TITULACIÓN DE INGENIERA EN GESTIÓN AMBIENTAL
Patrones de diversidad de la fauna de moluscos
(Gastrópoda) en un bosque tropical de montaña (Estación
Científica San Francisco) al sur de Ecuador.
Trabajo final de titulación.
AUTORA
Imaicela Ordóñez, María Augusta
DIRECTOR.
Saa, Luis Rodrigo, Ph.D.
Loja - Ecuador
II
CERTIFICACIÓN
Doctor
Luis Rodrigo Saa
DIRECTOR DEL TRABAJO DE FIN DE TITULACIÓN
CERTIFICA.
Que el presente trabajo de investigación denominado “Patrones de diversidad de la
fauna de moluscos (Gastrópoda) en un bosque tropical de montaña (Estación Científica San Francisco) al sur de Ecuador” realizado por la profesional en formación: María Augusta Imaicela Ordóñez; cumple con todos los requisitos establecidos por las normas generales para la graduación en la Universidad Técnica Particular de Loja, tanto en el aspecto de forma como de contenido, por lo cual me permito autorizar su presentación para los fines pertinentes.
Loja, marzo de 2013
III
CESIÓN DE DERECHOS
“Yo, María Augusta Imaicela Ordóñez declaro ser autora del presente trabajo y eximo expresamente a la Universidad Técnica Particular de Loja, y a sus representantes legales de posibles reclamos o acciones legales.
Adicionalmente declaro conocer y aceptar la disposición del Art. 67 del Estatuto Orgánico de la Universidad Técnica Particular de Loja que en su parte pertinente textualmente dice: “Forman parte del patrimonio de la Universidad la propiedad intelectual de investigaciones, trabajos científicos o técnicos de tesis de grado que se realicen a través, o con el apoyo financiero, académico o institucional (operativo) de la Universidad”.
Imaicela Ordoñez María Augusta
IV
DECLARACIÓN DE AUTORÍA
Las ideas, conceptos, procedimientos y resultados vertidos en el presente trabajo de fin de carrera, son de exclusiva responsabilidad de su autor.
V
DEDICATORIA
De una manera muy especial le dedico el presente trabajo a Dios, por darme la fuerza día a día para desarrollarlo.
A mi familia, a mis padres y hermanos, que son los pilares fundamentales en mi vida, sin su paciencia, confianza y sobre todo apoyo no hubiera podido realizar este trabajo.
A todos con cariño y afecto.
VI
AGRADECIMIENTO
A la Universidad Técnica Particular de Loja que por medio del Laboratorio de Sanidad Animal y Zoonosis (LABSA) del Departamento de Ciencias Agropecuarias y de Alimentos y a la Titulación de Gestión Ambiental que me ha dado la oportunidad de formarme y capacitarme para la elaboración del presente trabajo de investigación.
A mis padres y hermanos por su apoyo incondicional y sobre todo por la confianza depositada en mí, durante todo el proceso de mi carrera universitaria.
A mi director de tesis el Dr. Luis Rodrigo Saa, Ph.D. por el apoyo brindado a lo largo de la consecución del presente trabajo.
Al Ing. Carlos Iñiguez e Ing. Diana Maldonado que me ayudaron y guiaron a la elaboración de este proyecto.
Al Dr. Modesto Correoso, importante malacólogo en Ecuador, por su valiosa contribución y ayuda en la elaboración del presente trabajo.
VII
ÍNDICE DE CONTENIDOS
CERTIFICACIÓN DEL DIRECTOR DE TESIS ... II CESIÓN DE DERECHOS ... III DECLARACIÓN DE AUTORIA ... IV DEDICATORIA ... V AGRADECIMIENTO ... VI ÍNDICE DE CONTENIDOS ... VII ÍNDICE DE TABLAS ... IX ÍNDICE DE FIGURAS ... X ÍNDICE DE GRÁFICOS ... XI RESUMEN EJECUTIVO ... XII
1. INTRODUCCIÓN ... 1
2. OBJETIVOS ... 3
2.1. GENERAL ... 3
2.2. ESPECÍFICOS ... 3
3. MARCO TEÓRICO...4
3.1. PHYLUM MOLLUSCA ... 4
3.2. CLASE GASTRÓPODA ... 6
3.2.1. ORIGEN EVOLUTIVO DE LOS GASTRÓPODOS ... 8
3.2.2. ALIMENTACIÓN DE LOS GASTROPÓDOS ... 9
3.2.3. EXCRECIÓN ... 10
3.2.4. SISTEMA CIRCULATORIO ... 10
3.2.5. SISEMA NERVIOSO ... 10
3.2.6. REPRODUCCIÓN ... 11
VIII
3.4. GASTRÓPODOS EN ECUADOR ... 14
3.5. IMPORTANCIA DE LOS GASTRÓPODOS ... 16
3.6. PROTECCIÓN AMBIENTAL DE LOS MOLUSCOS EN ECUADOR ... 17
3.7. GASTRÓPODOS COMO PLAGAS ... 17
3.8. CARACOLES Y ENFERMEDADES QUE TRANSMITEN A LAS PERSONAS ... 19
3.9. LOS MOLUSCOS Y EL CAMBIO CLIMÁTICO ... 19
3.10. DECLIVE DE GASTRÓPODOS TERRESTRES ... 21
4. ÁREA DE ESTUDIO ... 22
5. METODOLOGÍA ... 24
5.1. MUESTREO DE GASTRÓPODOS TERRESTRES ... 24
5.2. ANÁLISIS DE DATOS ... 25
5.2.1. ÍNDICES DE RIQUEZA Y DIVERSIDAD ... 25
5.2.2. COMPOSICIÓN DE GASTRÓPODOS ... 26
6. RESULTADOS ... 27
6.1. RIQUEZA Y DIVERSIDAD ... 27
6.2. COMPOSICIÓN DE ESPECIES ... 30
7. DISCUSIÓN ... 32
8. CONCLUSIONES ... 34
9. RECOMENDACIONES ... 35
10. BIBLIOGRAFÍA ... 36
11. GLOSARIO ... 44
IX
ÍNDICE DE TABLAS
Tabla 1. Clasificación de la clase Gastrópoda en Ecuador ... 16
Tabla 2. Visión general de los gastrópodos encontrados en el BI y BNI ... 27
X
ÍNDICE DE FIGURAS
Figura 1. Phylum Mollusca, Systrophia heligmoida (Orbigny, 1835) ... 4
Figura 2. Terminología descriptiva de la concha de los gastrópodos ... 6
Figura 3. a) Fósil de gastrópodo, b) Fósil de ammonite ... 7
Figura 4. Partes principales de un gastrópodo... 8
Figura 5. Gastrópodo alimentándose ... 9
Figura 6. Anatomía de un gastrópodo ... 11
Figura 7. Cortejo, penetración, apareamiento, puesta, huevos, crías de gastrópodos terrestres ... 12
Figura 8. Gastrópodo terrestre ... 14
Figura 9. Achatina fulica ... 18
Figura 10. Mapa del área de estudio... 22
Figura 11. Tipos de bosque ... 23
XI
ÍNDICE DE GRÁFICOS
XII
RESUMEN EJECUTIVO
La biodiversidad es la variedad total de vida que existe sobre la tierra; como parte de esta se encuentran los moluscos terrestres (Gastrópoda), ha sido considerado como un fuerte indicador del estado de los ecosistemas naturales.
En el bosque tropical de montaña ubicado en la Estación Científica San Francisco en Zamora Chinchipe (Ecuador) durante julio - septiembre de 2012, se determinó los patrones de diversidad y composición de especies en dos zonas correspondientes a bosque intervenido y no intervenido.
Se colectó 1 898 gastrópodos correspondientes a 9 especies, Systrophia heligmoida
fue la más abundante con 1 767 especímenes. El esfuerzo de muestreo fue óptimo, la diversidad fue mayor para el bosque no intervenido.
A través del Análisis de Escalamiento Multidimensional no Métrico (NMDS) y del Análisis de similitudes (ANOSIM) se pudo distinguir en los tipos de bosque estudiados, composición de especies diferentes, el bosque intervenido desorganizada y dispersa, mientras que el bosque no intervenido homogénea.
1
1. INTRODUCCIÓN
En el transcurso del tiempo el clima global ha venido sufriendo un sinnúmero de cambios, asociados principalmente al incremento de los gases que causan el efecto invernadero, se calcula que la temperatura mundial de la superficie ha aumentado en 0,6 °C (IPCC, 2001). La atmósfera responde a este incremento de gases con un aumento de temperatura (Aalst, 2006). Estos cambios tienen efectos sobre especies vegetales y animales incluidos los moluscos, en diferentes vías como su distribución, tamaño poblacional, estructura física, metabolismo y comportamiento (IPCC 2001; IPCC 2002; Parmesan et al., 2003; Parmesan et al., 2004; Cumming et al., 2006), alterando los procesos naturales de los ecosistemas (Aalst, 2006).
La variedad total de vida existente en la tierra se denomina biodiversidad, la cual ofrece grandes beneficios directos como alimento, medicina, madera, energía primaria, materia prima, lugares de esparcimiento, entretenimiento, modelos de ingeniería y medicina, sustancias de uso agrícola, entre otros. Como parte de esta biodiversidad se encuentran los moluscos, los cuales hacen referencia al nombre común de los miembros de un filo de animales de cuerpo blando que suelen tener una envoltura externa dura de naturaleza calcárea, “Mollusca” (Camarda & Suarez, 2004).
El filo Mollusca está formado por unas 50 000 especies vivientes y unos 35 000 fósiles, su reproducción es sexual pero pueden ser dioicas o monoicas; siempre vinculadas a medios acuáticos o húmedos. La mayoría son marinos, algunos dulciacuícolas y unos pocos viven en ambientes terrestres pero siempre limitados por su necesidad de agua y humedad (Aycaguer, 2006).
2
Los moluscos terrestres tipifican desde el punto de vista biogeográfico a un país por sus especies endémicas, pues el genuino endemismo ocurre en especies con poca locomoción. Muchos moluscos pueden ser endémicos de algunas regiones y localidades del país, en general en Ecuador existen muchas especies de moluscos difíciles de encontrar y recolectar en su territorio, lo que contribuye a su baja presencia en colecciones de museos locales (Correoso, 2008).
El papel de los moluscos, en la dinámica ecológica de los ecosistemas donde habitan, es amplio, contribuyen a la formación del suelo tropical, cumplen funciones tróficas básicas en los ecosistemas y constituyen una parte importante de la biomasa de los invertebrados tropicales (Martínez & Piñango, 2008).
3
2. OBJETIVOS
2.1. GENERAL:
Conocer la diversidad de moluscos terrestres en un bosque tropical de montaña al sur de Ecuador.
2.2. ESPECÍFICOS:
Determinar la variación en los patrones de diversidad de moluscos (gastrópodos), en relación a dos tipos de bosques en un bosque tropical de montaña al sur de Ecuador.
4
3. MARCO TEÓRICO
3.1. PHYLUM MOLLUSCA
[image:16.595.240.385.212.353.2]El nombre del Phylum Mollusca deriva del latin “mollis” que significa suave o blando; se refiere al hecho de presentar un cuerpo suave el cual está protegido por una concha dura de naturaleza calcárea (Mille & Pérez, 2003).
Figura 1. Phylum Mollusca, Systrophia heligmoida (Orbigny, 1835)
Elaboración: Autora
Los primeros moluscos fósiles aparecieron en rocas del periodo cámbrico hace unos 600 millones de años; este es uno de los taxa más abundantes entre los invertebrados, tanto por el número de especies como por el número de individuos. Se encuentran en diversos hábitats terrestres y acuáticos y conforman aproximadamente 110 000 especies vivientes. Todas las clases vivientes son predominantemente marinas, solamente los caracoles y almejas han llegado a los hábitats terrestres. Especies comensales solo se observan entre los caracoles y las almejas y tan solo una familia de caracoles tiene representantes parásitos, estos últimos solo se reconocen por la larva, dadas las modificaciones que han sufrido los adultos como adaptación al parasitismo. La mayoría de especies vivientes de moluscos pertenecen a las clases Gastropoda, Pelecypoda y Cephalopoda (Russel, 1968).
5
concha se denomina manto o palio. El epitelio del manto forma, desde la base del saco visceral, un doblez de la piel, el pliegue del manto (a menudo denominado simplemente manto). En el interior de los espacios cubiertos por el manto, el surco del manto o la cavidad del manto, se localizan los órganos respiratorios, el osfradio (órganos sensoriales) y la glándula hipobranquial. En la cavidad del manto se abre el ano y los conductos del órgano excretor y reproductor (Storch & Welsch, 2001).
La piel presenta, siempre y cuando no esté cubierta por la concha, a menudo cilios, a veces espinas o escamas, carece casi siempre de cutícula y es rica en células glandulares grandes, y por eso es resbaladiza y blanda. La concha está formada al menos por tres capas: por el periostraco orgánico y externo y por dos capas calcáreas. Entre el manto y la concha se encuentran, además de las inserciones musculares, una estrecha cavidad extrapaleal, rellena de líquido. La reducción de la concha y su desplazamiento al interior del cuerpo no son fenómenos raros (Storch & Welsch, 2001).
El celoma de los moluscos se limita generalmente a una cavidad que comprende las gónadas y el corazón a veces también a un trozo de intestino. La mayoría de las veces solamente pertenece la parte que rodea el corazón como cavidad pericárdica. (Storch & Welsch, 2001).
Los moluscos son ecológicamente muy importantes en el ambiente marino y terrestre, y generalmente ocupan posiciones intermedias en las cadenas alimentarias (herbívoros, o carnívoros primarios) (Aycaguer, 2006).
En cuanto a la importancia económica de los moluscos, es cardinal en un país que tiene en sus hábitos alimentarios el consumo de importantes cantidades y variedades de moluscos marinos y terrestres, de alto valor proteico y digestibilidad reconocidos (Sánchez et al., 1991); destacándose algunas familias terrestres de caracoles que son parte importante de la dieta de grupos indígenas de la amazonía y de la sierra ecuatoriana (Patzelt, 2000).
6
Solenogastres, Scaphopoda y Cephalopoda con alrededor de 2 000 especies. Desde el punto de vista económico y alimentario, los taxa más importantes de moluscos son: Bivalvia, Cephalopoda y Gastrópoda. En particular, algunos representantes de la clase Gastrópoda son de especial interés médico; destacan entre ellos ciertos miembros de la familia Planorbidae y Melanidae, involucrados como intermediarios en ciclos de transmisión de enfermedades parasitarias. En la acuariofilia también se encuentran representantes de los moluscos, principalmente gastrópodos y bivalvos (Martínez & Piñango, 2008).
3.2. CLASE GASTRÓPODA
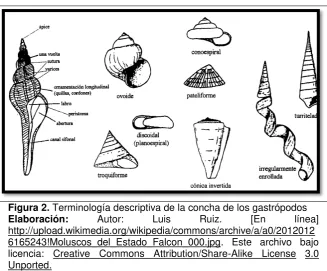
[image:18.595.148.476.437.708.2]La clase gastrópoda constituye un grupo muy diverso con la concha de una sola pieza, la cual puede ser enrollada, formando un caracol elíptico, helicoidal o turbinado, puede tener la forma de una espiral aplanada o de un cono corto, o incluso perderse secundariamente. Esta clase abarca aproximadamente 50 000 especies vivientes, la mayoría son marinos. Sin embargo, el grupo de moluscos más ampliamente desarrollados. La concha de algunas especies es muy apreciada por los coleccionistas del mundo, lo que ha propiciado que se vean en peligro de extinción (Mille & Pérez, 2003).
Figura 2. Terminología descriptiva de la concha de los gastrópodos
7
[image:19.595.85.578.234.428.2]La clase presenta un registro fósil ininterrumpido y ha tenido la radiación adaptativa más notable de todos los moluscos. Si tenemos en cuenta la enorme diversidad de hábitat que los gastrópodos han invadido, hemos de considerarlos como la clase con mayor éxito evolutivo de todos los moluscos. Las especies marinas se han adaptado a vivir sobre cualquier tipo de sustrato, así como también a la vida pelágica. Han invadido las aguas dulces, los pulmonados y otros grupos han conquistado el medio terrestre tras perder las branquias y convertir la cavidad peleal en un pulmón (Barnes, 1996).
Figura 3. a) Fósil de gastrópodo, b) Fósil ammonite
Elaboración: Autora
8
[image:20.595.123.503.133.401.2]Este mecanismo contribuye a la solución de los problemas de saneamiento implicados por la torsión, ya que el ano quedaba por encima de la cabeza y de la abertura con la corriente inhalante (Scarabino, 2004).
Figura 4. Partes principales de un gastrópodo.
Elaboración: Autor: Jürgen Schoner. [En línea] http://commons.wikimedia.org/wiki/File:Grapevinesnail_01.jpg. Este archivo bajo licencia: Creative Commons Attribution/Share-Alike License 3.0 Unported.
3.2.1. ORIGEN EVOLUTIVO DE LOS GASTRÓPODOS
Antiguamente todos los gastrópodos eran acuáticos, pero posteriormente, en el curso de la evolución una parte de ellos se han adoptado a vivir en tierra, son los denominados gastrópodos terrestres. El cambio a la vida terrestre tenía como problema principal el de la deshidratación. Los caracoles terrestres adoptan diversas soluciones para resolver este problema (Barnes, 1996).
9
3.2.2. ALIMENTACIÓN DE LOS GASTRÓPODOS
[image:21.595.171.412.242.445.2]Los gastrópodos terrestres siguen una dieta muy variada, alimentándose de plantas en descomposición, hongos, líquenes, algas y de plantas verdes, algunas especies son carnívoras, suelen ser dañinos para las plantas hortícolas ya que estas suelen ser más tiernas que las plantas silvestres, a este filo pertenecen las ostras, calamares, pulpos y caracoles, son animales acuáticos que se alimentan al mismo tiempo que se desplazan sobre un sustrato sólido por medio de un pie. (Forcelli, 2000).
Figura 5. Gastrópodo alimentándose
Elaboración: Autor: Tarquin. [En línea] http://commons.wikimedia.org/wiki/File:Iberus_gualterian us_alonensis2.JPG. Este archivo bajo licencia: Creative Commons Attribution/Share-Alike License 3.0 Unported
Aparato venenoso: Entre los moluscos, los carnívoros han desarrollado diferentes estrategias para atacar a sus presas. En el caso de los conos (superfamilia Conacea), cuentan con una probóscide muy larga que tiene en la punta un diente por medio del cual inyectan veneno neurotóxico. En el hombre pueden llegar a causar la muerte pocas horas después de ocurrido el accidente. (López & Urcuyo, 2009).
10
observa una migración de la apertura del esófago a la zona anterior y el ano a la posterior. Existen especies de gastrópodos herbívoros, carnívoros, detritívoros y carroñeros. Hay gastrópodos parásitos, ecto o endoparásitos, que presentan el cuerpo modificado para alimentarse de la especie parasitada. Los más modificados son de tipo vermiformes y viven en el interior de holoturoideos. (Forcelli, 2000).
3.2.3. EXCRECIÓN
En gastrópodos primitivos la excreción se realiza a través de los nefridios. Estos en uno de sus extremos desemboca en la cavidad pericárdica de la que recibe desperdicios que luego reduce a orina; el otro extremo se conecta a la cavidad paleal a donde envía la orina para que sea expulsada. En épocas secas o frías cierran su concha produciendo un moco que forma una capa dura al secarse para protegerse del ambiente y evitar la pérdida de agua en condiciones desfavorables; aun así, este tipo de especies, como las babosas comunes, pueden sobrevivir tras la pérdida de hasta un 80% de agua (López & Urcuyo, 2009).
3.2.4. SISTEMA CIRCULATORIO
Como resultado de la torsión el corazón se encuentra delante de la masa visceral. Está formado por dos aurículas en las formas primitivas, pero como consecuencia del enrollamiento de la concha y la desaparición de la branquia derecha, la aurícula derecha desaparece, ya que era la encargada de irrigar la estructura respiratoria. Desde el ventrículo salen dos aortas, una anterior y una posterior o una única que rápidamente se ramifica en dos. La rama anterior irriga la cabeza y el pie; debido al papel hidrostático que juega la sangre en el desplazamiento y movilidad del animal el sistema arterial desaparece para formar unos senos en los que se encuentra la sangre; esto no ocurre en la zona de la masa visceral, donde la sangre únicamente tiene función de transporte de sustancias. Antes de pasar por las branquias de retorno al corazón atraviesa el nefridio para eliminar las impurezas (Forcelli, 2000).
3.2.5. SISTEMA NERVIOSO
11
[image:23.595.166.444.217.410.2]Normalmente los ganglios que rodean el aparato digestivo han desarrollado un cerebro con varios órganos sensoriales. De hecho el sistema nervioso de los cefalópodos es tan complejo y está tan altamente organizado como el de los peces. El corazón se localiza en el extremo posterior del cuerpo y envía la sangre hacia un sistema circulatorio abierto que forma parte de la cavidad principal del cuerpo. Asociado al corazón existe un conjunto de órganos en el que se encuentran los riñones, las gónadas y a veces otras estructuras reproductoras. (Camarda & Suarez, 2004).
Figura 6. Anatomía de un gastrópodo
Elaboración: Autor: Vázquez. [En línea] http://commons.wikimedia.org/wiki/File:Scheme_snail_anatom y-numbers.svg. Este archivo bajo licencia: Creative Commons Attribution/Share-Alike License 3.0 Unported.
3.2.6. REPRODUCCIÓN
Los gastrópodos terrestres son monoicos o de sexos separados o bien son hermafroditas pero no son capaces de llevar a cabo la autofecundación, por lo que necesitan aparearse dos individuos maduros sexualmente para que se produzca la fecundación. En general, los gastrópodos terrestres hermafroditas no presentan ambos sexos al mismo tiempo, sino que son protándricos, es decir, que de jóvenes son machos, pero que al cabo de un año o dos se transforman en hembras. Cuando dos caracoles se encuentran entre ellos en la época de la reproducción, se produce un cortejo que consiste en rozamientos con los tentáculos, con el pie y con la cabeza, con los cuales el macho va estimulando a la hembra y comprueba su estado reproductivo. Si la hembra es receptiva y está activa sexualmente, entonces comienza el proceso de apareamiento (Barnes, 1996).
12
con otro, produciéndose el intercambio de fluido seminal desde el macho a la hembra con lo cual se produce la fecundación interna de los huevos. Tras el apareamiento, los dos caracoles se separan y la hembra busca un lugar adecuado para depositar los huevos. En algunos casos, la hembra excava un hoyo en la tierra húmeda donde depositará los huevos, de uno en uno por lo que tarda mucho tiempo en este proceso de puesta de los huevos. En otras ocasiones, la hembra deposita sus huevos en agujeros del terreno, hendiduras en la madera podrida o bajo las piedras. La mayoría de las especies ponen de 20 a 25 huevos, si bien algunas pueden poner hasta 100 huevos o más. Los huevos tienen normalmente forma esférica, son blandos, transparentes y en algunos casos están protegidos por una cubierta dura y opaca. El periodo de desarrollo embrionario depende exclusivamente de la temperatura, siendo por lo general seis semanas. Los nuevos caracoles tienen el aspecto de adultos en miniatura y alcanzan la madurez sexual en un año aproximadamente, aunque las especies de grandes dimensiones tardan de 2 a 4 años (Camarda & Suarez, 2004).
[image:24.595.91.524.492.683.2]El índice de mortalidad durante el primer periodo de vida es muy alto. Los huevos no están al cuidado de los progenitores, pueden secarse, ser devorados por otros animales o ser parasitados. Los pequeños, además, son muy vulnerables, sufren más las condiciones climáticas y son fáciles de cazar. Generalmente, sólo el 5% de los huevos depositados alcanza la edad adulta (Fernández, 2007).
Figura 7.a) Cortejo, b) Penetración, c) Apareamiento, d) Puesta, e) Huevos y f) Crías de gastrópodos terrestres.
13
En algunos casos, los adultos mueren después de la puesta de los huevos, pero en otros casos, pueden vivir varios años (hasta 8 o 10). Sin embargo, a lo largo de su vida, los caracoles terrestres corren numerosos peligros, ya que son muchos sus depredadores. El más conocido enemigo de los caracoles terrestres es el zorzal
(Turdus philomelos). Otros enemigos son la musaraña (Crocidura russula), el puerco
espín (Coendou prehensilis), muchos roedores y las larvas de la luciérnaga. Además, existen muchos parásitos que pueden atacar tanto a los huevos como a los individuos adultos. Incluso, los caracoles terrestres son portadores de muchos parásitos peligrosos para el hombre (Fernández, 2007).
3.3. GASTRÓPODOS TERRESTRES
Los gastrópodos se hallan representados entre 40 000 a 150 000 especies, mientras que la riqueza global de grupos terrestres se estima entre 30 000 y 35 000 especies de ese total (Aktipis et al. 2008). A pesar de la importancia en términos de riqueza y diversidad que tienen los gastrópodos terrestres, estudios de este tipo son escasos.
Antiguamente, todos los gastrópodos eran acuáticos, pero posteriormente, en el curso de la evolución, un parte de ellos se ha adaptado a vivir en tierra, son los denominados gastrópodos terrestres. El cambio a la vida terrestre tenía como problema principal el de la deshidratación. Los caracoles terrestres adoptan diversas soluciones para resolver este problema. En primer lugar, la mayoría de ellos vive en ambientes húmedos y lleva una vida activa principalmente nocturna, resolviendo de esta forma, al mismo tiempo, el problema añadido que suponen los posibles depredadores, ya que, en la oscuridad, las conchas resultan mucho menos visibles y por lo tanto, están más protegidas (Naranjo, 2003).
14
[image:26.595.203.424.356.574.2]Un factor que condiciona en gran medida a los caracoles terrestres es el tipo de terreno. La mayoría de ellos prefiere terrenos calcáreos, ya que el calcio resulta muy importante para estos animales, que lo utilizan en la formación de sus conchas. Por lo tanto, en los terrenos calcáreos siempre existe un mayor número de especies que en los ácidos. Las especies de caracoles que viven en terrenos ácidos se caracterizan por presentar conchas delgadas y frágiles. Un segundo factor importante que condiciona la presencia de los gastrópodos terrestres es el clima, en general, la humedad ambiental resulta importante para los caracoles terrestres, así como el calor, comparativamente, la diversidad de especies de caracoles terrestres aumenta progresivamente desde los polos hasta las zonas ecuatoriales; sin embargo, hay grandes zonas abiertas, secas y desprovistas de suficiente vegetación y humedad en las que no suelen vivir los caracoles terrestres, estos suelen ser más abundantes en zonas sombrías y húmedas, con abundante vegetación, cobijándose bajo piedras, madera o musgo (Naranjo, 2003).
Figura 8. Gastrópodo terrestre.
Elaboración: Autor: Stako. [En línea] http://commons.wikimedia.org/wiki/File:Slimaczek.jp g. Este archivo bajo licencia: Creative Commons Attribution/Share-Alike License 3.0 Unported.
3.4. GASTRÓPODOS EN ECUADOR
La biodiversidad de moluscos terrestres de Ecuador continental es poco conocida pero existe una riqueza relativamente alta de moluscos, con especies típicas y/o endémicas muy interesantes. Esta fauna tiene especies representativas, endémicos y moluscos de tamaños extremos: gigantescos como el caracol de Pope
(Strophocheilu spopelairianus), algunos prosobranquios muy interesantes así como
15
La fauna de moluscos del país abarca desde especies muy pequeñas casi insignificantes y de colores muy apagados hasta especies muy bonitas y llamativas
como en los géneros Corona, Sultana y Solaropsidae, de los bosques amazónicos.
Es notoria la familia Bulimulidae, la mejor representada en el país con más del 50% de especies; solo en Galápagos hay 65 especies endémicas. No es raro encontrar moluscos del género Bulimulus que viven en los fríos páramos a más de 4 000 m.s.n.m., entre las especies de moluscos, que viven a mayor altitud, además de especies de zonas áridas como Machalilla en la costa y otras de bosques nublados y del oriente (Correoso, 2008).
Los moluscos terrestres en general tienen en los trópicos, condiciones ideales de humedad vegetación y temperatura; del suelo que aporta el calcio para formar la concha: el carbonato de calcio es un gran factor limitante. Esto es tan importante que los caracoles tienen limitado su grado de dispersión, debido a la presencia de sales cálcicas del suelo, por eso hábitats como ríos y lagos pobres en esta sal pueden llegar hasta carecer de moluscos o presentar baja diversidad-densidad; o tener conchas pequeñas y frágiles (Correoso, 2008).
Los moluscos terrestres tipifican desde el punto de vista biogeográfico a un país por sus especies endémicas, pues el genuino endemismo ocurre en especies con poca locomoción. Muchos moluscos pueden ser endémicos de algunas regiones y localidades del país, por lo cual se han clasificado y nombrado con nombres locales
(topónimos) Bulimulus antizanensis, Bulimulus cotopaxense. En general en Ecuador
16 Tabla1. Clasificación de la clase gastrópoda en Ecuador
Clase Subclase Orden Superfamilia Familia
Gastropoda
Prosobranchia Achaeogastropoda
Architaenioglossa Helicinoidea Cyclophoroidea Helicinidae Poteriidae
Pulmonata Stylommatophora
Orthalicoidea Bulimulidae Orthalicidae Amphibulimulidae Achatioidea Subulinidae
Clausilidae Oleacinoidea Oleacinoidea Streptaxoidea Streptaxoidea
Systrophiidae Strophocheiloidea Strophocheilidae
Camaenoidea Camaenidae
Systrophiidae Succionoidea Succineidae Polygyroidea Polygyridae Gymnomorpha Limacoidea Limacidae Ariolimacidae
Soleolifera Veronicellidae
Prosobranchia
Achaeogastropoda Neritaceae Neritidae Mesogastropoda Ampullaroidea Cerithiacea Ampullaridae Melaniidae
Truncatelloidea Hydrobiidae
Pulmonata Basommatophora
Lymnaeoidea Limneidae
Ancyloidea Planorbidae
Physidae
Unioboidea Ancylidae
Muteloidea Ellobidae
Fuente: Correoso. M, 2008.
3.5. IMPORTANCIA DE LOS GASTRÓPODOS
17
3.6. PROTECCIÓN AMBIENTAL DE LOS MOLUSCOS EN ECUADOR
En Ecuador está la segunda especie de caracol terrestre de mayor tamaño del mundo, Strophocheilus popeleiranus, (solo superado por el caracol centroafricano
Achatina) vive en los bosques húmedos tropicales a ambos lados de los Andes, sirve
de alimento a las comunidades indígenas amazónicas, pero son cada vez más raros, y poco estudiados aunque tiene potencialidades de crianza y manejo en cautiverio, por su carne, rojiza y agradable que no tiene nada que envidiar a los conocidos Scargots que se importan de Francia e Italia. Actualmente ningún molusco terrestre del Ecuador continental aparece en las listas rojas de especies amenazadas de la UICN y desafortunadamente hemos comprobado una marcada disminución de moluscos terrestres en el Ecuador, debido a acciones humanas como la deforestación incontrolada, la construcción de ciudades y represas en algunas zonas, la quema de los páramos y bosque secos, además de causas naturales como inundaciones, erupciones volcánicas u otras que extinguirán muchas especies de moluscos y otras especies incluso antes de poder llegar a conocerlas (Correoso, 2008).
Desde los primeros estudios a finales del siglo de XIX hasta la fecha, se hace notoria la falta de información precisa sobre moluscos terrestres y fluviales en los diversos textos de fauna de Ecuador, además en universidades y museos nacionales. Ecuador presenta cifras sobre las 300 especies lo que lo ubica comparativamente entre Colombia y Perú (Correoso, 2008).
3.7. GASTRÓPODOS COMO PLAGAS
La introducción de especies exóticas representa un peligro para la biodiversidad, si bien algunas pueden ser controladas otras han significado una amenaza de muerte, para la fauna y flora endémicas y además se constituyen plagas agrícolas, problemas para la salud, económicos y medio ambientales en los países afectados (Correoso, 2006).
18
valor estético y mítico religioso con marcada influencia sobre el hombre desde sus orígenes hasta nuestros días (Correoso, 2008).
Correoso (2005) reportó el principal molusco plaga para Ecuador, así como también es una de las plagas más importantes de invertebrados a nivel mundial, el caracol
gigante africano Achatina fulica (Bowdich, 1822); considerada una de las 100 plagas
[image:30.595.188.435.287.491.2]más importantes por su peligrosidad. Achatina fulica es una plaga a nivel continental, está presente en los países andinos, y en un reciente diagnóstico de especies exóticas en Venezuela se reveló su presencia junto a otras especies de moluscos. Los moluscos aunque lentos y aparentemente frágiles son plagas frecuentes a escala mundial y a menudo figuran entre los invasores más agresivos (Ojasti, 2001).
Figura 9. Achatina fulica
Elaboración: Autor: Ahoerstemeier. [En línea]
http://commons.wikimedia.org/wiki/File:Achatina_fulica_T hailand.jpg. Este archivo bajo licencia: Creative Commons Attribution/Share-Alike License 3.0 Unported.
Achatina fulica (Bowdich, 1822), es una especie introducida y esta presente
19
3.8. LOS CARACOLES Y LAS ENFERMEDADES QUE TRANSMITEN A LAS PERSONAS
El caracol africano Achatina fulica (Bowdich, 1822), habría llegado a varios países de manera ilegal para comercializar su carne; ahora se han convertido en la causa de proliferación de enfermedades que afectan de manera significativa a la salud humana. Este caracol puede cobijar parásitos que se alojan en tejidos fibromusculares y secreciones de moco del animal, causando diversas afecciones desde enfermedades de tipo respiratorio, hasta meningoencefalitis eosinofílica, angiostrongiliasis abdominal en los seres humanos, transmitida por la
Angiostrongylus cantonensis, lombriz que parasita los pulmones de las ratas,
también es vector de la bacteria gramnegativa, Aeromonas hydrophila, que causa diversos tipos de síntomas, principalmente en las personas con sistemas inmunológicos delicados, el contagio se produce a través del contacto de ojos, nariz, o boca con la mucosidad del caracol, o por el consumo de su carne si ésta no es adecuadamente lavada (UAB, 2011).
Estos caracoles son hospederos de parásitos que causan enfermedades graves a humanos, y animales domésticos, lo que ocasiona muertes por problemas intestinales (reportada para Brasil). El caracol puede trasmitir el parásito a los roedores domésticos, lo que contribuye a mantener el ciclo biológico del nematodo; de esta forma puede llegar a constituir graves epidemias de muy difícil diagnóstico, como indican médicos y parasitólogos, donde los roedores desempeñan un papel muy importante en su epidemiología. Siendo sus vectores más frecuentes rata doméstica (Rattus rattus), rata arrocera pigmea (Oryzomys fulvescens) y ratón de
campo (Oryzomys caliginosus). Los síntomas en estos animales son poco
manifiestos (Correoso, 2006).
3.9. LOS MOLUSCOS Y EL CAMBIO CLIMÁTICO
Durante las últimas dos décadas, se ha evidenciado que el clima global está cambiando y los gases que causan el efecto invernadero siguen incrementándose alarmantemente, se calcula que la temperatura mundial de la superficie ha aumentado en 0,6 °C (IPCC, 2001). La atmósfera responde a este incremento de gases con un aumento de la temperatura (Aalst, 2006). Estos cambios tienen efectos secundarios en los sistemas hidrológicos, ecosistemas terrestres y marinos (Midgley
20
son sobre especies de plantas (Scholze et al., 2006); y animales en diferentes vías
como su distribución, tamaño poblacional, estructura física, metabolismo y
comportamiento (IPCC 2001; Parmesan et al., 2003; Parmesan et al., 2004;
Cumming et al., 2006; Aalst, 2006).
La biodiversidad es continuamente transformada por el cambio climático. Las condiciones de cambio a través de la superficie del planeta, algunas veces rápidas, algunas veces lentas, en grandes o pequeños incrementos, resultan en la reacomodación de las asociaciones biológicas. Es importante resaltar que el efecto del cambio global, al influir en procesos ecológicos y funcionales, también influye en los servicios ecosistémicos que aportan los bosques al bienestar humano, alterando procesos naturales de los ecosistemas (IPCC, 2002).
Desde sus inicios la civilización humana ha inducido en el ambiente cambios que afectan la biodiversidad, esto incluye la de los moluscos terrestres (Preece, 1998). Desde el punto de vista biológico, las variaciones ambientales llevan a sucesos de expansión, dispersión, invasión, migración, extinción, surgimiento de nuevas especies y alteraciones de los ámbitos de distribución de las especies (Monge, 2006), alterando y marcando los patrones biogeográficos naturales de las especies. En la actualidad las variaciones más dramáticas las producen la expansión de las fronteras agrícola y urbana y el cambio climático mundial. La deforestación asociada a la expansión de las actividades humanas ha producido espacios abiertos, con ámbitos de temperaturas y humedad más amplios (Nebel, 1999; Smith & Smith, 2001), lo que favorece a unas especies y desfavorecen a otras (Lundholm, 2006, Clergeau et al., 2006, Shochat et al., 2006). Los moluscos terrestres son un grupo muy susceptible a variaciones en la humedad por lo que también son afectados por la deforestación (Hyman, 1967; South, 1992; Cook, 2001).
21
3.10. DECLIVE DE GASTERÓPODOS TERRESTRES
Actualmente, la tasa de extinción de especies es una de las mayores en la historia de la Tierra (Lawton & May, 1995). Ésta pérdida de diversidad biológica es preocupante, ya que la biodiversidad tiene importantes beneficios para la humanidad (Costanza et al., 1997; Chapin et al., 2000; Balmford et al., 2002). Por este motivo, se están desarrollando actuaciones encaminadas a evitar la progresiva pérdida de especies (Margules & Pressey, 2000). Sin embargo, aún son relativamente escasos los esfuerzos de conservación sobre la fauna invertebrada, a pesar de que supone aproximadamente el 97% de la diversidad de especies conocidas y cumple un importante papel en los ecosistemas (Ponder & Lunney, 1999; Nee, 2004). De hecho, menos del 0,3% de los invertebrados han sido evaluados en las listas de la UICN (Baillie et al., 2004). Precisamente, los moluscos son el grupo animal que registra el mayor número de extinciones documentadas, con los moluscos terrestres mostrando el mayor descenso poblacional en los últimos años (Lydeard et al., 2004).
Es posible que la influencia humana esté condicionando el empobrecimiento de la diversidad malacológica; en general, se ha detectado una declinación a nivel
mundial en la diversidad de especies de moluscos no marinos (Lydeard et al., 2004),
22
4. ÁREA DE ESTUDIO
El estudio se lo llevó a cabo en la Estación Científica San Francisco (ECSF), ubicada al sur de Ecuador, 3 ° 58'18'' S, 79 ° 4 '45'' E (Fig. 10), comprende bosques de montaña de la cordillera oriental de los Andes (Göttlicher et al., 2009 & Thies et al., 2012); se encuentra entre un rango altitudinal que va desde los 1 800 a 3 140 m s.n.m. con un régimen climático que se divide en dos épocas, la época seca de octubre a abril y la época lluviosa de mayo a septiembre, con precipitaciones anuales de 2 180 mm, con un máximo en los meses de junio, julio y agosto (Bendix
[image:34.595.87.581.266.583.2]et al., 2008).
Figura 10. Mapa del área de estudio
Elaboración: Autora
El bosque de montaña de la ECSF posee las siguientes formaciones vegetales de
acuerdo a Göttlicher et al. (2009), bosque siempreverde montano y bosque nublado
montano y dentro de estos distingue algunas clases de cubierta vegetal terrestre: 1)
bosque pre – montano; 2) bosque tipo I, barrancos 1 900 – 2 100 m; 3) bosque tipo
II, crestas y laderas superiores 1 900 – 2 100 m; 4) bosque tipo III, crestas y laderas superiores 2 100 – 2 250 m; 5) bosque tipo IV, crestas y laderas superiores 2 250 –
23
a 2 700 m; 8) helechos; 9) pastos; 10) arbustos y 11) áreas sin vegetación (laderas y deslizamientos de tierra).
Además constituye área de distribución de algunas especies de mamíferos, entre ellos, oso de anteojos (Tremarctos ornatus), puma (Puma concolor), tigrillo
(Leopardus pardalis). Más de 600 especies de aves representan alrededor del
37.5% de las especies de aves endémicas de Ecuador, en las que distinguen la torcaza (Zenaida auriculata), el águila (Stephanoaetus coronatus), pava barbada
(Penelope barbata), lorito (Amazona autumnalis), gaviota andina (Larus serranus) y
mirlo (Turdus merula) (Emck, 1999).
[image:35.595.92.539.421.635.2]El muestreo de moluscos se llevó a cabo en dos coberturas diferentes, bosque: intervenido (BI) y bosque no intervenido (BNI) (Fig. 11), el primero localizado en los alrededores de las instalaciones de la ECSF, caracterizado por la escasa cobertura vegetal, presencia de pastizales, helechos y malezas; el segundo se caracteriza por albergar abundante flora y fauna nativa intacta, y el uso que se le da a este bosque es exclusivamente para la investigación por parte del Consorcio Alemán de Investigaciones (DFG, Deutsche Forschungsgemeinschaft).
Figura 11. Tipos de bosque
24
5. METODOLOGÍA
5.1. MUESTREO DE GASTRÓPODOS TERRESTRES
[image:36.595.89.564.270.449.2]En los meses de julio, agosto y septiembre de 2012, se realizó 8 muestreos por mes en cada tipo de bosque, dando un total de 48 muestreos. Se ubicó 10 cuadrantes de 1 m2 con una secuencia diagonal (Fig. 12), en los cuales se realizó el muestreo durante 15 minutos tanto en la hojarasca, suelo superficial (explorando hasta 5 cm de profundidad), sotobosque, como en otros elementos del entorno (rocas, troncos), para disminuir el margen de error o sesgo se realizó una réplica para cada cuadrante (Rangel & Gamboa, 2006).
Figura 12. Esquema de muestreo de gastrópodos terrestres en el área de estudio. A la izquierda se presenta la secuencia diagonal y disposición de los cuadrantes de muestreo. A la derecha una fotografía de los cuadrantes de muestreo en el campo.
Se determinó los cuadrantes según las preferencias de los gastrópodos tales como ambientes relativamente húmedos, sombríos y con un alto contenido de materia orgánica (Pérez, 1999). Todos los especímenes recolectados fueron depositados en tubos plásticos, se los conservó en alcohol al 70%, luego fueron transportados al Laboratorio de Sanidad Animal y Zoonosis (LABSA) de la Universidad Técnica Particular de Loja, en donde posteriormente fueron separados y contabilizados de acuerdo a sus rasgos o características morfológicas.
25 5.2. ANÁLISIS DE DATOS
Como atributos para realizar los análisis de la diversidad de gastrópodos en el BI y BNI se determinó varios índices que se detallan más adelante. Además, se analizó la composición de especies de gastrópodos para ambos tipos de bosque.
5.2.1. INDICES DE RIQUEZA Y DIVERSIDAD
Se utilizó el programa EstimateS® versión 8.2 (Colwell, 2009) para determinar los siguientes índices en la estimación de la riqueza de especies:
Curva de acumulación de especies: Evalúa la representatividad de especies con el esfuerzo de muestreo (Gotelli & Colwell, 2001; Jiménez y Hortal, 2003).
Ace: (Abundance – Based Coverage Estimator), se basa en datos de
abundancia que superestiman la riqueza de especies cuando el número de muestras es bajo (Colwell & Coddington, 1994).
Ice: (Incidence – Based Coverage Estimator) está basado en el concepto estadístico de cobertura de muestreo, que se refiere a la suma de las probabilidades de encontrar especies observadas dentro del total de especies presentes (Colwell, 2006).
Chao 1: Estima el número de especies esperadas considerando la relación entre el número de especies representada por un individuo y el número de especies representadas por dos individuos en las muestras (Colwell & Coddington, 1994).
Chao 2: Requiere solo datos de presencia y ausencia y es el que presenta menor sesgo cuando las muestras son pequeñas (Colwell & Coddington, 1994).
Jacknife 1: Que al tener en cuenta las especies únicas tiende a reducir el sesgo de los valores estimados (Colwell & Coddington, 1994).
Jacknife 2: Se basa en el número de especies que ocurren solamente en una muestra y en el número de especies que ocurren exactamente en dos muestras (Colwell & Coddington, 1994).
26
Se utilizó el programa estadístico R-Project (Ihaka & Gentleman, 1996) para determinar:
Índice de Diversidad de Simpson, el cual muestra la probabilidad de que dos individuos sacados al azar de una muestra correspondan a la misma especie.
Todos estos índices fueron calculados separadamente para cada cuadrante y luego fueron promediados para cada muestreo (n= 24 para cada tipo de bosque, i.e. 8 muestreos por 3 meses). Una vez obtenidos los índices, se aplicó una prueba t-Student para evaluar la significancia de los valores promedios de los índices entre ambos tipos de bosque (p= 0.05).
5.2.2. COMPOSICIÓN DE GASTRÓPODOS
Varios métodos estadísticos han sido utilizados para evaluar las relaciones entre el uso del suelo y biodiversidad (ej. Análisis multivariados o regresiones lineales). En este estudio se utilizó la función „metaMDS‟ del paquete „vegan‟ del programa estadístico R-Project (Ihaka & Gentleman, 1996) para aplicar un Escalamiento Multidimensional no Métrico (NMDS por sus siglas en inglés) que permita identificar comunidades de gastrópodos asociadas a ambos tipos de bosque. El NMDS es un método que crea una matriz de distancia (Bray – Curtis) entre los objetos investigados, creando un espacio que es cada vez mayor cuando aumenta las diferencias entre estos objetos. En otras palabras, esa matriz de distancia permite crear una separación que facilita detectar grupos asociados, en este caso, a diferentes tipos de bosque. Además, el NMDS es comúnmente considerado como el método de ordenación sin restricciones más robusto en el análisis de la ecología de comunidades (Minchin 1987).
27
6. RESULTADOS
6.1. RIQUEZA Y DIVERSIDAD
En general, se registraron 1898 individuos, pertenecientes a 7 familias, 8 géneros y 5 especies, 4 taxones (taxón 1= Streptaxidae, taxón 2= Opeas, taxón 3= Leptinaria, taxón 4= Euconulus) no fueron identificados hasta el nivel de especie. El BNI presenta el mayor número de individuos capturados (1706) así como también el mayor número de especies (9) (ver Tabla 2). Independientemente, tanto en el
bosque no intervenido como en el bosque intervenido S. heligmoida es la especie de
[image:39.595.92.563.372.566.2]mayor abundancia con 1 596 individuos para el BNI y con 171 para el BI; por otro lado Euconulus sp1 y E. striata registran el menor número de individuos, con 1 individuo cada una, exclusivamente para el BNI.
Tabla 2. Visión general de los gastrópodos encontrados en el BI y BNI durante todo el muestreo.
FAMILIA GÉNERO
ESPECIE
TIPOS DE
BOSQUE TOTAL
BNI BI
Systrophidae Systrophia Systrophia heligmoida 1596 171 1767
Streptaxidae Taxón 1 45 11 56
Subulinidae Opeas Taxón 2 21 5 26
Subulinidae Obeliscus Obeliscus cuneus riparius 17 3 20
Subulinidae Leptinaria Taxón 3 13 1 14
Scolodotidae Happia Happia guayaquilensis 7 0 7
Helicinidae Helicina Helicina rhynchostoma 5 1 6
Eucunulidae Euconulus Taxón 4 1 0 1
Spiraxidae Euglandina Euglandina striata 1 0 1
TOTAL 1706 192 1898
28
Gráfico 1. Curva de acumulación de especies para el total del área muestreada.
[image:40.595.157.468.53.232.2]La diversidad de especies de gastrópodos en ambos tipos de bosque fue baja. Sin embargo en promedio el BNI presenta mayor diversidad que el BI (Tabla 3), excepto para la diversidad de Simpson. Los resultados de la t de Student demuestran que existe diferencia altamente significativa (p > 0,001) entre BNI y BI, corroborando los resultados en la mayoría de los índices de riqueza y diversidad; adjudicando mayor riqueza y diversidad de especies al BNI, a excepción de la diversidad de Simpson. (Gráfico 2).
Tabla 3. Índices de diversidad calculados para BI y BNI.
Índices Bosque No Intervenido Bosque Intervenido Mínimo Media Máximo Mínimo Media Máximo
Ace 2.0 5.0 11.0 1.0 2.4 7.5
Ice 2.2 7.4 21.6 1.0 2.6 12.6
Chao1 2.0 4.1 11.0 1.0 1.8 4.5
Chao2 2.0 4.3 10.4 1.0 2.0 6.7
Jacknife1 2.9 5.2 10.6 1.0 2.3 6.7
Jacknife2 3.7 6.3 13.8 0.3 2.8 9.1
Bootstrap 1.2 4.0 8.5 1.0 1.9 5.1
Simpson 0.0 0.1 0.4 0.0 0.2 0.6
0 2 4 6 8 10
1 30 59 88
29
Gráfico 2. Comparación de los índices de riqueza y diversidad para bosque intervenido (BI) y bosque no intervenido (BNI). La diferencia estadística fue p > 0.001 (t-Student) para casi todos los índices
calculados. Únicamente el índice de Simpson no presentó diferencia entre ambos tipos de bosque (p
30 6.2. COMPOSICIÓN DE ESPECIES
En la ordenación NMDS, con un stress que indica alta calidad en la ordenación (0.03), en la comparación de los tipos de bosque se distinguen claramente 2 grupos, un grupo bastante desorganizado y disperso correspondiente al BI y otro grupo homogéneo correspondiente al BNI (Gráfico 3).
31
El análisis de similitud ANOSIM con un R global =0,916 y p=0,001 indica alta significancia estadística en los resultados del análisis, con una marcada diferencia entre las comunidades de gastrópodos de BI y BNI, baja diferencia en las muestras del BI y mínima diferencia en las muestras del BNI, siguiendo un patrón totalmente similar al análisis multivariante (NMDS) anteriormente aplicado (Gráfico 4).
32
7. DISCUSIÓN
Considerando que diferentes características del hábitat determinan la distribución de moluscos terrestres (Correa, 2007), el BNI registró el mayor número de especies de gastrópodos terrestres, debido a que proporcionan una considerable cobertura vegetal sobre el suelo, protección contra la desecación, abundante hojarasca y alimento, lo que favorece la presencia de estos organismos (Boycott, 1934; Berazain, 1981; Álvarez et al., 2003; Correa, 2007). Por el contrario, el menor número de especies de moluscos terrestres hallados en el BI, se explica principalmente por la escasa cobertura vegetal, y más iluminación sobre el suelo (Hunter, 1964; INEGI, 1980; INEGI 1981; Correa et al., 2007).
Destacando el trabajo de Álvarez et al., (2003) el cual encuentra la mayor parte de
especies de moluscos terrestres en hojarasca, en esta investigación las especies registradas estuvieron también presentes en hojarasca y humus. Esto se debe a que esta proporciona un clima adecuado, refugios y alimento a diversas especies, por lo que incide en su distribución (Petersen & Luxton, 1982; Solem & Climo, 1985; Locasciulli & Boag, 1987; Thompson & Correa, 1994; Correa, 2007).
Las curvas de acumulación de especies es un método importante en los estudios sobre biodiversidad, por la facilidad en la interpretación de resultados y por el uso de este método en numerosos trabajos sobre inventarios biológicos (Jiménez, 2003). En el caso de gastrópodos terrestres este método se ajustó eficazmente, mostrando un esfuerzo de muestreo óptimo, alcanzando la asíntota de especies esperadas.
Las zonas con algún grado de perturbación, soportan una menor riqueza y abundancia de especies en relación a los bosques nativos (Gasca & Ospina, 2000; Connell, 1978); enunciado que coincide con el presente estudio, el BI registra el menor número de especies y abundancia con respecto al BNI.
Con base en la diversidad de especies, tal como sugiere Pérez et al., (2006) en el
trabajo de Gonzales et al., (2010) las zonas conservadas de bosque poseen mayor
diversidad de especies en relación a las zonas con algún grado de perturbación, la mayor diversidad de especies en este estudio la registra en BNI en relación al BI.
33
métodos multivariantes que diferencian la composición de gastrópodos terrestres de acuerdo a la perturbación de hábitats.
Asumiendo que las especies del género Systrophia se distribuyen principalmente en
América del Sur hasta los 4 400 m de altitud (Romero & Ramírez, 2011), desempeñan una función ecológica como depredadores de otros moluscos, y son abundantes en ambientes húmedos, como el suelo de los bosque y zonas tropicales
(Correoso, 2008). En esta investigación la especie Systrophia heligmoida (Orbigny,
1835) registra el mayor número de individuos tanto para el BNI como para el BI.
34
8. CONCLUSIONES
El esfuerzo y la temporalidad del muestreo utilizado en este estudio fue el adecuado registrando un número considerable de especies presentes según las curvas de acumulación de especies.
La diversidad de especies fue mayor para el BNI con respecto al BI
presentado una variación en los patrones de diversidad.
La composición de especies es diferente entre los dos tipos de bosque estudiados, además es heterogénea en el BI y más homogénea en el BNI.
Ecológicamente el bosque tropical de montaña presenta una gran variedad de
hábitats y condiciones climáticas únicas, que en conjunto hacen que albergue una mayor cantidad de organismos o formas de vida con respecto a otros ambientes.
Los gastrópodos terrestres reflejan los niveles de perturbación, manifestando
un descenso de individuos y especies de acuerdo al tipo de bosque, lo cual corrobora su uso como indicadores de calidad ambiental.
Systrophia heligmoida, es una especie característica del bosque tropical, en
este ambiente encuentra las condiciones adecuadas para su desarrollo.
El conocimiento actual de la malacofauna terrestre en Sudamérica,35
9. RECOMENDACIONES
Realizar investigaciones con monitoreos en diferentes escalas tanto
espaciales como temporales para conocer más acerca de la composición de especies, en este tipo de ambiente.
Proponer la implementación de buenas prácticas ambientales, en el área de
bosque intervenido, que causen el menor daño ambiental posible.
36
10.
BIBLIOGRAFÍA
Aalst, M. 2006. The impacts of climate change on the risk of natural disasters. Disasters 30: 5 – 18.
Aktipis, S.; Giribet, R.; Lindberg, R. & Ponder, W. 2008. Gastropoda: An overview and analysis, p. 201- 237. In W.F. Ponder & D.R. Lindberg (eds.). Phylogeny and Evolution of the Mollusca.Universidad de California, Berkeley, EEUU.
Álvarez, J.; Naranjo, E. & Guevara, S. 2003. Perspectivas de estudios de acumulación de hojarasca en el subdosel y animales asociados. El caso de los moluscos. Pp. 80-87. Instituto de Ecología. Universidad Nacional Autónoma de México. Xalapa, México.
Aycaguer, C. 2006. Tecnología de los productos dela pesca, Malacología. Área de Ciencias del Mar. Uruguay.
Aycaguer, C. 2006. Elementos de Malacología, Aplicada a la Tecnología de Productos Pesqueros.
Baillie, M.; Hilton, C. & Stuart, N. 2004 IUCN red list of threatened species: A global species assessment. Gland y Cambridge.
Balmford, A.; Bruner, A.; Cooper, P.; Costanza, R.; Farber, S.; Green, E.; Jenkins, M.; Jefferiss, P.; Jessamy, V.; Madden, J.; Munro, K.; Myers, N.; Naeem, S.;Paavola, J.; Rayment, M.; Rosendo, S.; Roughgarden, J.;Trumper, K. & Turner, K. 2002. Economic reasons for conserving wild nature. Science, 297: 950-953.
Barnes, R. 1996. Zoología de los invertebrados, 6ª ed. Cap. 10: Moluscos (pp. 335494).
Barrientos, Z. 2003. Lista de especies de moluscos terrestres (Archaeogastropoda, Mosogastropoda, Archaeopulmonata, Stylommatophora, Soleolifera) informadas para Costa Rica. Revista Biológica Tropical. 51(3): 293-304.
37
Berazain, R. 1981. La Zona de Vegetación Serpentinícola del Jardín Botánico Nacional. Revista del Jardín Botánico Nacional 2: 151-181.
Boycott, A.1934. The habitat of land mollusca in Brittain. Journal of Ecology 22: 1-38.
Bücker, A., Sondermann, M.; Frede, H. & Breuer, L. 2010. The influence of land - use
on macroinvertebrate communities in montane tropical streams – a case study from
Ecuador.
Camarda, C. & Suarez, L. 2004. Ecología Acuática, Estudio avanzado de Moluscos.
Chapin, S.; Zavaleta, S.; Einer, T.; Naylor, L.; Vitousek, M.; Reynolds, L.; Hooper, V.;Lavorel, S.; Sala, E.; Hobbie, E.; Mack, C. & Díaz, S. 2000. Consequences of changing biodiversity. Nature, 405: 234-242.
Clergeau, P.; Jokimäki, J. & Snep, R. 2006. Using hierarchical levels for urban ecology. Trends Ecol. Evol. 21: 660-661.
Connel, J. 1978. Diversity in Tropical Rain Forest and Coral Reefs. Science. New Series. 199: 4335: 1302-1310.
Colwell, R. & Coddington, J. 1994. Estimating terrestrial biodiversity through extrapolation. Philos. Trans. R. Soc. Lond. B 345, 101-118.
Colwell, R. 2006. EstimateS: Statistical estimation of species richness and shared species from samples. Version 8. Available at: http://www.purl.oclc.org/estimates.
Colwell, R. 2009. EstimateS: Statistical estimation of species richness and shared species from samples. Version 8.2. User's Guide and application published at: http://purl.oclc.org/estimates.
Cook, A. 2001. Behavioural ecology: on doing the right thing, in the right place at the right time, p. 447- 488. InG.M. Barker (ed.). The biology of terrestrial mollusks. Nueva York, EEUU.
Correa, A. 2003. Gastrópodos terrestres del Noreste de México. Revista Biológica Tropical 51: 507-522.
38
Correoso, M. 2005. Los moluscos esos interesantes y valiosos invertebrados. Revista Clubing Magazín. Edición Sierra. Año 6.No 65. Ecuador.
Correoso, M. 2005. Lineamientos generales de la introducción de especies ¿enriquecimiento de la biodiversidad o problemas ecológicos?” Revista Geoespacial. No. 2. Escuela Politécnica del Ejército. Ecuador.
Correoso, M. 2006. Estrategia preliminar para evaluar y erradicar Achatinafulica (Gastropoda: Achatinaceae) en Ecuador. Sangolquí. Ecuador.
Correoso, M. 2008. Los Moluscos terrestres y fluviales del Ecuador. La biodiversidad desconocida. SIMBIOE. Quito Ecuador.
Costanza, R.; D‟Arge, R.; De Groot, R.; Farber, S.; Grasso, M.; Hannon, B.; Limburg, K.; Naeem, S.; O‟Neill, V.; Paruelo, J.; Raskin, G., Sutton, P. & Van Den Belt, M. 1997. The value of the world‟s ecosystems services and natural capital. Nature, 387: 253-260.
Cowie, R. & Robinson, A. 2003. The decline of native Pacific island faunas: changes in status of the land snails of Samoa through the 20th century. Biol. Conserv. 1: 55-65.
Cumming, G. & Vann, D. 2006. Will climate change affect ectoparasite species ranges? Global Ecology and Biogeography 15: 486 – 497.
Durance, I. & Ormerod, S. 2007. Climate change effects upland stream macroinvertebrates over a 25 – year period. Global Change Biology 13: 942 – 947.
Emck, P. 1999. Climatic conditions in the Cordillera de Numbala (Podocarpus National Park), Southern Ecuador.
Fernández, P. 2007. Características Generales de los Gastrópodos Marinos y Terrestres.
Forcelli, D. 2000. Moluscos Magallanicos Guía de Moluscos de Patagonia y Sur de Chile parte 1.
39
Gonzalez, N.; Ochoa, S.; Cambranis, E.; Lara, O.; Perez, I. & Ponce, A. 2010. Gasterópodos terrestres asociados a un paisaje agropecuario y a un referente ecológico en el sureste de México.
Gotelli, N. & Colwell, R. 2001. La cuantificación de la biodiversidad: Procedimientos y dificultades en la medición y comparación de la riqueza de especies Ecology Letters 4379-391.
Göttlicher, D.; Obregón, A.; Homeier, J.; Rollenbeck, R.; Nauss, T.; & Bendix, J. 2009. Land-cover classification in the Andes of southern Ecuador using Landsat ETM+ data as a basis for SVAT modeling. Department of Geography, Laboratory for Climatology and Remote Sensing, University of Marburg, 35032 Marburg, Germany. Plant Ecology, Albrecht-von-Haller-Institute for Plant Sciences, University of Göttingen, 37073 Göttingen, Germany.
Hunter, W.1964. Physiological aspect of ecology in non marine molluscs. Pp. 83-126. In: K. M. Wilbour & C. M. Yonge (Eds.). Physiology of Mollusca. Vol. 1. Academic Press. New York.
Hyman, L. 1967. The invertebrates: Mollusca 1(Vol VI). McGraw-Hill, Nueva York, EEUU.
Ihaka, R. & Gentleman, R. 1996. R: a language for data analysis and graphics. Journal of Computational and Graphical Statistics 5: 299–314.
Instituto Nacional de Estadística, Geografía e Informática. 1980. Carta de
precipitación total anual. Esc. 1: 1000,000. Secretaría de Programación y Presupuesto. México.
Instituto Nacional de Estadística, Geografía e Informática.1981. Carta de humedad de suelo. Esc. 1: 1000,000. Secretaría de Programación y Presupuesto. México.
Jiménez, A. & Hortal, J. 2003. Las curvas de acumulación de especies y la necesidad de evaluar la calidad de los inventarios biológicos. Revista Ibérica de Aracnología 8: 151 – 161.
Lawton, J. & May, R. 1995. Extinction rates. Oxford University Press. Oxford.
40
López, A. & Urcuyo J. 2009. Moluscos de Nicaragua. II Gastropodos Centro de Malacología/Biodiversidad Universidad Centroamericana-UCA. Nicaragua.
Lundholm, J. 2006. How novel are urban ecosystems. Trends Ecol. Evol. 21: 659-660.
Lydeard, C.; Cowie, W.; Ponder, A.; Bogan, P.; Bouchet, S.; Clark, K.; Cummings, T.; Frest, O.; Gargominy, D.; Herbert, R.; Hersheler, K.; Perez, B.; Roth, M.; Seddon, E.; Strong, E. & Thompson, F. 2004. The Global decline of nonmarine mollusks. BioScience 54: 321-330.
Margules, C. & Pressey, R. 2000. Systematic Conservation Planning. Nature, 405: 243-253.
Martínez, R. & Piñango, H. 2008. Aspectos y Características Generales del Filo Mollusca.
Midgley, F.; Hannah, L.; Millar, D.; Rutherford, M. & Powrie, L. 2002.Assessing the vulnerability of species richness to anthropogenic climate change in a biodiversity hotspot.Global Ecology & Biogeography 11: 445 – 451.
Mille, S. & Pérez A. 2003.Géneros más Comunes de Moluscos Gasterópoda y Pelecycopoda en México. México.
Minchin, P. 1987. An evaluation of the relative robustness of techniques for ecological ordination. Vegetatio 69:89–107
Miranda, M. & Cuezzo, M. 2010. Biodiversidad de gasterópodos terrestres (Mollusca) en el Parque Biológico Sierra de San Javier, Tucumán, Argentina. Revista Biológica Tropical 58: 3.
Monge, J. 2006. Ecological biogeography: a review with emphasis on conservation and the neutral model. Gayana 72: 102-112.
Moreno, C. & Halffter, G. 2000. Assessing the completeness of bat biodiversity inventories using species accumulation curves. J. Appl. Ecol., 37: 149-158.
41
Nebel, B. 1999. Ciencias Ambientales. Ecología y desarrollo sostenible. Prentice Hall, México D.F., México.
Nee, S. 2004. More than meets the eyes. Earth‟s real biodiversity is invisible, whether we like it or not. Nature, 429: 804-805.
Ojasti, J. 2001. Especies exóticas invasoras. Estrategia regional de biodiversidad para los países del trópico andino convenio de cooperación técnica no rembolsable. Caracas – Venezuela.
Panel Intergubernamental de Expertos sobre el Cambio Climático. 2001. Cambio Climático: Informe de síntesis. Evaluación del Grupo Intergubernamental de Expertos Sobre Cambio Climático.
Panel Intergubernamental de Expertos sobre el Cambio Climático. 2002. Cambio climático y biodiversidad. Gitay, H; Suárez, A; Watson, RT; Dokken, DJ. eds. 85 p. (Documento técnico V del IPCC). Consultado 11 dic. 2007. Disponible en: http://www.ipcc.ch/pub/tpbiodiv_s.pd.
Parmesan, C. & Yohe, G. 2003. A globally coherent fingerprint of climate change impacts across natural systems. Nature 421: 37 – 42.
Parmesan, C. & Galbraith, H. 2004. Observed impacts of global climate change in the U.S. Pew Center on Global Climate Change.
Patzelt, E. 2000. Fauna del Ecuador. 2 edición. Quito, Ecuador.
Pérez, A.; Vilaseca, J. & Zione, N. 1996. Sinecología básica de moluscos terrestres en cuatro formaciones vegetales de Cuba. Revista Biológica Tropical 44(1): 133-146.
Pérez, A. 1999. Estudio Taxonómico y biogeográfico preliminar de la malacofauna continental (Mollusca: Gastropoda) del Pacífico de Nicaragua. Tesis Doctoral, Universidad del País Vasco/Euskal Herriko Unibertsitatea, País Vasco, España.