Instituto Tecnológico y de Estudios Superiores de Monterrey
Campus Monterrey
Monterrey, Nuevo León a
Lic. Arturo Azuara Flores:
Director de Asesoría Legal del Sistema
Por medio de la presente hago constar que soy autor y titular de la obra titulada"
_ ", en los sucesivo LA OBRA, en virtud de lo cual autorizo a el Instituto Tecnológico y de Estudios Superiores de Monterrey (EL INSTITUTO) para que efectúe la divulgación, publicación, comunicación pública, distribución y reproducción, así como la digitalización de la misma, con fines académicos o propios al objeto de EL INSTITUTO.
El Instituto se compromete a respetar en todo momento mi autoría y a otorgarme el crédito correspondiente en todas las actividades mencionadas anteriormente de la obra.
De la misma manera, desligo de toda responsabilidad a EL INSTITUTO por cualquier violación a los derechos de autor y propiedad intelectual que cometa el suscrito frente a terceros.
Caracterización de la Biología de la Cotorra Serrana Oriental y
Desarrollo de Técnicas para la Detección de Nidos
Exitosos-Edición Única
Title Caracterización de la Biología de la Cotorra Serrana Oriental y Desarrollo de Técnicas para la Detección de Nidos Exitosos-Edición Única
Authors Simón Octavio Valdez Juárez Affiliation ITESM-Campus Monterrey Issue Date 2006-11-01
Item type Tesis
Rights Open Access
Downloaded 19-Jan-2017 10:26:39
INSTITUTO TECNOLÓGICO Y DE ESTUDIOS SUPERIORES DE MONTERREY CAMPUS MONTERREY
DIVISIÓN DE INGENIERÍA Y ARQUITECTURA PROGRAMA DE GRADUADOS EN INGENIERÍA
Caracterización de la biología reproductiva de la cotorra serrana oriental y desarrollo de técnicas para la detección de nidos exitosos.
TESIS
PRESENTADA COMO REQUISITO PARACIAL PARA OBTENER EL GRADO ACADEMICO DE:
MAESTRO EN CIENCAS
ESPECIALIDAD EN SISTEMAS AMBIENTALES
POR:
SIMÓN OCTAVIO VALDEZ JUÁREZ
INSTITUTO TECNOLÓGICO Y DE ESTUDIOS SUPERIORES DE MONTERREY
CAMPUS MONTERREY
DIVISIÓN DE INGENIERÍA Y ARQUITECTURA PROGRAMA DE GRADUADOS EN INGENIERÍA
Los miembros del comité de tesis recomendamos que el presente proyecto de tesis presentado por el Ing. Simón Octavio Valdez Juárez sea aceptado como requisito parcial para obtener el grado académico de:
Maestro en Ciencias en Sistemas Ambientales
Especialidad en Manejo Sostenible de Recursos Naturales
Comité de Tesis:
_________________________ Dr. Ernest C. Enkerlin Hoeflich
Asesor
________________________ _____________________ Dr. Diego Fabián Lozano García M.C. Sonia Gabriela Ortiz Maciel
Sinodal Sinodal
Aprobado:
_______________________ Dr. Francisco Ángel Bello
Resumen
Se analizó el comportamiento de 92 parejas exitosas de la cotorra serrana oriental (Rhynchopsitta terrisi) en
las dos principales colonias reproductivas de 1997 al 2005. El periodo de anidación fue dividido en 3 etapas (incubación, desarrollo, emplumado). Se encontró que la fecha de inicio de incubación varia anualmente en un rango de hasta 2 semanas. Se concluyó que las frecuencias de entradas al nido constituyen un indicador confiable del esfuerzo parental en las parejas exitosas, el cual varía en las distintas etapas de anidación con dos picos de actividad, el primero durante la incubación (2.37 entradas/día) y el mayor durante el emplumado (3.00 entradas/día) una semana antes de que vuelen los pollos. De la misma manera los patrones de actividad diaria por hora varían de acuerdo a la etapa de anidación. Se comprobó que en promedio el 73% de las parejas presentes en las colonias de anidación no son reproductivas y se estimo que el 86.17% de los nidos iniciados tienen éxito en la producción de al menos un pollo. Con los resultados se establecieron parámetros y
protocolos que indiquen a futuros investigadores o trabajadores de campo el éxito o fracaso de los nidos en las distintas etapas de anidación.
Abstract
We analyzed the behavior of 92 successful pairs of maroon-fronted parrot (Rhynchopsitta terrisi) on the two
De dic a to ria
Dedicado a mis padres que me apoyaron incondicionalmente en todos mis proyectos, me legaron su moral y valores e inculcaron en mi el gusto por la ciencia y la naturaleza
A Rubén y Alan por haberme enseñado tanto
A mis errores, pues de ellos aprendí más que de mis aciertos
A México
AGRADECIMIENTOS
♦ Mi más profunda gratitud a Sonia Gabriela Ortiz Maciel quien me dio la oportunidad de unirme al Programa de Manejo Sostenible de Ecosistemas, me dio su apoyo y confianza a lo largo de toda la Maestría y por haberme brindado su amistad.
♦ A mi director de tesis, Ernesto C. Enkerlin Hoeflich por su asesoría y consejo que mejoraron enormemente la calidad del presente trabajo.
♦ A mis sinodales, Sonia Gabriela Ortiz Maciel y Fabián Lozano por su tiempo dedicado, aportaciones y comentarios.
♦ A Rene Valdes Peña, con quien pasé mucho tiempo como compañero de campo y quien siempre se mantuvo en mejor disposición y con gran paciencia para los contratiempos encontrados en un proyecto anterior de tesis.
♦ Centro de Calidad Ambiental (CCA) del ITESM, por el apoyo y la beca otorgadas en mi calidad de asistente de investigación, así como por permitirme hacer uso de la información recabada por el Programa de Manejo Sostenible de Ecosistemas a lo largo de 9 temporadas de monitoreo de la cotorra.
♦ Voluntarios del programa de manejo sustentable de Ecosistemas, que recabaron mucha de la información utilizada por el presente proyecto, en particular a Lucas Foester que fue mi compañero durante el monitoreo intensivo de la temporada 2005.
♦ Museo de las Aves de México, por todas las facilidades otorgadas para realizar el monitoreo de la colonia de anidación El Taray.
♦ CONABIO, Parque Nacional Cumbres de Monterrey y el ITESM, por el apoyo financiero otorgado al proyecto general de las cotorras, lo que permitió cubrir muchos de los costos operativos y de equipo del estudio.
♦ A los compañeros y amigos que estuvieron conmigo a lo largo del estudio de la Maestría. Agradezco particularmente a mis amigos de “la casa de los serios” y a los de “la casa de los ñoños”, con quienes compartí momentos muy gratos.
♦ A los maestros Internacionales de las Preparatorias Tec., particularmente de los campus Santa Catarina y Garza Sada que me brindaron su amistad y con quienes viví muy gratas experiencias.
♦ A mi tía Lucila López de Cabañas y a mi prima Adriana Cabañas por haberme recibido por primera vez en Monterrey brindándome su compañía y afecto durante los periodos más difíciles de mi llegada.
CONTENIDO
Página
RESUMEN...iii
DEDICATORIA…...v
AGRADECIMIENTOS...vi
CAPITULO I INTRODUCCION Antecedentes………...2
Psitácidos en México...2
Género Rhynchopsitta ...3
La cotorra serrana oriental ...3
Distribución geográfica...3
Hábitos alimenticios ...4
Hábitos reproductivos...4
Amenazas para la conservación de la especie ...5
Importancia de conservación de la especie ...5
Importancia del estudio de la biología reproductiva de la cotorra... 6
Objetivo general………....6
Estructura de la tesis………. 7
CAPITULO II CARACTERIZACIÓN DE LOS NIDOS EXITOSOS DE LA COTORRA Introducción ...8
Antecedentes...8
Objetivo específico del capítulo...8
Área de estudio………...8
Metodología ...9
Caracterización de nidos exitosos…………..………...9
Variación en la fecha de inicio de la temporada reproductiva ...10
Frecuencia de entradas y salidas al nido...11
Variación temporal en las visitas efectuadas por 1 ó 2 de los padres ...11
Resultados ...12
Variabilidad en la fecha de inicio de la temporada reproductiva ...12
Comparación de la frecuencia de entradas y salidas del nido...13
Cambios temporales en la frecuencia de visitas al nido…………,.14
Cambios en los patrones de actividad diarios...17
Discusión de resultados...22
Diferencia en el registro de entradas y salidas…...22
Diferencias encontradas respecto a trabajos anteriores...23
Frecuencia de visitas al nido.,,,,,,,,...24
Variabilidad en la fecha de salida de los volantones...25
Incubación...26
Desarrollo...27
Emplumado...28
Conclusiones...30
CAPITULO III DESARROLLO DE CRITERIOS PARA LA DETECCIÓN DE NIDOS EXITOSOS, NIDOS QUE FRACASAN Y PAREJAS NO REPRODUCTORAS Introducción ...31
Antecedentes ...31
Objetivos específicos del capítulo...32
Metodología...32
Criterio I...32
Criterio II... ..33
Nidos exitosos no detectados……….………....33
Nidos que fracasan...-...34
Cavidades utilizadas por parejas no reproductoras...33
Resultados...35
Criterio I...35
Criterio II...39
Nidos exitosos no detectados………40
Cavidades utilizadas por parejas no reproductoras...42
Discusión de resultados...44
Efectividad del criterio I...44
Efectividad del criterio II...45
Estimación de nidos exitosos no detectados...46
Estimación de nidos que fracasaron………...47
Proporción de parejas no reproductoras,,,,,,,,..,,,,,,,,,,,,,,,,,,,,,,……..,,,. 50
CAPITULO IV.
DISEÑO DE UNA METODOLOGÍA DE MONITOREO
Introducción ...52
Antecedentes ...52
Objetivo específico del capítulo...53
Metodología...54
Fecha de inicio del monitoreo...54
Número de sesiones de observación...54
Hora de inició y duración de las sesiones de observación ...55
Resultados...55
Fecha de inicio del monitoreo...55
Número de sesiones de observación...57
Hora de inició y duración de las sesiones de observación ...58
Discusión de resultados...59
Número de sesiones de observación...59
Porcentaje de entradas registradas en relación a la duración de las sesiones de observación…….,...59
Recomendaciones...60
Conclusiones...61
LITERATURA CITADA ...63
INDICE DE TABLAS
Página TABLA 1. Días de observación por año en las 2 principales colonias de anidación...10 TABLA 2. Fecha de salida de los pollos en el Taray gris durante las temporadas
1997-2005...12 TABLA 3. Frecuencia de entradas en las 3 etapas de la temporada de anidación…...15 TABLA 4. Frecuencia de visitas diarias en los nidos de varias especies de psitácidos...24
TABLA 5.Estimaciones de productividad utilizando la frecuencia de entrada y
Porcentaje de error en las 3 etapas de anidación...38 TABLA 6.Nidos exitosos no detectados en el Taray Rojo...41 TABLA 7. Cavidades con frecuencia de entrada por debajo del IC que cumplieron con el criterio II y estimado de nidos exitosos dentro de esta fracción...41 TABLA 8. Nidos exitosos estimados para el Taray Rojo utilizando los criterios I y II...42 TABLA 9. Porcentaje anual de parejas no reproductoras en las 3 etapas de anidación...43 TABLA 10. Causas de falla total en los nidos de R. pachyrhyncha por año
(Tomada de Monterrubio 2000) ...48 TABLA 11.Estimado anual de nidos que fracasaron en el Taray Rojo...50 TABLA 12.Colonias de anidación de la cotorra de acuerdo a la metodología de
monitoreo utilizada...53 TABLA 13. Número de días de observación y criterio para considerar como exitosa a una cavidad...54 TABLA 14. Nidos exitosos estimados en el Taray Rojo en base a un número limitado de sesiones de observación...57 TABLA 15.Disminución en las entradas registradas de acuerdo a la duración de las
INDICE DE FIGURAS
Página FIGURA 1. Promedio de entradas y salidas registradas por día para los nidos
exitosos(N=92) ...14 FIGURA 2.Promedio de entradas diarias registradas en la temporada de
anidación (N=92) ...…...15
FIGURA 3.Porcentaje de entradas realizadas por 1 ó 2 padres (N=92)... 17 FIGURA 4.Entradas y salidas por hora durante la incubación (N=71) ... 18
FIGURA 5. Hora de ocurrencia de las tres primeras entradas durante la
incubación (N=71) ... 19 FIGURA 6.Entradas y salidas por hora durante el desarrollo (N=84) ... 20
FIGURA 7. Hora de ocurrencia de las tres primeras entradas durante el
desarrollo (N=84) ... 20 FIGURA 8.Entradas y salidas por hora durante el emplumado (N=92) ... 21
FIGURA 9. Hora de ocurrencia de las tres primeras entradas durante el
emplumado (N=92) ... 22 FIGURA 10. Número de visitas efectuadas a los nidos exitosos del risco rojo a lo largo de la temporada 1997 (N=10) (Tomada de Macias-Caballero 1998) ... 23 FIGURA 11. Clasificación de los nidos del Taray Rojo de acuerdo a su frecuencia de entrada durante la incubación (N=106) ...37 FIGURA 12. Clasificación de los nidos del Taray Rojo de acuerdo a su frecuencia de entrada durante el desarrollo (N=153) ...37 FIGURA 13. Clasificación de los nidos del Taray Rojo de acuerdo a su frecuencia de entrada durante el emplumado (N=76) .-...38 FIGURA 14. Total de nidos registrados durante el emplumado en el Taray Rojo
clasificados de acuerdo a su frecuencia de entrada (N=208)………....39
FIGURA 15. Nidos registrados en mas del 30% de las sesiones de observación durante el emplumado en el Taray Rojo clasificados de acuerdo a su
FIGURA 16. Fluctuación temporal en la proporción de parejas reproductivas y no
reproductivas………...……43 FIGURA 17.Diagrama de flujo a utilizar para la aplicación de ambos criterios en la detección de nidos exitosos………...……...……47 FIGURA 18.Días de monitoreo considerando la variación en el inicio de la
Capítulo I
Introducción
La cotorra serrana oriental (Rhynchopsitta terrisi) es habitante de los bosques de
coníferas de la Sierra Madre Oriental localizados entre 2000 y 3500msnm (Collar et al.,
1992). En 1947 fue reconocida por Moore como una especie diferente de su especie hermana, la cotorra serrana occidental Rhynchopsitta pachyrhyncha. Ambas especies
difieren en los patrones de coloración (Hardy 1967), están separadas geográficamente y presentan diferentes hábitos de anidación: la cotorra serrana oriental anida en riscos de piedra caliza, mientras que la cotorra serrana occidental anida en huecos de árboles maduros de diversas especies de pino (Lawson y Lanning 1981).El rango de distribución de la cotorra serrana oriental, en lo sucesivo cotorra, se restringe a bosques de pino en elevaciones superiores a los 2000msnm dentro de la Sierra Madre Oriental, en el Sureste de Coahuila, Sur de Nuevo León y Suroeste de Tamaulipas. Recientemente se ha registrado en la Sierra Gorda, Querétaro durante el invierno (SEMARNAP, 2000)
La SEMARNAT (Anteriormente SEMARNAP, Secretaria de Medio Ambiente y Recursos Naturales y Pesca) incluye a la cotorra dentro de su proyecto para la conservación, manejo y aprovechamiento sustentable de los psitácidos en México. La Norma Oficial Mexicana NOM-059 considera a la cotorra serrana oriental como una especie amenazada, a nivel internacional la cotorra se encuentra listada en el apéndice I del CITES (Convención Internacional sobre el Trafico de Especies en Peligro), (SEMARNAP 2000).
Las principales amenazas antropogénicas a las que se enfrenta la cotorra son los cambios de hábitat, particularmente los incendios forestales y la destrucción de bosques para crear espacios para la agricultura y pastoreo. La tala de bosques representa una amenaza pero en menor escala (Enkerlin-Hoeflich et al. 1997). Una amenaza reciente la constituye el
desarrollo inmobiliario desordenado que genera tanto aumentos en los riesgos de incendio como efectos directos por desmontes. (Enkerlin-Hoeflich, comm. pers. 2004).
El presente estudio tipificará el comportamiento de anidación de las parejas exitosas de la cotorra tomando en cuenta la información acumulada por 10 años de esfuerzo continuo del programa de manejo sostenible de ecosistemas en el Centro de Calidad Ambiental del Instituto Tecnológico de Estudios Superiores de Monterrey.
Antecedentes
Los primeros estudios realizados con Rhynchopsitta terrisi fueron hechos por Lawson y
Lanning en 1981. Ellos reportaron información general de la especie, mencionaron la manera cómo localizaron las cotorras, los conteos que hicieron, observaciones respecto a la anidación colonial en los riscos, la migración, la descripción de los pollos, vocalización y depredadores (Lawson y Lanning, 1981)
De 1994 a la fecha el Centro de Calidad Ambiental del ITESM, como parte del proyecto de investigación “Status distribución y ecología de las Cotorras Serranas (Rhynchopsitta pachyrhyncha y R. terrisi) en el Norte de México ha monitoreado todas las colonias de
anidación conocidas para la cotorra, reuniendo un registro extenso del comportamiento de anidación de la especie y de los polluelos producidos cada año en las 2 principales colonias de anidación, “El Taray y Condominios”. Dicho acervo de información fue utilizado para la elaboración del presente trabajo.
Durante la temporada reproductiva 1997 Macias-Caballero estimó que hubo un total de 30 nidos exitosos en la colonia de anidación El Taray,, cada uno produciendo en promedio 2.0 pollos, por lo que la productividad de la colonia para esa temporada fue de 60 pollos. Durante la temporada de 1998 se encontró una disminución de 30% de nidos exitosos y un 25% en el éxito reproductivo por pareja (Macias-Caballero 1998.)
El uso del paisaje de la cotorra en su rango de distribución reproductivo es altamente dependiente en los bosques de la Sierra Madre Oriental ya que duerme y come principalmente en bosques de pino-oyamel-ayarin (Pinus, Pinus cembroides y Quercus),y
en menor medida bosques de pino piñonero y pino encino (Pinus y Quercus). Todos estos
localizados con exposición norteña. En el mismo estudio se encontró que la especie es muy selectiva, ya que nunca se le encontró en bosques de encino, matorral, pastizal, zona agrícola o suelos desnudos (Ortiz-Maciel 2000).
Psitácidos en México
La Cotorra Serrana Oriental (Rhynchopsitta terrisi) pertenece al orden de los
psitaciformes y a la familia psitácidos, la cual es particularmente carismática para los humanos, siendo ésta la principal causa de lo difundido de la práctica de su captura para ser usadas como mascotas. Este hecho aunado a las presiones ocasionadas por la destrucción y transformación de su hábitat natural ha convertido a la familia psitácidos en una de las más vulnerables y con más alto riesgo de declinación y/o extinción. De las 140 especies de psitácidos neotropicales 42 se encuentran amenazadas o en peligro de extinción (Collar y Juniper 1992). Mundialmente de todas las familias de aves, los psitácidos tienen la mayor proporción de especies amenazadas o en peligro de extinción (Juniper y Parr, 1998).
internacional todas las especies consideradas en peligro de extinción por la NOM-059 también son incluidas en el apéndice I del CITES (SEMARNAT 2000).
Género Rhynchopsitta
Aún cuando los psitácidos se distribuyen principalmente en ambientes tropicales, el género Rhynchopsitta es una de las pocas excepciones, ya que solamente habita en los
bosques templados de pino encino del norte de México. Ambas especies de Rhynchopsitta (R. terrisi y R. pachyrhyncha) se consideran de talla grande, siendo la más pequeña la
occidental. Difieren también en color, y en sus requerimientos de anidación: la Cotorra Serrana Occidental anida en huecos de árboles mientras que la Cotorra Serrana Oriental anida en cavidades de los riscos, sin embargo muestran similitudes en su dieta y en las vocalizaciones (Snyder. 1996) y no presentan dimorfismo sexual.
Cotorra Serrana Oriental (Rhynchopsitta terrisi)
La cotorra serrana oriental presenta una coloración vistosa, con el cuerpo verde oscuro, frente marrón, ojos color ámbar, anillo ocular amarillo y pico negro en adultos y blanquecino en juveniles. Presenta coloración roja en la parte superior de las plumas cobertoras y parte superior de las patas de color rojo oscuro. Mide aproximadamente de 40.5 a 45.5 cm. y su peso promedio es de 460gr. (Howel y Webb, 1995).
La cotorra presenta hábitos de anidación gregarios, anida en los huecos y grietas de los riscos de piedra caliza en la Sierra Madre Oriental, formando colonias que pueden contener desde 1 o 2 parejas hasta más de 100. Las cavidades en las que anida se encuentran entre los 7 y los 200m de altura (Enkerlin-Hoeflich et al. 1997), hasta la fecha se tiene conocimiento de 22 colonias de anidación en los estados de Coahuila y Nuevo León (Enkerlin-Hoeflich et al. 1999).
Distribución geográfica
El rango de distribución de la especie es muy pequeño, debido a la reducción de los bosques de confieras en el Holoceno, razón por la cual se considera una especie relictual (Rea, 1997). Su principal rango actual de distribución está confinado a una pequeña área del noreste de México, que comprende una estrecha sección de la Sierra Madre Oriental de aproximadamente 300 Km. de largo por 60 Km. de ancho (Collar et al., 1992). Dicha área
está conformada por una pequeña parte del sureste de Coahuila, el centro y el sur de Nuevo León, y el suroeste de Tamaulipas. De los 18,000 Km2 contenidos en esta franja sólo de un 20 a un 40% (3,500-7,000 Km.2) tienen las características apropiadas para servir de hábitat
a la cotorra (Lanning y Lawson, 1979) Es muy probable que debido a la destrucción de hábitat esta cifra sea menor en la actualidad. El punto más al sur de su área de anidación es el cerro del Potosí, (Collar et al., 1992) mientras que el sitio más sureño en que se han
(SEMARNAT, 2000). Aunque los registros históricos de avistamientos en Veracruz no necesariamente corresponden a esta especie (Collar et al., 1992).
La especie presenta movimientos estacionales dentro de su rango en respuesta a la disponibilidad de las semillas de pino que constituyen su principal alimento (Juniper y Parr 1998). La reproducción de la especie ocurre en la parte norteña de su rango de distribución (centro de Nuevo León y sureste de Coahuila) durante los meses de Abril a Noviembre; mientras que en los meses de Diciembre a Marzo se encuentra en la parte sureña de su rango (sur de Nuevo León y Suroeste de Tamaulipas) en donde pasa el invierno (Macias-Caballero 1998)
Hábitos alimenticios
La cotorra se alimenta principalmente de semillas en las piñas de varias especies de pinos y en menor grado de bellotas, brotes de coníferas y ocasionalmente de néctar de flores de maguey (Agave macroculmis) (Enkerlin-Hoeflich, et al. 1999). Las especies de pino más
utilizadas son: Pinus ayacahuite, P. strobiformis, P. montezumae, P. greggi, P. cembroides y P. culminicula. (Ortiz-Maciel, 2000). La producción de semillas de pinos y encinos
tiende a ser errática regionalmente, las aves se desplazan grandes distancias en busca de alimento, esto tiene como consecuencia que exista una gran variabilidad en sus patrones de movimiento anuales (Enkerlin-Hoeflich et al. 1999).
Hábitos reproductivos
La especie presenta hábitos de anidación gregarios formando colonias que varían en tamaño desde 1 o 2 parejas hasta más de 100 (Enkerlin-Hoeflich et. al. 1997). Por observación de las colonias de Rhynchopsitta terrisi se sabe que su ciclo de anidación inicia
durante el mes de Junio y termina alrededor de la última semana de Octubre, pudiéndose prolongar hasta mediados de Noviembre (CCA-ITESM Información no publicada). Hasta la fecha las colonias de anidación mas grandes conocidas son el santuario de aves “El Taray” y la colonia de anidación “Condominios”, las cuales seran descritas posteriormente en el presente trabajo.
Debido a que los nidos de la cotorra son inaccesibles para los investigadores, actualmente no existe información sobre los parámetros que requieren de la inspección directa de los nidos de la cotorra en estado silvestre. Dada su cercanía taxonómica con Rhynchopsitta pachyrhyncha se puede utilizar a esta especie como referencia, en dicha especie la postura
Amenazas para la conservación de la especie
La principal amenaza para la cotorra es la fragmentación del hábitat, ocasionada por incendios, actividades agropecuarias y fraccionamientos campestres (SEMARNAT 2000). Una gran cantidad de parches de hábitat ha sufrido incendios y la regeneración del bosque es muy lenta o es sustituida por vegetación característica de disclimax edáficos e hídricos, por lo que muchas áreas previamente boscosas ahora son chaparral de encino con poco valor para la especie (Enkerlin-Hoeflich et al., 1997).
Debido a la disminución de las zonas boscosas en la Sierra Madre Oriental, la cotorra se encuentra en una especie de “archipiélago” formado por pequeñas islas de bosque en buenas condiciones, las cuales se reducen y fragmentan cada vez más. Como consecuencia de su condición relictual, endémica y especialista de los bosques templados del Noreste de México la cotorra es altamente vulnerable a los cambios de paisaje (Ortiz-Maciel 2000). La caza o captura furtiva, auque en menor escala, también constituye una amenaza para la especie, ya que por lo lento de su dinámica poblacional, la comercialización puede tener grandes consecuencias. (SEMARNAT 2000) considera como una amenaza a los coleccionistas y criadores de EUA, Europa y Japón, que pretenden obtener pie de cría .
Importancia de conservación de la especie
La cotorra es una especie indicadora de la salud de los bosques de pino donde habita, ya que se alimenta sólo de las piñas de pinos maduros, disponibles en grandes cantidades sólo en bosques con poca o nula alteración. Debido a esto las acciones de conservación enfocadas hacia la cotorra benefician a una gran diversidad de especies que cohabitan con ella, tales como: el búho moteado (Strix occidentalis), el búho pigmeo (Glaucidium gnoma), el trogón elegante (Trogan elegans) y el oso negro (Ursus americanus eremicus)
(Snyder 1996).
Adicionalmente los ecosistemas forestales de la Sierra Madre Oriental brindan una serie de servicios ambientales de vital importancia para las poblaciones urbanas y rurales de Nuevo León y Coahuila. Los servicios ambientales derivados de los ecosistemas forestales están ligados con la regulación de procesos naturales, tales como la provisión de agua y la recarga de acuíferos, la mejora de la calidad del aire mediante la absorción de dióxido de carbono, el control de la erosión del suelo y como soporte esencial en la mitigación de riesgos naturales. En el 2004 la Comisión Nacional Forestal (CONAFOR) aprobó el pago de $56,400 pesos anuales durante un periodo de 5 años por concepto de servicios ambientales hidrológicos prestados por 188 hectáreas de bosque de pino ubicadas dentro de los límites del santuario de aves el Taray. (CONAFOR 2004)
últimos 20 años (Enkerlin-Hoeflich et. al. 1997); estudios realizados por Ortiz-Maciel
(2000) sugieren que la población se encuentra cercana al límite mínimo de viabilidad de la especie, de alrededor de 1500 individuos. En Diciembre del 2004 el equipo del trabajo del ITESM realizó un conteo de entre 3100 y 3500 cotorras en una zona cercana a Zaragoza N.L. en el 2004 y en el 2006 en la misma área se registró una parvada de más de 3600 individuos, siendo hasta la fecha los conteos más grandes reportados para la especie (CCA-ITESM información no publicada).
Importancia de estudio de la biología reproductiva de Rhynchopsitta terrisi
Dado que la población de la cotorra muestra gran vulnerabilidad por los factores anteriormente mencionados, la información relacionada con su biología de anidación, factores de mortalidad y medidas de productividad pueden ser utilizadas como indicadores de la calidad del hábitat y pueden ayudar a establecer medidas de conservación más efectivas (Enkerlin-Hoeflich et. al. 1997).Uno de los mayores obstáculos para la
conservación de los psitácidos es la escasa información sobre la ecología, biología y densidades poblacionales de sus especies en condiciones silvestres y la inexistencia de una metodología estandarizada para el monitoreo de sus poblaciones (Desenne y Sanz 1995). Como se mencionó con anterioridad la cotorra anida en cavidades prácticamente inaccesibles para los biólogos responsables de monitorear el comportamiento de anidación de las cotorras, por lo que deben de basar sus observaciones en el comportamiento de las parejas, para determinar su éxito o fracaso en el nido. Actualmente la evaluación del comportamiento de una pareja depende en gran medida de los observadores y su experiencia, debido a que no existen lineamientos que permitan identificar de manera concisa cuando una pareja se encuentra en comportamiento de anidación. El presente trabajo pretende servir como una guía para la detección de nidos exitosos.
Objetivo general
Estructura de la tesis
Capítulo II
Caracterización de los nidos exitosos de la cotorra
Introducción
Una de las medidas más comunes para medir la actividad de los nidos exitosos es el monitoreo del esfuerzo parental. El esfuerzo parental se refiere a la cantidad de recursos tales como el tiempo y/o energía que los padres invierten en la producción y el cuidado de su descendencia (Siegel et. al. 1998). El cual puede tomar diversas formas, tales como la
preparación de los nidos, producción de gametos o asistencia a la descendencia (Clutton-Brock 1991). Por lo tanto el esfuerzo parental puede ser medido en varias maneras, siendo una de las más comunes el índice de aprovisionamiento al nido por parte de los padres. El estudio del esfuerzo parental con la cotorra presenta dificultades y limitaciones por el hecho de que no es posible observar el interior de los nidos. Esta limitación tiene como consecuencia que el éxito de un nido sólo se pueda comprobar mediante la salida de un pollo.
El primer capitulo del presente trabajo se enfocará en caracterizar el esfuerzo parental en aquellos nidos en los que se ha registrado la salida de al menos un pollo. Dicha caracterización tiene como finalidad el establecer criterios y parámetros que de ser observados en otros nidos en futuras temporadas, puedan servir para inferir el éxito o fracaso de un nido en particular.
Antecedentes
En las dos últimas décadas la biología reproductiva de los psitácidos neotropicales ha sido objeto de numerosos estudios que surgen ante la necesidad de profundizar más el conocimiento de esta familia severamente afectada por los factores antropogénicos anteriormente mencionados. El presente trabajo representa el segundo esfuerzo encaminado a la comprensión de la biología reproductiva de la cotorra, haciendo uso de la nueva información recabada desde la última publicación en este tema en 1998.
Objetivo específico del capítulo
Caracterizar el esfuerzo parental de las parejas que se reproducen exitosamente.
Área de estudio
Tamaulipas. La Sierra Madre Oriental presenta un declive generalmente abrupto con alturas de hasta 3,600 msnm. Las estribaciones de la Sierra Madre Oriental en el Oeste se nivelan entre los 1,000 y 1,800 msnm., formando parte del altiplano Mexicano en sus porciones llamadas Mesa del Norte en Coahuila (Contreras-Balderas 1995).
El presente estudio se llevó a cabo en las dos principales colonias de anidación conocidas para la cotorra; el Santuario de aves “El Taray”, y el risco de anidación conocido como “Condominios 4”, ambas ubicadas en la parte Oeste de la Sierra Madre Oriental.
Santuario de aves “El Taray”
Operado como santuario de aves a partir de 1995 y en proceso de decreto, esta reserva de 335 ha. es la principal colonia de anidación de la cotorra. Se localiza en el municipio de Arteaga en Coahuila, a 4.5 Km. de Ciénega La Purísima, UTM 0350658E, 2801227N, a una altitud de 2160 msnm. (Enkerlin et.al.1997). Esta colonia consta de 2 riscos,
nombrados de manera informal por su coloración como “Taray gris” y “Taray rojo”. En el periodo de 1997-2005 se han registrado 188 cavidades potencialmente activas como nidos en el Taray Gris y 177 en el Taray Rojo (CCA-ITESM información no publicada)
Colonia de anidación “Condominios 4”
Se le conoce con este nombre de manera informal, a la segunda colonia de anidación más grande conocida para la cotorra. El risco se localiza en el municipio de Santiago N.L. en el ejido Laguna de Sánchez, a 9 km. al suroeste de San Juan Bautista, Coordenadas UTM. 0365043, 2804287N, con una elevación de 2100 msnm. (Enkerlin et.al. 1997). La colonia
se divide en 4 riscos, a cada uno de los cuales se le asignó un número de acuerdo a su ubicación, siendo el 1 el que se localiza en el extremo Este y el 4 en el Oeste. Para este estudio sólo se considero el risco 4, en el cual se han registrado 88 cavidades potencialmente activas como nidos (CCA-ITESM información no publicada).
Metodología
Caracterización de nidos exitosos
El monitoreo de la actividad en las colonias de anidación fue llevado a cabo por personal del programa de manejo sostenible de ecosistemas y por voluntarios capacitados siguiendo la metodología de Macias-Caballero (1998). La información fue recopilada a lo largo de 497 sesiones de observación, que variaban en duración dependiendo de las condiciones climáticas y de la logística, generalmente la hora de inicio fue 08:30-09:00 y la hora fin 17:00-18:00. La información recopilada durante estas sesiones fue utilizada en el presente trabajo tomando en cuenta las siguientes consideraciones:
monitorea la actividad de la colonia diariamente hasta que todas las parejas abandonan para dirigirse a su rango invernal. La frecuencia con la que se realizaron las sesiones de monitoreo no fue constante anualmente por razones de logística. En la tabla 1 se muestra el número de días que fueron llevados a cabo los monitoreos por año y por colonia.
Tabla 1. Días de observación por año en las principales colonias de anidación. Días de visita por año
Nombre del
risco 1997 1998 1999 2000 2001 2002 2003 2004 2005 Total Taray Gris 30 40 17 20 15 59 50 41 47 304
Condominios 4 13 38 3 5 7 24 52 19 32 193
Las observaciones fueron hechas desde la base de los riscos a simple vista o utilizando binoculares para las cavidades más distantes del punto de observación. En el monitoreo se procuró registrar todas las entradas y salidas que fueron observadas en todas las cavidades de la colonia. Sólo se tomó en cuenta la información recopilada en sesiones cuya duración excediera 7:00 horas.
Para determinar un comportamiento de anidación típico diario se utilizó una muestra de 92 nidos pertenecientes a “El Taray Gris” y “Condominio 4”, los cuales se muestran en el anexo 1. Se incluyeron dentro de la muestra todos los nidos en los cuales fue confirmada la salida de al menos un pollo durante las sesiones de monitoreo. No se consideraron dentro de la muestra a aquellos nidos en los cuales se escuchó uno o más pollos en su interior pero no fue posible confirmar fecha de salida.
Debido a que la fecha de vuelo de los pollos presenta una gran variabilidad, (lo que será analizado posteriormente en este trabajo), se le asignó un valor de 0 al día en el que el pollo abandonó el nido y a partir de este día se le asignó un valor en retrospectiva a cada día anterior a la salida del pollo que va desde 0, el día en el que se registró el vuelo del pollo, hasta -93 el cual corresponde a 93 días antes del vuelo del pollo. La temporada de anidación se dividió en 3 etapas (1) incubación: días -93 a -64, (2) desarrollo temprano: días -63 a -32, y (3) desarrollo tardío: días -31 a 0.
Variabilidad en la fecha de inicio de la temporada reproductiva
El comportamiento de variabilidad anual en el inicio de la temporada de anidación ha sido documentado en otros psitácidos tales como Myopsitta monachus, (Navarro et.al.
registrados para determinar si existía una diferencia anual en la fecha de salida de los pollos y para establecer un promedio anual para la fecha de salida de los volantones.
Frecuencia de entradas y salidas al nido
Como primer paso en la caracterización de los nidos exitosos se analizó la diferencia entre el número de entradas y salidas para comprobar la hipótesis de que ambos eventos son registrados con la misma frecuencia, ya que al ser dependientes se esperaría que para ambos eventos se observaran frecuencias muy similares. Se tomaron en cuenta todas las entradas y salidas registradas durante las sesiones de observación descritas con anterioridad, con la siguiente modificación; no se contabilizaron las entradas tipo ST (Aves que se encontraron y permanecieron paradas en la orilla de una cavidad todo el tiempo que se les observó) y N1 (aves que llegan y se quedan paradas en la orilla de la cavidad por varios minutos), criterios establecidos por Macias-Caballero (1998). Debido a que en este tipo de entrada el o los individuos no ingresan a la cavidad sino que permanecen en la orilla, por lo tanto este tipo de entrada no representa un indicador confiable de presencia de pollos en el interior de los nidos.
Una vez que se comprobó la falsedad de la hipótesis de que las entradas y salidas se registran con la misma frecuencia (los resultados se muestran posteriormente en el presente capítulo), se procedió a analizar las fluctuaciones en la frecuencia de entradas que se registran en las diferentes etapas a lo largo de la temporada. Tanto el comportamiento de entradas y salidas diarias como la frecuencia de entradas por etapa fueron analizados utilizando el paquete estadístico MINITAB versión 12.
Para el análisis de frecuencia de entradas y salidas por hora se utilizó la misma metodología de observación, sin embargo la actividad se agrupó en horas después del amanecer. Se utilizó como hora promedio de amanecer para cada semana, la hora del amanecer del Jueves, por ser este el día medio de la semana. Las entradas y salidas que ocurrían dentro de cada hora de las sesiones de observación fueron promediadas en cada etapa de la temporada de anidación con el objetivo de comprender los cambios en los patrones de conducta diarios de las parejas exitosas a lo largo de la temporada.
Variación temporal en las visitas al nido realizadas por 1 ó 2 de los padres
los pollos es suficiente como para que la hembra pueda abandonar el nido sin que exista riesgo de hipotermia. La frecuencia y la duración de los viajes de la hembra en busca de alimento en compañía del macho se incrementan conforme avanza la temporada de anidación. El macho a su vez entra con mayor frecuencia al nido una vez que la hembra lo acompaña. De tal manera que durante las últimas semanas del desarrollo y todo el emplumado ambos padres entran y salen del nido con regularidad.
Tomando en cuenta lo anterior se procedió a analizar la frecuencia de entradas al nido realizadas por 1 ó 2 individuos, en las distintas etapas de la temporada de anidación con el fin de determinar si el comportamiento de la cotorra presenta similitud con patrón de conducta descrito con anterioridad. Debido a que no se puede observar el interior de los nidos de la cotorra, se plantea la hipótesis de que el índice de entradas realizadas por 1 o 2 individuos constituye un indicador de la etapa de anidación en la que se encuentra un determinado nido, esperando encontrar un mayor porcentaje de entradas realizadas por un sólo individuo (presumiblemente el macho) durante la etapa de incubación.
Resultados
Variabilidad en la fecha de inicio de la temporada reproductiva
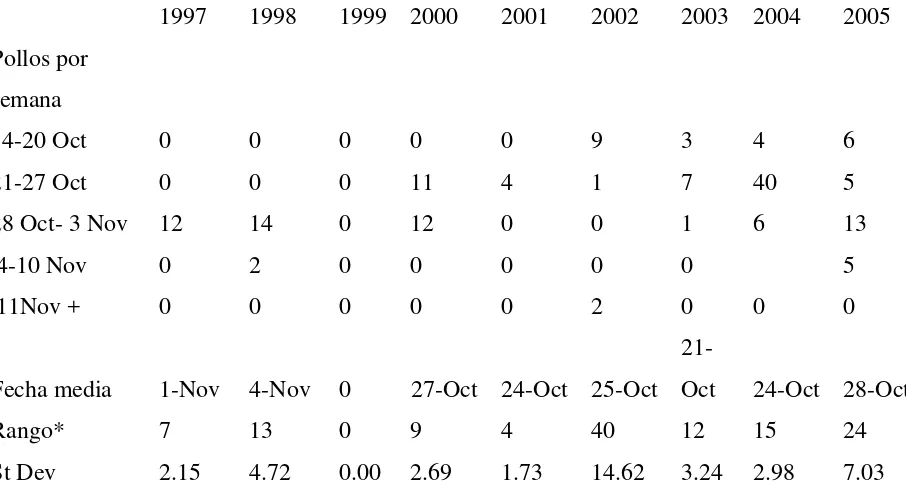
[image:27.612.90.543.457.705.2]De acuerdo con los resultados obtenidos se comprobó la hipótesis de la existencia de variaciones en la fecha de salida anual de los volantones (prueba de Wilcoxon Chi2= 14.067, p =0.000,α=0.05.). La tabla 2 muestra el resumen de lo observado para la variación en la fecha de salida de los volantones.
Tabla 2. Fecha de salida de los pollos en el Taray gris durante las temporadas 1997-2005
1997 1998 1999 2000 2001 2002 2003 2004 2005 Pollos por
semana
14-20 Oct 0 0 0 0 0 9 3 4 6
21-27 Oct 0 0 0 11 4 1 7 40 5
28 Oct- 3 Nov 12 14 0 12 0 0 1 6 13
4-10 Nov 0 2 0 0 0 0 0 5
11Nov + 0 0 0 0 0 2 0 0 0
Fecha media 1-Nov 4-Nov 0 27-Oct 24-Oct 25-Oct
21-Oct 24-21-Oct 28-Oct
Rango* 7 13 0 9 4 40 12 15 24
Se obtuvieron valores diferentes de fecha de salida promedio para cada año, siendo la temporada 2003 la que presenta la fecha más temprana, el 21 de Octubre, mostrando una marcada diferencia de 2 semanas con respecto al 4 de Noviembre, la fecha promedio más tardía, correspondiente a la temporada de 1998.
Las fechas de vuelo más tardías registradas son el 25 y 26 de Noviembre del 2002, cuando 2 volantones abandonaron el nido. El registro más temprano ocurrió durante la temporada 2005 con la salida de un volantón el 15 de Octubre.
El año que presentó una mayor sincronía fue el 2001 donde todos los pollos salieron en un rango de 4 días, mientras que el año en el que se presentó una menor uniformidad fue en el 2002 con un rango de 40 días de diferencia en la salida de los pollos. Cabe mencionar sin embargo que sólo hubo 4 nidos exitosos para la temporada 2001, por lo que lo reducido de la muestra puede influenciar la longitud del rango.
Comparación de frecuencia de entradas y salidas del nido
Se comprobó como falsa la hipótesis de la igualdad entre el número de entradas y salidas, observándose una menor cantidad de registro de salidas durante la incubación y el desarrollo. Durante la incubación se detecta un promedio de 0.333 salidas por cada entrada, (T-Test con p valor de 0.000α=0.05,), en el desarrollo se detectan 0.4469 salidas por cada entrada (T-Test con p valor de 0.000, α=0.05). Es de notarse que en las 2 primeras etapas de anidación se detecta menos de la mitad de salidas con relación a las entradas, por factores que se discutirán más adelante en el presente trabajo. En contraste el número de entradas y salidas registradas no mostró diferencia durante el emplumado, en la cual se registran entre 0.802 y 0.9405 salidas por entrada, lo que no representa una diferencia significativa, (T-Test con p valor 0.291,α=0.05,)
Figura 1.Promedio de entradas y salidas registradas por día para los nidos exitosos, las líneas verticales representan la división entre las 3 etapas de anidación.(N=92).
Cambios temporales en la frecuencia de visitas al nido
Figura 2.Promedio de entradas diarias registradas en la temporada de anidación, las líneas verticales representan la división entre las 3 etapas de anidación.( (N=92)
Se comprobó estadísticamente la existencia de diferencias en el comportamiento de la cotorra durante las diferentes etapas de anidación. (Prueba Kruskal-Wailis,P-valor=0.000, 2 grados de libertad). Durante la incubación se obtuvo un promedio de 2.36 visitas por día, durante el desarrollo, el número de visitas por día disminuyó a 1.98, para después incrementarse paulatinamente durante el emplumado a un promedio de 3.00 visitas por día. Adicionalmente se obtuvieron intervalos de confianza para las medias en las 3 etapas, los resultados se muestran en la tabla 3.
Tabla 3. Frecuencia de entradas en las 3 etapas de la temporada anidación.
INTERVALO DE CONFIANZA
95% PARA LA MEDIA
Etapa PROMEDIO DES.ESTANDAR INFERIOR SUPERIOR
Incubación 92-64 2.367816092 1.757658338 2.10482 2.63082
Desarrollo 63-32 1.948497854 1.473007091 1.7495 2.12981
[image:30.612.82.510.556.662.2]La incubación es la segunda etapa más activa de la temporada de anidación, con un promedio de 2.36 entradas por nido por día, presentando hasta un máximo de 9, lo cual sólo se registró en 3 ocasiones, siendo lo más común que regresaran 1 ó 2 veces después de la primer visita. La actividad alcanzó su punto más bajo durante el desarrollo, con un promedio de 1.95 entradas por día y un registro máximo de 8. El emplumado es la etapa que presenta mayor frecuencia, con un promedio de 3.00 entradas por día y un registro máximo de 14 entradas.
Variación temporal en las visitas efectuadas por 1 ó 2 de los padres
Se llevaron a cabo análisis estadísticos para determinar si existe diferencia en el número de entradas hechas por 1 ó 2 individuos en las diferentes etapas de anidación. Se encontró que efectivamente existe una diferencia, habiendo un mayor registro de entradas realizadas por ambos padres durante las etapas de desarrollo y emplumado (T-Test con p valor de 0.000, α=0.05). Contrario a lo que se esperaba encontrar, no se observó diferencia significativa entre el número de entradas hechas por 1 ó 2 individuos durante la etapa de incubación (T-Test con p valor de 0.160, α=0.05).
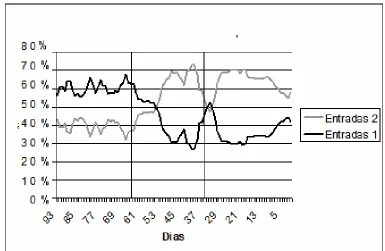
Como se puede apreciar en la figura 3, durante la incubación alrededor del 60% de las visitas al nido son realizadas por un solo individuo. A partir del día 53 y hasta el 46 el porcentaje desciende a valores cercanos al 50% y a partir del día 46 el número de entradas hechas por 2 individuos es mayor con respecto a las realizadas por 1 individuo. Durante el desarrollo y el emplumado aproximadamente el 70% de las entradas son realizadas por ambos padres.
Figura 3. Porcentaje de entradas realizadas por 1 ó 2 padres (N=92), las líneas verticales representan la división entre las 3 etapas de anidación.
Cambios en los patrones de actividad diarios
Antes de proceder al análisis de los resultados obtenidos para los patrones de actividad diaria de la cotorra, cabe mencionar que la información obtenida fue sujeta a los siguientes factores de corrección.
Primeramente debido a que no todas las sesiones de observación abarcan la totalidad de las 12 horas necesarias para la caracterización, se utilizó la regla de 3 para estimar el número de entradas o salidas que hubieran sido registrados en aquellas horas en las que no se llevaron a cabo monitoreos en el 100% de las sesiones.
Incubación
El patrón de actividad diaria durante la incubación muestra una mayor frecuencia de entradas durante la 2ª y 3ª horas que el resto de las etapas de anidación (Figura 4). En la incubación el 25% del total de las visitas al nido ocurren durante la segunda hora, lo cual se debe al arribo de los machos a alimentar por primera vez a las hembras. Sin embargo , al parecer no existe una hora predilecta para la segunda transferencia de alimento, si bien la actividad en la 4ª y 5ª hora es considerable, no llega a formar un pico de actividad. A partir de la 6ª hora la actividad disminuye hasta la hora 11, durante la cual se registran más salidas que entradas. Durante la incubación se observaron 2 comportamientos atípicos, el primero un traslape de la primera y segunda entrada durante la segunda hora (Figura 5) y el segundo un aparente mayor registro de salidas durante las ultimas horas del día, particularmente durante la hora 11, ambos comportamientos serán analizados a detalle en la discusión de resultados.
[image:33.612.104.500.283.536.2]
Hora de ocurrencia de las 3 primeras entradas (incubación)
0 5 10 15 20 25 30
1 2 3 4 5 6 7 8 9 10 11
Hora despues del amanecer
N
o
. d
e en
tr
ad
as
[image:34.612.117.499.70.360.2]Primera Segunda Tercera
Figura 5. Hora de ocurrencia de las tres primeras entradas durante la incubación (N=71) Desarrollo
En el desarrollo registró una mayor cantidad de entradas con respecto a las salidas durante la 1ª hora del día. Presentando un pico de actividad en el transcurso de la 3ª y 4ª horas, en este lapso se registraron el 30% del total de las entradas (Figura 6). Existe un pico de segundas entradas durante la cuarta hora a partir de la cual se observa un descenso constante en la ocurrencia de 2ª y 3ª entradas (Figura 7).
Figura 6. “Entradas y salidas por hora durante el desarrollo (N=84)”
Hora de ocurrencia de las 3 primeras entradas (desarrollo)
0 5 10 15 20 25 30 35
1 2 3 4 5 6 7 8 9 10 11
Horas despues del amanecer
N
o
. d
e en
tr
ad
as
Primera Segunda Tercera
[image:35.612.116.497.379.633.2]Emplumado
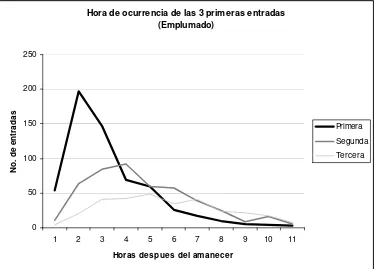
El pico de actividad durante el emplumado coincide con las 2 etapas anteriores respecto a la 2ª y 3ª hora después del amanecer, cuando se registra alrededor del 30% de la actividad diaria. Sin embargo a partir de la 4ª hora, la actividad disminuye de manera más uniforme que en las etapas anteriores (Figura 8).
[image:36.612.115.495.239.472.2]
En cuanto a las horas a las que ocurren la primera y la segunda entrada los resultados coinciden con lo observado en la etapa de desarrollo, la mayoría de las primeras visitas ocurren durante la segunda hora y la segunda visita presenta un pico en la 4ª hora. La 3 visita se distribuye de manera homogénea a partir de la 3ª hora (Figura 9).
Hora de ocurrencia de las 3 primeras entradas (Emplumado)
0 50 100 150 200 250
1 2 3 4 5 6 7 8 9 10 11
Horas despues del amanecer
N
o
. d
e
en
tr
ad
as
[image:37.612.120.495.74.343.2]Primera Segunda Tercera
Figura 9. Hora de ocurrencia de las 3 primeras entradas durante el emplumado (N=92)
Discusión de resultados
Diferencia en el registro de entradas y salidas
Se rechazó la hipótesis de que las entradas y salidas se registran con la misma frecuencia, dicha diferencia puede atribuirse a los siguientes fenómenos:
El primero es que las cotorras al entrar a su nido usualmente lo hacen vocalizando, lo que facilita para el observador seguirlas hasta que aterrizan en la entrada del nido donde perchan brevemente antes de entrar, lo que hace sumamente fácil la detección de las entradas, mientras que en las salidas tienden a ser repentinas y aun cuando en ocasiones sí vocalizan antes de salir del nido, en la mayoría de los casos salen en silencio, por lo que resulta considerablemente más difícil detectar el evento.
considerable de salidas. Este fenómeno no afecta de manera significativa a las entradas ya que al entrar no todas las parejas lo hacen al mismo tiempo.
El comprobar que existe una diferencia entre la entradas y salidas permitió hacer una evaluación de la eficacia del esfuerzo de monitoreo en las distintas etapas, que proveyó de los argumentos necesarios para diseñar una metodología de monitoreo (Capítulo III) basada en un perfil más preciso del comportamiento de la cotorra durante su ciclo de anidación.
Diferencias encontradas respecto a trabajos anteriores
[image:38.612.118.494.367.608.2]Al comparar el comportamiento de la frecuencia de entradas obtenida por el presente trabajo y el trabajo de Macias-Caballero (1998) ambos coinciden en encontrar picos de actividad durante la incubación y las últimas semanas del emplumado, con el punto de actividad más bajo durante la etapa de desarrollo. La principal diferencia encontrada entre ambos trabajos radica en la intensidad de dichos picos, mientras que en el trabajo de Macias-Caballero el pico más alto de actividad es de 7.4 entradas durante la segunda semana de incubación (Figura 10), en el presente trabajo es de 3.5 entradas (Figura 2)durante los últimos días de el emplumado.
Figura 10. Número de visitas efectuadas a los nidos exitosos del risco rojo a lo largo de la temporada 1997 (n=10) (Tomada de Macias-Caballero 1998)
datos. Al hacer uso de una fecha promedio asume que todos los nidos se encuentran en la misma etapa de anidación de manera simultánea y se corre el riesgo de que dentro de un mismo día se considere a nidos que se encuentran en diferentes etapas de la anidación. Cabe aclarar que el trabajo de Macias-Caballero (1998) unicamente utilizó datos de las temporadas de anidación de 1995 al 1997, mientras que el presente trabajo hizo uso de los datos de las temporadas 1997-2005 por lo que el mayor tamaño de muestra tiende a diluir los nidos con alta frecuencia de entradas, ocasionando que en general se observe una menor frecuencia de entradas en el presente trabajo. Sin embargo debido al mayor tamaño de muestra el resultado del presente estudio se considera más representativo del comportamiento de anidación en los nidos exitosos de la cotorra.
Frecuencia de visitas al nido
[image:39.612.81.533.370.548.2]Como se mencionó con anterioridad, la cotorra presenta un patrón de visitas múltiples al nido, numerosos estudios han documentado que dentro de la familia de los psitácidos existe una gran variabilidad en el número de visitas diarias al nido por parte de los padres. En la tabla 4 se presentan los resultados encontrados para el número de visitas diarias en estudios con otras especies de psitácidos neotropicales.
Tabla 4.Frecuencia de visitas diarias en los nidos de varias especies de psitácidos.
Autor Especie Número de Visitas/ Día
Snyder, N.F. Amazona vittata
Enkerlin-Hoeflich, E. (1995) Amazona autumnalis 2.09
Enkerlin-Hoeflich, E. (1995) Amazona oratrix 2.18
Enkerlin-Hoeflich, E. (1995) Amazona virdigenalis 2.08
Iñigo-Elias, E (1996) Ara macao 5.00-9.00
Gnam, R. Amazona leucocephala
bahamensis
0.50 – 5.55 Koeing, S.E. (1991) Amazona agilis 3.40
Macias-Caballero, C. (1998) Rhynchopsitta terrisi 2.40-7.40
Renton, K. (2000) Amazona finschi 2.08-2.60
Valdez-Juarez, S. (2006) Rhynchopsitta terrisi 1.97 – 3.00
Debido a que no existe información en los factores que determinan la variabilidad de las entradas al nido en el género Rhynchopsitta, se utilizó como referencia a la información
existente para el género Amazona.
Los factores que influencian la variabilidad intraespescífica en el número de visitas diarias al nido dentro del género Amazona no están bien definidos, se ha observado una
elevaciones requieran de menores cantidades de alimento ya que experimentan un menor gasto energético en la termorregulación, alternativamente puede tratarse de una adaptación para prevenir el sobrecalentamiento durante las horas mas calientes del día (Luescher 2006).
Lo anterior puede apreciarse en la tabla 4, donde las especies de hábitats relativamente elevados y húmedos (Ara macao, Amazona vitata y A. agilis) presentan una tendencia hacia
las entradas múltiples, mientras que las especies de hábitats secos y bajos, (Amazona finschi, A oratrix, A. virdigenalis A. autumnalis), presentan una patrón constante de 2
entradas al día.
El hábitat de la cotorra se conforma principalmente de pino-oyamel-ayarín, de pino, pino piñonero y de pino-encino, en un rango de altitud que va desde los 2300 hasta los 3000 msnm (Ortiz-Maciel 2000), con una precipitación anual promedio que varia entre los 230 y 1100 mm (CCA-ITESM 1994). Los climas en la Sierra Madre Oriental van de áridos a semiáridos, no obstante la altitud en la que habita la cotorra impide que las temperaturas alcancen extremos que pudieran actuar como una fuente de estrés limitando el número de entradas diarias como presumiblemente ocurre con especies de tierras más bajas, sin embargo no se debe considerar que las condiciones climáticas son el único factor actuando sobre el los patrones de visitas diarios de la cotorra, adicionalmente las entradas múltiples se pueden deber a defensa contra depredadores, aun cuando debido a lo inaccesible de los nidos es poco probable que mamíferos o reptiles puedan llegar a depredar un nido, aunque se ha observado a cuervos y halcones cola roja ingresar a cavidades. Adicionalmente el número de visitas puede verse influenciado por el valor energético y nutricional de la dieta de los pollos, aunque son necesarios estudios adicionales en la dieta de los pollos para sustentar dicha hipótesis. En la experiencia del autor el acceso a los nidos es extremadamente difícil y riesgoso, por lo que es poco probable que se llegue a contar con dicha información para la cotorra, sin embargo estudios de la dieta de los pollos de R. pachyrhyncha pudieran extrapolarse para R. terrisi dado lo similar de su dieta, con lo cual
se podrían fundamentar hipótesis más sólidas sobre la razón de las entradas múltiples que pudieran llevar a una mejor comprensión de los hábitos reproductivos de la cotorra.
Variabilidad en la fecha de salida de los volantones
Se comprobó que la cotorra presenta variaciones en la fecha de salida de los volantones, dicha variación puede ser ocasionada por una variación en la fecha de inicio de incubación y/o por la influencia de los factores ambientales en el desarrollo de los pollos.
Estudios con Myopsitta monachus (Navarro et. al.1992) encontraron que el inicio de la
pachyrhyncha en pobres condiciones de salud presentan una mayor estadía en el nido y por
lo tanto tienen una mayor longitud alar en el momento de realizar el primer vuelo. Renton (2002) encontró que la tasa de crecimiento de los pollos de A. finschi está relacionada
proporcionalmente con la precipitación debido a que a mayor precipitación hay una mayor abundancia de alimento.
La Sierra Madre Oriental (en adelante SMO) presenta condiciones climáticas muy cambiantes anualmente, dentro del periodo de estudio se incluyen años inusualmente secos, como 1998 y 1999 que tuvieron una precipitación anual de 390.5mm y 435.5mm respectivamente y años inusualmente húmedos como el 2005 en el cual debido a la presencia del huracán Rita se presentó una precipitación anual record de 1111.4.mm (CNA 2006). Lo altamente variable de las condiciones anuales en la SMO puede jugar un papel muy importante en el acomodo temporal del ciclo de anidación de la cotorra, sin embargo aun no se cuenta con información suficiente como para establecer la influencia que las condiciones ambientales pudieran ejercer sobre la temporada de anidación de la cotorra. Con base en lo observado se propone que la variación en la fecha de salida de los pollos anualmente puede estar relacionada con varios factores ambientales actuando de manera simultánea en la selección de fecha de postura y en el desarrollo de los pollos, generando la variabilidad interanual observada en los nidos de la cotorra.
En el caso nidos excepcionalmente tardíos, como los presentandos en la temporada 2002, la demora en su fecha de vuelo se puede deber a que fueran producto de un segundo intento de postura posterior a uno fallido como se ha reportado para nidos excepcionalmente tardíos en otros psitácidos (Enkerlin-Hoeflich 1995).
Incubación
Durante esta etapa se observa una elevada frecuencia de entradas al nido que contrasta fuertemente con lo observado para otros psitácidos que también realizan entradas múltiples y en los cuales durante la incubación se registra la menor frecuencia de entradas para toda la temporada (Wilson et. al. 1995). Adicionalmente no se encontró una mayor frecuencia de
entradas realizadas por 1 sólo individuo, lo cual se esperaría si como ocurre en otras especies únicamente uno de los padres ingresara al nido.
refleja la frecuencia en la transferencia de alimento de macho a hembra. La conducta anterior también provee de una explicación de por qué contrario a lo esperado no se observó una diferencia en el número de entradas realizadas por 1 ó 2 individuos, ya que es frecuente que después de efectuada la transferencia de alimento el macho ingrese junto con la hembra al nido. Es de destacarse que en otras especies de psitácidos durante la incubación el macho no entra al nido o lo hace en raras ocasiones (Wilson 1995, Enkerlin-Hoeflich 1995).
Aun tomando en cuenta lo anterior se considera que el número máximo de visitas registradas durante la incubación, es suficiente como para que se pueda pensar que la cotorra realiza visitas múltiples durante la incubación, más aun considerando que también se ha observado que la hembra sale del nido a recibir el alimento, cuando es llamada por el macho sin que este ingrese a la cavidad.
Patrones de actividad diaria durante la incubación
Como se mencionó con anterioridad, dentro de los patrones de conducta diarios se registraron 2 tendencias que no parecen corresponder con los hallazgos realizados con otras especies de psitácidos. En la figura 5 se puede apreciar que durante la incubación la mayoría de las segundas entradas se producen durante la misma hora que la primera entrada, debido a lo corto del lapso transcurrido entre la primera y segunda entrada es poco probable que en este caso la segunda entrada se trate de una visita de alimentación, la coincidencia de ambos picos se debe al comportamiento de transferencia de alimento descrito anteriormente.
El segundo comportamiento atípico consta del registro de una mayor cantidad de salidas durante la hora 11 podría explicarse por una mayor salida de individuos hacia el final del día, lo cual significaría que uno o ambos padres durmieran fuera de la cavidad. Esta hipótesis no coincide con lo reportado para R. pachyrhyncha en donde ambos padres
duermen en el nido durante la incubación (Lanning y Shiftlet 1983). Los resultados se deben de tomar con cautela ya que durante la incubación solamente 7 sesiones cubrieron hasta la hora 11, durante las cuales se registraron 2 salidas, lo que ocasionó que el porcentaje relativo de salidas por sesión sea elevado, sin que el tamaño de la muestra llegue a ser representativo del comportamiento de la cotorra.
Desarrollo
El hecho de que estos 2 fenómenos coincidan se debe a que los pollos comienzan a desarrollar capacidad de termorregulación y pueden ser dejados solos por periodos de tiempo más prolongados, por lo que la frecuencia de la transferencia de alimento macho-hembra disminuye hasta desaparecer alrededor de la 6ª o 7ª semana y la macho-hembra comienza a salir por periodos más prolongados de hasta 6 horas, lo anterior coincide con lo observado para otros psitácidos en donde la hembra abandona el nido por periodos más prolongados conforme los pollos adquieren la capacidad de termorregulación (Snyder et. al. 1987, Gnam
1991, Waltman y Beissenger 1992, Wilson et. al. 1995, Enkerlin-Hoeflich 1995)
Patrones de actividad diaria durante el desarrollo
Al analizar el patrón de conducta durante el desarrollo se debe tener en consideración que ésta es una etapa de transición, por lo que durante las primeras semanas de esta etapa se observan patrones de conducta similares a los de la incubación, mientras que hacia el final se esperaría observar patrones de conducta similares al emplumado.
En las primeras semanas, cuando los pollos carecen de capacidad de termorregulación los padres duermen en la cavidad, por lo que se observa un mayor número de entradas durante las ultimas horas, producto de los padres regresando a dormir al nido, una vez que los pollos pueden pasar la noche sin cuidado parental, los padres no duermen en el nido, lo que produce el elevado número de salidas observado durante la 6a hora que corresponde a los padres realizando la última transferencia de alimento del día y saliendo del nido para no regresar sino hasta el día siguiente, lo que a la vez explica el elevado número de entradas durante la primera hora.
A diferencia de la incubación, se observa que el pico de segundas entradas no es coincidente con el pico de primeras entradas, esto se debe a que las hembras comienzan a abandonar el nido a partir de la quinta o sexta semana de iniciada la incubación, por lo que ya no se realizan transferencias de alimento de macho a hembra. Si bien se sabe que la duración de los viajes en busca de alimento varían enormemente en distancia recorrida y duración del viaje (Ortiz-Maciel 2000), durante el desarrollo parece haber una tendencia hacia viajes con 2 horas de duración, tomando en cuenta que el pico de primeras entradas se observa en la hora 2 y el de segundas entradas en la hora 4, lo cual coincide con las observaciones realizadas para la especie hermana (R. pachyrhyncha), donde los padres se
ausentaban del nido por periodos de 2 a 5 horas (Lanning y Shiflett 1983).
Emplumado
Basado en observaciones de campo del autor durante la temporada 2003 y 2005 se plantea la posibilidad de que el incremento en las entradas en la semana final de la temporada no sea exclusivamente con el objetivo de alimentar a los pollos. Se observó que en repetidas ocasiones 1 ó 2 padres entraban a la cavidad, vocalizando de manera continua mientras permanecían dentro de la cavidad (por alrededor de 1 o 2 minutos) y después abandonaban el nido para exhibir el comportamiento de “sacar pollo” descrito por Macias-Caballero (1998), este comportamiento llegó a repetirse varias veces al día. Debido a lo breve de la permanencia de los padres en el nido y al comportamiento exhibido una vez fuera de éste se puede inferir que las visitas hacia el final de la temporada tienen el propósito de motivar a los pollos a salir del nido.
La ocurrencia de un gran número de entradas cuya función no es la transferencia de alimento denota que la cotorra sufre de poco estrés por depredación ya que a diferencia de otros psitácidos, no limita el número de entradas al nido en respuesta a la presencia de depredadores.
Otro aspecto que cabe destacar es que durante el desarrollo, la transferencia de alimento a los pollos no necesariamente es llevada a cabo por los 2 padres, en el caso de un nido en particular durante la temporada 2005, se observó en numerosas ocasiones que ambos padres llegaban al área pero sólo uno ingresaba a la cavidad mientras que el otro permanecía perchado en un agave cercano, esperando hasta por 1 ó 2 horas que su pareja saliera del nido, una vez que ésta salía ambos abandonaban el área en busca de alimento.
Patrones de actividad diaria durante el emplumado
Coincidente con las etapas anteriores se observó el mayor pico de actividad durante la segunda y tercera hora del día, a partir de la cual se produjo una disminución paulatina de actividad, sin embargo a diferencia de las etapas anteriores, el emplumado es la única etapa en la que de manera constante se registró la misma cantidad de entradas y salidas por hora, producto de que ambos padres tienen patrones de conducta similares a lo largo de toda la etapa de emplumado.
Al igual que durante el desarrollo, se observa una tendencia de 2 horas en la duración de los viajes en búsqueda de alimento, ya que el pico de primeras entradas se produce durante la hora 2, con el pico de segundas entradas durante la hora 4 con la tercera entrada distribuida de manera homogénea a partir de la 4a hora.
Conclusiones
(1) La cotorra presenta un patrón flexible de visitas diarias al nido en las 3 etapas de anidación, variando desde 0 a 14 visitas diarias. Sin embargo es altamente probable que no ocurra transferencia de alimento en todas las visitas.
(2) Cada una de las tres etapas de anidación presenta una frecuencia de entradas característica. Siendo el emplumado la etapa más activa (3.00), seguida por la incubación (2.67) y con la frecuencia más baja durante el desarrollo (1.97).
(3) A diferencia de otros psitácidos el macho de la cotorra sí entra al nido durante la incubación, pudiendo realizar transferencia de alimento tanto fuera como dentro del nido. (4) La transferencia de alimento macho-hembra cesa alrededor de la sexta y séptima semana de anidación, cuando la hembra comienza a abandonar el nido en compañía del macho.
(5) Los horarios de visita diarios para las tres etapas de anidación coinciden en picos de actividad durante la segunda y tercera hora con un descenso de actividad paulatino a partir de la cuarta hora.
(6) La fecha de vuelo de los pollos de la cotorra varía anualmente probablemente en respuesta a la variación anual en las condiciones ambientales en la SMO lo que pudiera influir de manera simultánea en la selección de la fecha de inicio de la incubación y en el desarrollo de los pollos.