-.. --,-+.*...- ,.""^_c~ ._.- 4 1 _ _ 1 _ _ - - + , - " . _ ~ ( _ 1 - ~ . . ~ ~ -
Universidad Autónoma
Metropolitana-Iztapalapa.
.
/Ciencias Básicas e Ingeniería
'Aplicación de la teoría de procesos estocásticos
en
la
configuración del ADN.
/Doctorado en Ciencias
'Física)
Tesis que presenta:
J
M.C.
Osbaldo Resendis Antonio.
Asesor:
J
Dr.
Leopoldo
García-Colín
S.
Índice.
General
1 Introducci6n 3
1.1 Antecedentes
. . .
31.2 Aspectos generales
. . .
51.3 propiedades Físic+Químicas del ADS
. . .
71.3.1 Estructura.Molecular.
. . .
7
1.3.2 Fenomenología de la desnaturalización térmica
. . .
:. . .
92 Antecedentes Teóricos de la desriaturalizaci6n
.
2.1 Introducción. . .
13 13 2.2 Enfoque Estadístico. . .
132.2.1 Modelo de Ising
. . .
132.2.2 hfétodo matricial de Gibbs y 1)iMnrzio . . . 16
2.2.3 Modelo de Reiss-.lcQuarrie-ll[.Tn.ii(.-..}ieti . . . 19
2.2
.. 1 ,\ Iodelo de Poland . . . 232.
.$ Enfoque termodinámico . . . 27Termostática fundanientd t’ii lit (I[. ..riiiiii l.alizacii>n di 1 ADN
. . .
27. . ~ Termodinámica de Oiigóniero~ . . . 29
‘2 :I 1
,
. > 3 \lodelos estocasticos CII la descripcicjii (IC la dcLiiaturaliziicih térmicade1
-2DN 35 .I . I Iiitroduccióri 35:5.2
~ ~ O d e l o s
Teóricos de la desnaturalizaci6ri t6rrriica con mtlependercia temporal . . 363.2.1
.j,2.2 8.2.3
Método de Matrices Est ocitst'cas.
.
, , ,.
,. . .
,. . .
. , .. .
..
, , , 36 Desnaturalizaci6n de w a cadena con secuencia simple.. . .
, , ,.
.
38 Desnaturalización en cadenas de ADN correspondientes a organismos SU- periores...
.
.. .
.. . . . . . . .
.. .
. . , , , ,. .
,. . .
, ,.
42
3.3 hlodelos Teóricos de la drsnati ralización térmira con dependencia temporal. , . 513.3.1 Iritroducción .
.
. . . . .. .
, ,. .
. . . ..
. . . ..
. .. . . .
51
3.3.2
52
3.3.3 Formalismo de Fokker-Planck.
.
. . . . , . . , . . . , , , , . , . j; Relajación térInic? cn i'na cadena simple de t'o1yC':Polg.G. . . . . , . , , ,4
TeoríaEstadística
de ladesnatiiralizaci6ri
de
cadena triple
de
ADN. 06 4.1 Introducción. . . .. .
. . .. . . . .
. . . ..
. . . . . , , , , , , , . 66-1.2 Estructura Molecular del ADN Triple y feriomeriulogía. . , . , , , , , , , , , . . . 67
-1.3 >lodelo de Matrices Estocástic:as.
. . . .
. . . . , , ..
, , , , , , ,. .
,.'
, 694.4
Resultados y conclusiones. . . . .
. . . . . . . .. .
, ,. .
, , , , . ,.
, . , , , 72Capítulo
1
Introducción
1.1 Antecedentes
La incursión de la física en el campo de la biología molecular fue inaugurado por
E.
Schr6edinger en septiembre de 1943, cuando él impartió una serie de conferencias, tituladas ?What is life?”, en el Trinity College de Dublín [l]. Un año después, la editorial Cambridge Unibersity Press edit6un
pequeño libro que condensa las ideas esenciales de estas conferencias [2] en torno a dos temas que hasta nuestros dias son centrales en la biología: la naturaleza de la herencia y la termodinámica de los sistemas vivos.En el primer tema, Schrtkdinger expres6 sus reflexiones sobre el mecánismo que permite a los organismos transmitir la información de generación en generación. En 1935, Timoféeff- Ressovsky, Zimmer y DelbrUck [3] presentaron un artículo en el cual, a partir del análisis de
las mutaciones en la mosca de
la
fruta (Drosophila melanogaster),
inferían el tamaño de la”unidad genética” (denominada gen). 1,os resultados de éste articulo condujeron a Schrüedinger a inducir que la naturaleza del gen wms stfa en una especie de cristal aperi6dico el cual codificaba
la información a través de su estrucíu b. La importancia de esta aseveración no fue confirmada sino hasta la aparición de dos ies~.lt:dcs cruciales en la Biología molecular. El primero se dio en el año de 1944, en el que los e, pe ,irentos realizados por 0.Avery sugerian firmemente que la información genética
sc
alnacenaba en el ácido deJoxirribonucleico; (ADN) y no en las>
este campo fue en
1953,
año en el que Watson y Crick pr, delADN
e implidtamente el mecanismo de replicación de resultados, pilares de la biologíc molecular, confirmaron, de Schroedinger sobre la unidad genética de los seres viv de estehecho
han permitido una serie de avancessorp
molecular, la ingeniería genética y la medicina genómicaLa segunda cuestión planteada por Schroedinger con da de un sistema vivo en
Lérminos
de las leyes termcd segunda ley. Debido a que la condición del sistema en erio, una respuesta concluyente en esta linea continua sie trrmodinámica irreversible de este siglo
[l].
Los trabajos pioneros de Delbrtick y Schroedinger h matemáticos incursionen en la investigación de los fenón las contribuciones científicas para su análisis cientffico e esta tesis. Sin embargo, en la mayoría de estos trabaj común, representar matemáticamente
un
sistema biológ leyes fundamentales que gobiernan los fenómenos de la terrogante dista mucho de ser definitiva. En este contc contribuciones'que el autor y el Prof. Leopoldo Garcia-' muy específico de la biología molecular, la desnaturaiizac experimental, la desnaturalización consiste en el rompimilas
bases nitrogenadas de nucleótidos complementarios, 1( tener la estructura secundaria de la biomolécula. Con la rsta tesis el lector encontrará modelos físicos originales qroii las mediciones experimentales reportadas comunmenl
ebperanios que estas contribuciones cientificas contenids
c k i . mriquecer y a estimular s u estudio desde un marco
el de la Física.
4
pusieron la estructura tridimensional
I
i la informaci6n genética[5].
Estos dose manera contundente la aseveración
s. Desde entonces, las consecuencias
mdentes en el campo de la biología >or citar algunas [6].
ste en explicar la estructura ordena- Amicas, con especial atención en la tudio rn altamente fuera del equiiib-
do uno de loa principales retos de la
n estimulado a que muchos físicos y
enos biologicus, cuya compilacihn de iistorico queda fuera del alcance de
s podemos distinguir una tendencia'
iocon la finalidad de vislumbrar las
iateria viva. La respuesta a esta in- :to, ésta tesis contiene las modestas bun S. han desarrollado en un tema 6n del ADN. Desde el punto de vista nto de los enlaces de hidrógeno entre
i cuales son los responsables de man-
inaiidad de entender éste proceso, en
ie conducen a resultados consistentes
: en la literatura. Sin m&s preambulo
1.2 Aspectos generales.
La predicción experimental del comportamiento de la desnaturalizaci6n t&mica del ADN (&ido desoxirribonucléico) ha sido
un
tema que desde hace tiempo se ha tratado de fundamentar con base en la Termodinámica y la Mdnica Estadística en equilibrio[7,8].
Los
estudios realizados desde el enfoque termodinámico, el cual ea completamente fenomenol6gico, han permitido ciertogrado de avance en la predicci6n de la temperatura de fusión de cadena? cortas de A DN (&I
orden de 20 pares de bases)
[Q].
Por
temperatura de "fusión" nos refairnos a la tern,\- W ~ W Q iila cual un conjunto de moléculas idénticas de ADN, inmersas en una solución wuosa, se separin en dos cadenas poiiméncas
(figura
1.1).. + I Figura 1-1: Desnaturalización térmica del ADN
Por otra parte, el enfoque estadístico contiene dos limitaciones :mportantes. En la primera, se asume la condición de equilibrio térmico entre la cadena de ADN J la solucióii acuosa en
partición de la cadena de
ADN
se aproxima asumiendo que los nucleótidos interaccionan única y exclusivamente entre primeros vecinos. E s característico de este desarrolloteórico
introducir varios parámetros ajustables, los cuales representan una serie de contribucionesasociadas
apropiedades de la cadena polimérica. En esta categoría entran la entropía poiimérica y el
grado de nucleación, por citar algunas
[lo,
111. En este contexto, dos aspectos han motivdo al desarrollo de esta tesis. Por una parte, la aplicación de la teoría de las matrids estocásticos [I21a este problema, y por otra, los recientes datos experimeiitales de la termcdinAliir:.i di? pares
de nucleotidos acoplados [9]. Ambos aspectos
nos
han conducido a cuestionar la neíssir(ad de incluir parámetros ajustables en la descripción, o bien si la descripción de la desnaturalización del ADN es pmible realizarla considerando solo la inforniación de la estabilidad termodinámica entre pares de nucleótidos. Proponer una respuesta a esta cuestión constituye uno de los ejes centrales de esta tesis.Por otra parte, al momento de revisar la bibliografía referente a este problema, es evidente
que una descripción de cómo el ADN logra su equilibrio termodinámico con el medio acuoso
durante el proceso de desnaturalización es inconclusa. Desde el punto de vista experimental,
la condición de relajación térmica se logra cuando la velocidad de calentamiento'es "suficiente-
mente lenta"
,
típicamente del orden de 0.8 'C/min, como para que a cada tiempo de medición el equilibrio térmico se haya alcanzado.A
juicio del autor, la caracterización de estos tiempos derelajación es de suma importancia para establecer el límite de validez de los desarrollos estadís- ticos y tal caracterización es posible desde el enfoque de la teoría de los prooesos estochticos. De esta manera, otro de los ejes, de esta tesis se centra en sustentar teóricamente esta línea de investigación.
'
Finalmente, deseo comentar que los estudios de la cadena de ADN doble estan muy desar- rollados y avanzados, sin embargo no es así para cadenas triples de ADN [13,14]. La estructura espacial de esta biomolécula es de vital importaricia en los fenomenos de transcripcion molecular y en las tknicas médicas y biotecnológicas actuales [fi. 15.16). Los modelos que pretenden de-
scifrar las propiedades de la triple hélice son rralriirrite esca~os y las mediciones experimentales aiúi estan incompletas [17]. El iíltimo eje de esta tesis. se concentra en sugerir una metodología simple de la desnaturalización de esta estructura particular del ADN, la cual es de suma ini- portancia para estudios actuales y futuros. en el campo de la Biologfa Molecular.
El
número de interrogants respecto a las contribuciones de esta thises
reaimente superior,sin embargo, estas propuestas constituyen un modesto avance en la comprensi6n integral del comportamiento de la molécula que esta iduyendo en la ciencia de este siglo, el ADN.
1.3 Propiedades FísiceQuímicas del
ADN.
1.3.1
Estructura Molecular.
La molécula de ADN es un macromolhla constituida por dos hebras poliniérimxw que se en.. lazan formando una estructura espacial de doble hélice [5]. A
su
vez, cada una de estns hebraspoliméncas es el resultado de la unión de varias unidades moleculares fundamentales Uamadas nucleótidos.
Los
nucleótidos son estructuras moleculares formadas por el enlace de tres elemen--
tos químicos: un grupo fosfato, un azúcar (pentosa) y una base nitrogenada(Fzgi.2).
Figura
1-2:
Estructura simplificada de un nucleútido.Las bases nitrogenadas que existen en la naturaleza se clasifican como purinas: Adenina
y Guanina; y pirimidinas: Citocina, Timina y Lrracilo. Cada una de las bases forman un nucleósido cuando estas se enlazan a una ribosa mediante un enlace glucosido entre un átomo de nitrogen0 de la base y uno del carbono del azúcar
[13,18].
Cada una de las hebras que constituyen el ADN se forma debido a que un p i p o fosfato simultaneamente se enlaza a doscarbonos de distinto nucleósido
(Fig
1.3).
Figura 1-3: Estructura de cada una de las cadenas polimérica que constituyen el ADN.
nucleótidos es una consecuencia de esta reactividad bidireccional del grupo fosfato. Todos los organismos vivos están constituidos de estos bloques fundamentales de la vida. Sin embargo, una propiedad que caracteriza al ADN de cada organismo es
su
secuencia. La escritura de la secuencia de un ADN par.ticuiar se inicia en la dirección del carbono 5' al carbono 3 de los azúcares al borde de la secuencia. Para facilitar la notación normalmente se emplean las letras de los nucleótidos leidos en esa dirección, por ejemploCGGTA.
LL
estabilidad estructural de la doble hélice del ADN se debe fundamentalmente a dos clases de interacciones [13,18]. Laprimera se asocia a los enlaces de hidrógeno generados entre las bases nitrogenadas de las hebras
opiiwtas (Fig
1.4).
Estos puentes de hidrógeno entre las bases nitrogenadas no son azarosos,sino más bien siguen las reglas de acoplamiento de \Vatson y Crick [5,13], es decir desde el
piinto de vista termodinámico, el acoplamiento más estable entre las bases nitrogenadas se logra
ciiando 1:~ Adexiina se enlaza con la Timina y la Citocina se une preferentemente con la Guanine. La segunda, se debe a interacciones cruzadas entre pares de nucleótidos vecinos
[lQ].
Existen otros fact,ores que iduyen en la estructura secundaria del ADN, es decir la estructura particulaide doble hélice, que son aspectos propios del medio en el que la molécuia se disuelve. Estos
factores pueden ser el
pH,
las concentraciones de sales en la solución acuosa y la temperatura.La variación de alguno de essos factores externos produce como consecuencia un cambio en la estructura secundaria de la molécula. La más simple de estas transiciones es una que pasa de un estado ordenado, identificado p o r la doble hélice, a uno desordenado, caracterizado por
dos hebras total o parcialmente separadas (fzgura 1.1). Esta transición coastituye el objeto de estudio central de esta tesis. En l a siguiente sección presentamos ;a caracterización fíwa y
experimental de este fenómeno, asi como la principal terminologia de este campo.
Figura 1-4: Estructura del ADN
1.3.2
La desnaturalización térmica de la molécula del
ADN
consiste en el rompimiento de los enlacesde hidrógeno. los cuales son responsables de mantener la estructura de doble hélice, inducidos por un incremento en la temperatura. La metodología experimental de este proceso consiste en diluir varias cadenas idénticas de ADN en una solución acuosa a condiciones ñsioiógicas bien definidas. Desde el punto de viste fe,iomenológico esta transición se manifiesta con cambios en
cantidades que son medibles experimentalmente y las cuales permiten caracterizar dicha traosi- ción. por ejemplo cambios en la absorción de UV 6 en la densidad de la solución. Es importante rcsaltar que, de acuerdo con la mecánica es' adístiwi, estas mediciones son el resultado de pro- nicdiar sohw iin ensamble estadístico de sistema [8]. La más común de estas propiedades es~el
Ilariiatlo Pfecto hipercrómfco. Este efecto consiste, en un incremento en la, absorción de UV (280 nm) en las bases nitrogenadas cuando estas se er.mentran en un estado libre con respecto ii su estado enlazado a través de los puentes de hidr6:erio. De esta manera la hipercrórzicihc: de
las bases nitrogenadas, es una propiedad que permite cuantizar el número promedio di. p.tres
de bases de Watson y Crick que se encuentran en estado enlazado como función de la tem-
peratura en la solución aci.osa [5,
lo].
Con la finalidad de caracterizar la transición se defineuna temperatura de fusi6n
T,,
la cual identifíca el estado en el que la fracción total de pares- de bases acopladas es;.
Desde el punto de vista experimental es un hecho innegable que T, depende de las propiedades del medio acuoso en que el ADN esta inmerso, es decir del pH, dela concentración de iónes y de la temperatura en la solución acuosa
[20].
La pregunta inmediata en este contexto es cómo influyen estas variables en el proceso de desnaturalizaci6n delADN.
La respuesta irice&a a esta cuestión continua siendo un problema abierto, sin embargo se han logrado avances significativos al respecto [6,7].
Entre las propiedsdes más importantes asociadas a cadenas largas de
ADN,
podemos señalar la dependencia lineal entre la concentracinn de pares de bases Citocina-Guanina y la temperatu-ra de fusión T,. El entendimiento de este proceso constituye un paso importante para entender los mecanismos primordiales de la biologia molecular, por ejemplo la repiicación, reparación,
mutación y transcripción del
ADN
[16,18]. Por otra parte, la predicción certera de la desnat- iiralización térmica del ADN es de suma importancia para algunas técnicas empleadas en la biología molecular, entre las que sobresalen la r e a x i 6 n de polimeraza en cadena (PCR) [21],st~.iienciación por hibridación [22], blanco ariti&;ro 1231, Southern Bloth [24], y también de
ii1teri.s en el campo de la nanotecnología [25].
Bibliografía
[I] Murphy M. P. and A. J. O ~ e i l l Luke (Editors).What is life? The next fifty years. Speculations on the future of bicilogy. Cambridge University Press (1995).
[2] Schroedinger
E..
What is life?.The physical aspects of the living cell. Camiridge Uni-versity Press (1944).
[3] Timoféeff-Ressovsky N. W., Zirnmer K. G. and Delbrttck M., Nachrichten til? Lr I. Fi,-logiv
der Gesellschaft der Wissenschaften Gottingen 1 (1935).
[4]
Bolivar
ZapataF.
Obra cientifica IV. La genética nioderna: fundamentos y horizontes.Capítulos 2-4. El colegio Nacional, México (2000).[5] Watson J. D. and Crick
F.
H. C.'Molecular structure of nucleic acids. Nature,171
,4356, 737-738. (1953).[6] Blake
R.
D.,
Encyclopedia of Molecular Biology and Molecular Medicine, V01.2, pp.l, VCH, New York, ed. MeyerR.A.,
(1996).[7] Cantor, C.
R.
and cchimmel,P.
R..
Biophysical Chemistry part I11 the behavior of biological macromolecules. W.H.
Freeman and Company, San Francisco CA.(1980).[8] Poland D. and Sheraga H.A.. Theory of helix-coil transition in Biopolymers: Statistical Mechanics theory of order-disorder transition in Biological Macromolecules. Academic Press, New York, (1970).
191 SantaLucia J., Proc.Nat1. Acad. Sci. USA 95, 1460 (1998).
[lo] Warteli R.M. and Benight A.S., Physics Reports 126, 67 (1985).
[li] Gelbart W.M., Bruinsma
R.,
Pincus P.A. y Parsegian V.A. Physics Today, septiembre38 (2000) .
1121 Kerner
R..
in :J.L.
Morán-idpez (Ed.). Current Problems in Condensed Matters:Theory and Experiments, Plenum Press, ;>. 323, ( 1 W X ) .
[13] Sinden, R.. DNA structure and fimction. Academic Press, San Diego (1994).
[14] Poland D. Cooperative E.quili1iriir in Physical Hiochemistry. Oxford University Press, London (1978).
[16] Brown,
T.A.,
Genomes., John Wiley & Sons Inc., New York, (1999).1171 Soyfer V.
N.
and Potaman V.N.,
Triple-Helical Nucleic Acids. Springer-Verlag, New York. inc. (Is%).[is] Lt-hIli.:iger
A.í~.7
Biochemistry.iVort. New York(1970).
[l!)] Saiw;:t'i.
\V.
1'ririciplt.s of Xiic:lt%: . k i ( l s C t r i x t , i m . Springer-Cérlag,Xew
York (198.1).[lo]
http:, ,'nxwibt .iinairi.nLu/sintr.5is:1211 Saiki '1.k.. (klfarid D.EI.. Stoffd
C..
Scliarf C . . EIigiirlii íi.fI.EIor~i G.T.! lliillis i<.l'i. z:,dgrlich
F1,:L.
Scicricr 239. 487 (, 1998).[22]
k'citlor S . l ' . L fhva KP.. Iliiang X.('..k~as- :I.('.. Il~~lriics C.P. arid .-\dams C.i,.. XatlIrc364. i.5.5 .[199:3).
["3] Frcicr S.11. Antisense Rescarch and Applicatioris.
('RC;
press Boca Raton, FL. pp. fj7,Crooke S.T. and Lebleu
R.
eds.1241 Southern E . X . J. 1101. Biol. 98, 503 [1975),
Capítulo
2
Antecedentes Teóricos de
la
desnaturalize ión.
2.1
Introducoión.
La caracterizaci6n cuantitativa de la desnatiiialización ha sido un problema estudiado desde hace unas decadas [I]. Desde el punto de vista teórico el reto de este fenómeno consiste en proponer un modelo capaz de predecir las propiedades que caracterizan la transicidn en términos de las condiciones ñ~ioi6gicas que prevalecen en el medio acuoso, por ejemplo la temperatura, el pH y la fuerza ionica. En este capitulo presentamos los modelos físicos más representativos que se han propuesto en la literatura para describir la desnaturalización térmica del ADN, los cuales han sido clasificados en dos ramas En la primera, presentamos las ideas centrales de la desnaturalización vista desde un enfoque mechico estadístico.
En
la segunda, clasificamos aquellos desarrollos termcdinárnicos que se han propursto en la literatura.2.2
Enfoque
Estadístico.
2.2.1
Modelo
de
Ising.
mecánico estadístico se considera que inicialmente existe una distribución aieatória del estado de spin de los electrones. Conforme un campo magnético externo se introdurn en el sistema, los electrones tienen cada vez más preferencia a alinearse con la direccbjn del campo externo hasta que eventualmente todos estos adoptan un momento dipolar
+1.
Es
en este estado cuando se alcanza la transición ferromagnética[2].
El modelo de Ising se traslada de manera inmediata al problema de la transición helice-hebra delADN
cuando consideramos que cada electron de la red unidimensional representa un par de bases en la secuencia de la cadena deADN
donde los posibles estados asociados a cada electrón[+1
,-I] corresponden a un estado enlazado6
libre' respectivamente entre las bases nitrogenadas de hebras opuestas [3]. Notemos que ahora la temperatura de la solución acuosa induce la tra:i:>ición y juega el papel del campo magnético externo. De acuerdo con la notación matricial propuesta por Kramers y Wannier[2],
el cambio de la energía libre de Gibbs asociada a una de las configuraciones posibles para una secuencia de N bases de nucleótidos en una aproximación a primeros vecinos es:.N
AG(PI,PLZiPLJ...CLN)
=-pG(CLi?PLi+l)
i=l
donde pi
=
fl
define el estado deli
-esirno
par de bases y se cumple la condición cíclicapN+l
=
pl. En el resto de este de esta tesis denotaremos porAG(,U¡+~,~J
el cambio de energía libre de Gibbs entre el estado arbitrario depi+l
ypi
y el estado de referencia en dondeambos pares de bases se encuentran libres, es decir
AG(pi+l,
p,)
=
G(P,+~,
pi)-G(-1, -1).
Por
otra parte, definimos un operador matricial
P
cuya propiedad esencial es la de realizar unaproyección sobre los estados del i-esimo y el
i
+
1 par de bases, de manera tal que ai proyectar sobre estos obtenemos el factor de Boltzmann entre ellos, es decir )I'na de las ventajas de este formalismo consiste rn observar que la función de partición, la cual nos permite obtener las propiedades termodinámicas de: sisttma en estudio, se ob- tiene directamente de la traza del operador matricial
P
[4]. Coli la finalidad de ejemplificar({+I, +l}{+i,
-1)
{-l,+-}
{-I, --I}). En este c m , el operador matriciaiP
es
una matrizcuadrada de
2
z 2 cuya función de partición se escribe como [4]:2 =
T r ( P N )
=q
+
A? (2.1)donde XI y A2 son los eigenvalores de la matriz
P.
En la situación en que el ADN seconstituya por una secuencia muy grande de nucleótilos, la función de partición se aprouma por la 1%' potencia del eigenvalor de mayor magnitud
[1,4].
El conocimiento explícito cAe la función de partición Z nos perm'te calcular la fracción de bases nitrogenadas que se encuentranen un estado enlazado, 0, conocida en la literatura como la helicidad
6
el parámetro de orden. Cuandoun
par de bases nitrogenadas enlazadas ubicadas dentro de un segmento helicoidal deADN
contribuye conun
factor deAG
RT
s = exp(--)
donde AG representa el cambio de energía libre de Gibbs entre el estado libre y el enlazado de las bases complementarias y es el mismo para todos los pares de bases acoplados, la helicidad
se obtiene por
(2.'3)
donde s = exp(-$$) y AG es el cambio de energía libre de Gibbs y constituye
un
parametrotermodinámico ajustable
[l].
Desde el punto de vista experimental, la helicidad 8 y su comportamiento con los cambios en la temperatura de la solución acuosa, es una propiedad medible en el laboratorio, y por lo tanto comparable con el desarrollo teórico correspondirnte (51. Es un hecho innegable que la
natiiraleza de la helicidad 8 esta fuertemente influenciada por las condiciones fisiologicas de
la solución amosa y por la estabilidad termodinámica entre pares de bases vecinos [I]. DP
ésta manera, la operatividad de este esquema teórico se concentra en dos ejes principales. Por
entre nucleótidos enlazados (ver
tabla1 sección
2.3.2
).
Estas dos cuestiones merecen de manera independienteun
análisis detallado.A
juicio del autor, en la literatura existe una ausencia de conectividad entre ambos esquemas. Por una parte se encuentran lm modelos teóricos sujetos a diversos parametros externos, y por otra, se encuentra una cantidad considerable de mediciones experimentales de la termodinámica de duplex de pares de bases [6-
91. La unión entre amboaenfoques constituye una de las principales contribuciones de esta tesis. Los siguientes capitulos están centrados en el análisis y discusión de algunos de los métodos teóricos que, a
ni
juicio, son los más representativos en sugerir un criterio que evalue de manera eficiente la función de partición representada en la ecuación (2.1). De manera genhrica, podemos identificar dos diferencias entre los distintos modelos que presentamos. La primera es!riba en la forma de aproximar la función de partición del sistema, y la segunda, en el grado de descripción del tratamiento expresado mediante parametros intrínsecos del modelo.2.2.2
El modelo que aquí presentamos fue desarrollado de manera independiente por un lado p&
Zimm y Bragg
[lo],
y por otro lado por Gibbs y DiMarzio[ll],
de manera que, sin restartratos
asus
creadores, ambos son igualmente reconocidos. La idea central consiste en proponer que un par de bases nitrogenadas enlazadas, ubicadas dentro de un segmento helicoidal de ADN, contribuye conun
factor deMétodo matricial
de
Gibbs
yDiMarzio.
AG
RT
s = exp(-
-)
en la fiinción de partición. Aqui
AG
es el cambio de energía libre de Gibba entre el estado librey
el enlazado de dos nucleótidos acoplados por enlaces de hidrógeno,R
es la constanteunivrrsal de los gases y
T
es la temperatura termodinámica. De esta manera, la contribución a la función de partición de un segmento helicoidal de j pares de bases es de s3. Además, existe una~ontribución adicional asociada a cada región desnaturalizada en la cadena polimérica. Ebta contribución se representa a través de un factor LT
<
1 (Llamado parametro de cooperatividad). Desde el punco de vista ffsico, el parametro u cuantifica 108 cambios a la energía libre de Gibbsasociados a las interacciones enxre los primeros vecinos de un par de nucleótidm acoplados.
En la literatura, esta interacción se co~oce conlo en
rgia
de empaquetamiento y contribuye de manera fundamental en la descripción teórica de la desnaturalización térmica delADN.
De
esta iiiaiiera. en est? marco de referencia, la función de partici6n asociada a una secuencia conN
pares de bases enlazados y un número
h
de zonas desnaturalizadas se expresa comodonde la sumatoria se extiende sobre todas las configuraciones para las cuales N y
h
soncompatibles, es decir h
I
N.
De
la mecánica estadística fundamental se deduce que el número promedio de pares de bases enlazados correspondiente a esta función de partición esla cual queda completamente definida cuando se conoce la funci6n de partición Z.
Con
lafinalidad de mostrar el método matricial de Zimm-Bragg, primero consideraremos una secuencia
simple de nucleotidos, PolyAPolyT (podría ser cualquier otra secuencia, sin embargo ésta es
una de las más simples). Asumiremos que sa representa la función de partición interna para un par de nucleótidos acoplados Adenina-Timina, la cual se caracteriza por el cambio de energía libre entre $1 estado libre y el enlazado, es decir
A G ~ ~ so = exp(--)
RT
Por convención denotemos a Z$ como la fiinción de partición de una molécula de N seg- nientos asociada a configuraciones que finalizan con un par de bases enlazado y a
Zg
comola fiiricióri de pártición de segmentos que se i<i<~itificari por tener el último par de base en un
estado libre. Empleando esta notación. la fiiiicióri de partición del segmento de N elementos
la siguiente forma
El primer térmiro de estas relaciones cuantifica todas las configuraciones que se caracterizan por tener el úitimo par de nucleotidos en la secuencia en un estado enlazado t l d o que cl par
de bases anterior también se encuentra en estado enlazado. El siguiente térnzi:ia d,i le, ris:.na
relación resulta de considerar todas las posibles configuraciones en donde se tiene un par de nucldtidos en estado enlazado seguido de un par de nucleótidos libres, lo cual da origen al factor usa. La segunda relación de la ecuación (2.6) justifica el hecho de que un par de
nucleótidca libres seguido de un par de nucleótidos libres o enlazados no contribuye a la función de partición, simplemente es la unidad.
Por
otra parte, de acuerdo a Zimm y B r a g la ecuación(2.6)
se
pueden reescribir en forma compacta mediante la siguiente notación matricialdonde
De acuerdo a esta notación la función
d?
parti<,i<jnZN
para moléculas formadas por Nsegmentos es simplemente la suma de las conipoiirntrs del vector
Zg.
Es interesante resaltarque
si
el número de pares de bases es grande, m (leyir N-
00, las técnicas estandares delalgebra matricial demuestran que
1
-in Z ~ = i n
Tr
(A)zzln XN
donde X es el eigenvalor mayor de la matriz
A,
la cual es consistente con la ecuación (2.1). Elcálculo de la función de partición mediante este método matricial constituye la piedra angular
de
la mayoría de los desarrollos te6ricos que se !ian presentado en la literatura [1,4]. Finalmente, a
manera de resumen podemos afirmar que la cuestión fundamental de esta descripción mecánico eztadística es, en general, el cálculo explícko de la función de partición 2, la cual contiene información de la estabilidad termodinámica consecuente de la interacción a primrros vecinos entre los distintos pares de bases a lo largo i e la secuencia genética.
2.2.3
Modelo de
Reiss-McQuarrie-McTagucCohen.Uno de los métodos para evaluar la función de partición 2, expresada en la ecuación (2.7),
consistente con las mediciones experimentales hechas para la desnaturalización del ADN de organismos superiores, fue el propuesto por Reiss, McQuarrie, McTague y &hen [12]. La prin- cipal contribución de ellos fue la de establecer una conexión entre el método matricial de Gibbs y DiMarzio y las observaciones experimentales realizadas por Marmur y Doty en 1962 [13]. El resultado de s u modelo fue consistente con la correlación empírica entre la. temperatura de fusión y la concentración molar de pares de bases Citocina-Guanina a lo largo de cadenas de ADN inmersas en una solución acuosa de 0.01 M de Na+.
Los
experimentos realizados por Marmur y Doty [13] muestran que el ajuste estadístico de esta correlación se representa porT,(K)
= 342.6+
0.41[C+
G] (2.9)donde [C
+
C] es la fracción molar de pares de bases Citocina-Guanina. Es necesario hacer algunos comentarios respecto a este resultado. Primero, la validez de la ecuación anterior se limita a la situación en que vanas replicas identicas de .ADN se encuentran diluidas en una soliición acuosa con 0.01 M de Na+ y pH7.
Sqquido, esta relación lineal es consistente conc1 cixnportamiento de cadenas de ADN formadas p o r lo nienos de
IOB
nucleótidos. Tercera,el a.juste de los resultados experimentales ciertarritmte es lineal en el intervalo de temperaturas
de 85°C a 100°C. Fuera de este régimen de trniprratiiras el ajuste lineal es completamente trrhneo [13] ( U W figura 3.3). Finalmente, los efectos de
la
correlación en la secuencia de ADNEn este contexto, dos preguntas surgen de manera inmediata. Primero, partiendo del mod- elo estadístico descrito, cómo
es
posible reproducir el comportamiento lineal expresado en laecuación (2.9). Segundo, cómo depende la ecuación (2.9) de la concentración de contraiones Na+ presentes en la solución acuosa. La respuesta a la primera cuestión fue encontrada por Reisa et ai 1121. Ellos consideraron una molecula de AI IN con una secuencia deñnida
por
ABABAB....
ABen donde A representa un par de nucleóticos acoplados formado por Adenina-limina y
B
rep resenla un par de nucleótidos acopla<los Citocina-Cuanina. De acuerdo a la ecuacióri (2 i ) , !rifunción de partición de esta molecula se calcula por
_ -
- -
2 = Traza(ABABAB
....
AB)
donde
donde SA y sg son las funciones de partición internas asociadas a un par de base Adenina-Timina y Citocina-Guanina respectivamente, y matematicamente se expresan siguiendo la ecuación
(2.4).
Además, u es el parámetro de nucleación y se asume que es el mismo para ambos paresde bases. Por otra parte, aplicando la ecuación (2.1), la función de partición 2 es la traza de la
matriz resultante del producto de matrices
A
y en el orden especificado en la secuencia. Sin embargo existe una difucultad. Como consecuencia de que en general las matricesÁ
y no coinnutan, la función de partición depende de la wcuencia en particular la cual para cadenas lai gas era desconocida a priori. La idea central de ese trabajo consistió en sugerir que los datos repoi:ados por Marmur y Doty son el resultado de una función de partición resultado de un promedio entre las posibles permutaciones de los pares de bases que conforman la cadena de AI)N. %pongamos que la secuencia ABABAB- - - -
_ -
....
AI?
contiene N, elementosÁ
yNb
unidadesde B.
En
general, el número posible de permutaciones entre estos pares de bases esPor otra parte, escribivndo
B = A + E K
donde
(2.10)
(2.11)
y sustituyendolas en la función de partición 2 para una permutación posible, se demuestra que ésta se puede escribir como una serie de potencias en
cuyos
coeficientes,Tp,
son
la traza delproducto de matrices factorizadas por EP, es decir
Nh
De esta manera el logaritmo de la función de partición es la suma de las trazas de las dis-
tintas contribuciones de las potencias de EP. La cuestión esencial que k i s s et al. plantearon fue calcular la distribución de las trazas, o mejor dicho de los logaritmos de las trazas, as- ciadas a todas estas permutaciones posibles. El resultado fue que la distribución asociada a las trazas no fue una gaussiana sino fue una distribución agudamente centrada alrededor de ckwo valor promedio [12]. iledizamio una distribiicióri de los eigenvalores obtenidos para cada
p i w i i i i ! aci6,n, ellos enconlraron que existe eigerivalorrs <pie son representativos de la mayoria de IRE pwniitaciones. Eligiendo 621 mayor de los eigrnvalores de esta distribución, calculando 2 y
aplicanch la ecuación (2.3) deino.itraron que la helicidad se escribe como
(w
-
O ) [slU(i-
w)
-
u)+
O ]e=--
donde se emplea la siguiente notación
donde
Ti
es la temperatura de fi 3ión asociada al par de basesA
6E ,
ademásdonde x.4 y z~ representan las fracciones de parrs de bases enlazados adenina-timiria y
citocina-guan:iia respectivamente. Finalmente, u) debe de satisfacer la siguiente condición
w - u
W 2
L-
-
W l l - , W ~ l -con:
Los resultados numéricos de la temperatura de fusi6n obtenidos a partir de la heiicidad, ecuación
(2.12),
resultaron consistentes con el ajuste hecho por Marmur y Doty, ecuaci6n(2.9) [12].
Esta congruencia se obtuvo imponiendo que u = en la ecuación (2.12), y que los cambios entrópicos es una invanante en cada par de nucldtidos acoplados, es decir
A H 4
-AH8
T4
I."~~ -
hipótesis que fue propuesta en bas; i los ríwiltados (IC los experimentos de calorimet,ría real-
izados hasta ese momento [14].
Así, concluinios que el métod,> proiniestú por I2eiss vt al. para evaluar la función de partición
de cadenas grandes es consisten1 e con lo:. rrsiiltulos obtenidos por Marmur y Doty. Sin embar-
go, es necesario mencionar alpmoos as-mtw de i r i i d s . Primero, la idea de elegir una secuencia promedio, obtenida de entre Loc.as la:, pmibles perniiitaciones entre los pares de bases, merece
22
~ ;* ..
un análisis exhaustivo de la naturaleza de las correlaciones asociadas a esta secuencia promedio.
Recientemente, el estudio de las correlaciones en las secuencias de organismos eucariotes han mostrado que los intrones, regiones que no codifican, carecen completamente de correlación a diferencia de los exones, regiones que codifican, que pres?ntai! una correlación de largo alcance
[lij]. Sin tmbargo, esta afirmación aún es cuestionable en la Itteratura [16].. A juicio del autor,
la consistencia de la metodología empleada por Reiss et al es el resultado de una abnmadora cantidad de secuencias de intrones que se encuentran sin comdación en los orgmiwrs? i m r x i - . otes, sin embargo este es un tema aún por explorar. Segundo, en el artículo de h1ar:our y Coty
[13] no se dan suficientes detalles sobre la funcioiiali(1ad qiic presenta T,,, y [C+C] a diferentes
concentraciones de iónes inmersos en 1;. .olución. por lo que queda en tela de jui,rio
la
linealidadde la ecuación (2.9). De hecho en la literatura se piiede encontrar que a menudo hay autores que crean condiciones fisiológicas en donde la concentración de ambos pares de bases presen- tan contribuciones importantes en la transición estructural [17,18]. Finalmente, es importante mencionar que los parámetros ajustables en este modelo son las entalpías y entropias de ambos pares de bases, las temperaturas de fusión, y por último, el parametro de nucleación o. En la siguente sección mostraremos un modelo más estructurado sobre el fenomeno de transición helice-hebra en el ADN.
2.2.4
Modelo de Poland
El modelo de Poland consiste en un refinamiento al modelo de king
[l,
31, en donde la heiicidad se escribe como el producto de dos contribuciones asociadas a grados de libertad independientesdonde
Oint
es la fracción promedio de pan's (IC, Iiaws <pie re encuentran enlazados y Oat es lafracción de hebras que al menos tiene un par de basrs CII cl estado enlazado. ,,,O esta relacionado
UIII
PI
tqirilibrio de la disociación entre diiplrx y iiiia siri,plc c.idrna de ADN, Cz+
2C1. Este proceso de disociación generalmente se ignora ritari(1o si' a na:izF* la desnaturalización de cadenasde
ADX
largas. Sin embargo, la inclusión del paso <it,(ii
io<.ia<;ióii es esencial para describir lapartición interna &,t.
A
cierta temperatura, la cadena de AdN tiene un número determinadode configuraciones asociadas al número de pares de bases enlazados. Si c'enotamos p o r
k
a la configuración qiie presenta Nk pares de base enlazados. entonces 1 t fraocih promedio de paresde bases enlazados se deterniina por
tlomle Z, rs la liini:ióri de partición de la
k
- isirrin <:orifi~iración 1.a fiinción de parii&jniriterria
Zk
se eipec:ifica mediante tres paránittros adic:iorialcs. El primero es el paráriietro tieestabilitlaii s i (iridexado para distirigiiir l o s
pd>'
. .viri?s tlt, hasps enlazados), el ciial representala constante de ecpulibrio asignada a formar el i- , . ' N O par de bases enlazados. De esta manera,
s1 contribuye una vez en
Zk
para cada i - ésirrio par <le riricleótidos enlazado que aparezca enla configuración
k.
Esta constante de eqiulibrio se deterniina en base a dos contribuciones. Laprimera contiene los cambios
en
la energi'a libre de Gibhs,ACi,
correspondiente a los enlacesde hidrógeno del
i
- esirno par de nucleótidos complementarios. La segiuida contribiieiónrepresenta los cambios en energía libre debida a la fuerza de empaqiietaniiento entre prinieros vecinos de pares de niicleótidos,
es
decirAC,
+
AGg-
I,i1
RT
si = exp[-E1
segiindo paránietro. es el llamado de niieleación oi.. l.i, el cual cuantifica el grado der»operatividad qiie caracteriza la transición estructural [3].
A
diferencia de la sección anterior,r s t v partíriic~tro esta indexado debido a q i i ~ las irittwmhes entre vecinos de una srciieiicia
lii~tt~iii~:i.riea son distintas dependiendo del i - 1 y
i
- í s i n i o par de bases. I 3 término 0 ~ - . 1 , ~< T vritritiitj-e ii fa fitricii>ri de partición cada vez r l i i c apirw<> iiri lazo iiit,errio
fomiado
por pares de i ) ; t . i , q libres. ('iiarititativarrirrite esta coiitril~ii(~i~'~ii rrprcscnta porSottwios que la exprtsión anterior para ( T ~ I . j i:iirrt.sponde a Zainhios de la enrrgía lihrr ,correspondiente a las fuerzas de eriir>ailiietaiiiierito, es decir la fuerTa tie interacción
Figura 2-1: Representación esquemática de las interacciones entre dos pares ae base3 de nu-
cleótidos. Las iíneas verticales representan los enlaces fosfatos.
Los
enlaces de hidró:rno srrepresentan por
(=).
Las líneas cruzadcm representan las interacciones que dan origcn al e f k t i s de empaquetamiento del ADN.cruzada entre un duplex de DNA (ver
Figura
2.1).Por otra parte, los lazos internos formados por m pares de bases libra contribuyen de
manera importante en
la
desnaturalización de las cadenas de ADN (ver figura 1.1). Con lafinalidad de tomar en cuenta estas contribuciones, se define a
f(m)
comola
probabilidad deque dos hebras complementarias unidaa en un punto se encuentren en un arreglo espacial tal que después de m pares de beses libres permitan la formación de un par de
bases
enlazados.La dependencia matemática de este parámetro entrópico ha sido abordada con la teoría de los caminantes aleatorios y con aigunaa formuiaciones empíricas. Sin embargo, la cuestión principai
de encontrar una funcionaiidad váiida para m grandes y pequeñas aun permanece inconclusa
en este campo [3,19].
Para ñnaliiar esta sección, es importante señalar que este esquema teórico permite calcular
6,,t de una manera exacta en función de los tres parámetros descritos anteriormente, es decir,
cr,,s* y
f(m).
El tercero de estos puede estimarse, con cierto grado de aproximación, de lateoría de los caminantes aleatorios [19]. Sin embargo, los dos primeros dependen fuertemente dr las mediciones experimentales asociadas a la estabilidad termodinámica de las diez posibles combinaciones por pares entre los pares de bases Adenina-Timina y Citocina-Guanina (Tabla
1 pag.31). Hasta 1995 esta información no estaba disponible debido a las limitaciones de las tknicas experimentales. Por esa razón las entalpías y entropfas empleadas hasta ese entonces eran promedios obtenidos de las mediciones hasta ese momento disponibles [ZO].
otros casm los resultados teóricos difieren completamente de las mediciones experimentales,
por ejemplo revisar [20,21]. En resumen, a pesar del formalismo establecido en la literatura, presentado de manera concreta en las m i o n e s anteriores de esta teak, aún permanecen
muchas
cuestiones abiertas por resolver, entre las que sobresalen la mrrelación que existente entre las propiedades intrínsecas de la cadena
(
la fraoción de pares de bases y s u correspondiente secuencia genética) y la iníluencia que ejercen las propiedades externas del medio (concentrwióii de Nat y pH en la solución acuosa) sobre la temperatura de fusiónT
,
.
Es importante resaltar que los modelos hasta ahora revisados son estrictamente rne!~um5pi,. cos, es decir no consideran la naturaleza microscópica del ADN, sino m á s bien emplean la
informaci6n termodinámica obtenida experimentalmente para las distintas combinaciones entre pares de nucleótidos vecinos. Desde hace menos de un lustro, en la literatura se han presentado
nuevos enfoques para estudiar este fenómeno. Entre estos resaltamos principalmente cuatro. En el primero, agnipanios las varias extension- al modelo de Poland [1,22,23] las cuales dekiiben la desnaturalización térmica del
ADN
tomando en cuentaaspectos
más finos de la molécula del ADN, por ejemplo las interacciones de volumen excluido entre las hebras de polinucleóti- dos y las p r o p i d e s mecánicas como la rigidez y la torsión de la cadena polimérics [24,25].El segundo enfoque incluye el formalismo de la termodinámica propia de la desnaturalización mecánica del ADN [26
-
281. Esta, es una línea de investigación reciente la cual es de inter&actual para entender las propiedades estmcturales de la biomolécda [29].
Por otra parte, en la literatura se han propuesto algunos formalismos que abordan la desnat- uralización del ADN d@e un enfoque microscópico,esta manera de abordar el problema con- stituye la etiqueta que caracteriza el tercer grupo. En este enfoque las descripciones emplean un formalismo dinámico, es decir proponen un Hamiltonian0 asociado a la cadena polimérica el cual contiene los potenciales de interacción entre las bases enlazadas de las cadenas comple- mentarias [30 - 321. A partir de este enfcque se han obtenido logros importantes en explicar y
reproducir la desnaturalizwió~! para algunas secuencias de ADN, por ejemplo mencionamos s u consistencia con el comportamiento experimental de cadenas de oligonucleótidos heterogéneas de
ADN
[21,23]. Sin embargo, es importante resaltar que aún cuando estos modelos han rep> ducido algunos residtados experimentales, la dificultad principal con que se enfrentan estriba en la forma de modelar el Randtoniano del s i s t e k , muy particularmente la energía potenciali
de interacCión entre los nucleotidos.
Finalmente, el cuarto enfoque lo constituyen los modelos
baeados
en los estudios termod- inámicos h d o s para predwir la temperatura de fusión de oligonucle6tidos [6 -- 9,331. Esteenfcque es completaraentr fenomenol6gico y lo trataremos en detalle en la siguiente sección
2.3
Enfoque terrnodinhico.
2.3.1
Termostática fundamental en la desnaturahacibn del
ADN.
Desde el punto de vista fenomenológico,
la
desnaturalización térmica del ADN puede carac- teriierse con la ayuda del potencial termodinámico asociado al prweso, en este caso la energíalibre de Gibbs [34,35]. En consistencia con la segunda ley de la termodinámica, la energía libre de Gibbs nos proporciona
un
criterio para que la desnaturalización se logre en funciónde las propiedades externas del medio, muy particularmente la temperatura. De acu6rdo a la termostática, la energia libre de Gibbs
(AG)
se especifica enbase
a
los cambios entálpicosAH
y entrópicos
AS
del sistema de acuerdo a la siguienterelación
AG= AH-T AS
(2.15)donde
T
es la temperatura termodinámica.En
el caso de la desnaturalización del ADN, la temperatura de fusiónT,
corresponde a la temperatura en que los cambios entr6picoese
equi-libran con los cambios entálpicos de forma tal que
AG
=O
[35]. Por otraparte,
de acuerdo a la termodinámica fundamental, los cambios de entalpía y entmpía en general dependen del calor especificoACp
asociado al procesoAH
=AH"+ACp(T-TO)
AS =
AS"+ACp
In(3
-
*
donde
To,
AH"
yASD
representan la temperatura de fusión, los cambios entálpicos y entrópicosen condiciones estandard respect:vamente [26,27]. De esta forma, el enfoque termodinámico es-
Sin embargo, es necesario resaltar algunos
aspectos
finos
de este enfoque. Primero, la ecuación(2
15) es estrictamente váiida cuando existe equiiibrio termodinámico entre la cadena poliméri-I
ca y la solución acuosa en la cual esta inmen a, condición que algunas veces se viola sobre todo
a bajas concentraciones de iones en la soluciá 1 aci ma [36]. Segundo, esta relación es independi-
ente de la correlaci6n en la secuencia genética del ADN en estudio. Experimentalmente ee ha verificado que cuando se estudian cadenas poliméricas cuyo perffl de desnaturalización corre- sponde a un sistema de dos niveles, es decir un sisíema que solo tiene la posibilidad de e s t a en un estado completamente enlazado 6 en otro completamente desnaturalizado, la temperatura
de fusión T, se determina empleando la ecuaci6n (2.15)
AH
AS
Tm =
-
(2.16)Esta consistencia resulta ser la correspondiente a
AD”;
con secuencias simpies y repetitivas[7
-
91. Cuando la secuencia en el ADNes
heterogénea y correlacionada, los efectos en laconcentraci6n de las hebras.diluidas en la solución acuasa [C] empiezan a contribuir de manera considerable en la termodinámica de Ia transición. En este cam, la relación empfrica aceptada comunmente para describir
la
desnaturalizaci6n de oligonucledtidoses
A H
R
AS AS
T,(K)
=
-
+
-
In[q
donde
R
es la constante universal de los gases. Una particularidad de estos sistemases
queel perfil de desnaturalización no se modela como un sistema de dos niveles, sino más bien existen
algunos estados intermedias entre el estado completamente acoplado y el desnaturalizado. Estos estados intermedios corresponden a desnaturalizaciones locales a lo largo de
la
cadena polimérica 11,371 que en principio interkionan con otra? cadr nas de ADN cercanas a esta, contribuyendo así a la entropía de la desnaturalización.Finalmente, deseo resaltar que la mayoría de 108 trabajos experimentales que miden las propiedades termodinámicas durante la dcsnatiiraiizsci6n de una cadena de ADN asumen que
ACP
= O. La consecuencia inmediata di: estr srip<..sici6n se refleja en la invariancia de los camb.ios entálpicos y entrópicos ante caribios de tt:mperatura[G
-
81. Esta supocisión ha permitido obtener datos experimentales que predicen la T, para varios oligonuclótidos [6-
81, :-sin embargo, su validez e8 aún un aspecto cuestionable en la literatura
[XI.
A
juicio del autor, lavalidez de esta relación no es general, sino m&s bien depende de
las
propiedades que caracterizanla solución acuosa, por ejemplo el pH y las ccncentraciones de sales. La manera explicita de
esta dependencia es aún una cuestih por reso’ver.
En la siguiente sección presentaremos las ideas fundamentales que prevalecen alrededor de los estudios sobre la desnaturalización de oligonucli ótidos, tema que ha sido uno de las linea3 de investigación de mayor relevancia en los últimos 850s debido a sus implicaciones tecnologicas
[5,% 391.
2.3.2
Termodinámica
d?
Oligómeros.
Los primeros trabajos que abordaron el problema de
la
desnaturalización desde un enfoquetermodinámico tuvieron como fundamento la idea central de que
para
cadenascortas
la
ter- modinámica del sistema se especifica a partir de la termodinámica local, que a primeraaprox-imaci6n se determina principalmente por las interacciones a primeros vecinos
[6
-
91. En esteesquema la energía libre de Gibbs de una secuencia particular se determina por la suma de tres
contribuciones
AG
=
AG;-i+i
+
AG8im
+
A G I ~ ~
(2.17)El primer término representa la suma de todas las posibles interacciones entre el
i
-
ésimo
par de bases y su vecino próximo. El siguiente t é d o es una contribuci6n adicional a
la
energía de Gibbs debido a la simetría en la secuencia. Finalmente, cuantifica los efectos de borde en la secuencia, es decir los extremos de la cadena polimérica.De
manera análoga, la entalpía y la entropía se escriben como,(2.18)
conocemos
AHi,i+i, AHinic, AHInic? ASi,i+i, AS&m Y
AS~nie
p o d e m estimar la Tm eu basea la ecuación (2.16). Notemos que este enfoque tiene la peculiaridad de considerar la secuencia de la cadena polimérica en estudio.
Por otra parte, en base a una serie de experimentos ingeniosos hechos con cadenas de oligonucleótidos Rreslauer et al.[6], inspirados por los trabajos pioneros de Tinoco et al
[40],
re- portaron los parámetros termodinamicos asociados a las diez posibles intera.cciones entre pares de nucleótidos en una aproximación de primeros vecinos [6]. Estos experimentos consixt' ,ieron en construir sintéticamente cadenas de oligonuclétidos con secuencias conocidas de oligonu- cle6tidos [ver referencia 6 para detalles]. Posterior al trabajo de Breslauer, se han reportado mediciones más precisas de estos parámetros que predkcn la T, para una cantidad considerable de oligómeros sujetos a distintas condiciones [6-9,411.
En esta tesis hemos adoptado la versión llamada unificada, la cual asocia los parámetros termodinámicos reportados en la Tabla 1.Tabla 1. Psrámetros de estabilidad termodinámica
a 37°C.
AH"
ASo
kd/mol cal/K.mol duplez
AAITT -7.9 -22.2
ATIT.4
-7.2 -20.4TAIAT
-7.2 -21.3E
CAIGT
-8.5 -22.7CTIGA
-7.8 -21.0GAICT
-8.2 -22.2GTICA
-8.4 -22.4CG/GC -10.6 -27.2
GCICG -9.8 -24.4
GGICC
-8.0 -19.9AH"
ASo
kcallmol callK.mo1 Termino de iniciación G o C 0.1 -2.8 Termino de iniciación
A
oT
2.3 4.1Corrección de simetría 0 -1.4
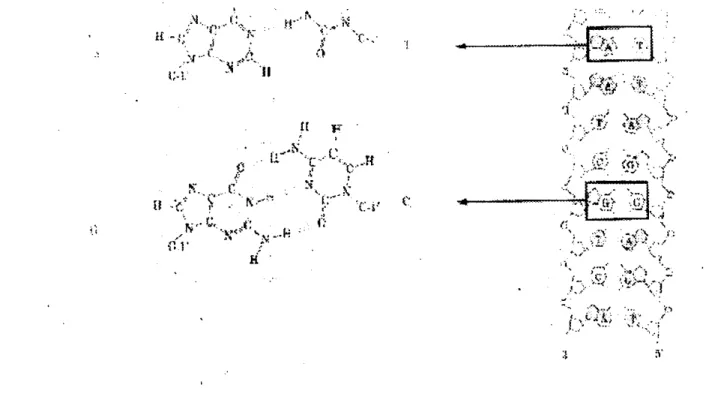
A manera de ejemplo, consideremos la secuencia de ADN mostrada en la figura 2.2
5'
C-G-T-T-G-A
3'* * * t .
3 '
G-C-A-A-C-T
5.
Figura 2-2: CBlculo de las energía libre de Gibbs empleando los parámetros terrndn8micos reportados en la Tabla 1.
mediante la siguiente expresión
I
A@,(predicha)
=
ARi7(CG/GC)+
AG7(GT/CA)+
+ASZ!7(TT/AA)
+
A@,(TG/AC)+
+A@,(GA/cT)+
ARi7(inic)(2.19)
donde
R
piiede ser la entalpía o la entropía a condiciones estandar. La forma explícita de ambosparámetros se logra al sustituir la información contenida en la tabla 1 en la ecuación (2.19).
Además, amifiándonos de la ecuación (2.15), es posible mwtrar que la energfa libre de Gibbs es AG%&(predicha) = -5.35 kcal/mol, la cual no difiere en forma significativa de la cantidad observada experimentalmente Aq7(obs.) = -5.20 kcal/mol.
Esta térmodinhica de oligómeros ha resultado ser una técnica sencilla y útil para el es-
tudio de la desnaturalización, no unicamente de oligonucleótidos heterogéneoe, sino tambien, como defendemos en esta tesis, en el estudio de cadenas largas con secuencia heterogenea y aleatóna
[42].
Recientemente el empleo de estas ideas se ha extendido a situaciones en donde las hebras complementarias que forman el oligómero presentan un defecto en el acoplamiento de los pares de bases[41].
Sin embargo, esta descripción empieza a fallar cuando se analiza el comportamiento en cadenas formadas por un número mayor de 20 bp, situación enla
que lasinteracciones entre las hebras de ADN producen un incremento en los cambios entrópicos de la transición. En este caso los métodos estadísticos son los m& más adecuados para predecir la
Tm
[81.3ibliografía
[ i ] I'oland, D. and Scheraga,
H.
Theory of helix-coil transition in Bioplymer. Academic Press,Yew o r k (1970).
121
Garcia-Colin S., Termodinámica Estadistica.UAh1-Iztapalapa, MéxicoD.F.
(1995). 131 WartellR.M.
and Benight A.S.. Physics Reports 126, 67 (1985).[4]
C a t o r , C. R. and Schimrnel,P.
K.. Biophysical Chemistry part111:
the behavior ofbiological macromolecules. W. H. Freeman and Company, San Francisco CA.(1980).
[5]
BlakeR.
D., Encyclopedia of Molecular Biology and Molecular Medicine, V01.2, pp.l,VCH, New York, ed. Meyer
R.A.,
(1996).
[6]
BreslauerK.
J., Frank R.: BlockerH.
y MarkyL.
A,, Proc. Natl. A d . Sci., 833746
(1986).
[7]
SantaLucia J., Proc.Nat1.Awl.
Sci. USA 95,1460 (1998).
[SI
Allawi H.T. y SadaLucia J., P.:.oc. Natl. Acad. s i . ,57
26 (1998).
[9]
SantaLucia J., Allawi H.T. and Seneviratne P. A., Proc. Natl. Acad. Ski., :35 7.1(L996).
[lo]
Zimm B.H.
and BraggJ
R, J. Chem. Phys. 31526 (1959).
[ll]
Gibbs, J.H.
and Dimarzio E.A.,
J. Chem. Phys. 30271 (1959).
[12]
Reiss €K, McQuarrie D.A,,
Mctague J. P. and CohenE.
R., J. Chem. Phys.44
638
(1966).
[13]
MarmurJ.,
Doty P.,J.
Mol. Biol., 5109
(1962).
[14]
Ross P.D., Scniggs R.L., Biopolymers3
491 (1965).
[15]
Bunde A. Havlii S.(Eds.),
fiactala in Science. Springer-Verlang, first edition(1994).
[16]
Tavaré S., Giddings. Mathematical methods for DNA sequences,,CRC
Press, BocaRaton
(1989).
[17]
http://www.ibt.unam.mx/sintesis/[is]
RudnickJ.
and BruinsmaR.,
http://xxx.lanl.gob/: arXv: cond-mat/O105102(2001).
[I91
Weiss G., Aspects and Applications of the random walk. North Holland, New York(1994).
[20]
Tachibana H., Uen-Nishio S., Gotoh O. and Wada A,, J.Biochem. 92,623
(1982f.
121)
CuleD.
and HwaT.,
Phys. Rev. Lett. 792375 (1997).
[22]
GareiT.,
Monthus C. and OrlandH.,
http://xxx.lanl.gob/:cond-mat/0101058
(2001).
[23]
Campa A. and Giansanti A,: Phys. Rev. E.,58 3
(1998).
[24]
Theodorakopouloe N., Dauxois. Peyrard &l.,
Phys. Rev. Lett.85,
6 (2000).
[25]
Gelbart W.M., Bruinsma R , , I'incus P.A. y Parsegian V.A. Physics Today, septiembre(2000)