (2011). Modificada por la autora
1.3 Variables Microambientales.
Las termitas muestran una alta sensibilidad a las condiciones ambientales, tanto bióticas como abióticas, así como los procesos de los ecosistemas (Jones & Eggleton, 2000). Los principales desafíos ambientales de las colonias de termitas son las precipitaciones y la temperatura. Dentro de las selvas tropicales las temperaturas se almacenan, pero las lluvias pueden ser torrenciales y continúas (Eggleton, 2011a).
1.3.1 Temperatura
14 de manera diferente a la temperatura debido a sus diferentes hábitos de alimentación (Jones, 2000; Davies et al., 2003b; Palin et al., 2011). Las termitas que se alimentan del suelo dependen de los sustratos de alimentos energéticos más bajos siendo afectadas negativamente por las bajas temperaturas del suelo (Jones, 2000; Davies et al., 2003a; Girardin & Rozas, 2011).
1.3.2 Precipitaciones.
Los cambios en la diversidad de termitas están influenciados por las precipitaciones, particularmente en los bosques lluviosos tropicales pueden tener un efecto negativo en la riqueza y abundancia de especies de termitas, debido a que los niveles muy altos de precipitaciones pueden desencadenar inundaciones en los microhábitats y la consecuente muerte de colonias (Dibog et al., 1998; Bignell & Eggleton, 2000).
1.3.3 Altitud
Las termitas están preferentemente distribuidas en bosques tropicales, siendo más diversas en bosques ecuatoriales (Collins, 1983). Pero habitualmente su riqueza, abundancia y biomasa disminuye mientras hay un aumento en la altitud, similar a la mayoría de invertebrados de bosques tropicales (Eggleton, 2000). La razón se debe a que las termitas son ectotérmicas y mientras existan una mayor altitud la temperatura desciende, por tanto una notable disminución en su riqueza y abundancia (Jones, 2000; Gathorne-Hardy et al., 2001)
1.3.4 Microhábitats
15
CAPITULO II
16
2.1 Área de estudio.
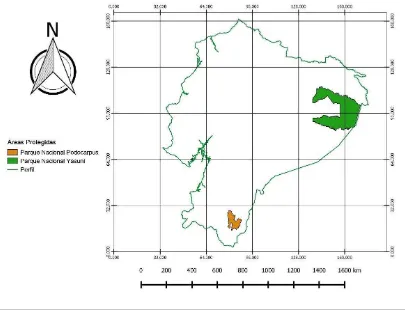
[image:25.595.97.502.169.479.2]El presente estudio se desarrolló en dos Áreas Protegidas del Ecuador, en el Parque Nacional Yasuní y Parque Nacional Podocarpus (Figura 1).
Figura 1. Áreas Protegidas de estudio. Parque Nacional Yasuní y Parque Nacional Podocarpus
2.1.1 Parque Nacional Yasuní.
17 Figura 2. Zona de estudio - Parque Nacional Yasuní, Bosque Siempreverde de Tierras bajas
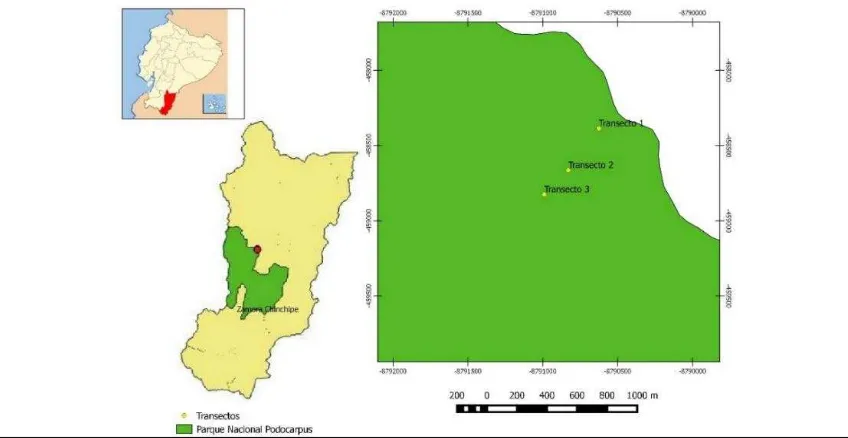
2.1.2 Parque Nacional Podocarpus
El segundo sitio de estudio corresponde al Parque Nacional Podocarpus (PNP) (Figura 3) concretamente en el Sector Bombuscaro, ubicado en el cantón Zamora, Provincia de Zamora Chinchipe, Ecuador. El sitio posee un clima con temperaturas que fluctúan de 21°C a 29°C y precipitaciones de 1350 a 2261 mm/año. La vegetación del sector Bombuscaro corresponde a Bosque siempre verde montano bajo con una altitud de 1000 a 1900 m snm (Sierra, 1999; MAE, 2013; Plan de Manejo de Parque Nacional Podocarpus, 2014).
[image:26.595.86.510.529.748.2]18
2.2 Muestreo.
2.2.1 Muestreo de termitas.
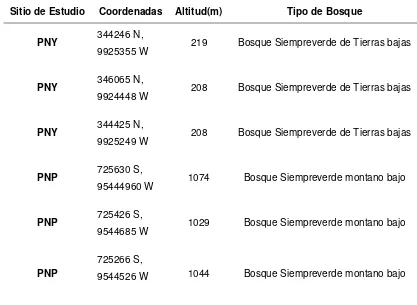
En los dos bosques de estudio se establecieron 6 puntos de muestreo, para el Parque Nacional Yasuní (PNY) tres puntos de muestreo en Mayo/2016 y para el Sector Bombuscaro, Parque Nacional Podocarpus (PNP), tres puntos en Febrero/2016. Cada punto clave estuvo espaciado con una distancia de 300 – 500 metros (Tabla 2).
[image:27.595.85.502.270.575.2]
Tabla 2. Datos de zonas de estudio- Parque Nacional Yasuní y Parque Nacional Podocarpus
Sitio de Estudio Coordenadas Altitud(m) Tipo de Bosque
PNY 344246 N,
9925355 W 219 Bosque Siempreverde de Tierras bajas
PNY 346065 N,
9924448 W 208 Bosque Siempreverde de Tierras bajas
PNY 344425 N,
9925249 W 208 Bosque Siempreverde de Tierras bajas
PNP 725630 S,
95444960 W 1074 Bosque Siempreverde montano bajo
PNP 725426 S,
9544685 W 1029 Bosque Siempreverde montano bajo
PNP
725266 S,
9544526 W 1044 Bosque Siempreverde montano bajo
19 ocurrencia de una colonia de termitas de una especie en un punto de excavación) fue colectado y registrado (Davies et al., 2003a).
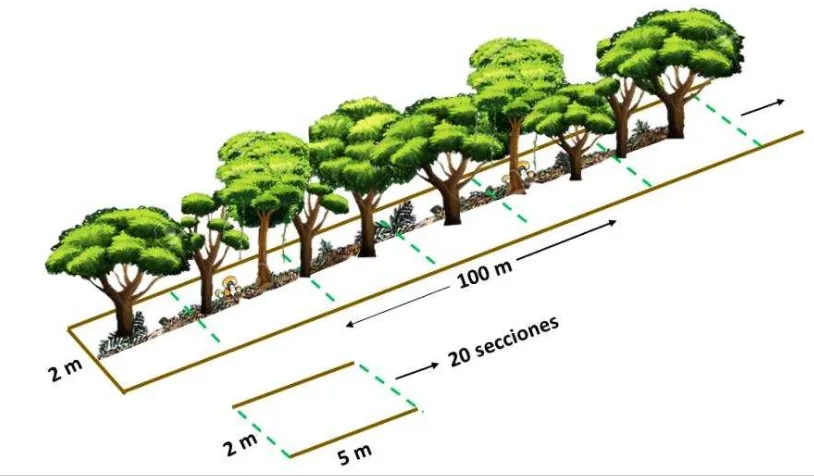
El método de muestreo se basó en un transecto de 100 metros dividido en 20 cuadrantes continuos, con dimensión de 5 × 2 , en cada cuadrante se colectó los representantes de termitas (varios individuos entre trabajadores y soldados, si están presentes). Se recogió de forma activa y manual por una hora/persona en todos los microhábitats de termitas, se incluyó el suelo, madera en descomposición, hojarasca, nidos, también para registrar la ocurrencia de especies de termitas en el suelo, en cada cuadrante se colectó doce muestras de suelo superficial ( ×
[image:28.595.93.500.366.604.2]� � � � � y se exploró la existencia de termitas. Inmediatamente todas las termitas se almacenaron en alcohol al 80% y se etiquetaron con el número de cada cuadrante y transecto.
Figura 4. Método estandarizado según Jones & Eggleton (2000) Fuente: Autora
2.2.2 Variables Microambientales
20 movilizó en bolsas plásticas a los laboratorios del Instituto de Ecologia- UTPL y Estación Científica Yasuní – PUCE con el fin de evaluar el peso húmedo y seco
Seguidamente en cada cuadrante se estimó la radiación solar y cobertura del dosel mediante fotografías hemisféricas (ojo pez), los datos de altitud con un GPS GARMIN y datos de pendiente con ayuda de un clinómetro.
2.3 Fase de Laboratorio
2.3.1 Identificación
Para la identificación de especies de termitas se contó con la colaboración de una especialista en éste taxón Ph.D Cecilia Dahlsjö (Oxford, Inglaterra). Los soldados termitas se identificaron a nivel de género utilizando la clave para géneros neotropicales de Constantino (2002), mientras que las especies fueron identificadas utilizando claves para géneros específicos (Bourguignon & Roisin, 2011) o a través de la colección de referencia en el Museo de Historia Natural (NHM) en Londres. Las especies de termitas soldados y castas de obreras se identificaron por su morfología intestinal y estructuras de válvulas entéricos descritos en Bourguignon et al., (2010, 2015), y en la clave inédita de LM Hernández (no publ.).
2.3.2 Hojarasca y Madera muerta.
La hojarasca y madera muerta colectada en el Parque Nacional Podocarpus y Parque Nacional Yasuní, se pesó en estado húmedo en una balanza electrónica, luego se la secó en la estufa a una temperatura de 80° C durante 48 horas, posterior se pesó en estado seco (Anexo 2).
2.4 Análisis de datos.
2.4.1 Diversidad, Riqueza y Abundancia.
21 de especies (número de especies), abundancia relativa (número total de apariciones especie por cuadrante muestreado) y diversidad (índices de diversidad).
Para determinar la eficacia de la estimación de la diversidad de especie con el método de muestreo, se estableció curvas de rarefacción para generar la acumulación de especies con respecto al esfuerzo de muestreo y la riqueza de especies observadas, para lo que se utilizó el paquete ‘vegan’ implementado por el entorno de programación R (R Development Core Team, 2016)
Para estimar la riqueza y estructura de la comunidad de termitas se utilizaron estimadores no paramétricos de riqueza como ACE 1, que utiliza datos de abundancia para cuantificar la rareza de especies y CHAO 1 que estima la riqueza de especies que solo aparecen en una muestra y la riqueza de especies que son compartidas en dos muestras (Colwell et al., 1994; Magurran, 2004) todos estos métodos estadísticos son implementados por el entorno de programación R (R Development Core Team, 2016).
Para determinar cuán diverso es un sitio se utilizó el índice de diversidad Simpson el cual muestra la probabilidad de que dos individuos sacados al azar de una muestra correspondan a la misma especie (Marrugan, 2001) y el Índice de similitud de Jaccard que relacionan el número de especies compartidas con el número total de especies exclusivas (Southwood, 1978; Krebs, 1998), éstos índices fueron implementados por el entorno de programación R (R Development Core Team, 2016), usando el paquete ‘vegan’ (Oksanen et al., 2011).
2.4.2 Variables Ambientales.
Para determinar la asociación entre las variables microambientales y el ensamblaje de comunidad de termitas, se utilizó el coeficiente de correlación de Pearson, usando el paquete ‘Stat’ en el entorno de programación R (R Development Core Team, 2016).
22
CAPITULO III
23
3.1 Resultados
En los dos Bosques Amazónicos de Ecuador se registró una riqueza total de 69 especies de termitas distribuidas en 23 géneros. El 63,75% de la termitofauna registrada corresponde al Parque Nacional Yasuní con un total de 16 géneros y 44 especies, el 20,5% al Parque Nacional Podocarpus con un registro de 4 géneros y 14 especies y el 15,94% restante corresponde a las especies compartidas entre los dos bosques con un total de 7 géneros y 11 especies.
3.1.1 Registro Taxonómico.
[image:32.595.82.509.561.773.2]Utilizando la metodología propuesta por Jones & Eggleton (2000), en el Parque Nacional Yasuní en el muestreo Mayo/2016, las termitas colectadas corresponden a dos familias (Termitidae y Rhinotermitidae), cuatro subfamilias de Termitidae (Apicotermitinae, Nasutitermitinae, Termitinae y Syntermitinae), y dos subfamilias de Rhinotermitidae (Heterotermitinae y Rhinotermitinae). Las dos familias registraron 20 géneros y 56 especies de termitas, mientras que en el muestreo realizado en Febrero/2016 en el Parque Nacional Podocarpus, en los tres transectos establecidos la colecta de termitas pertenecen a dos familias (Termitidae y Rhinotermitidae), cuatro subfamilias de Termitidae (Apicotermitinae, Nasutitermitinae, Termitinae y Syntermitinae), y a dos subfamilias para Rhinotermitidae ( Heterotermitinae y Rhinotermitinae), las dos familias registraron 11 géneros y 24 especies de termitas (Tabla 3).
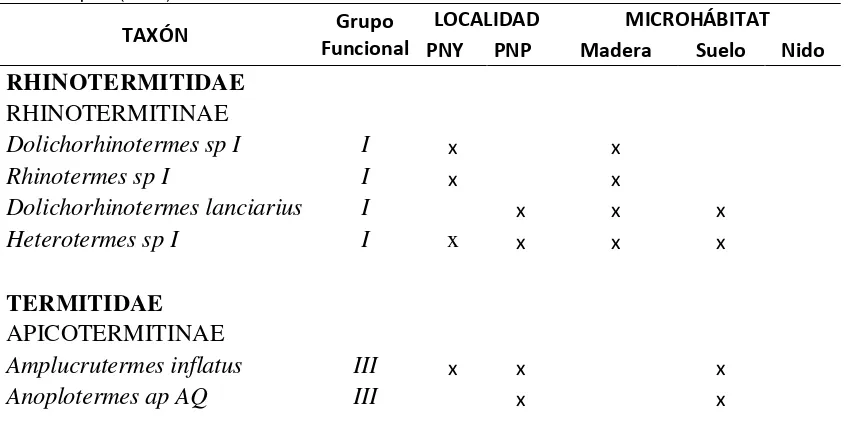
Tabla 3. Registro de especies de termitas en Parque Nacional Yasuní (PNY) y Parque Nacional Podocarpus (PNP)
TAXÓN Grupo
Funcional
LOCALIDAD MICROHÁBITAT
PNY PNP Madera Suelo Nido
RHINOTERMITIDAE
RHINOTERMITINAE
Dolichorhinotermes sp I
I
x xRhinotermes sp I
I
x xDolichorhinotermes lanciarius
I
x x xHeterotermes sp I
I
x
x x xTERMITIDAE
APICOTERMITINAE
Amplucrutermes inflatus
III
x x x24
Anoplotermes banksi
III
x x x x xAnoplotermes grp AC
III
x xAnoplotermes grp AD
III
x xAnoplotermes grp AG
III
x xAnoplotermes grp AM
III
x xAnoplotermes grp AP
III
x xAnoplotermes grp DE
III
x xAnoplotermes grp EL
III
x x xAnoplotermes grp JU
III
x xAnoplotermes grp L
III
x xAnoplotermes grp NA
III
x xAnoplotermes grp OI
III
x xAnoplotermes grp RH
III
x x xAnoplotermes grp TUC
III
x x xAnoplotermes grp XY
III
x xAnoplotermes sp AB
III
x x x x xAnoplotermes sp BU
III
x x xAnoplotermes sp F
III
x x x xAnoplotermes sp K
III
x xAnoplotermes sp V
III
x x x xAnoplotermes sp XY
III
x xx xEmbiratermes neotenicus
III
x x xEmbiratermes sp I
III
x x xEmbiratermes sp IV
III
x x xHumutermes krishnai
III
x x x x xHydrotermes ariensho
III
x xHydrotermes kawaii
III
x xPatawatermes nigripunctatus
III
x x x xRuptitermes sp A
III
x xNASUTITERMITINAE
Atlantitermes snydari
III
x x xAtlantitermes sp II
III
x xEreymatermes sp I
III
x x xNasutitermes sp I
II
x xNasutitermes sp II
II
x xNasutitermes sp III
II
x x xNasutitermes sp IV
II
x xNasutitermes sp V
II
x xNasutitermes sp VI
II
x x x xNasutitermes sp VII
II
x x x xNasutitermes sp VIII
II
x x x xNasutitermes sp X
II
x x25
Nasutitermes sp XII
II
x x x xNasutitermes sp XIII
II
x xNasutitermes sp XIV
II
x xSubulitermes sp I
III
x x xSubulitermes sp II
III
x x xConstrictotermes sp I
III
x xSYNTERMITINAE
Silvestritermes sp I
III
x xSilvestritermes sp IV
III
x x xSilvestritermes sp II
III
x x x xSilvestritermes sp III
III
x x x xEmbiratermes neotenicus
III
x xEmbiratermes sp I
III
x xTERMITINAE
Crepititermes verruculosus
III
x xCylindrotermes flangiatus
II
x x x xCylindrotermes saprianga
II
x x x xCyrillotermes sp I
III
x xNeocapritermes talpa
III
x xNeocapritermes talpoides
III
x xTermes sp I
III
x xTriangulartermes spII
III
x x xCornicapritermes sp I
III
x xCylindrotermes macrognathus
II
x x x x3.1.2 Riqueza y Abundancia de termitas en dos Bosques Amazónicos.
Dentro del estudio una diferencia muy marcada se presenció en el total de encuentros (abundancia relativa) durante el muestreo de los dos bosques. En los tres transectos establecidos en el Parque Nacional Podocarpus hubo una menor ocurrencia de termitas en comparación con los transectos en el Parque Nacional Yasuní.
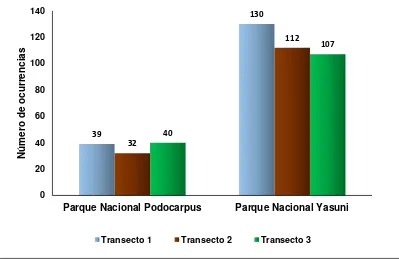
26 Figura 5. Abundancia relativa en los transectos de los dos sitios de estudio.
Una alta diversidad fue encontrada en los dos bosques amazónicos, en el Parque Nacional Yasuní D = 0.93) y Parque Nacional Podocarpus (Simpson_1-D= 0.689) (Tabla 4).
Tabla 4. Riqueza y Abundancia relativa (número de encuentros por cada especie) de especies
registradas en los dos Bosques Amazónicos.
Bosque Parque Nacional Podocarpus (Bosque Siempreverde Montano bajo)
Parque Nacional Yasuní (Bosque Siempre verde de tierras
bajas)
Abundancia Relativa 111 349
Riqueza 24 56
Diversidad (Simpson 1-D) 0,63 0,93
El índice de similitud de Jaccard expresa un valor de disimilitud de (J =0.27), determinando que el 73% de riqueza de especies entre los dos parques tienen similitud.
39
130
32
112
40
107
0 20 40 60 80 100 120 140
Parque Nacional Podocarpus Parque Nacional Yasuni
Nú
mero
d
e ocurr
encias
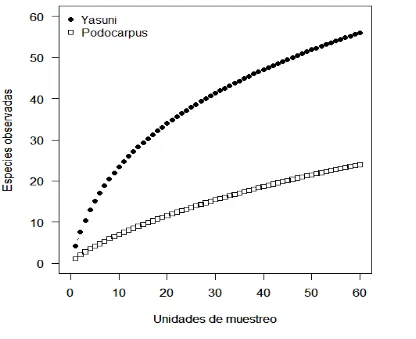
27 En la curva de rarefacción de especies establecida para el total del área muestreada, se puede deducir que la curva de las especies observadas en los dos tipos de Bosque casi logra alcanzar la asíntota. Con el esfuerzo de muestreo se obtuvo una parte representativa de especies, sin embargo en el Parque Nacional Yasuní y Parque Nacional Podocarpus se ve que aún la diversidad encontrada podría incrementarse, sobre todo en el Parque Nacional Podocarpus (Fig.6).
Figura 6. Curvas de rarefacción de especies para el Parque Nacional Yasuní (PNY) y Parque Nacional Podocarpus (PNP).
28 Tabla 5. Estimadores no paramétricos de riqueza en los dos tipos de bosque.
Parque Nacional Podocarpus Parque Nacional Yasuní
Estimadores Especies
Estimadas Error Estándar Especies Observadas Especies Estimadas Error Estándar Especies Observadas
ACE 1 50.8 4.19 24 87.4 3.98 56
CHAO 1 37 4.19 90.5 3.98
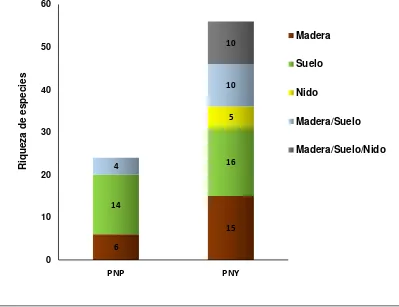
Los microhábitats que resultaron frecuentemente colonizados fueron el suelo, madera y nidos, siendo el suelo el microhábitat con mayor registro de especies observadas para ambas localidades.
En el Parque Nacional Yasuní 36 especies fueron muestreadas en un solo microhábitat, mientras que 20 especies compartieron hasta tres microhábitats, por otra parte en el Parque Nacional Podocarpus 20 especies se muestrearon en un solo microhábitat y 4 especies compartieron dos microhábitats (Fig.7).
Figura 7. Microhábitats ocupados por termitas en el Parque Nacional Podocarpus (PNP) y Parque Nacional Yasuní (PNY), en proporción al número total de encuentros.
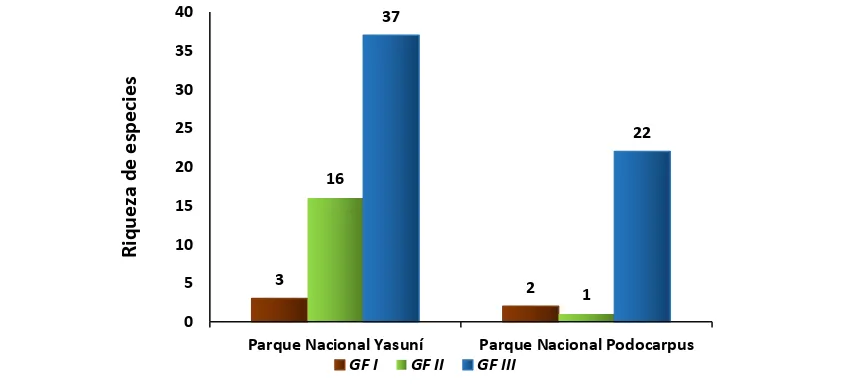
[image:37.595.98.497.431.738.2]29 En los dos sitios de estudio se identificó a tres grupos de alimentación. El GFII se alimenta de madera en descomposición / hojarasca, y es el gremio con el mayor número de ocurrencias para ambas localidades. El GFI que se alimenta exclusivamente de madera resultó ser el gremio con menor registro de ocurrencias para ambos sitios (Fig.8)
Figura 8. Grupos funcionales de termitas en el Parque Nacional Yasuní y Parque Nacional Podocarpus, en proporción al número de ocurrencias (Abundancia relativa).
Por otra parte los grupos de alimentación en relación a la riqueza de especies obtuvieron otro resultado. El GFIII que se alimenta de madera en descomposición/suelo humificado, es el grupo con el mayor número de especies para ambos sitios, mientras que el GFII y el GFI, obtuvieron un menor registro de especies en ambas localidades (Fig.9)
Figura 9. Grupos funcionales de termitas en el Parque Nacional Yasuní y Parque Nacional
Podocarpus, en proporción al número de ocurrencias (Abundancia relativa).
28 11 163 62 158 38 0 20 40 60 80 100 120 140 160 180
Parque Nacional Yasuní Parque Nacional Podocarpus
N ú mero d e o cu rr en ci as
GF I GF II GF III
3 2 16 1 37 22 0 5 10 15 20 25 30 35 40
Parque Nacional Yasuní Parque Nacional Podocarpus
Ri q u ez a d e espe ci es
[image:38.595.88.518.539.731.2]30
3.1.3 Variables ambientales.
Podemos determinar de acuerdo al coeficiente de correlación de Pearson que, la abundancia relativa y riqueza de especies de termitas registradas en el Parque Nacional Podocarpus, no se correlacionan con la mayoría de variables ambientales, a excepción de la variable “Peso Seco de Madera y Hojarasca” expresando que a medida de que existe un aumento en esta variable la abundancia de termitas incrementa. No obstante en el Parque Nacional Yasuní, las variables ambientales no se correlacionan significativamente con la riqueza de especies y abundancia relativa (Tabla 9).
Tabla 6. Coeficiente de Pearson (Correlación) entre variables microclimáticas y abundancia relativa en el Parque Nacional Yasuní y Parque Nacional Podocarpus.
Abundancia Relativa Riqueza
Variables Microambientales Correlación P- valor Correlación P- valor
Parque Nacional Podocarpus
Peso húmedo de hojarasca 0.8943 0.1874 1.136 0,103
%Hojas -0.997 0.8387 -0.95503 0.828
% Madera 0.989 0.1633 0.963 0.1697
Peso Seco de Hojarasca 35.567 <0.001 * 0.489 0.3133
% de Cobertura Vegetal 0.0169 0.507 0.83137 0.2046
Pendiente -0.0169 0.5067 11.809 0.1213
Parque Nacional Yasuní
Peso húmedo de hojarasca -0.6768 0.7494 -0.4147 0.6601
%Hojas 0.3823 0.3518 0.1311 0.4481
% Madera -0.2164 0.585 -0.00635 0.5025
Peso Seco de Hojarasca 10.417 0.1509 11.548 0.1265
% de Cobertura Vegetal 1.0417 0.1509 1.0417 0.1509
[image:39.595.73.517.315.755.2]31
3.2 Discusión.
El presente estudio abarcó un análisis del ensamblaje de la comunidad de termitas en dos bosques amazónicos de Ecuador, ubicados al noreste y al sureste, éste último es un sitio que aún no ha sido estudiado y no posee registros de termitas. Entre los seis puntos de muestreo se registró un total de 69 especies, este número manifiesta una alta diversidad de termitas para los dos bosques amazónicos, según Bahder et al., (2009), quien en su estudio realizado en ecosistemas similares encontró cifras de diversidad coincidentes (72 especies). A pesar de esto los estimadores de riqueza arrojaron valores elevados para cada sitio y disimiles a lo observado en los dos bosques de estudio Seguramente el conocimiento de riqueza de termitas aumentará si se incrementan más muestreos en los sitios de estudio y en otras localidades del país.
Dentro de este estudio las subfamilias con mayor número de especies se atribuyó a Apicotermitinae y Nasutitermitinae, similar a lo reportado por Davies et al., (2003) en un Bosque tropical inundable de Guyana Francesa y por Dahsljö et al., (2014b) en un Bosque tropical de tierras bajas en las selvas de Perú, en donde los mayores registros taxonómicos se atribuye a estos dos grupos. Eggleton (1999) y Bourguignon et al., (2011) señalan que la abundancia y riqueza de Apicotermitinae tiende a ser muy alta en los bosques tropicales, lo cual confirma nuestros resultados de composición taxonómica, 31 especies para Apicotermitinae y 19 para Nasutitermitinae. Nuestros resultados difieren a lo encontrado por Bahder et al., (2009), en los Bosques siempre verdes de Tierras Bajas de Ecuador (Orellana y Napo) donde se reporta a Nasutitermitinae como la subfamilia más abundante, seguido de Apicotermitinae, es posible que estas diferencias en la dominancia de las subfamilias se deba a que en el estudio desarrollado por Bahder y colaboradores se utilizaron protocolos no estandarizados, los encuentros fueron al azar en diferentes microhábitats exceptuando el suelo, y en un tiempo muy variado.
32 Un interesante resultado que resalta en este estudio es el hecho de que el 72,5% de especies reportadas para los dos parques (50 de 69 Podocarpus y Yasuní respectivamente) correspondan al Grupo alimenticio III, coincidiendo con lo encontrado por Palin et al., (2011) en un estudio realizado en un gradiente altitudinal de Perú. El Grupo alimenticio III está constituido por especies especialistas que se alimentan de madera en descomposición/suelo humificado, que generan un gran aporte en la fertilidad del suelo (Eggleton et al., 1995, 1996, 2002; Davies et al., 2003b; Jones et a.l, 2003). Por otro lado, las especies especialistas que corresponden al Grupo de alimentación I (5,7%) y Grupo de alimentación II (21,8%) registraron un menor número de especies. Este resultado surge como un efecto de la dinámica que presentan los bosques tropicales, en donde los árboles maduros proyectan continuamente ramas, que favorecen al aumento de madera, hojarasca y acumulación de humus en la capa superficial del suelo (Donovan et al., 2007). Basado en esta dinámica los gremios de alimentación GF I y GF II, producirían materia orgánica que contribuirían al aumento de nutrientes en el suelo, sirviendo como fuente de alimento para el GF III; grupo que resalta por su gran colonización en los dos bosques de estudio. No hay duda de que la participación de estas especies especialistas en los diversos procesos ecosistémicos del Parque Nacional Yasuní y Parque Nacional Podocarpus contribuye a la prestación de servicios esenciales como la descomposición de materia orgánica y el reciclaje de nutrientes (Jouquet et al., 2011).
34
CONCLUSIONES
Los dos Bosques Amazónicos estudiados albergan una alta diversidad de especies de termitas, pese a las diferencias estructurales de cada ecosistema.
El esfuerzo de muestreo para la estimación de la diversidad mediante transectos tuvo un efecto positivo en la determinación de la riqueza de termitas para los dos parques nacionales, sin embargo aún existen especies que no están registradas, por lo que se debe incrementar el muestreo a futuro.
Las subfamilias Apicotermitinae con el género Anoplotermes y Nasutitermitinae con el género Nasutitermes son el taxón con mayor dominancia en los dos bosques amazónicos de estudio.
La comunidad de termitas en el Parque Nacional Yasuní obtuvo un alto índice en abundancia relativa y riqueza de especies a diferencia de la comunidad de termitas en el Parque Nacional Podocarpus.
Las variables microambientales tomadas en el Parque Nacional Yasuní no influyen en el ensamblaje de la comunidad de termitas.
35
RECOMENDACIONES
Considerando la importancia de las termitas en los bosques tropicales y la falta de estudios en Ecuador, se recomienda continuar realizando investigaciones en los diversos biomas del país, para así comprender su comportamiento en los ecosistemas tropicales y obtener un listado taxonómico más amplio.
Para futuros estudios sería necesario agregar variables microambientales diferentes a las utilizadas en el presente estudio.
36
REFERENCIAS BIBLIOGRÁFICAS
Abadía L., J. C., Arcila, Á. M., & Chacón, P. (2013). Incidencia y distribución de termitas (Isoptera) en cultivos de cítricos de la costa caribe de Colombia. Revista Colombiana de Entomologia, 39(1), 1–8. Retrieved from
http://www.scopus.com/inward/record.url?eid=2-s2.0-84897887739&partnerID=tZOtx3y1
Abadía, C. J. & A. M. Arcila. 2009. Termitas en cultivos de limón en los departamentos del Atlántico y Magdalena, Colombia. Boletín del Museo de Entomología de la Universidad del Valle 10: 36–46
Ackerman, I. L., Constantino, R., Gauch, H. G., Lehmann, J., Riha, S. J., & Fernandes, E. C. M. (2009). Termite (Insecta: Isoptera) species composition in a primary rain forest and agroforests in central Amazonia. Biotropica, 41(2), 226–233. doi:10.1111/j.1744-7429.2008.00479.x
Adller, P & Foottit ,R.(2009) Introduction. In: Foottit, R.; P. Adler, eds. Insect Biodiversity. Science and Society. UK: Blackwell Publishing Ltd.; p. 1-6.
Araújo, V., Silva, M. P., & Vasconcellos, A. (2015). Soil-sampled Termites in Two Contrasting Ecosystems within the Semiarid Domain in Northeastern Brazil: Abundance, Biomass, and Seasonal Influences. Sociobiology, 62(1), 70–75. doi:10.13102/sociobiology.v62i1.70-75
Artigas, J. (1994). Entomología económica, insectos de inte- rés agrícola, forestal, médico y veterinario (nativos, intro- ducidos, y susceptibles de ser introducidos). Concepción, Chile. Ed. Universidad de Concepción, 1 v, 1126 p.
Bahder, Brian W.,Scheffrahn, Rudolf H., Křeček, Jan, Keil, Clifford & Whitney-King, Susan. (2009). Termites (Isóptera: Halotermitadae, Rhinotermitidae, Termitidae) of Ecuador”. Annales de la Société Entomologique de France 45: 529-36
37 Barros, E.; Pashanasi, B.; Constantino, R.; Lavelle, P. (2002). Effects of land-use system on the soil macrofauna in western Brazilian Amazonia. Biology and Fertilization of Soil 35: 338-347.
Bignell, D. E. (2000). Introduction to symbiosis, p. 169-187. In Abe, T., Higashi, M. & Bignell, D. E. (eds), Termites: Evolution, Sociality, Symbiosis, Ecology. Kluwer Academic Publications, Dordrecht.
Bignell, D. E., Roisin, Y., & Lo, N. (2011)a. Biology of Termites: A modern Synthesis.
Springer Science+Business Media B.V. 2011 Vol. 1.
doi:10.1017/CBO9781107415324.004
Bignell, D. E., Constantino, R., Csuzdi, C., Karyanto, A., Louzada, J., Susilo, F.-X., … Zanetti, R. (2011)b. Macrofauna. Manual de Biología de Suelos Tropicales, 91–148.
Bignell, D.E. & Eggleton, P., (2000). TERMITES IN ECOSYSTEMS. In Termites: Evolution, Sociality, Symbioses, Ecology. pp. 363–387.
Bignell, D.E. (2006). Termites as Soil Engineers and Soil Processors. In H. König & A. Varma (Eds.), Intestinal Microorganisms of Termites and Other Invertebrates (pp. 183–220). Berlin: Springer.
Blas, M., & Hoyo, J. Del. (2013). Entomología cultural y conservación de la biodiversidad . Los insectos en las Artes Mayores, 42, 1–22.
Bourguignon T, & Roisin Y. (2011). Revision of the termite family Rhinotermitidae (Isoptera) in New Guinea. Zookeys, 148, 55–103.
Bourguignon T, Lo N, Cameron SL, Šobotn'k J, Hayashi Y, Shigenobu S, Watanabe D, Roisin Y, Miura T, & Evans TA .(2015). The evolutionary history of termites as inferred from 66 mitochondrial genomes. Molecular Biology and Evolution, 32: 406–421
38 Bourguignon, T., Leponce, M. & Roisin, Y. (2011). Beta-diversity of termite assemblages among primary French Guiana rain forests. Biotropica 43:473–479.
Calderon, R. A.; Constantino, R. (2007). A survey of the termite fauna (Isoptera) of an eucalypt plantation in Central Brazil. Neotropical Entomology 36 (3): 391-395
Camousseight, A. (1999). Las termitas y su presencia en Chile. Santiago, Chile. CONAF (Corporación Nacional Forestal). Nota Técnica (37), 8 p.
Cancello, E. M., Silva, R. R., Vasconcellos, A., Reis, Y. T., & Oliveira, L. M. (2014). Latitudinal Variation in Termite Species Richness and Abundance along the Brazilian Atlantic Forest Hotspot. Biotropica, 46(4), 441–450. doi:10.1111/btp.12120
Coleman, D.C., D.A. Crossley, y P.F. Hendrix. (2004). Fundamentals of soil ecology, second edition. Elsevier Academic Press, Burlington, MA.
Collins, M. S. (1969). Water relations in termites. In K. W. Krishna & F. M. Weesner (Eds.), Biology of termites (Vol. 1, pp. 433–458). New York: Academic Press.
Colwell, R. ., Mao, C. ., & Chang, J. (2005). Interpolando, extrapolando y comparando las curvas de acumulación de especies basadas en su incidencia. Sobre Diversidad Biológica: El Significado de Las Diversidades Alfa, Beta Y Gamma, 73–84. doi:84−932807−7−1
Colwell, R. K., A. Chao, N. J. Gotelli, S.-Y. Lin, C. X. Mao, R. L. Chazdon, and J. T. Longino. (2012). Models and estimators linking individual-based and sample-based rarefaction, extrapolation, and comparison of assemblages. Journal of Plant
Ecology5:3-21
Colwell, R.K., & Coddington, J.A. (1994). Estimating terrestrial biodiversity through extrapolation. Philos. T. Roy. Soc. B, 345: 101–118. doi: 10.1098/rstb.1994.0091
Constantino, R. (2002). The pest termites of South America: Taxonomy, distribution and status. Journal of Applied Entomo- logy 126: 355-365.
Constantino ,R (2010) On-line termite database.
39 Constantino, R.; Acioli, A. N. S. (2006). Termite diversity in Brazil (Insecta: Isoptera). pp. 117-128. In: Moreira, F.; Siqueira, J. O.; Brussaard, L. (Eds.). Soil biodiversity in Amazonian and other Brazilian ecosystems. CAB International, Wallingford. 28 p.
Constantino, Reginaldo. (1998). “CATALAF OF THE LIVING TERMITES OF THE NEW WORLD (INSECTA: ISOPTERA)”. ARG.ZOO.S.PAULO: 35(2):135-230
Costa-Leonardo A. (1997). Métodos para coleta e estudo das populacões de cupins subterrâneos Naturalia Sao Paulo 22: 199-206.
Dahlsjö, C. a L., Parr, C. L., Malhi, Y., Rahman, H., Meir, P., Jones, D. T., & Eggleton, P. (2014)a. First comparison of quantitative estimates of termite biomass and abundance reveals strong intercontinental differences. Journal of Tropical Ecology, 30, 143–152. doi:10.1017/S0266467413000898
Dahlsjö, C. A. L, Parr, C. L., Malhi, Y. Meir,P. & Eggleton, P. (2014)b. Describing Termite Assemblage Structure in a Peruvian Lowland Tropical Rain Forest: A Comparison on Two Alternative Methods” Insects Sociaux: 141-50
Dangles, O., Nowicki, F. (2010). Biota Maxima. Pontificia Universidad Católica del Ecuador, Institut de Recherche pour le Développement. Imprenta Mariscal. Quito– Ecuador,
Darlington, J. (1985). Lenticular soil mounds in the Kenya Highlands. Oecologia 66:116-121.
Darlington, J. P. E. C., Zimmerman, P. R., Greenberg, J., Westberg, C., & Bakwin, P. (1997). Production of metabolic gases by nests of the termite Macrotermes jeanneli in Kenya. Journal of Tropical Ecology, 13(4), 491. doi:10.1017/S0266467400010671
40 Davies RG, Eggleton P, Jones D. (2003)a . Evolution of termite functional diversity: analysis and synthesis of local ecological and regional influences on local species richness. J Biogeogr 30:847–877
Davies, R. G., L. M. Hernandez, P.Eggleton, R.K. Didham, L.L.Fagan, And N. N. Winchester. (2003)b . Environmental and spatial influences upon species composition of a termite assemblage across neotropical forest islands. J. Trop. Ecol. 19: 509–524.
De Franca Alves, W. (2009). Térmitas Como Bioindicadores De Qualidade De Habitat Na Caatinga , Brasil : Há Uma Sintonia Entre As Variáveis Estruturais Dos Habitats E As. Universidade Federal Do Rio Grande Do Norte.
Dibog L, Eggleton P, Forzi F (1998) Seasonality of soil termites in a humid tropical forest, Mbalmayo, southern Cameroon. J Trop Ecol 14:841-850
Donovan S., Jones D., Sands W., Eggleton P. (2000) Morphological phylogenetics of termites (Isoptera) Biological Journal of the Linnean Society 70: 467- 513.
Donovan, S. E., Eggleton, P. y Bignell, D. E. (2001). Gut content analysis and a new feeding group classification of termites. Ecological entomology, 26:356-366.
Donovan, Sarah E., Paul Eggleton, and Andy Martin. (2002). “Species Composition of Termites of the Nyika Plateau Forests, Northern Malawi, over an Altitudinal Gradient.” African Journal of Ecology 40: 379–85.
Donovan, S. E., Griffiths, G. J. K., Homathevi, R., & Winder, L. (2007). The spatial pattern of soil-dwelling termites in primary and logged forest in Sabah, Malaysia. Ecological Entomology, 32, 1–10. doi:10.1111/j.1365-2311.2006.00856.x
Ebeling, W. (2000). Wood destroying insects and fungi.. Disponible en http://insects.ucr.edu/ entl33/ebeling/ebel5-l.html
Eggleton P, Williams PH, Gaston KJ (1994) Explaining termite global richness: productivity or history? Biodivers Conserv 3:318–330
41 Eggleton, P. (2000) Global patterns of termite diversity. Termites: evolution, sociality, symbioses, ecology (ed. by T. Abe, D.E. Bignell and M. Higashi), pp. 25–51. Kluwer Academic Publishers, Dordrecht, The Netherlands.
Eggleton, P. (2011). An introduction termites: Biology, taxonomy and funtional morphology. pp. 1-26. In: Bignell, D. E.; Roisin, Y.; Lo, N. (Eds.). Biology of termites: A modern synthesis. Springer Science + Business Media B.V. 592 p.
Eggleton, P., Bignell, D. E., Sands, W. A., Mawdsley, N. A., Lawton, J. H., Wood, T. G., & Bignell, N. C. (1996). The diversity, abundance and biomass of termites under differing levels of disturbance in the Mbalmayo Forest Reserve, southern Cameroon. Society, The Royal Transactions, Philosophical Society, Royal Sciences, Biological, 351, 51–68.
Eggleton, P., Bignell, D.E., Sands, W.A., Waite, B., Wood, T.G. & Lawton, J.H. (1995) The species richness of termites (Isoptera) under differing levels of forest disturbance in the Mbalmayo Forest Reserve, southern Cameroon. Journal of Tropical Ecology, 11, 85-98.
Eggleton, P., Homathevi, R., Jeeva, D., Jones, D.T., Davies, R.G. & Gut content analysis in termites 365 Maryati, M. (1997) The species richness and composition of termites (Isoptera) in primary and regenerating lowland dipterocarp forest in Sabah, east Malaysia. Ecotropica, 3, 119-128.
Eggleton, P., Homathevi, R., Jones, D. T., Macdonald, J. A., Jeeva, D., Bignell, D. E., … Maryati, M. (1999). Termite Assemblages, Forest Disturbance and Greenhouse Gas Fluxes in Sabah, East Malaysia. Philosophical Transactions: Biological Sciences, 354(1391), 1791–1802. doi:10.1017/CBO9781107415324.004
Eggleton, P., Homathevi, R., Jones, D. T., Macdonald, J. A., Jeeva, D., Bignell, D. E., Maryati, M. (1999). Termite Assemblages, Forest Disturbance and Greenhouse Gas Fluxes in Sabah, East Malaysia. Philosophical Transactions: Biological Sciences, 354(1391), 1791–1802. doi:10.1017/CBO9781107415324.004 London.
42 Fontes L. R. (1996). Controle de cupins em ambientes urbanos. Simposio
Latino-Americano
Gathorne-Hardy, F., Syaukani, And P. Eggleton. (2001). The effects of altitude and rainfall on the composition of the termites (Isoptera) of the Leuser Ecosystem (Sumatra, Indonesia). J. Trop. Ecol. 17: 379–393
Geiger, R., Aron, R. H., & Todhunter, P.(2003). The climate near the ground (6a ed.). Maryland, USA: Rowman y Littlefield Publishers, Inc
Girardin, a J. & Rozasda, A., (2011). Termite Diversity along an Amazon – Andes Elevation Gradient , Peru. Natural History, 43(1900 m), pp.100–107
Gillot, C. 2005. Entomology. 3 ed. Springer, Dordrecht, The Netherland.
Jones, C.G., Lawton, J.H. & Shachak, M., (1994). Organisms as Ecosystem Engineers Organisms as ecosystem engineers. , 69(3), pp.373–386.
Jones, C.G., Lawton, J.H. & Shachak, M., (1997). Jones et al 1997 Positive Ecology 78 1946-1957.pdf. Ecological Society America, 78, pp.1946–1957.
Jones, D, & Eggleton, P. (2011) Global Biogeography of Termites: A Compilation of Sources. Biology of Termites: A modern Synthesis pp, 477-498
Jones, D. T. (2000). Termite assemblages in two distinct montane forest types at 1000 m elevation in the Maliau Basin, Sabah. Journal of Tropical Ecology, 16(2), 271– 286. doi:10.1017/S0266467400001401
Jones, D.T. & Eggleton, P., (2000). Sampling termite assemblages in tropical forests: Testing a rapid biodiversity assessment protocol. Journal of Applied Ecology, 37(1), pp.191–203.
Jouquet, P., Traoré, S., Choosai, C., Hartmann, C., & Bignell, D. (2011). Influence of termites on ecosystem functioning. Ecosystem services provided by termites. European Journal of Soil Biology, 47(November), 215–222. doi:10.1016/j.ejsobi.2011.05.005
43 Kimmins, J. P. (1987). Forest ecology. A foundation for sustainable management (2a
ed.). New Jersey, USA: Prentice Hall
Korb J (2008). Termites, hemimetabolous diploid white ants? Front Zool 5:15
Krebs, C. J. (1998) Ecological Methodology, 2nd edition, Pearson Education, London.
Krishna K., Weesner F. (1969) Biology of termites volume 1 Academic Press New York 598 pp
Krishna, K., Grimaldi, D.A., Krishna, V. & Engel, M.S. (2013) Treatise on the Isoptera of the world. Bulletin of theAmerican Museum of Natural History, 377(Volumes 1–7), 1–2704.http://www.bioone.org/doi/abs/10.1206/377.1
Lavelle, P. and A.V. Spain. 2001. Soil Ecology, Kluwer Scientific, Amsterdam
Lavelle, P., Bignell, D., Lepage, M., Wolters, W., Roger, P., Ineson, P. & Dhillion, S. (1997). Soil function in a changing world : the role of invertebrate ecosystem engineers. European Journal of Soil Biology, 33(May 1996), 159–193.
Lee E, Wood TG. (1971). Physical and chemical effects on soils of some Australian termites and their pedological significance. Pedo- biologia 11:376-409
Lepage, M. (1983) Foraging of Macrotermes spp. (Isoptera: Macrotermitinae) in the tropics. Social Insects in the Tropics 1, 205-217.
M.R. Spiegel; J. Schiller; R. A. Srinivasan (2007). «9. Análisis de la varianza». Probabilidad y Estadística [Schaum's Outline of Theory and Problems of Probability and Statistics]. Schaum (2ª edición). México D.F.: McGraw-Hill. pp. 335-371.
Magurran, A.E. (2001).Ecological diversity and its measurement. Princeton University Press,New Jersey, 179 pp
Magurran, A.E. (2004). Explaining the excess of rare species in natural species abundance distributions Nature 422, 714-716