Participación de los queratinocitos en la respuesta inmunitaria contra
actinomicetoma
Alejandro Palma Ramos,* Laura Estela Castrillón Rivera,* Mirna Gabriela Encinas Parra,* Carmen Padilla
Desgarennes,** Roberto Arenas Guzmán***
Artículo original
REsUMEN
Antecedentes: el micetoma es un síndrome clínico que consiste en lesiones inflamatorias deformantes, no dolorosas y fístulas que afectan
los tejidos cutáneo y subcutáneo, la aponeurosis y el hueso. Generalmente aparece en el pie o en la mano. Para su estudio de ha dividido en tres zonas: la zona que rodea al grano se denomina la zona 1, compuesta por neutrófilos y macrófagos; la zona 2, por linfocitos T y la zona 3, por linfocitos B. Asimismo, se demostró la existencia de IgG, IgM y complemento en la superficie de los granos y en los filamentos que rodean al grano. Los neutrófilos y macrófagos recluidos en la lesión por el complemento participan en el ataque al grano. En la última
década se ha observado que los queratinocitos participan activamente en la respuesta inmunitaria innata, ya que expresan receptores tipo Toll (TLRs), y son una fuente importante de citocinas, quimiocinas y péptidos antimicrobianos, y son capaces de expresar antígenos de clase II. Con esto ha quedado claro que los queratinocitos pueden no sólo participar en la respuesta inmunitaria epidérmica, sino que tienen un papel importante en el inicio de la respuesta.
Objetivo: mostrar a los queratinocitos como células que participan de manera importante en la respuesta inflamatoria, contra los actino -micetomas por Nocardia brasiliensis y Actinomadura madurae.
Material y método: las biopsias de los pacientes con micetoma (por N. brasiliensis, A. madurae) las proporcionaron el Departamento
de Micología del Centro Dermatológico Dr. Ladislao de la Pascua y del Hospital Dr. Manuel Gea González. En el estudio histopatológico se realizó primero la técnica de hematoxilina y eosina, para la búsqueda y caracterización del grano; el marcaje se realizó con el equipo
comercial Cell and tissue staining kit, HRP-AEC System anti-cabra (catálogo CTS009, R&D Systems), (avidina-biotina y 3,3’ diamino -benzidina), proyectado para la localización de antígenos en un rango de amplio espectro de especímenes histológicos y citológicos. Los anticuerpos utilizados fueron: 1) anti-IL-1β humano producido en cabra por R&D Systems y 2) anti- TNF-α humano producido en cabra
por R&D Systems.
Resultados: en los actinomicetomas por N. brasiliensis y en los actinomicetomas por A. madurae se observaron citocinas (TNF-α e IL-1β) unidas al grano, y en la epidermis (zona de queratinocitos) se intensificó la marca que manifiesta las altas concentraciones de estas
citocinas.
Conclusión: la existencia de los polimorfonucleares neutrófilos y su activación obedece a la secreción de los mediadores proinflamatorios provenientes de la activación de los queratinocitos, por lo cual se observa una respuesta de tipo innato en este síndrome.
Palabras clave: actinomicetoma, queratinocitos, IL-1β, TNF-α, N. brasiliensis, A. madurae.
AbsTRACT
background: Mycetoma is a clinical syndrome that consists in deformant, inflammatory, painless lesions, and fistulae affecting cutaneous and
subcutaneous tissues, aponeurosis and bone; usually in feet. Histologically, the lesion has been described in three zones: first closer to grain
or zone 1 composed mainly by neutrophils and macrophages; the zone 2 by lymphocytes T, and the distant zone or zone 3 by lymphocytes B. Macrophages and neutrophils are recruited by chemoattractants derived by complement molecules possibly as consequence of IgG,
IgM and complement present in surface and filaments surrounding the grain. In last decade, it has been demonstrated that keratinocytes
participates actively in innate immune response, because they express Toll-like receptors (TLRs) and are an important source of cytokines, chemokines and antimicrobial peptides. Besides, these cells are able to express class II MHC antigens. In consequence, keratinocytes have an important role in epidermic immune response and now they are considered an inductor to adaptative immune response. Objective: To demonstrate keratinocyte activation as a producer of proinflammatory cytokines (IL-1β and TNF-α) in actinomycetoma by Nocardia brasiliensis and Actinomadura madurae.
Material and methods: Biopsies from patients with mycetoma by N. brasiliensis and A. madurae were provided by Mycology Department
from Dermatological center Dr. Ladislao de la Pascua and Dr. Manuel Gea González Hospital. The histopathological study was made by H-E stain to find and characterization of grains. For detection of interleukins (IL-1β and TNF-α) in situ, immunohistochemical analysis was
done using Cell and tissue staining kit, HRP-AEC, R&D Systems.
Results: IL-1β and TNF-α were observed bounded to grains and in epidermis (keratinocyte zone), where the stain was intensified due the
presence of high concentration of these cytokines.
Conclusion: The presence of polymorphonuclear neutrophils activated is consequence of proinflammatory mediators secretion derived from keratinocytes. As a result of this fact, the immune innate response is evident in this syndrome.
* Profesor investigador del Laboratorio de Inmunopotencia-dores. Departamento de Sistemas Biológicos, Universidad
Autónoma Metropolitana, Unidad Xochimilco.
** Laboratorio de Micología, Centro Dermatológico Dr. Ladislao de la Pascua, Servicios de Salud Pública del DF.
*** Servicio de Micología, Hospital General Dr. Manuel Gea
González, Secretaría de Salud.
Correspondencia: Dr. Alejandro Palma Ramos. Departamento de Sistemas Biológicos, Universidad Autónoma Metropolitana, Unidad
Xochimilco. Calzada del Hueso 1100, colonia Villa Quietud, CP
04960, México, DF.
Correo electrónico: [email protected]. Recibido: agosto, 2009. Aceptado: septiembre, 2009.
Este artículo debe citarse como: Palma RA, Castrillón RLE, Encinas PMG, Padilla DC, Arenas GR. Participación de los queratinocitos en la respuesta inmunitaria contra actinomicetoma. Dermatol Rev Mex 2009;53(5):225-33.
La versión completa de este artículo también está disponible en:
www.nietoeditores.com.mx
E
l micetoma es un síndrome clínico que consisteen lesiones inflamatorias deformantes, no dolo -rosas y fístulas que afectan los tejidos cutáneo y subcutáneo, la aponeurosis y el hueso.1 Afecta
generalmente la extremidad inferior y de ésta el pie, ya que se trata de áreas expuestas directamente a la infección en 70 a 75% de los casos, pero puede aparecer en cualquier región del cuerpo. Su evolución es crónica y asintomática durante los primeros meses de aparición. La enfermedad es el resultado de la implantación traumática de un micro-organismo del suelo dentro de los tejidos. Las lesiones se componen de abscesos que supuran, granulomas y fístulas que drenan, así como granos que son característicos de los agentes etiológicos. Éstos incluyen una amplia variedad de bacterias (actinomicetomas) y hongos (eumicetomas) del suelo.2
Predomina en varones, con una proporción de 4:1; excepto el micetoma por Actinomadura madurae, que predomina en las mujeres en relación de 2:1. Ocurre con mayor frecuencia a la edad de 16 a 45 años. Los estados de la República Mexicana con más casos de micetoma en orden de frecuencia son: Morelos, Guerrero, Veracruz, Michoacán, Oaxaca, Guanajuato, Puebla, Hidalgo, San Luis Potosí, Estado de México, Sinaloa y Jalisco.
En México, son más frecuentes los actinomicetomas que los eumicetomas (97.9 vs 2.1%). De los actinomi-cetomas, el más frecuente es Nocardia con 86.5%, y de ésta la especie brasiliensis es la más frecuente; después
Actinomaduramadurae con 10% (incluida A. pelletieri),
y por último Streptomyces somaliensis, con 1.3%. Los eu-micetomas representan 2.1%, predominan los producidos por granos negros y la especie más frecuente es Madurella mycetomatis.3
En eumicetomas causados por Madurella mycetomatis
se han encontrado tres tipos de reacción inflamatoria.4 La reacción tipo I se caracteriza por una zona de neutrófilos
que rodean al grano, una zona intermedia de macrófagos, células gigantes y una zona periférica que contiene linfo-citos y células plasmáticas. La reacción tipo II no tiene la
zona de neutrófilos y el grano está rodeado por macrófa -gos y células gigantes. La reacción tipo III consiste en un granuloma epitelioide discreto, con células gigantes de Langhans sin granos bien formados.5
Mediante el uso de técnicas de inmunohistoquímica se mostró que en la reacción tipo I, la zona que rodea al
grano dio positiva para CD15 (neutrófilos); la zona 2 fue
positiva para CD68 (macrófagos) y CD3 (linfocitos T), mientras que la zona 3 contiene células positivas para CD20 (linfocitos B). Asimismo, se demostró la existencia
de IgG, IgM y complemento en la superficie de los granos y en los filamentos que rodean al grano. Los neutrófilos
y macrófagos recluidos en la lesión por el complemento
participan en el ataque al grano. El perfil de citocinas en
la lesión y en los ganglios linfáticos regionales fue prin-cipalmente las citocinas IL-4 e IL-10 provenientes de las células Th2.6
En la piel, la epidermis provee la primera línea de de-fensa contra los patógenos, los queratinocitos forman 95% de las células de la epidermis; se creía que su función era únicamente mantener la estructura de la epidermis vía la producción de citoqueratina y como barrera física a una gran variedad de microorganismos exógenos. En la última década, se ha observado que los queratinocitos expresan receptores tipo Toll (TLRs), y son una fuente importante de citocinas, quimiocinas y péptidos antimicrobianos, que son capaces de expresar antígenos de clase II. Con esto ha quedado claro que los queratinocitos pueden no sólo participar en la respuesta inmunitaria epidérmica, sino que tienen un papel importante en el inicio de la respuesta.7,8 La
tarea de la respuesta inmunitaria innata de la piel consiste en movilizar rápidamente los componentes de la respuesta
del huésped, incluidos los queratinocitos, neutrófilos, mas
-tocitos, eosinófilos y macrófagos. Estas células expresan
ac-tivados por componentes de microorganismos patógenos.9 Los receptores son específicos de patrones moleculares
asociados con patógenos (PAMPs), como el lipopolisacári-do (LPS) de bacterias gramnegativas, mananos y zymosan en la pared celular de levaduras; los lipoarabinomananos (LAM) son un componente de la pared celular de micobac-terias, y el peptidoglucano (PGN) o el ácido lipoteicoico (LTA) de bacterias grampositivas, así como lipopéptidos de varios microorganismos.9 El reconocimiento a patrones
de patógenos fúngicos, como el lipopéptido tripalmitoila-to, ocurre a través del heterodímero TLR-2/TLR-1, el peptidoglucano, el ácido lipoteicoico y lipoproteínas de grampositivas (Staphylococcus aureus o Borrelia burg-dorferi), el lipoarabinomanano de las micobacterias y el lipopolisacárido de Leptospira y Porphyromonas se unen mediante los TLR-2,10 TLR-6/TLR-2 se unen al zymosan
y al diacil lipopéptido.11 El TLR-3 se une al ARN doble
cadena, los lipopolisacáridos se unen a TLR-4 unidos con CD-14 y MD-2 (detectado en queratinocitos),12 la flagelina
por 5, ARN de una cadena (ARNss) viral por TLR-7 y TLR-8, y el ADN (CpG ADN) bacteriano o viral por TLR-9.13 El reconocimiento de los patrones moleculares
asociados con patógenos por los receptores tipo Toll inicia una señal en cascada mediada por el receptor citoplásmi-co Toll-interleucina 1 (TIR), que inicia la producción de
mediadores proinflamatorios, péptidos antimicrobianos,
moléculas de adhesión y enzimas inducibles.14
También, el reconocimiento de los patrones molecula-res asociados con patógenos por los receptomolecula-res tipo Toll inicia una señal en cascada mediada por el receptor Toll-interleucina 1 citoplásmico que interactúa con una proteína adaptadora MyD88, y ésta recluta una serin/treonina qui-nasa (la IL-1R-asociada con la quiqui-nasa, lo que se conoce como IRAK). IRAK se activa por fosforilación y entonces se une con el receptor de TNF asociado con el factor 6
(TRAF-6), activando dos rutas distintas, JNK y NF-kB. La actividad de NF-kB es regulada por IkB, (que es degradada cuando el receptor tipo Toll se une con MyD88). De esta manera, NF-kB se une a la región promotora de los genes
de citocinas proinflamatorias, péptidos antimicrobianos,
enzimas inducibles y moléculas de adhesión.14
Son muchos los papeles de los queratinocitos en la respuesta inmunitaria contra las infecciones. Los patrones moleculares asociados con patógenos de
microorganis-mos inducen la producción de citocinas proinflamatorias
(TNF-α, IL-1β quimiocinas IL-8/CXCL8), y péptidos an-timicrobianos (hBD-2), por la activación de los receptores tipo Toll en queratinocitos. Los péptidos antimicrobianos derivados de los queratinocitos matan los patógenos y pre-vienen la diseminación de infecciones sistémicas.
En resumen, la función antimicrobiana de las β-defensinas regulan el reclutamiento de las células T de memoria y células dendríticas inmaduras a través de la interacción con CCR6. Además, los péptidos antimicrobianos son capaces de activar células dendríticas inmaduras por medio del TLR-4 e iniciar la respuesta inmunitaria adaptativa. Las citocinas derivadas de los queratinocitos (TNF-α) y quimiocinas (IL-8/CXCL8) median el reclutamiento de células inmunes efectoras
(neu-trófilos) a los sitios de infección, aumentan la extravasación
y forman un gradiente químico en la dermis.15
En estudios anteriores de nuestro grupo de trabajo hemos observado pocas citocinas, como el INF-γ, IL-4 e IL-1016 en actinomicetomas, en un estudio longitudinal en
la formación de actinomicetoma inducido por Nocardia brasiliensis en ratón (figura 1).
La poca presencia del INF-γ puede explicarse por la participación de células NK (natural killer), que producen esta citocina en la respuesta inmunitaria de tipo innata por la existencia de la IL-12 e IL-15 provenientes de quera-tinocitos (IL-12) y de las células de Langerhans (IL-12
e IL-15); la IL-10 de algunos macrófagos activados, con una muy baja o casi nula participación de los linfocitos, ya que en estos síndromes las células que se encuentran
en la reacción son preferentemente neutrófilos y algunos
histiocitos con predominio de los primeros.17
Nuestra propuesta se basa en la poca participación de los linfocitos T CD4 y de los linfocitos T CD8,18 porque
el grano en el actinomicetoma está formado por bacterias rodeadas de un polisacárido neutro en el caso de actinomi-cetomas por Nocardia brasiliensis, y un polisacárido ácido sulfatado en el caso de actinomicetomas por Actinomadura madurae,19,20 y no hay presentación de antígeno ya que
ésta se da cuando el antígeno es un péptido y aquí son polisacáridos los que están en la capa que forma el grano, de tal manera que la respuesta se mantiene por medio de la activación de los queratinocitos que se encuentran en la epidermis, los cuales, aunque su función principal es la síntesis de queratina, colágeno y factores de crecimiento, son células accesorias del sistema inmunitario, y expresan moléculas de adhesión que regulan el movimiento de célu-las a través de la piel. Son reservorio de IL-1β, IL-3, IL-6, IL-8, IL-12, GM-CSF (factor estimulante de colonias de granulocitos), TGF-β (factor de crecimiento transforman-te), M-CSF (factor estimulante de colonias de monocitos), PDGF (factor de crecimiento/diferenciación plaquetario), ETAF (factor activador de timocitos derivado de células epidérmicas), TNF-α (factor de necrosis tumoral). Ex-presan moléculas MHC I y MHC II, así como moléculas de adhesión ICAM-1; y también receptores tipo Toll, que son moléculas de reconocimiento a patrones moleculares asociados con patógenos, lo que resulta con la producción
de mediadores proinflamatorios como las citocinas TNF-α, IL-1β21 y quimiocinas como la IL-8/CXCL8, y otras con
propiedades antimicrobianas como las CCL18, CCL19, CCL20, CCL25, CXCL1, CXCL10.8
ObJETIVO
Mostrar a los queratinocitos como células que participan
de manera importante en la respuesta inflamatoria en los
actinomicetomas por N. brasiliensis y A. madurae.
MATERIAL y MéTODO
Biopsias: las biopsias de los pacientes con micetoma (por N. brasiliensis, A. madurae) las proporcionaron el
Departamento de Micología del Centro Dermatológico Dr. Ladislao de la Pascua y del Hospital Dr. Manuel Gea González. Se estudiaron tres cortes de cada paciente y fueron tres pacientes por tipo de actinomicetoma. Se practicó el marcaje por histoquímica para la demostración de la existencia de TNF-α e IL-1β.
Estudio histopatológico: se realizó primero la técnica de hematoxilina y eosina, para la búsqueda y caracteriza-ción del grano.
Hematoxilina-eosina:22 desparafinar, colorear con
hematoxilina de Harris durante un minuto y lavar. Diferen-ciar con alcohol-ácido, lavar y virar con agua amoniacal; lavar otra vez, colorear con eosina durante 30 segundos, deshidratar y montar. Se observaron núcleos azules y citoplasma en color rosa o naranja.
Histoquímica: el marcaje se realizó con el equipo comercial Cell and tissue staining kit, HRP-AEC System anti-cabra (catálogo CTS009, R&D Systems), (avidina-biotina y 3,3’ diaminobenzidina), proyectado para la localización de antígenos en un rango de amplio espectro de especímenes histológicos y citológicos.
Anticuerpos: 1) IgG anti-IL-1β humano producido en cabra por R&D Systems, 2) IgG anti-TNF-α humano producido en cabra por R&D Systems.
REsULTADOs
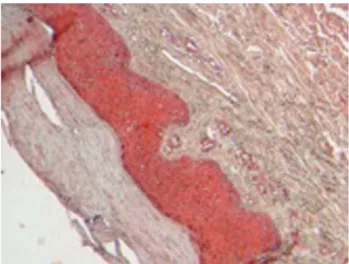
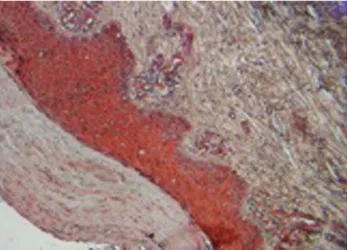
Se analizó como control piel humana sana proporcionada por el Centro Dermatológico Ladislao de la Pascua, a la cual se le marcó a un corte con IgG anti- NF-α humana hecho en cabra y otro con IgG anti-IL-1β humana hecho en cabra y tratados con un conjugado anti-cabra (equipo comercial Cell and tissue staining kit, HRP-AEC System anti-cabra, catálogo CTS009, R&D Systems, avidina-biotina y 3,3’ diaminobenzidina), y contrastados con hematoxilina. Se observó que no hubo coloración (roja) en ninguno de los dos cortes tratados con los anticuerpos y, por tanto, se presume la ausencia de estas citocinas
(figuras 2 y 3).
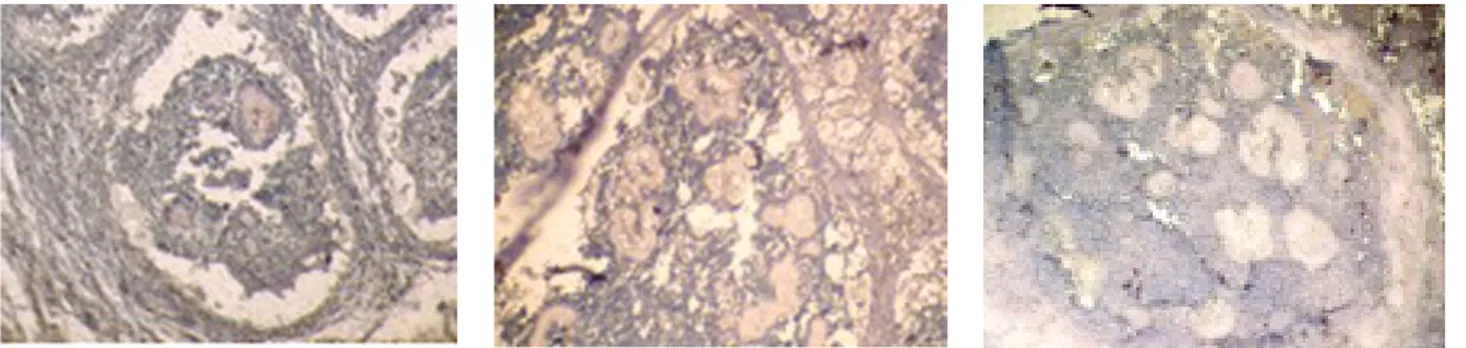
Se estudiaron tres biopsias de pacientes con actinomi-cetoma por Nocardia brasiliensis y actinomicetoma por
Actinomadura madurae.
Al primer corte de cada biopsia de paciente se le prac-ticó la tinción de hematoxilina y eosina para corroborar la presencia del grano y realizar la caracterización del
Figura 2. Corte histológico de piel humana sana tratada con IgG anti-TNF-α humana hecho en cabra y con un conjugado anti-IgG de cabra conjugado con avidina-biotina y 3,3’ diaminobenzidina. No se observó el TNF-α.
Figura 3. Corte histológico de piel humana sana tratada con IgG anti-IL-1β humana hecho en cabra y con un conjugado anti-IgG de cabra conjugado con avidina-biotina y 3,3’ diaminobenzidina. Casi no se observó la IL-1β.
Primero analizamos los actinomicetomas por Nocardia brasiliensis, se marcó con anti-TNF-α. Se encontró el TNF-α en el grano (figura 6) y en los queratinocitos de la
epidermis (figura 7).
En la figura 7 se observó una coloración roja intensa en
la zona epidérmica en donde se encuentran los
queratino-citos, lo que manifiesta altas concentraciones de TNF-α en esta zona.
Para analizar los actinomicetomas por Nocardia brasiliensis se marcó con anti-IL-1β, se encontró esta
Figura 4. Actinomicetoma por Nocardia brasiliensis. H y E 40x.
Figura 5. Actinomicetoma por Actinomadura madurae. H y E 40x.
citocina en el grano (figura 8) y en los queratinocitos de la epidermis (figura 9).
En el actinomicetoma causado por Actinomadura madurae, primero se realizó la tinción de hematoxilina y eosina para observar el grano y caracterizar el micetoma
(figura 5).
Posteriormente se realizó la histoquímica, se marcó con IgG anti-TNF-α humana hecho en cabra y Cell and tissue staining kit, para observar la presencia de TNF-α
Figura 7. Intensa coloración roja que demuestra las altas concen-traciones de TNF-α en la parte superior derecha, perteneciente a la capa epidérmica (zona de queratinocitos). También se observó marca en el grano, característico de Nocardia brasiliensis. Se tra-tó con IgG anti-TNF-α humana hecho en cabra y Cell and tissue staining kit 40x.
Figura 8. Color rojo intenso en el grano que demuestra la existencia de IL-1β en el mismo. Se trató con IgG anti-IL-1β humana hecho en cabra y Cell and tissue staining kit 40x.
Figura 9. Intensa coloración roja que demuestra las altas concen-traciones de IL-1β en la capa epidérmica (zona de queratinocitos). También se observó marca en el grano, característica de Nocardia
brasiliensis. Se trató con IgG anti-IL-1β humana hecho en cabra y Cell and tissue staining kit 40x.
en la lesión, se encontró unido al grano y en la epidermis
en donde están los queratinocitos (figura 10).
En la piel, encontramos el TNF-α en la epidermis, zona
de queratinocitos (figura 11).
En el actinomicetoma por Actinomadura madurae, la IL-1β se observa en el grano, ya que el péptido se adhiere
directamente a éste (figura 12).
Figura 11. Intensa coloración roja que demuestra las altas concen-traciones de TNF-α en la capa epidérmica (zona de queratinocitos). Se trató con IgG anti-TNF-α humana hecho en cabra y Cell and tissue staining kit 40x.
Figura 12. Color rojo intenso en el grano de Actinomadura madurae, que demuestra la existencia de IL-1β en el mismo. Se trató con IgG anti-IL-1β humana hecho en cabra y Cell and tissue staining kit 40x.
Al observar la epidermis podemos corroborar el hallaz-go de la IL-1β en ésta, y lo más probable es que provenga
de los queratinocitos (figura 13).
DIsCUsIóN
Es muy probable que la participación de las células dendríticas en este síndrome se encuentre gravemente mermada, ya que la principal función de éstas es la de presentar antígenos a las células T en el ganglio linfático más cercano, y en los sitios de la lesión cuando estos linfocitos T son de memoria; una restricción importante
Figura 13. Intensa coloración roja que demuestra la existencia de IL-1β en la capa epidérmica (zona de queratinocitos), en el actino-micetoma por Actinomadura madurae. Se trató con IgG anti-IL-1β humana hecho en cabra y Cell and tissue staining kit 40x.
es que para funcionar como células presentadoras de antí-geno (CPA), estas células sólo presentan péptidos unidos a su complejo principal de histocompatibilidad (MHC) ya sea de clase I o de clase II, y la secreción importante de citocinas como la IL-12, IL-1β, IL-6, IL-15, IL-18, TNF-α, y quimiocinas. Por otra parte, en estas células hay preferencia para el reconocimiento de antígenos unidos a anticuerpos o a complemento, ya que tienen receptores para el Fce (RFce), Fcγ (RFcγ)y C3b (CR1) y no usan el receptor de manosa,23 tomando en cuenta que en
el actinomicetoma las bacterias se encuentran envueltas por polisacáridos en los granos que produce Nocardia brasiliensis y los que produce Actinomadura madurae,
por lo que son pocas las bacterias fagocitadas como la existencia de péptidos secretados por el microorganis-mo en el medio, lo que activaría las células dendrítricas
(figura 14).
La función inmunológica de los queratinocitos es prevenir las infecciones sistémicas por patógenos poten-ciales, como primera línea de defensa y tienen un papel activo en la inmunidad de tipo innato. Se ha descrito que expresan de manera constitutiva e inducible receptores como TLR-1, TLR-2, TLR-3, TLR-4, TLR-5, TLR-6 y TLR-9, lo que les facilita el reconocimiento a patrones moleculares asociados con patógenos, lo que da como resultado que moléculas como los lipoarabinomananos o peptidoglucano se unan a TLR-2, provocando la
quimiocinas como la IL-8/CXCL8 y péptidos antimicro-bianos como CCL18, CCL19, CCL20, CCL25, CXCL1, CXCL10,24 estén presentes en el actinomicetoma y tal vez
en el micetoma en general.
CONCLUsIóN
La existencia de los polimorfonucleares neutrófilos y
su activación obedece a la secreción de los mediadores
proinflamatorios provenientes de la activación de los
queratinocitos. Por lo cual se observa una respuesta de tipo innato en este síndrome.
REFERENCIAs
1. González OA. Mycetoma. In: Cañizares O, editor. Clinical tropical dermatology. Oxford, UK: Blackwell Scientific, 1975;pp:24-29.
Figura 14. Modelo de respuesta inmunitaria ante el actinomicetoma.
2. Magaña M. Mycetoma. Int J Dermatol 1984;23(4):221-36. 3. Padilla C, Novales J, Juárez V, Flores A. Minimicetoma.
Presentación de un caso. Rev Cent Dermatol Pascua 2004;13(1):41-44.
4. El Hassan AM, Fahal AH, Veress Bela. Cell phenotypes, im-munoglobulins and complement in lesions of eumycetoma caused by Madurella mycetomatis. J Sudanese Dermatology 2006;4(1):2-5.
5. Fahal AH, Toum El, Gumaa EA, Maghoub SA, et al. Host tissue reaction to Madurella mycetomatis: new classification. J Med
Vet Mycol 1995;33:103-6.
6. Fahal AH. Mycetoma: a thorn in the flesh. Trans R Soc Trop
Med Hyg 2004;98:3-11.
7. Barker JN, Mitra RS, Griffiths CE, Dixit VM, Nickoloff BJ. Keratinocytes as initiators of inflammation. Lancet 1991;337(8735):211-4.
8. Barker JN, Jones ML, Mitra RS, et al. Modulation of keratino-cyte derived interleukin-8 which is chemotactic for neutrophils and T limphocytes. Am J Pathol 1991;139(4):869-76. 9. Janeway CA, Medzhitov R Jr. Innate immune recognition. Annu
10. Takeda K, Kaisho T, Akira S. Toll-Like receptors. Annu Rev Immunol 2003;21:335-76.
11. Baker BS, Ovigne JM, Powels AV, Corcoran S, Fry L. Normal keratinocytes express Toll-like receptors (TLRs) 1, 2 and 5: modulation of TLR expression in chronic plaque psoriasis. Br J Dermatol 2003;148(4):670-9.
12. Song PI, Park YM, Abraham T, et al. Human keratinocytes express functional CD14 and Toll-like receptor 4. J Invest Dermatol 2002;119(2):424-32.
13. Akira S, Takeda K. Toll-like receptor signaling. Nat Rev Immunol 2004;4(7):499-511.
14. Zhang G, Ghosh S. Toll-like receptor-mediated NF-kappaB activation: a phylogenetically conserved paradigm in innate immunity. J Clin Invest 2001;107(1):13-19.
15. Iwasaki A, Medzhitov R. Toll-like receptor control of the adaptive immune responses. Nat Immunol 2004;5(10):987-95.
16. López BA, Arenas GR, Vega MM, Castrillón RL, Palma RA.
Identificación de células y mediadores inflamatorios en le -siones de pacientes con diagnóstico de micetoma. Dermatol Rev Mex 2008;52(6):247-5.
17. Palma RA, Cuevas MM, Arenas GR, Vega MM, Castrillón RL. Diseminación hematógena por Nocardia brasiliensis en tres
ratones Balb/c a partir de un micetoma en el cojinete plantar.
Dermatol Rev Mex 2008;52(2):70-76.
18. Palma RA, Castrillón RL, Pizaña CA, Vega MM, et al. Subpo -blaciones de linfocitos T en el micetoma. Dermatol Rev Mex 2007;51(6):212-8.
19. Palma RA, Castrillón RL, Padilla DC, Reyes FF. Caracterización histoquímica de micetomas por Actinomadura madurae,
Nocar-dia brasiliensis y Madurella mycetomatis. Dermatol Rev Mex 2005;49:51-58.
20. Palma RA, Castrillón RL, Padilla DC, Rosas HL, Márquez
C. Purificación y determinación de la estructura de los
polisacáridos que forman el cemento de unión en granos de
actinomicetomas ocasionados por Actinomadura madurae y Nocardia brasiliensis. Dermatol Rev Mex 2006;50:165-73. 21. Castrillón RL, Palma RA, Padilla DC. La función inmunológica
de la piel. Dermatol Rev Mex 2008;52(5):211-24.
22. Cormack HD. La histología y sus métodos de estudio: His-tología de HAM. 1987;pp:1-28.
23. Ayala GI, Hernández SAM, Castell RA, Álvarez PSJ, et al. Par -ticipation of epidermal Langerhans cells in human pathology and their potential as targets for drug development: a review of literature. Proc West Pharmacol Soc 2005;48:13-20. 24. Pivarcsi A, Nagy I, Kemey L. Innate immunity in the skin: how
keratinocytes fight against pathogens. Curr Immunol Rev
2005;1:29-42.
Páginas de la sociedad Mexicana de Dermatología, A.C.
www.promedicum.org (para la comunidad médica)