Estudio de mutantes de Burkholderia sacchari incapaces de crecer en propionato (prp) y afectados en el uso de intermediarios de la α-oxidación de
este substrato.
Yeimy Paola Galindo Rozo
TRABAJO DE GRADO
Presentado como requisito parcial para obtener título de:
MICROBIÓLOGA INDUSTRIAL
PONTIFICIA UNIVERSIDAD JAVERIANA FACULTAD DE CIENCIAS
MICROBIOLOGÍA INDUSTRIAL BOGOTÁ D.C.
Estudio de mutantes de Burkholderia sacchari incapaces de crecer en propionato (prp) y afectados en el uso de intermediarios de la α-oxidación de
este substrato.
Yeimy Paola Galindo Rozo
TRABAJO DE GRADO
Presentado como requisito parcial para obtener título de: MICROBIÓLOGA INDUSTRIAL
DIRECTORA
Dra. Luiziana Ferreira da Silva
ASESOR DE TESIS Ana Carolina Suzuki Dias Cintra
PONTIFICIA UNIVERSIDAD JAVERIANA FACULTAD DE CIENCIAS
MICROBIOLOGÍA INDUSTRIAL BOGOTÁ D.C.
NOTA DE ADVERTENCIA
ARTÍCULO 23 DE LA RESOLUCIÓN Nº 13 DE JULIO 1946.
“ La Universidad no se hace responsable por los conceptos emitidos por sus alumnos
en sus trabajos de tesis. Sólo velará porque no se publique nada contrario al dogma
y a la moral católica y porque las tesis no contengan ataques personales contra
persona alguna, antes bien se vea en ellas el anhelo de buscar la verdad y la
Estudio de mutantes de Burkholderia sacchari incapaces de crecer en propionato (prp) y afectados en el uso de intermediarios de la α-oxidación de
este substrato
Yeimy Paola Galindo Rozo
APROBADO
_____________________________ Dra. Luiziana Ferreira da Silva
Farmacia y Bioquímica PhD. Ciencias Biológicas
(Microbiología) Directora
_____________________________ Dr. José Gregorio Cabrera Gomez
Ciencias Biológicas PhD. Ciencias Biológicas
(Microbiología) Jurado 1
_____________________________ Ana Carolina Suzuki Dias Cintra
Ciencias Biológicas M. Sc. Ciencias Biológicas
(Microbiología) Asesora
_____________________________ Dra. Érica Mendes Pereira
Química
PhD. Ciencias Biológicas (Biotecnología)
Estudio de mutantes de Burkholderia sacchari incapaces de crecer en propionato (prp) y afectados en el uso de intermediarios de la α-oxidación de
este substrato
Yeimy Paola Galindo Rozo
APROBADO
_____________________________ Dra. Angela Umaña M. Phill
Decana Académica Facultad de Ciencias
_____________________________ Dra. Janeth Arias
DEDICATORIA
Dedico este trabajo a Dios por ser mi fuerza espiritual, por guiarme y brindarme
sabiduría para cada uno de los momentos vividos y especialmente, por darme la
oportunidad de tener a mis seres queridos.
A mi padres Joaquín y Myriam por todo su amor, apoyo, por cada uno de los
sacrificios que hicieron para que realizara mis sueños, por su constante entrega y
confianza, porque me ayudaron a seguir adelante frente a cada una de las
adversidades presentadas.
A mis hermanos Fernando y Laura, por su cariño, apoyo y compañía en todos los
momentos de mi vida, especialmente durante mi formación, por ser parte importante
de éste proceso y ser incondicionales en cada una de las etapas vividas.
A mis Marías hermosas por su valiosa y sincera amistad, por ser parte importante
de mi vida y por permitirmen aprender que existe la verdadera amistad… Las adoro
niñas… Gracias por ser las mejores amigas del mundo.
A mi novio Edgar Leonardo, por todo su apoyo, comprensión, amor, cariño,
dedicación, entrega, por estar presente en todos los momentos de mi vida y por ser
tan incondicional.
AGRADECIMIENTOS
Agradezco especialmente a mi directora de Tesis Dra. Luiziana Ferreira da Silva
por el apoyo brindado, por la confianza depositada, por la oportunidad brindada y
por generar un mayor gusto por la biología molecular, haciéndome crecer con bases
sólidas en el inicio como investigadora.
Agradezco de forma especial también, al Dr. José Gregorio Cabrera Gomez por la
ayuda y acompañamiento a lo largo del desarrollo de mi trabajo de grado, así como
por cada una de las discusiones y sugerencias valiosas, que fueron de gran ayuda
para el esclareciendo de dudas.
A la Universidad de Sao Paulo (USP) ubicada en Sao Paulo (Brasil), especialmente
al Instituto de Ciencias Biomédicas, por permitirme realizar el presente trabajo, así
como por el préstamos de sus instalaciones y equipos, y demás materiales para el
desarrollo de la investigación.
A Ana Carolina Suzuki Dias Cintra y a Renata de Melo Peixoto, agradezco por
todas las enseñanzas profesionales y personales, que me hicieron madurar y crecer
como profesional, también por todo el apoyo brindado durante la realización de la
investigación, así como por su valiosa amistad y en general por cada uno de los
momentos vividos dentro y fuera del laboratorio. Meninas eu adoro vocês!
Agradezco de forma muy especial a mis amigos y compañeros de laboratorio Marco
Antonio, Érica, Mateus, Tathi y Dani, que colaboraron para la ejecución de éste
trabajo.
A mis mejores amigas Diana, Prunella, Viviana, Carolina y Rosa Carolina, por su
valiosa amistad y cariño desde el inicio de la carrera, así como por enviarmen desde
Colombia todo su apoyo, brindándome su cariño, dándome ánimos para seguir
adelante y realizar mis sueños; niñas gracias por todos los momentos vividos, las
adoro!!!
Finalmente quiero agradecer a mi familia, a mi abuelita y a mi novio Edgar
Leonardo por la paciencia, apoyo, cariño, confianza, consejos, amor y por estar
TABLA DE CONTENIDO
1 INTRODUCCIÓN ... 16
2 REVISIÓN DE LITERATURA... 19
2.1 QUÉ SON LOS POLIHIDROXIALCANOATOS (PHA)? ...19
2.2 CARACTERÍSTICA DE Burkholderia sacchari LFM 101. ...19
2.3 SÍNTESIS DE PHA EN BACTERIAS. ...21
2.4 UTILIZACIÓN DE SUBSTRATOS DE BAJO COSTO EN LA PRODUCCIÓN DE PHA...23
2.5 PRODUCCIÓN DEL CO-POLÍMERO P3HB-co-3HV ...25
2.5.1 UTILIZACIÓN DE ÁCIDO PROPIÓNICO COMO PRECURSOR DE UNIDADES 3HV. ...25
2.6 CATABOLISMO DE PROPIONATO EN DIFERENTES ORGANISMOS. .. ...26
2.6.1 UTILIZACIÓN DE MICROORGANISMOS RECOMBINANTES INCAPACEZ DE CRECER EN ÁCIDO PROPIÓNICO. ...32
3 JUSTIFICACIÓN ... 35
4 OBJETIVOS... 37
4.1 OBJETIVO GENERAL...37
4.2 OBJETIVOS ESPECÍFICOS...37
5 MATERIALES Y MÉTODOS. ... 38
5.1 TIPO Y LUGAR DE ESTUDIO...38
5.2 POBLACIÓN Y MUESTREO. ...38
5.3 MICROORGANISMOS Y PLÁSMIDOS UTILIZADOS EN EL ESTUDIO.. ...38
5.4 MEDIOS DE CULTIVO...39
5.4.1 Medio Luria-Bertani - LB ...40
5.4.2 Caldo nutriente (CN) ...40
5.4.3 Medio Mineral (MM) ...40
5.4.3.1 Solución de elementos traza...40
5.4.3.2 Medio mineral con sacarosa...41
5.4.3.3 Medio mineral con propionato...41
5.4.3.4 Medio mineral con sacarosa y canamicina...41
5.4.3.5 Medio mineral con sacarosa e tetraciclina. ...41
5.4.3.6 Medio mineral con glucosa...41
5.4.3.7 Medio mineral con glucosa y canamicina...41
5.5 ANTIBIÓTICOS EMPLEADOS. ...41
5.7 PRESERVACIÓN DE CEPAS...42
5.8 REACTIVACIÓN Y VALIDACIÓN DE LA BIBLIOTECA GENÓMICA Y DE LOS MUTANTES DE B. sacchari...42
5.9 ENSAYO FENOTÍPICO DE MUTANTES. ...43
5.10 COMPLEMENTACIÓN DE MUTANTES. ...43
5.11 EXTRACCIÓN DE DNA PLASMIDIAL (MINIPREP) ...44
5.11.1 Solución de GETL (Birnborim e Doly, 1979) ...45
5.11.2 Solución de acetato de potasio (Birnboim e Doly, 1979) ...45
5.12 GEL DE AGAROSA. ...45
5.12.1 Solución TBE ...46
5.12.2 Solución TAE ...46
5.12.3 Colorante de corrida ...46
5.13 MANIPULACIÓN DEL DNA ...47
5.13.1 Digestión del DNA ...47
5.14 EXPERIMENTO DE ACUMULO DE PHA PARA EVALUAR EL EFECTO DE LAS COMPLEMENTACIONES OBTENIDAS. ...47
5.15 DETERMINACIONES ANALÍTICAS. ...48
5.15.1 Concentración celular. ...49
5.15.2 Proporción de polímero ...49
6 RESULTADOS Y DISCUSIÓN. ... 50
6.1 ENSAYO FENOTÍPICO ...50
6.2 VALIDACIÓN DE LA BIBLIOTECA GENÓMICA DE Burkholderia sacchari LFM 101. ...53
6.3 BÚSQUEDA DE CLONES QUE RESTITUYEN EL FENOTIPO prp+ EN MUTANTES AFECTADOS EN INTERMEDIARIOS DE LA ALFA OXIDACIÓN.. ...55
6.4 ENSAYO DE ACUMULO. ...59
7 CONCLUSIONES... 61
LISTA DE ABRVIATURAS Y SÍMBOLOS 2 MCC: Ciclo de 2-metilcitrato
2MCS: 2- metilcitrato sintasa 3HV: 3- hidroxivalerato Amp: Ampicilina AN: Agar Nutriente CN: Caldo Nutriente CoA: Coenzima A CS: Citrato sintasa
DNA: Ácido desoxirribonucléico
EDTA: Ácido etileno diamino tetracético IPTG: Isopropil tio-β-D-galactósidos Kan: Canamicina
Kb: Kilo bases
LB: Medio de cultivo Luria Bertani
LBK: Medio de cultivo Luria Bertani con Kanamicina (Canamicina) M.S: Masa seca
MCS: Sitio de multiclonage MM: Medio mineral
MMS: Medio mineral con sacarosa
MMPK: Medio mineral con propionato y canamicina MMSK: Medio mineral con sacarosa y canamicina MMG: Medio mineral con glucosa
MMAA: Medio mineral con ácido acrílico MMAAc: Medio mineral con ácido acético MMAL: Medio mineral con ácido láctico MMF: Medio mineral con fructosa MMPir: Medio mineral con piruvato n: Variación del grupo hidroxilo ORF: Open Reading Frame P3HB: Poli-3-hidroxibutirato pb: Pares de bases
PCR: Reacción en cadena de la polimerasa PHAs: Polihidroxialcanoatos
prp: Propionato
SDS: Dodecil sulfato de sodio
TCA: Ciclo de los ácidos tricarboxílicos Tris: Tri-hidroximetil- amoniometano U.V: Radiación ultravioleta
ÍNDICE DE LAS FIGURAS
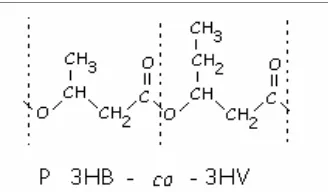
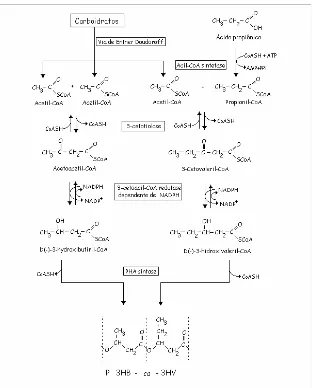
Figura 1. Estructura del copolímero de poli-3- hidroxibutirato-co-3-hidroxivalerato (P3HB-co-3HV). ... 17 Figura 2. Síntesis de P3HB-co-3HV a partir de carbohidratos y ácido propionico. 22 Figura 3. Esquema de los experimentos de acumulo de polímero, realizados a los
mutantes y mutantes complementados de la cepa B. sacchari LFM 101... 48 Figura 4. Via metabólica para la alfa-oxidación de propionato para los mutantes en estudio. ... 53 Figura 5. Gel de agarosa después de una corrida electroforética, indicando los
diferentes padrones de bandas de algunos de los 28 clones seleccionados al azar de la biblioteca genómica de B. sacchari: (1)(14) Padrón: DNA λ digerido con Hind III; (2 a 13) clones escogidos aleatoriamente y digeridos con EcoRI... 55 Figura 6. Gel de agarosa, después de una corrida electroforética, indicando los
ÍNDICE DE LAS TABLAS
Tabla 1. Cepas bacterianas y plásmidos (vectores de clonage) empleados. ... 38 Tabla 2. Antibióticos utilizados. ... 42 Tabla 3. Validación del crecimiento de los mutantes prp de B. sacchari LFM 183,
184, 185, 186 y 198 en diferentes compuestos. ... 50 Tabla 4. Validación del crecimiento de los mutantes prp de B. sacchari LFM 183,
184, 185, 186 y 198 en ácido acrílico. ... 51 Tabla 5. Nueva clasificación de los mutantes en estudio de acuerdo a los sustratos
consumidos... 52 Tabla 6. Mutantes prp de B. sacchari complementados fenotípicamente por los
fragmentos P1 y P2... 56 Tabla 7. Tamaños estimados de los subfragmentos estudiados en cada clone capaz
RESUMEN
Burkholderia sacchari es una bacteria capaza de acumular gránulos intracelulares de polihidroxialcanoatos (PHA) que son utilizados en la producción de plásticos biodegradables. A partir de la sacarosa, ésta cepa acumula el homopolímero poli-3-hidroxibutirato (P3HB). Cuando el propionato es adicionado como cosubstrato, la bacteria acumula un copolímero de 3-hidroxibutirato-co-3-hidroxivalerato (P3HB-co -3HV), con mejores características físico-químicas que el P3HB. Sin embargo, apenas un pequeño porcentaje del propionato suministrado es convertido en unidades 3HV, debido a la presencia de vías más eficientes del catabolismo; que utilizan el propionato para la generación de biomasa (sin PHA), CO2 y agua, reduciendo el porcentaje de 3HV en el copolimero. Mutantes UV previamente obtenidos, incapaces de crecer en propionato (prp), se tornaron más eficientes en la producción del copolímero, teniendo también el fenotipo alterado para el crecimiento en intermediarios de la α-oxidación de propionato. En este trabajo, fueron estudiado 5 de éstos mutantes prp de B. sacchari: LFM 183, 184, 185, 186 y 198. Posterior a la validación de una biblioteca genómica de la cepa salvaje, compuesta por cerca de 2000 clones, fueron seleccionados y caracterizados preliminarmente en gel de agarosa dos fragmentos diferentes de DNA (en los clones P1 y P2) capaces de restituir el crecimiento en propionato a todos éstos mutantes. Estos fragmentos deben contener genes envueltos en el catabolismo de propionato en B. sacchari, cuya inactivación llevó al aumentos en la eficiencia bacteriana en acumular el copolimero.
ABSTRACT
Burkholderia sacchari is a bacterial strain able to accumulate intracellular granules of polyhydroxyalkanoate (PHA), which are applied in the production of biodegradable plastics. From sucrose this strain accumulates the homopolymer poly-3-hydroxybutyrate (P3HB). When propionate is supplied as co-substrate the copolyester of 3-hydroxybutyrate-co-3-hydroxyvalerate (P3HB-co-3HV) is accumulated. However, a low percentage of the propionate supplied is converted to 3HV units, which can be attributed to the presence of more efficient catabolic pathways that convert this substrate do CO2, water and non-PHA biomass, thus reducing the 3HV content. UV mutants previously obtained were unable to grow in propionate (prp) and also showed the phenotype affected concerning grow on intermediates of propionate α-oxidation. In the present work 4 of those B. sacchari prp mutants were studied: LFM 183, 184, 185, 186 y 198. After a screening in a B. sacchari genomic library (c.a. 2000 clones), the prp+ phenotype was restored to the mutants by two different DNA fragments harbored by clones P1 and P2, which were preliminary analyzed by agarose gel electrophoresis. P1 and P2 contain genes involved in propionate catabolism and their inactivation led to bacterial improvement on P3HB-co-3HV accumulation.
1
INTRODUCCIÓN
Los Polihidroxialcanoatos (PHA) constituyen una familia de poliésteres acumulados intracelularmente por microorganismos, en forma de gránulos, que presentan importancia biotecnológica por ser empleados en la producción de plásticos, elastómeros biodegradable y biocompatibles.
Burkholderia sacchari LFM 101 es una cepa bacteriana Gram.negativa, aislada de suelo brasilero en un programa de selección de microorganismos capaces de producir PHA a partir de sacarosa, materia prima abundante en Brasil, proveniente del bagazo de caña. Entre las diferentes cepas testadas, ésta se destacó por su capacidad de rápido crecimiento en esta fuente de carbono, así como en el acumulo de altos porcentajes de PHA en su interior.
LFM 101 viene siendo estudiada bajo diversos aspectos con el objetivo de conocer su metabolismo y su potencial para la utilización biotecnológica. Recientemente, estudios taxonómicos, incluyendo laboratorios de Brasil, Alemania y Bélgica, indicaron que la cepa LFM 101 representa una nueva especie en el género Burkholderia, proponiendo así el nombre de Burkholderia sacchari.
Al ser cultivada esta nueva cepa en sacarosa, bajo una limitación de un nutriente esencial para el crecimiento como el nitrógeno, fósforo, oxígeno, etc., B. sacchari
acumula el homopolímero poli-3-hidroxibutirato (P3HB) y, al ofrecerse como co-substrato el propionato, la bacteria acumula un co-polímero de
Figura 1. Estructura del copolímero de poli-3- hidroxibutirato-co-3-hidroxivalerato (P3HB-co-3HV).
La incorporación de unidades 3HV, obtenidas por el abastecimiento de propionato como co-substrato, genera un co-polímero más flexible y con mayor variedad de aplicaciones. Dado el costo de este co-substrato y al bajo aprovechamiento del mismo por bacterias para la síntesis del co-polímero, inclusive por B. sacchari, fueron obtenidos por UV mutantes de esta bacteria más eficientes en la conversión de propionato en 3HV (mutantes prp), habiéndose aumentado entre tres y ocho veces este factor de conversión (o Y3HV/ Prop se evaluó de 0,10 para 0, 35 hasta 0,80 g/g), indicando su alto potencial para su uso industrial en procesos desarrollados por el IPT y permitido para la producción en una mayor escala. De modo general, los mutantes pueden ser clasificados en dos grandes grupos: aquellos que además de estar afectados en prp, están afectados en el uso de intermediarios de la α-oxidación de propionato y aquellos prp no afectados en el uso de estos intermediarios. Este último grupo ya fue ampliamente estudiado y engloba mutantes que pasaron a presentar Y3HV/ Prop en torno de 0,80 g/g. En cuanto al primer grupo, los valores de Y3HV/ Prop están en torno de 0,30 g/g y éstos fueron objetos de éste trabajo.
Se estudiaron los mutantes prp LFM 183, 184, 185, 186 y 198 de Burkholderia sacchari
2
REVISIÓN DE LITERATURA.
2.1 QUÉ SON LOS POLIHIDROXIALCANOATOS (PHA)?
Los polihidroxialcanoatos (PHA) están compuestos por un grupo bastante diverso de poliésteres, acumulados por un número significativo de bacterias (Steinbüchel & Valentin, 1995). Son termoplásticos, biodegradables e biocompatibles, cuya producción y aplicabilidad vienen siendo estudiadas en todo el mundo. Estas características hacen que estos materiales sean una alternativa ambientalmente viable para mudar el uso de los plásticos de origen petroquímico, dado al impacto ambiental que generan. Para las bacterias, los PHA actúan como reserva de carbono y energía (Steinbüchel & Valentin, 1995).
En Brasil, a partir de 1992 la producción de algunos tipos de PHA esta siendo estudiada, destacándose por sus propiedades, P3HB y P3HB-co-3HV, en el cual se emplean materias primas renovables por la agricultura, tales como sacarosa, melaza e hidrolizado de bagazo de caña, entre otros (Bueno Netto et al., 1993, Gomez & Bueno Netto, 1997; Silva, 2000; Gomez & Bueno Netto, 2001, Silva et al., 2002). Uno de los resultados más importantes de este desarrollo en el país es la producción en escala piloto, que se inició empleándose derivados da caña de azúcar (Nonato et al., 2001). Otro resultado importante fue el aislamiento e identificación de la nueva especie bacteriana Gram. negativa Burkholderia sacchari LFM 101 (Brämer et al., 2001) más eficiente y, recientemente, mejorada para la incorporación de unidades 3HV a partir del uso del propionato, así como su capacidad de usar xilosa e hidrolizado de bagazo de cana-de-azúcar (Silva et al., 2002).
2.2 CARACTERÍSTICA DE Burkholderia sacchari LFM 101.
Colonias de la cepa LFM 101 crecen sobre medio caldo o agar nutriente (CN), presentando color blanco, siendo opacas debido a la acumulación de P3HB y P3HB-
co-HV. Las células son cocobacilos Gram. negativos (0,5 – 0,8 µm de ancho, 1,5 – 3,0 µm de largo) y lisadas con KOH al 3%, móviles debido a la presencia de flagelos polares y con temperatura óptima de crecimiento entre 25 – 37 ºC, y con un contenido de 63,7 mol % de G + C. No formador de esporas, oxidasa y catalasa positiva, así como reductor de nitrato a nitrito, presentando ausencia de licuefacción de gelatina e hidrólisis de esculina. (Silva et al., 2000)
Los sustratos asimilados por la cepa son los siguientes : acetato, N-acetilglucosamina, adipate, adonitol, DL-arabinosa, D-arabitol, citrato, D-fructosa, DL-fucosa, galactosa, gluconato, D-glucosa, glicerol, inositol, 2-cetogluconato, lactato, D-lixose, malato, manitol, D-manosa, fenilacetato, propionato, piruvato, D-rafinosa, sorbitol, sacarosa and
D-xilosa. Las siguientes Fuentes de carbono no son usadas: almidón, amigdalin, L -arabitol, arbutin, caprate, celobiosa, dulcitol, eritritol, metil α-D-glucósido, metil α-D -manósido, β gentiobiosa, glicógeno, inulina, 5-cetogluconato, lactosa, maltosa, melecitosa, melobiosa, metil β-xilósido, ramnosa, ribosa, salicina, L-sorbosa, D -tagatosa, trehalosa, D-turanosa, xilitol y L-xilose. Los siguientes carbohidratos son oxidados por la cepa: adonitol, DL-arabinosa, D-arabitol, D-fructosa, DL-fucosa, galactosa, gluconato, D-glucosa, glicerol, inositol, 2-cetogluconato, D-lixosa, manitol,
D-manosa, D-rafinosa, ribosa, sorbitol, sacarosa and D-xilosa. Los siguientes carbohidratos no son oxidados: N-acetilglucosamina, almidón, amigdalina, L-arabitol, arbutina, celobiosa, dulcitol, eritritol, β-gentiobiosa, glucógeno, inulina, 5- cetogluconato, lactosa, maltosa, melecitosa, melibiosa, metil α-D-glucósido, metil α-D -manósido, metil β-xilosido, ramnosa, salicina, L-sorbosa, D-tagatosa, trehalosa, D -turanosa, xilitol y L-xilose. (Brämer et al.,2001).
Las cepa LFM 101 es susceptible a los antibióticos tetraciclina (100 µg mL-1), canamicina (50 µg mL-1), cloranfenicol (100 µg mL-1) e ampicilina (15 µg mL-1) (Silva
2.3 SÍNTESIS DE PHA EN BACTERIAS.
En diversas bacterias, a partir del acetil-CoA obtenido del catabolismo de carbohidratos, por ejemplo, la síntesis del homopolímero 3HB ocurre envolviendo la acción de tres enzimas (Oeding & Schlegel, 1973; Senior & Dawes, 1973). Primero, por acción de la 3-cetotiolasa, dos moléculas de acetil-CoA se condensan, generando acetoacetil-CoA. Esta, al seguir, sufre una reducción, formando 3-hidroxibutiril-CoA por acción de la acetoacetil-CoA reductasa. La PHA sintasa, finalmente, incorpora cada molécula de 3-hidroxibutiril-CoA a la cadena polimérica en crecimiento.
En la presencia de un co-substrato, como el propionato, por ejemplo, una molécula de propionil-CoA podrá entonces condensarse como una de acetil-CoA y, siguiendo las etapas posteriores, formar 3-hidroxivaleril-CoA para finalmente ser incorporada por la PHA sintasa para formar el co-polímero P3HB-co-3HV (Figura 2).
relativamente caro para la producción de PHA, habiendo sido desarrollados programas de obtención de mutantes más eficientes en la canalización de propionato a unidades 3HV por parte de empresas productoras (Byrom, 1990). Para la obtención de mutantes más eficientes en la conversión de propionato a unidades 3HV, es de fundamental importancia el conocimiento de las vías metabólicas envueltas en el catabolismo de este substrato y que pueden estar actuando de modo competitivo con la vía de síntesis de P3HB-co-3HV.
2.4 UTILIZACIÓN DE SUBSTRATOS DE BAJO COSTO EN LA
PRODUCCIÓN DE PHA
Según Choi y Lee, el mayor obstáculo en la comercialización de PHA es su alto costo, aproximadamente el 40% del total del costo de producción de PHA son debidos a los costos de los substratos. Así, para conseguir reducir los costos de producción es necesaria la utilización de fuentes de carbono más baratas
Gomez e colaboradores (1996), validaron la eficiencia de producción de PHA por diversas bacterias Gram. negativas, aisladas del suelo de plantaciones de caña de azúcar. Los medios de cultivo fueron realizados con limitación de nitrógeno y utilizando como carbohidratos sacarosa, glucosa y fructuosa. Cuando se utilizó la sacarosa, algunos microorganismos mostraron valores de rendimiento global de carbohidrato en producto, YP/C, mayores que cuando se utilizó glucosa y fructosa. Los autores explicaron el hecho, afirmando que la misma masa de sacarosa genera 5% más acetil-CoA y, por consiguiente PHA, que glucosa y fructuosa. Entre los microorganismos aislados,
Burkholderia sp LFM 101 fue capaz de acumular 75% de su peso seco en P3HB a partir de glucosa más fructosa y 69% a partir de sacarosa, con una eficiencia mayor que 80% del máximo rendimiento teórico, que es de 0.48g/g cuando se utiliza glucosa y/o fructosa, de acuerdo con Yamane (1993), y 5% mayor, de 0.5g/g cuando se usa sacarosa. La bacteria todavía fue capaz de crecer con velocidad específica de crecimiento (0.4-0.45h-1), mayor que R. eutropha (0.3 h-1)
Según Wong y Lee 1998, el suero de leche proveniente de la industria de lacticíneos, resultante de la fabricación de queso o caseína, representa cerca del 80 – 90% del volumen de leche transformado. Una vez que el suero de leche es rico en nutrientes, siendo la lactosa su mayor constituyente, se presenta como otra alternativa de substrato de bajo costo en la producción de PHA.
Otros estudios se han ido realizando con el fin de reducir los costos de producción de PHA, usando aceites vegetales, los cuales son productos agrícolas renovables y de bajo costo. Fukui y Doi (1998), estudiaron la capacidad de R. eutropha H16 de crecer y producir PHA a partir de aceites vegetales como aceite de oliva, maíz, dende y ácido oleico como únicas fuentes de carbono. Los resultados mostraron que la bacteria crece muy bien en dichos substratos, acumulando altas cantidades de P3HB, entre 79 – 82% de su peso seco.
2.5 PRODUCCIÓN DEL CO-POLÍMERO P3HB-co-3HV
Una vez que el P3HB es un material rígido y quebradizo, la incorporación de unidades 3HV es responsable por la mejoría de las características del polímero P 3HB-co-3HV. Como resultado de la incorporación de unidades 3HV, ocurren alteraciones de parámetros, como niveles de cristalinidad y punto de fusión, acarreando en la disminución de la rigidez del polímero y en el aumento de la resistencia al impacto. Cantidades de 3HV entre 17 y 20 mol% permiten la obtención de un producto con propiedades termoplásticas, que llevan a una mayor aplicación del polímero (Holmes, 1985). A partir de fuentes de carbono como glucosa y fructuosa, Ralstonia eutropha
produce a penas homopolímero 3HB. Para la producción del co-polímero, son necesarios precursores como ácido propiónico o valérico, que pueden ser metabolizados a unidades 3HV (Byrom, 1990).
2.5.1 UTILIZACIÓN DE ÁCIDO PROPIÓNICO COMO PRECURSOR DE UNIDADES 3HV.
La producción industrial del co-polímero P(3HB-co-3HV) por R. eutropha, es realizada adicionando ácido propiónico al cultivo para la incorporación de unidades 3HV. El valor de 3HV incorporado al polímero es controlado por la variación de la ración de glucosa y acido propiónico de la alimentación. Es necesario un control apropiado de la concentración de propionato adicionado pues, si se presenta en el medio en concentraciones mayores que 0.1%, puede ser toxico para el microorganismo el inhibir la síntesis del polímero (Byrom, 1990).
también puede ser asimilado para el crecimiento celular (Doi, 1987) El acido propiónico es rápido y eficientemente degradado por las células bacterianas, y su uso, durante la fase de multiplicación celular, implica el mayor desperdicio de este substrato, impidiendo que cumpla su principal papel en este proceso, o sea, la síntesis de unidades 3HV (Byrom, 1990).
La baja cantidad teórica de P(3HB-co-3HV) y la productividad son atribuidas al hecho tóxico del co-substrato, las estrategias inadecuadas de alimentación de ácido propiónico a los cultivos, y al hecho de que la mayoría de los microorganismos presentan bajo factor de conversión de ácido propiónico en unidades 3HV (Silva, 2000).
Marangoni (2000), estudió diferentes estrategias de alimentación de ácido propiónico en diferentes substratos en la producción de P(3HB-co-3HV) como R. eutropha DSM 545, en cultivo alimentado. Cuando utilizó suero de leche, las estrategias de alimentación de ácido propiónico por pulsos obtuvo baja productividad, sin embargo, con grandes fracciones de 3HV, cerca de 38%, con conversión de propionato en 3HV de 0.22 molHV/mol ac prop, sugiriendo la posibilidad de la existencia de precursores 3HV en el suero. Con azúcar invertido, se utilizó alimentación con ácido propiónico, con regulación de pH, en pulsos de 1g/L y de forma continua, siendo que las fracciones molares de 3HV obtenidas fueron de 17.5 y 26 mol% 3HV, respectivamente. En cuanto a los factores de conversión fue obtenido 0.03 molHV/mol ac prop para las estrategias de regulación de pH y pulsos, y 0.13 molHV/mol ac prop para la alimentación continua, manteniendo la concentración de ácido propiónico en 0.4g/L en el medio.
2.6 CATABOLISMO DE PROPIONATO EN DIFERENTES ORGANISMOS.
como resultado de la fermentación de carbohidratos por diversos organismos y, junto con acetato, constituyen la fuente de energía para estos animales. Es también producido y consumido en ambientes metanogénicos (Stams et al., 1998). El Ácido propiónico también puede ser producido por fermentación del almidón o azúcar por
Propionibcterium acidipropionici (Carrondo et al., 1988, Blanc & Goma, 1989, Raecker
et al., 1992) y puede ser utilizado para fragancias y agentes flavorizantes (Samel et al., 1993), además de tener un papel en la producción de quesos.
En bacterias, el catabolismo de propionato viene siendo estudiado por ser el último compuesto oriundo de la degradación de ácidos grasos de cadena impar a través de la α -oxidación, teniendo una estructura que no es blanco de las reacciones que a él dieran origen. Recientemente, el uso de propionato, junto con azúcares, adquirió importancia en la obtención de polihidroxialcanoatos (PHA), polímeros biodegradables, como P3HB-co-3HV, justamente para introducir la fracción 3HV y mejorar las propiedades del polímero (Steinbüchel, 1991). Algunos microorganismos, mostraron ser capaces de sintetizar polímero con unidades 3HV a partir de substrato no relacionado estructuralmente a este monómero. Este hecho despertó el interés por el estudio de vías capaces de formar propionil-CoA para la síntesis de unidades 3HV en Nocardia coralina, Rhodococcus ruber y otros organismos relacionados (Valentin & Dennis, 1996). Recientemente, Aldor et al. (2002) desarrollaron un sistema de expresión, en los cuales están envueltos los genes del ciclo de 2-metil citrato (prpC) y los genes de formación de propionil-CoA en E. coli (sbm e ygfG) y genes de biosíntesis de PHA de
Acinetobacter (phaBCA), que permitió obtener una cepa recombinante de Salmonella entérica capaz de producir 3HV sin la formación exógena de propionato. Entretanto, se trata de producir propionil-CoA a partir de succinato, que es la vía con reacciones que se procesan en el sentido inverso a las del catabolismo de propionato en humanos y otros eucariontes.
comprobar la presencia de pocas vías, en algunos microorganismos, principalmente en actinomicetos, sin embargo, diversas vías propuestas anteriormente todavía requieren ser comprobadas. En el momento, la literatura revisada aporta un grupo en la Universidad de Madison (Horswill & Semerena, 1997; Horswill & Escalante-Semerena, 1999; Horswill & Escalante-Escalante-Semerena, 2001) estudiando el catabolismo de propionato en Salmonella en los últimos 8 años, seguido del Instituto de Microbiología de Münster, que inició en este tema con el estudio realizado por la propia proponente en aquella institución, extendiéndolo a Ralstonia eutropha
Independiente de la vía propuesta para el catabolismo del propionato, la activación a propionil-CoA es la primera etapa descrita, la reacción está inicialmente atribuida a las dos rutas de síntesis de acetil-CoA (Rhie & Dennis, 1995 a,b), una catalizada por la acetil-CoA sintetasa y otra, por la acetato quinasa y transacetilasa. Kumari et al. (1995) concluyeron que las células de E. coli requieren ambas vías de activación de acetato. Recientemente, fueron publicadas evidencias del papel del gene prpE (presente en el operon prpBCDE) en la síntesis de propionil-CoA sintetasa (prpE) en Salmonella typhimurium (Horswill & Escalante-Semerena, 1999).
1998, Brämer et al., 2002), las bases genéticas están bien establecidas. En estas bacterias, los genes de 2MCC están organizados en un operonprpBCD .
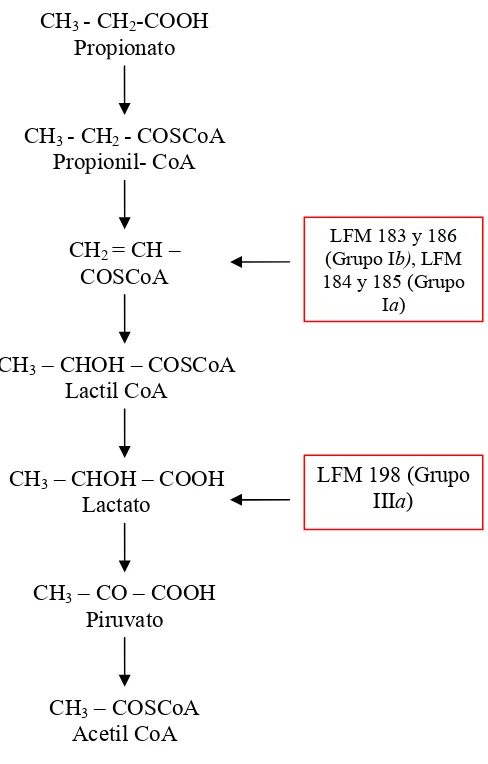
Estudios fisiológicos y moleculares de mutantes prp de B. sacchari sugieren que la baja conversión de propionato a 3HV por la vía de síntesis del polímero, resulta de la existencia de por lo menos dos vía de catabolismo que compiten, y son más eficientes, por este substrato: la α-oxidación y el ciclo de 2metilcitrato (2MCC) (Silva et al., 2000, Gregorio et al., 2001). Por lo tanto, la propuesta de la existencia de esta primera vía fue basada apenas en evidencias fenotípicas de mutantes estudiados. Así, la existencia del ciclo de 2MCC fue recientemente comprobada en B. sacchari por estudios moleculares (Brämer et al., 2002).
Hasta ahora diversas vías del catabolismo de propionato han sido propuestas en la literatura, la 2MCC es la única comprobada en el nivel molecular, habiendo sido descrita en bacterias al final de la década pasada (Textor et al., 1997).
por la 2-metilisocitrato liasa (PrpB.). Resultados recién publicados comprueban la existencia del 2MCC en B. sacchari LFM101 (Brämer et al., 2002) que serán comentados a seguir.
El uso del propionato viene siendo estudiado en B. sacchari LFM 101, demostrado que, de modo similar a otras bacterias, la conversión de propionato en unidades 3HV es restricta a menos que 10% del máximo teórico (Silva et al., 2000; Brämer et al., 2001). Mutantes de B. sacchari, incapaces de crecer en propionato (prp), que acumulan unidades 3HV a partir de este sustrato, fueron obtenidos, clasificados y comparados, buscando mejorar el rendimiento obtenido (Silva et al., 2000). Fueron definidos dos grupos de mutantes UV: un grupo que se encuentra afectado en la utilización de intermediarios de la α-oxidación y otro no afectado. En cuanto a los mutantes del primer grupo alcanzaban al máximo, valores del factor de conversión del propionato a unidades 3HV (Y3HV/Prop) de 0,37 g/g, mutantes no afectados en esta vía alcanzaban hasta 0,80 g/g. La suma de estos valores corresponde a 1,17 g/g, bastante próximo del rendimiento máximo teórico de 1,35 g/g. Fue propuesto entonces que el ácido propiónico sería consumido por lo menos tres vías: dos vías de degradación (cada una bloqueada en cada grupo de mutantes), las cuales competirían todavía con una tercera vía, la de síntesis de 3HV. El mutante LFM 189, no afectado en la α-oxidación, continua siendo estudiado por ser aquel que presentó el mejor rendimiento (Y3HV/Prp). Mutantes con inativación sitio específica de genes del 2MCC fueron recientemente estudiados, cuyos resultados corroboran la hipótesis de que existen dos vías para el catabolismo de propionato en B. sacchari (Pereira, 2007).
sintasa y los dos últimos genes codifican productos implicados en la conversión de 2-metilcitrato a 2-metilisocitrato (Brämer et al., 2002).
Partiendo de la cepa salvaje, fue clonado un fragmento genómico Sal 1 (S1) de 1 Kb, capaz de restituir el fenotipo prp al mutante LFM 189 (Silva, 1998). Sin embargo, S1 no fue capaz de complementar otros 4 (LFM 192, 193, 195 y 196) de los 6 mutantes del mismo grupo de LFM 189, complementando apenas LFM 197, también de LFM 189. A partir de resultados del secuenciamiento de S1 y regiones adyacentes, Brämer et al. (2002) demostraron, del punto de vista molecular, la presencia del 2MCC en B. sacchari. El operon prp descrito está compuesto de 4 genes (prpB, prpC, acnM y a
open reading frame -ORF5) que muestran identidad con los genes de los loci prp de muchas bacterias. El gen prpC codifica la 2-metilcitrato sintasa, prpB codifica la 2-metilisocitrato liasa, y los genes acnM y ORF5 probablemente son requeridos para la conversión de ácido 2-metilcítrico en ácido 2-metilisocítrico. Otro gene (prpR) posicionado uspstream al operon, con una orientación invertida, codifica un probable regulador transicional del operonprp. Tres ORFs adicionales (6, 7, 8), cuyas funciones son desconocidas, fueron localizados downstream a la ORF5 en el locus prp de B. sacchari y no fueron encontradas en ningún otro operon. El gene prpD, probablemente envuelto en la reacción de isomerización en el 2MCC en S. enterica, no fue localizado en el locus prp de B. sacchari. Si, en B. sacchari, los productos de los genes acnM y ORF 5 estuvieren también envueltos en la conversión de 2-metilcitrato a 2 metilisocitrato, como ocurre en R. eutropha HFR39 (Brämer & Steinbüchel, 2001), los autores infieren que el producto del gen prpD no es requerido para la funcionalidad de este ciclo en esta bacteria. Considerando que los productos das ORFs 6, 7, 8 pueden no ser necesarios para la funcionalidad de la 2-metilcitrato sintasa en B. sacchari, fue propuesto un conjunto mínimo de genes estructurales para el locus prp de esta cepa, responsables por la funcionalidad del ciclo de 2-metilcitrato.
En lo que se refiere a los mutantes afectados en la utilización de intermediarios da α -oxidación, se especula que ésta posible vía podría asumir un papel más importante bajo condiciones de crecimiento celular, y no de acumulo, una vez que el 90% de los mutantes prp obtenidos en estudios anteriores, presentarán este fenotipo (Silva et al., 2000). Más allá de estas características fenotípicas y de la alteración característica de los valores de Y3HV/Prop, se sabe que muchos de estos mutantes no fueron complementados ni por E2 ni por S1, fragmentos de DNA genómico de B. sacchari portadores de regiones de genes de 2MCC, ni por otras regiones capaces de complementar otros mutantes prp del grupo IV (Cintra et al., 2005). Otro dato a ser considerado es que los resultados de eficiencia de conversión de propionato a unidades 3HV de los mutantes afectados en 2MCC y en la α-oxidación indican que la deleción de ambas vías podría resultar en la generación de mutantes presentando valores próximos al máximo teórico de Y3HV/Prop (Silva et al., 1996, Silva et al., 2000).
A pesar de estos avances, hay todavía lagunas a ser llenadas en cuanto a la presencia de otras vías en B. sacchari, ya que todavía no fueron estudiados los mutantes afectados en el uso de intermediarios de la α-oxidación aquí propuesto. El conocimiento de las vías actuantes y su posible deleción, por modificaciones genéticas, podrían generar mutantes con eficiencia de conversión de propionato a 3HV más próximas del valor máximo teórico de Y3HV/Prop = 1,35 g/g.
2.6.1 UTILIZACIÓN DE MICROORGANISMOS RECOMBINANTES
INCAPACEZ DE CRECER EN ÁCIDO PROPIÓNICO.
El ácido propiónico es adicionado como co-substrato precursor de unidades de 3HV en la producción del co-polímero, sin embargo, solamente una pequeña fracción de éste es incorporada de esta forma al polímero, siendo que el restante es desviado para otras rutas metabólicas, como crecimiento celular y formación de unidades 3HB. (Squio & Falcão, 2003).
Lee et al, desenvolvieron a partir de A.eutrophus NCIMB 11599, una cepa mutante de A.eutrophus BK-23, que es incapaz de asimilar el acido propiónico para el crecimiento celular. Las dos cepas fueron comparadas en cuanto a la habilidad de acumular P(3HB-co-3HV) y a la composición del polímero producido. Los experimentos fueron realizado en cultivo alimentado, utilizando la ración de alimentación P/G de 0.21 y limitación en nitrógeno. Niveles finales similares de contenido de células y del co-polímero fueron obtenidos para los cultivos, entretanto, se observó que para la cepa mutante, una fracción de unidades 3HV dos veces mayor (22 mol%), en cuanto para la cepa original fue menor que 10 mol%. La tasa de conversión de ácido propiónico en unidades 3HV durante la fase de acumulo fue de 0.4 gHV/g ac proppara el mutante, contra 0.1 gHV/g ac
prop para la cepa original. Así, los autores relatan que con esa cepa se torna posible producir P(3HB-co-3HV) con mayor fracción de unidades 3HV, usando menos ácido propiónico.
En estudios semejantes, Sartori (1998) obtuvo una cepa mutantes A. eutrophus UV1, derivada de R. eutropha DSM 545, incapaz de crecer en propionato, visando su mejor aprovechamiento para la biosíntesis de las unidades 3HV del polímero. La cepa mutante UV1 acumuló 37 mol% de 3HV, en cuanto a la cepa original acumuló a penas 8.6 mol% y se mostró capaz de convertir 60.7% de todo el propionato suministrado a unidades 3HV, siendo que, en las mismas condiciones, la cepa original sólo convirtió 18.5%. Más allá de eso, la cepa mutante todavía mantiene, o hasta supera, la capacidad de producción de polímero total de la cepa original. De esta forma, el nuevo mutante es presentado como un óptimo potencial de aprovechamiento industrial.
Otro microorganismo que está siendo estudiado para la producción de PHA´s es
Burkholderia sp. Burkholderia sp. LFM 101 es presentada como una buena productora del polímero, entre tanto, la conversión de acido propiónico en unidades 3HV es restricta a menos de 10% del rendimiento máximo teórico, que es de 1.35g 3HV/g ácido propiónico de acuerdo con Gómez et al (1996). La intención de aumentar esta eficiencia de conversión y utilización del ácido propiónico, Silva et al, desenvolvieron mutantes de
original fueron comparadas creciendo en un biorreactor con raciones de ácido propiónico y sacarosa (P/S) de 0.44 para la cepa original y de 0.09 para el mutante, ya que era esperado que el mutante tuviese mejor utilización del ácido propiónico. Con el mutante LFM 189 obtuvieron una alta tasa de conversión del ácido propiónico en unidades de 3HV, de 1.2 gHV/g ac.prop, en cuanto para la cepa original obtuvieron apenas
0.05 gHV/g ac.prop. Entre tanto, las células acumularon poco polímero, siendo cerca del 50%
3
JUSTIFICACIÓN
La mayoría de los plásticos producidos durante décadas son de origen petroquímico, es decir, de fuentes naturales no renovables, que por el acelerado crecimiento urbano y la necesidad de consumo han ido agotando rápidamente dicha materia prima. Esto ha generando en la población y especialmente en los investigadores un interés particular por hallar soluciones rápidas y concretas de fuentes naturales y por ende, de organismos capaces de solucionar el problema ambiental que están generando los plásticos producidos a partir de petróleo, ya que la degradación de éstos se lleva a cabo en un tiempo mayor a 100 años (Duarte, 2004).
Por lo anteriormente mencionado, en la actualidad, se están desarrollando diversas investigaciones que conllevan a la producción de plástico a partir de materias primas renovables, que minimizan el consumo de petróleo y por ser fabricadas a partir de fuentes naturales, son fácilmente degradadas y producidas.
Debido a los altos costos de éste plástico, a la participación mínima en el mercado internacional y a las dificultades de procesamiento, se están buscando cepas altamente eficientes en la conversión de sustratos en el producto deseado, así como en el desarrollo de procesos que permitan explorar al máximo el potencial de dichas cepas y del desenvolvimiento de procesos extracción-purificación, de forma que los costos sean reducidos drásticamente. (Franchetti, 2006)
Según antecedentes bibliográficos, se están desarrollando metodologías que investigan la utilización de sacarosa con propionato como fuente de carbono, para obtener un plástico mucho más maleable y de fácil degradación por diferentes organismos, que conllevan a la disminución del impacto ambiental. Por ende es de gran importancia restablecer el fenotipo de los mutantes LFM 183, 184, 185, 186 y 198, conocer los genes afectados, restituyendo la expresión de los intermediarios de la vía de la α-oxidación, con el fin de alcanzar un rendimiento teórico cercano al de la cepa salvaje, es decir, de 0,10 g/g.
Una vez obtenido el fragmento que restituyó el fenotipo salvaje a éstos mutantes, debe ser validada la complementación integral por medio de experimentos en frascos agitados. Si esto se lleva a cabo, quiere decir que los fragmentos contienen genes relacionados en el catabolismo de propionato en éstos mutantes afectados en el uso de intermediarios de la α-oxidación. Siendo así, los fragmentos de DNA con esta propiedad constituyen una importante herramienta para investigar en trabajos posteriores si existe la vía en cuestión e propones nuevos experimentos para mejorar aún más la formación de 3HV.
4
OBJETIVOS
4.1 OBJETIVO GENERAL
El objetivo de este trabajo fue validar mutantes de B. sacchari incapaces de crecer en propionato como única fuente de carbono (prp), buscando en una biblioteca genómica, construida a partir de la cepa salvaje, fragmentos de DNA capaces de restituir el fenotipo prp+ a los mutantes afectados en el uso de intermediarios de la alfa-oxidación de propionato.
4.2 OBJETIVOS ESPECÍFICOS
] Analizar una biblioteca genómica de B. sacchari salvaje, para seleccionar clones conteniendo fragmentos de DNA originales, capaces de restituir la capacidad de crecimiento en propionato a los mutantes en estudio.
] Corroborar la agrupación de los mutantes LFM 183, 184, 185, 186 y 198, por medio de repeticiones ensayos fenotípicos, como se encuentra reportado en la literatura.
] Evidenciar en cuál de los posibles intermediarios de la α – oxidación se encuentran afectados los mutantes en estudio.
5
MATERIALES Y MÉTODOS.
5.1 TIPO Y LUGAR DE ESTUDIO.
El presente trabajo de Grado fue de carácter investigativo, el cual se realizó en los laboratorios de Fisiología de Microorganismo, en el Instituto de Ciencias Biomédicas II de la Universidad de Sao Paulo ubicada en Sao Paulo (Brasil).
5.2 POBLACIÓN Y MUESTREO.
Las muestras fueron manejadas en la cámara de flujo laminar, en condiciones controladas, de acuerdo con la reglamentación empleada en el laboratorio de Fisiología de Microorganismos, ubicado en el Instituto de Ciencias Biomédicas de la Universidad de São Paulo.
5.3 MICROORGANISMOS Y PLÁSMIDOS UTILIZADOS EN EL ESTUDIO.
[image:39.612.100.541.473.687.2]Los microorganismos y plásmidos (vectores de clonage) utilizados en el estudio fueron las cepas enunciadas en la Tabla 1, donde se enuncian las características principales de las mismas. Las cepas anteriormente mencionadas fueron suministradas por el laboratorio de Fisiología de Microorganismos, del Instituto de Ciencias Biomédicas de la Universidad de Sao Paulo, en forma liofilizada y/o preservada en glicerol al 10%.
Tabla 1. Cepas bacterianas y plásmidos (vectores de clonage) empleados.
CEPA CARACTERÍSTICAS ORIGEN
Burkholderia sacchari
LFM 101 Sac+, PHA+, prp+, Kans, Amps, Tcs, acet+ lac+ pir+ Gómez et al.,
1996
Burkholderia sacchari LFM 183
Sac+, PHA+, prp-, Kans, Amps, Tcs, acet- lac -pir -
Silva et al., 2000
Burkholderia sacchari LFM 184
Sac+, PHA+, prp-, Kans, Amps, Tcs, acet- lac? pir
LFM 185
Burkholderia sacchari LFM 186
Burkholderia sacchari
LFM 198 Sac
+
PHA+ acet- prp- lac+ pir- Kans Amps Tcs
Escherichia coli S17-1 PHA
-, Tcs, recA, genes tra do plásmido RP4 integrado al DNA genómico
Simon et al., 1983
pVK100 Cósmido 23,0Kb Tcr, Kmr Nester,1982Knauf e
pVK100::P1 Cósmido 23,0Kb Tcr, Kmr, abrigando inserto en estudio Este trabajo pVK100::P2 Cósmido 23,0Kb Tcr, Kmr, abrigando inserto en estudio Este trabajo
LFM- Laboratorio de Fisiología de Microorganismos del ICB-USP; Sac+- crecimiento en sacarosa; PHA+- acumulo de PHA; acet- -ausencia de crecimiento en acetato; prp- - ausencia de crecimiento en propionato; lac-- ausencia de crecimiento en lactato; pir-- ausencia de crecimiento en piruvato; Kans – sensibilidad a canamicina; Amps – sensibilidad a ampicilina; Tcs – sensibilidad à tetraciclina.
Cada una de las cepas que fueron halladas en el presente trabajo fueron preservadas en glicerol al 10% y mantenidas en un freezer con temperatura de -80ºC.
5.4 MEDIOS DE CULTIVO
Los medios de cultivo empleados en cada uno de los ensayos, fueron esterilizados en autoclave por 20 minutos (min) a 121ºC, a 1 atmósfera de presión (atm).
Al preparar los diferentes medios minerales que contienen glucosa, sacarosa, propionato, entre otros, la fuente de carbono fue esterilizada por separado y adicionada después a la esterilización de las soluciones de sales.
En el preparo de medios minerales sólidos, la solución de sales minerales y de agar fueron esterilizados separadamente de la fuente de carbono.
5.4.1 Medio Luria-Bertani - LB
(Sambrook et al., 1989).
• Triptona 10g
• Extracto de levadura 5g
• NaCl 10g
• H2O 1000mL
5.4.2 Caldo nutriente (CN)
• Extracto de carne 3g
• Peptona 5g
• H2O 1000mL
5.4.3 Medio Mineral (MM)
(MM-Ramsay et al., 1990)
• Na2HPO4 3,5g
• KH2PO4 1,5g
• NH4SO4 1,0g
• Solución de elementos traza 1,0mL • Citrato férrico 6% 1,0mL • CaCl2. 2H2O 1% 1,0mL
• MgSO4. 7H2O 20% 1,0mL 5.4.3.1 Solución de elementos traza
• H3BO3 0,3g
• MnCl2.4H2O 30,0mg
• NaMoO4.2H2O 30,0mg
• NiCl2.6H2O 20,0mg
• CuSO4.5H2O 10,0mg
• H2O 1000mL 5.4.3.2 Medio mineral con sacarosa.
Medio mineral descrito en el ítem 5.3.3 conteniendo 1g/L de sacarosa.
5.4.3.3 Medio mineral con propionato.
Medio mineral descrito en el ítem 5.3.3 conteniendo 1g/L de propionato.
5.4.3.4 Medio mineral con sacarosa y canamicina.
Medio mineral descrito en el ítem 5.3.3 conteniendo 1g/L de sacarosa e canamicina (100μL/mL).
5.4.3.5 Medio mineral con sacarosa e tetraciclina.
Medio mineral descrito en el ítem 5.3.3 conteniendo 1g/L de sacarosa e tetraciclina (12,5μL/mL).
5.4.3.6 Medio mineral con glucosa
Medio mineral descrito en el ítem 5.3.3 conteniendo 1g/L de glucosa.
5.4.3.7 Medio mineral con glucosa y canamicina
Medio mineral descrito en el ítem 5.3.3 conteniendo 1g/L de glucosa y canamicina (100 g/mL).
5.5 ANTIBIÓTICOS EMPLEADOS.
Tabla 2. Antibióticos utilizados.
5.6 CONDICIONES DE CULTIVO
Las cepas de E. coli fueron cultivadas en medio Luria Bertani (LB) (Sambrook et al., 1989) e incubadas a 37 ºC durante 18 – 24 horas. Los antibióticos fueron adicionados al medio conforme al ítem 5.4.
Burkholderia sacchari LFM 101 y los mutantes obtenidos de ella, con los cuales se realizó el presente estudio, fueron cultivados en caldo nutriente (CN), medio LB, o medio mineral (MM), con la respectiva fuente de carbono, por 24 horas durante 30 ºC a 200 rpm, conteniendo antibióticos adecuados, cuando fuese necesario.
5.7 PRESERVACIÓN DE CEPAS
Todas las cepas bacterianas empleadas en la investigación, fueron preservadas y almacenadas a través de dos métodos (Silva et al., 1992): a) Criopreservación en glicerol
al 10% (v/v), almacenando a – 80 ºC y b) Liofilización en solución de leche al 10 % y glutamato de sodio al 5 %, siendo almacenadas finalmente en refrigerador doméstica.
5.8 REACTIVACIÓN Y VALIDACIÓN DE LA BIBLIOTECA GENÓMICA Y DE LOS MUTANTES DE B. sacchari.
La biblioteca genómica existente, creada por Cintra e colaboradores (2007) fue reactivada en medio LB con kanamicina (LBK), la cual se construyó con el DNA genómico de la cepa salvaje (cepa B. sacchari LFM 101), adecuadamente digerido, en fracciones específicas del mismo, ligándolos posteriormente al cósmido pVK100 (Knauf & Nester, 1982), para luego ser empacotado en capsides del fago y transducido para E. coli S17-1 (Simon et al., 1983). Como una alternativa, el vector con inserto (s), se introdujo en esta bacteria por transformación. Los clones se seleccionaron en medio Luria
Antibiótico Solución stok (mg/mL) Concentración final (μg/mL)
Ampicilina (Amp) sal sódica 100 (en H2O) 100
Se realizó la recuperación de la biblioteca genómica construida en estudios anteriores, así como de los mutantes liofilizados o criopreservados, de acuerdo a la cepa bacteriana, se realizó la semeadura en un medio LBK y LB, los cuales aseguraban un buen crecimiento y permitiesen validar la viabilidad de las cepas, así como para verificar que todos los mutantes a lo largo de su almacenamiento mantuvieran las características fenotípicas (perfil de utilización de intermediarios de la α-oxidación, así como de propionato) anteriormente observadas, así como la ausencia de contaminantes con la que fueron preservados.
Así mismo, fue realizada una validación de la biblioteca genómica por medio de un test de eficiencia, con el fin de verificar si la biblioteca construida poseía fragmentos de DNA genómico de la bacteria (B. sacchari) conteniendo un numero suficiente de clones para ser representativo de todo el genoma bacteriano, y así mismo evidenciar si el cósmido pVK100 había sido empacotado con éxito, conteniendo insertos y segundo, si éstos fragmentos de DNA eran iguales, o si por el contrario el tamaño de las bandas era diferente y aseguraba una buena representación del genoma de Burkholderia. Para lo cual, 28 clones de la biblioteca fueron escogidos aleatoriamente para extraer su DNA plasmidial y realizar la digestión con endonucleasas de restricción Eco RI.
5.9 ENSAYO FENOTÍPICO DE MUTANTES.
Al tener todos los mutantes reactivados y conferir que no tuviesen contaminantes, se procedió a confirmar las características fenotípicas de cada uno de ellos, como fue realizado anteriormente (Silva, 1998). Cada uno de los mutantes fue semeado en medio mineral sólido, con diferentes fuentes de carbono, con concentración final de 1g/L como sacarosa, fructosa, glucosa, propionato, lactato, acetato y piruvato como se encuentra descrito en el ítem 5.4.3, y el medio con ácido acrílico con concentraciones finales de 1 g/L, 0,5 g/L y 0,25 g/L para posterior incubación a 30 ºC por un periodo de tiempo indeterminado, es decir, hasta observar crecimiento de alguna de las cepas.
5.10 COMPLEMENTACIÓN DE MUTANTES.
186 y 198, fueron seleccionados por conjugación, empleándose medio mineral sólido, conteniendo propionato (1g/L) y canamicina (MMPK). En una placa de medio mineral conteniendo sacarosa y kanamicina (MMSK) y otra conteniendo propionato y kanamicina (MMPK), fueron semeados con asa de Drigalski 200 μL de una cultura del mutante, después de 5 minutos, fueron estriados los clones de la biblioteca genómica. Posterior a la conjugación, los clones que presentaron un crecimiento significativo en medio MMPK, fueron transferidos a un nuevo medio MMPK, con el fin de confirmar, la restitución del fenotipo prp a los mutantes, para finalmente ser semeados en medio LB con canamicina (LBK), con el objetivo de realizar posteriores análisis.
Es importante resaltar, que cada una de las culturas por separado, no puede crecer en estos medios, debido a que E. coli S17-1, necesita factores de crecimiento para obtener un aumento significativo de células, por ende no crece en un medio mínimo con sacarosa como fuente de carbono. Así mismo B. sacchari no crece en medio que contiene antibiótico, por tanto en este medio de cultivo, sólo crecen los mutantes de B. sacchari
que recibieron el cósmido pVK100 por conjugación. Los clones de E. coli capaces de complementar los mutantes, así como cada uno de los mutantes complementados fueron cultivados en medio LBK. Posteriormente fueron sometidos a extracción de DNA plasmidial, digeridos con endonucleasa de restricción EcoRI y analizados por electroforesis en gel de agarosa para verificar si ambas cultivos contenían el mismo conjunto de cósmido e inserto.
5.11 EXTRACCIÓN DE DNA PLASMIDIAL (MINIPREP)
incubadas en baño de hielo por 5 minutos. Posteriormente se adicionaron 200 L de SDS en NaOH 0,2 M; se mezclo por inversión y se incubó por 5 minutos en baño de hielo. Para la precipitación del DNA cromosomal e de las proteínas, se adicionaron 150 L de solución de acetato de potasio previamente enfriada y, después una nueva incubación por 5 minutos en baño de hielo, se centrifugó por 10 minutos, a 6000 rpm, con una temperatura de 4ºC. El sobrenadante, el cual contiene el DNA plasmidial fue transferido para un nuevo tubo, donde se procedió a realizar la extracción con fenol/cloroformo (250 L de fenol y 250 L de cloroformo), se centrifugó por 10 min, a 13000 rpm, a 4 ºC. En seguida, fue realizada la precipitación con etanol absoluto (750 L), posteriormente se incubó durante un periodo de 15 – 30 min en baño de hielo, centrifugándose por 10 minutos a 13000 rpm, 4 ºC y el “pellet” fue seco al vacío durante 20-30 min y resuspendido en agua con 0,01 % del volumen de la solución de RNAse y la mezcla fue incubada por 1 hora a 37 % y mantenida a -20 ºC. a continuación se encuentran descritas las soluciones empleadas para la extracción de DNA:
5.11.1 Solución de GETL (Birnborim e Doly, 1979)
Lisozima 4 mg/mL Glucosa 50mM
EDTA de calcio y sodio (etileno diamino tetracetato) 10mM
Tris/HCL (Hidroclorato de hidroximetil aminometano) 25Mm pH 8,0
5.11.2 Solución de acetato de potasio (Birnboim e Doly, 1979)
• Acetato de potasio 5M 60mL
• Acido acético 11,5mL
• H2O 28,5mL
5.12 GEL DE AGAROSA.
Para el análisis del DNA, se utilizó gel de agarosa en las concentraciones de 0.8% y 1,0 % (m/v). Se utilizaron las soluciones TBE, TAE y TAE modificado como tampón de corrida (Sambrook et al., 1989).
Las muestras de DNA fueron aplicadas en los gels, con 1/10 del volumen de colorante de corrida. Las corridas electroforéticas en mini-gels fueron realizadas a 90 V, 80 mA, 80 W durante 1 hora.
Los fragmentos fueron observados después de la coloración con brometo de etidio (0,1mg/100mL de TBE) durante 20 minutos, lavándolo posteriormente en el mismo tampón, con el fin de mejorar la visualización sobre luz UV (254 nm), en el transiluminador (Ultralum 100)
5.12.1 Solución TBE
Tris (hidroximetil aminometano) 54g Ácido bórico 27,5g
EDTA de calcio e sodio (etileno diamino tetracetato) 0,5M (20mL) H2O 1000mL
5.12.2 Solución TAE
Tris-base 242g Acido acético glacial 57,1mL
EDTA de calcio y sodio (etileno diamino tetracetato) 0,5M (100mL) H2O 1000mL
5.12.3 Colorante de corrida
• Glicerol 5g
• H2O 100mL
5.13 MANIPULACIÓN DEL DNA 5.13.1 Digestión del DNA
Para la digestión del DNA, se adicionó la enzima de restricción según las instrucciones del fabricante (Invitrogen Corporation) incluyendo el tampón adecuado. Para cada 1μL de DNA, se utilizaron de 1 a 2 unidades de la enzima, de forma que esta representase máximo 10 % del volumen. La digestión del DNA fue incubada a 37 ºC “overnight”. Después de la digestión, se inactivo la enzima de restricción a 65 ºC por 10 min.
5.14 EXPERIMENTO DE ACUMULO DE PHA PARA EVALUAR EL EFECTO DE LAS COMPLEMENTACIONES OBTENIDAS.
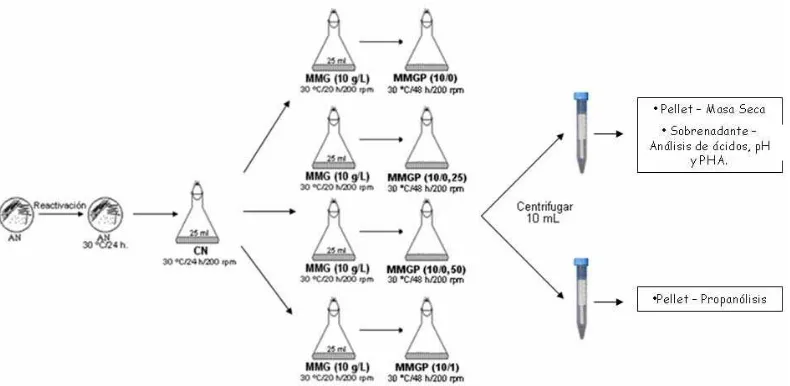
glucosa residual , fue adicionado ácido propiónico con el fin de asegurar la síntesis de P3HB-co-3HV, manejando 4 concentraciones diferentes, siendo a) sin adición de propionato (0 g/L), b) concentración de 0,25 g/L, c) concentración de 0,75 g/L, y d) concentración de 1,00 g/L. Posterior a la adición del propionato, fueron retiradas 4 muestras de 1,5 mL para respectivos análisis de biomasa, azúcares, ácidos, entre otros, para finalmente incubar las culturas en shaker a 200 rpm, 30 ºC durante 48 horas. Finalizadas las 72 horas, fueron retiradas 2 muestras de 10 mL, centrifugadas, donde los “pellets” obtenidos fueron empleados para determinar masa seca celular y para ser liofilizado con el fin de realizar propanólisis para medir PHA formado, como se muestra en la figura 3. Fueron determinados biomasa, concentración de fuentes de carbono y de PHA en el inicio y final del ensayo. Los diferentes análisis fueron realizados como se encuentra descrito en Silva (1998).
Figura 3. Esquema de los experimentos de acumulo de polímero, realizados a los mutantes y mutantes complementados de la cepa B. sacchari LFM 101.
5.15 DETERMINACIONES ANALÍTICAS.
5.15.1 Concentración celular.
Para medir la concentración celular se determinó la masa seca celular, después de la centrifugación, filtrando en una membrana con diámetro de poro de 0,45 μm y dejándolas secar en una estufa a 105 ºC hasta obtener un peso constante, así mismo, se colocó como control una membrana sin filtración de ninguna muestra para evaluar la variación de la humedad.
5.15.2 Proporción de polímero
Los polihidroxialcanoatos producidos en las diferentes condiciones testadas fueron cuantificados por cromatografía gaseosa, como fue descrito por Riis & Mai (1988) modificado como se encuentra descrito en Silva (1998).
Se centrifugaron 10 mL de la suspensión celular (siendo éste un volumen de acuerdo a la concentración de PHB en la célula), a 7000 rpm durante 10 minutos. Posteriormente se descartó el sobrenadante y se mantuvo la biomasa, para ser liofilizada. Al tener las células en dicha condición se procedió a pesar 10 mg en tubos de vidrio de 10 mL con tapa rosca y fueron adicionados 0,1 mL de la solución ácido benzoico más propanol, más 1,5 mL de la solución de HCl más propanol y 1,5 mL de 1,2 dicloroetano. Seguidamente, los tubos fueron cerrados fuertemente con ayuda de teflón y se agitaron vigorosamente, para colocarlos finalmente en baño termostatado a 105 ºC por 3 horas, agitándolos en la primera media hora. Después de ser retirados y enfriados, se adicionaron 4 mL de agua destilada y se agitaron vigorosamente, con el fin de recuperar la fase orgánica en el fondo del tubo para análisis; finalmente se retiró el residuo de agua con sulfato de sodio anhidro.
Para calcular la concentración de la muestra, se realizó el siguiente cálculo:
FR amostra Masa benzóico A Masa benzóico A Área PHB Área
PHB . 100
.
6
RESULTADOS Y DISCUSIÓN.
6.1 ENSAYO FENOTÍPICO
[image:51.612.153.487.340.534.2]Los resultados de la validación de las características fenotípicas de los mutantes prp se representan en la Tabla 3, donde se muestra el consumo de los diferentes sustratos, pertenecientes a los intermediarios de la α-oxidación, así como la agrupación de los mismos de acuerdo a los sustratos consumidos. Se observa que el mutante LFM 198 a diferencia de los otros mutantes en estudio, presenta un buen crecimiento en medio con ácido láctico, lo cual puede indicar que posiblemente se encuentra afectado en los anteriores intermediarios de la α-oxidación.
Tabla 3. Validación del crecimiento de los mutantes prp de B. sacchari LFM 183, 184, 185, 186 y 198 en diferentes compuestos.
MMG MMS MMF MMP MMAL MMPir MMAAc GRUPO FENOTIPICO
LFM 101 +++ +++ +++ +++ +++ +++ +++ Salvaje
LFM 183 +++ +++ +++ --- --- --- --- I
LFM 184 +++ +++ +++ --- --- --- --- I
LFM 185 +++ +++ +++ --- --- --- --- I
LFM 186 +++ +++ +++ --- --- --- --- I
LFM 198 +++ +++ +++ --- ++ --- --- III
+++: Crecimiento altamente visible. ++: Buen crecimiento. +: Crecimiento difícilmente notable. ---: Ausencia de crecimiento. MMG (medio mineral con glucosa), MMS (medio mineral con sacarosa), MMF (medio mineral con fructosa), MMP (medio mineral con propionato), MMAL (medio mineral con ácido láctico), MMPir (medio mineral con Piruvato), MMAAc (medio mineral con ácido acético), todos con concentración final de 1 g/L.
Al comparar los resultados del presente ensayo con el experimento realizado cuando se agruparon los mutantes de B. sacchari obtenidos por UV como se encuentra
Medio
Los mutantes fueron evaluados en presencia de ácido acrílico, como se encuentra representado en la Tablas 4, donde se observa que se realizaron tres ensayos, en los cuales fue ofrecido como fuente de carbono ácido acrílico correspondientes a las siguientes concentraciones finales en el medio: 1 g/L, 0,5 g/L y 0,25 g/L, para cada uno de los mutantes en estudio.
Tabla 4. Validación del crecimiento de los mutantes prp de B. sacchari LFM 183, 184, 185, 186 y 198 en ácido acrílico.
MMAA
1 g/L 0,5 g/L 0,25 g/L
LFM 101 --- + ++
LFM 183 --- --- ++
LFM 184 --- + ++
LFM 185 --- + ++
LFM 186 --- --- ++
LFM 198 --- + ++
++: Buen crecimiento. +: Crecimiento difícilmente notable. ---: Ausencia de crecimiento. MMAA (medio mineral con ácido acrílico).
Como se observa en la tabla anterior, ninguna de las cepas creció en presencia de 1 g/L de ácido acrílico, lo cual está de acuerdo con los resultados de Silva (1998). Las cepas LFM 101, 184, 185 y 198 presentaron crecimiento difícilmente visible incluso después de 10 días de incubación, comparado con 0,5 g/L de ácido acrílico. Al analizar las cepas LFM 183 y 186, éstas no crecieron en ésta condición. Entre tanto, cuando se trabajó con una concentración de 0,25 g/L, todas las cepas presentaron un buen crecimiento. Estos resultados deben ser presentados por la actividad tóxica del acrilato, el cual puede actuar como inhibidor de la β-oxidación. La literatura presenta trabajos que utilizaron el ácido acrílico para inhibir la β-oxidación de ácidos grasos en organismos como especies de
Aeromonas (Kobayashi et al 1994) y en E. coli (Lu et al 2003). Los autores anteriormente mencionados, trabajaron con concentraciones de 0,1 g/L de ácido acrílico
Medio