TRABAJO DE GRADO
Presentado como requisito parcial para otorgar el titulo de:
Doctor en Ciencias Biológicas
ESTUDIO MOLECULAR DE ESTRUCTURA GENÉTICA EN POBLACIONES
INDÍGENAS Y AFROCOLOMBIANAS MEDIANTE MARCADORES NUCLEARES
Y MITOCONDRIALES
Por
MARÍA CLAUDIA NOGUERA SANTAMARÍA
Director
JAIME EDUARDO BERNAL VILLEGAS. MD. PhD
Director Instituto de Genética Humana,
Pontificia Universidad Javeriana
Codirector
IGNACIO BRICEÑO BALCAZAR MD. PhD
Médico genetista y profesor titular Pontificia Universidad Javeriana
Facultad de Medicina
Pontificia Universidad Javeriana
ESTUDIO MOLECULAR DE ESTRUCTURA GENÉTICA EN
POBLACIONES INDÍGENAS Y AFROCOLOMBIANAS
MEDIANTE MARCADORES NUCLEARES Y MITOCONDRIALES
Por
MARÍA CLAUDIA NOGUERA SANTAMARÍA
ESTUDIO MOLECULAR DE ESTRUCTURA GENÉTICA EN
POBLACIONES INDÍGENAS Y AFROCOLOMBIANAS
MEDIANTE MARCADORES NUCLEARES Y
MITOCONDRIALES
Por
NOTA DE ADVERTENCIA
ESTUDIO MOLECULAR DE ESTRUCTURA GENÉTICA EN
POBLACIONES INDÍGENAS Y AFROCOLOMBIANAS MEDIANTE
MARCADORES NUCLEARES Y MITOCONDRIALES
Por
MARÍA CLAUDIA NOGUERA SANTAMARÍA
APROBADO
Concepción Judith
Puerta Bula. PhD
Decana Académica
Facultad de Ciencias
Alba Alicia Trespalacios
PhD
DEDICATORIA
A los hombres que más quiero, mi padre y hermano, porque son mi
ejemplo y mi vida, por su apoyo en todo momento, porque me han
acompañado en el ascenso de cada escalón para lograr mis grandes
metas y principalmente porque sin ellos mi vida no estaría tan llena de
amor y compañía.
AGRADECIMIENTOS
A Dios, por la vida, la familia y por cada bendición recibida desde el
momento en que nací, por las virtudes otorgadas y por día a día de su
mano, llevar mi camino en la vida.
Al Doctor Jaime Bernal Villegas por darme la oportunidad de hacer
parte de su grupo de trabajo, por su enseñanza y sabiduría al
introducirme en el mundo de la genética, por sus lecciones de vida,
por ser como un padre, un amigo; gracias por compartir sus
conocimientos, su tiempo, y por facilitarme los medios para poder
llevar a cabo mi proyecto de tesis doctoral.
Al Doctor Ignacio Briceño, quien fue un apoyo para encontrar el
camino de un investigador; gracias por su guía, sus enseñanzas, su
amistad y por siempre estar dispuesto a dedicarme el tiempo
necesario.
A Daniel Uricoechea Patiño, por el apoyo brindado en la ejecución de
las gráficas de análisis, por su apoyo incondicional, el tiempo
invaluable que me ha regalado y lo más importante, su amistad.
A la Pontificia Universidad Javeriana, en cabeza de la Facultad
de Ciencias y el Instituto de Genética Humana por el apoyo en este
proceso.
A la Universidad de la Sabana por prestarme sus instalaciones para
realizar parte de mi proyecto.
Al Doctor Jairo Osorno, por sus sabios y oportunos consejos.
Finalmente al Instituto de Patología e inmunología Molecular de Porto.
“IPA
TIMUP
”
, en Porto, Portugal, por su colaboración, su apoyo, todas
Sus ilustraciones y por darme todo para realizar mis análisis
moleculares en Portugal; principalmente a Leonor Gusmao y Verónica
Gómes, quienes me brindaron más que eso, su amistad.
Tabla de contenido
Tabla de contenido ... 1
Indice de Tablas ... 6
RESUMEN ... 17
ABSTRACT ... 19
1. INTRODUCCIÓN ... 20
2. JUSTIFICACIÓN ... 22
3. OBJETIVOS ... 23
3.1. Objetivo general ... 23
3.2. Objetivos específicos: ... 23
4. MARCO TEÓRICO ... 24
4.1. Origen y poblamiento ... 24
4.2. Aspectos históricos y Arqueológicos ... 26
4.3. El tráfico de esclavos africanos ... 28
4.4. Etnografía de las poblaciones estudiadas ... 29
4.4.1. Chimila ó Ette ennaka ... 30
4.4.2. Guahibo ... 30
4.4.3. Guane ... 30
4.4.4. Guayabero ... 30
4.4.5. Inga ó Ingano ... 31
4.4.6. Pastos ... 31
4.4.7. Tukano ... 31
4.4.8. Uitoto ... 31
4.4.10. Zenú ... 32
4.4.11. Paez ... 32
4.4.12. Guambiano ... 32
4.5. Clasificación de Lenguas Indígenas de Colombia ... 33
4.5.1. Lenguas aisladas: ... 34
4.5.2. Tronco Lingüístico Arawak ... 34
4.5.3. Familia Caribe ... 35
4.5.4. Familia Quechua ... 35
4.6. Población Afrocolombiana del Caribe ... 36
4.6.1. Palenque de San Basilio ... 40
4.7. Reseña histórica de la expedición humana ... 41
4.7.1. Antecedentes de Expedición Humana ... 42
4.8. Marcadores Moleculares ... 52
4.8.1. La Mitocondria ... 52
4.8.2. ADN Mitocondrial ... 53
4.8.3. Cromosoma Y ... 67
4.8.4. Similitudes entre el cromosoma Y y el ADN Mitocondrial (ADNmt) ... 80
4.8.5. Inserciones ALU ... 82
5. METODOLOGIA ... 83
5.1. Muestreo Poblaciones Indígenas ... 83
5.1.1. Tipificación de Haplogrupos mitocondriales ... 84
5.1.2. Estudio mitocondrial de haplogrupos por RFLPs ... 85
5.1.3. Estudio Mitocondrial de Haplotipos por secuenciación directa ... 88
5.1.4. Amplificación PCR ... 88
5.1.5. Estimadores Genético Poblacionales. ... 89
5.1.6. Diversidad Génica ... 91
5.1.7. Clasificación de las secuencias en haplogrupos ... 91
5.2. Muestreo de poblaciones Afrocolombianas del Caribe. ... 93
5.2.1. Tipificación de Cromosoma Y ... 94
5.2.2. Análisis STR ... 95
5.2.3. Análisis de Alelos. ... 97
5.2.4. Analisis genético – poblacionales. ... 99
6. RESULTADOS Y DISCUSIÓN ... 105
6.1. Análisis Mitocondrial por RFLPs ... 105
6.1.1. Agrupación Lingüística Amovas ... 106
6.1.2. Agrupación Geográfica Amovas ... 107
6.1.3. Distancias Genéticas ... 108
6.1.4. Análisis Mitocondrial (Región Hipervariable HVS-I) ... 123
6.1.5. REDES MEDIANAS ... 129
6.1.6. Análisis SNaPShot muestras totales ... 131
6.1.7. SNaPShot y Análisis STR ... 132
6.2. Análisis Componente Europeo, Africano y Amerindio en la población Afrocolombiana y en comparación con Uganda , África ... 140
6.3. Análisis de Frecuencias Alélicas en la región Afrocolombiana ... 143
6.3.1. Sistema DYS19 ... 143
6.3.2. Sistema DYS389-I ... 144
6.3.3. Sistema DYS389-II ... 144
6.3.4. Sistema DYS390 ... 144
6.3.5. Sistema DYS391 ... 144
6.3.6. Sistema DYS392 ... 145
6.3.7. Sistema DYS393 ... 145
6.3.8. Sistema DYS438 ... 145
6.3.9. Sistema DYS439 ... 145
6.3.10. Sistema DYS437 ... 145
6.3.11. Sistema DYS448 ... 146
6.3.12. Sistema DYS456 ... 146
6.3.13. Sistema DYS458 ... 146
6.3.14. Sistema DYS635 ... 146
6.3.15. Sistema YGATA-H4 ... 147
6.4. Determinación de haplotipos. ... 157
6.5. Analisis comparativo con otras poblaciones en el mundo ... 167
6.6. Analisis Networks ... 169
7. CONCLUSIONES ... 174
8. BIBLIOGRAFIA ... 177
9. ANEXOS ... 189
9.1. Análisis de marcadores sanguíneos definidos por la Expedición Humana .. 189
A continuación se presentan tablas y árboles filogenéticos contruidos a partir de los datos obtenidos por los investigadores asociados a la Expedición Humana entre 1989 y 1993. Se incluyen dendrogramas, cladogramas y filogramas. ... 189
9.1.1. Frecuencias Análisis de Expedición Humana y árboles filogenéticos .. 189
9.2. Estrategia de Montaje de muestras ... 258
9.3. SNaPShot Mx2 ... 259
9.4. MULTIPLEX 1 ... 260
9.5. SNaPShot MxQ ... 261
9.6. SNaPshot Mx1 + M13 ... 262
9.7. SNaPshot Mx1 + M13 ... 264
Indice de Tablas
Tabla 1 Distribución de Antígenos HLA A, B, C, DR y DQ en poblaciones amerindias
que habitan las costas del Pacífico y Atlántico Colombiano ... 45
Tabla 2 Sistemas analizados en Expedición Humana (cortesía del Doctor Jaime Bernal) ... 47
Tabla 3 Sistemas sanguíneos y sus alelos ... 51
Tabla 4 Sistema ABO ... 52
Tabla 5 Identificación de haplogrupos por enzimas de restricción ... 59
Tabla 6 Tipificación de los haplogrupos A-E. ... 60
Tabla 7 Características de los marcadores Y-STR utilizados para este estudio ... 75
Tabla 8 Tabla comparativa de Haplogrupos de ADN Mitocondrial y Cromosoma Y .... 81
Tabla 9 Primers utilizados. ... 85
Tabla 10 Estandarización PCR ... 86
Tabla 11 Primers HVRI ... 88
Tabla 12 Condiciones de PCR HVRI ... 88
Tabla 13 Programas estadísticos más utilizados en Genética de poblaciones. ... 89
Tabla 14 Cambio en las posiciones que definen los haplogrupos mitocondriales de la Región Hipervariable. ... 91
Tabla 15 Acceso de los loci STR estudiados ... 96
Tabla 16 Condiciones de PCR STRs ... 97
Tabla 17 Frecuencias de los haplogrupos ADNmt en poblaciones colombianas ... 107
Tabla 18 Distancias Genéticas Fst en las poblaciones estudio. ... 109
Tabla 19 Correlación entre genes y lengua ... 116
Tabla 20 Análisis de Frecuencias y Diversidad estimada en poblaciones indígenas. 120
Tabla 21 Diversidad por Haplogrupos de ADNmt en poblaciones de Norte, Centro y
Suramérica ... 122
Tabla 22 Polimorfismo encontrados en el análisis de secuencias ... 124
Tabla 23 Frecuencias haplotípicas observadas ... 125
Tabla 24 Frecuencias de haplogrupos mitocondriales encontrado en Rodas 2003 ... 128
Tabla 25 frecuencia de haplogrupos en cromosoma Y ... 137
Tabla 26 Consenso STRs + SNaPShot Afrocolombianos. ... 158
Tabla 27 consenso de haplotipos totales por STRs Cromosoma Y ... 160
Tabla 28 Consenso Haplotipos Afrocolombianos ... 163
Tabla 29 Distribución de Frecuencias Alélicas de haplotipos Cromosoma Y ... 164
Tabla 30 Population pairwise FSTs ... 166
Tabla 31 Haplotipos Otras partes del mundo ... 167
Tabla 32 Matriz FTz sistema transferrina ... 189
Tabla 33 Matriz FTz PGM1. ... 194
Tabla 34 Matriz FTz MITOCONDRIAL ... 198
Tabla 35 Matriz FTz LUTHERANS ... 203
Tabla 36 Matriz FTz LEWIS ... 207
Tabla 37 Matriz FTz KPA-B ... 211
Tabla 38 Matriz FTz KIDD ... 215
Tabla 39 Matriz FTz IgSA2 ... 219
Tabla 40 Matriz FTz IgSA1 ... 220
Tabla 41 Matriz FTz HLA DR ... 221
Tabla 42 Matriz FTz HLA DQ ... 225
Tabla 43 Matriz FTz HLA C ... 229
Tabla 44 Matriz FTz HLA A ... 233
Tabla 45 Matriz FTz HLA B ... 237
Tabla 46 Matriz FTz HAPTOGLOBINAS ... 241
Tabla 47 Matriz FTz DUFFY ... 245
Tabla 48 Matriz FTz C3 ... 249
Tabla 49 Matriz FTz ABO ... 253
Tabla 50 Matriz FTz ALFA 1 ANTITRIPSINA ... 257
Índice de figuras
Figura 1. Áreas Arqueológicas y su cronología [64] ... 27
Figura 2 Mapa de poblamiento de América ... 28
Figura 3 Familias lingüísticas del mundo antes de Greenberg ... 36
Figura 4 Localización Geográfica de la población del Caribe. ... 37
Figura 5 Esquema de ADN mitocondrial humano ... 55
Figura 6 Migraciones con base en el ADNmt ... 62
Figura 7 Mapa de los marcadores STRs del Cromosoma Y humano ... 71
Figura 8 SNPs ... 73
Figura 9 Repeticiones cortas en tándem (STRs) ... 75
Figura 10 Posiciones de STR a través del cromosoma Y ... 78
Figura 11 Distribución mundial de la frecuencia en haplogrupos Y ... 79
Figura 12 Distribución de la frecuencia haplotipica del cromosoma Y en Europa, África, Asia y Australia. ... 81
Figura 13 Secuencias Alu (SINE’s) ... 83
Figura 14 Regiones de ADN mitocondrial amplificadas. ... 89
Figura 15 Electroferogramas de haplotipos STRs ... 98
Figura 16 Agrupación lingüística de poblaciones de estudio ... 106
Figura 17 Agrupación geográfica de poblaciones de estudio ... 107
Figura 18 Mapa de frecuencias de haplogrupos mitocondriales en poblaciones nativas Colombianas. ... 108
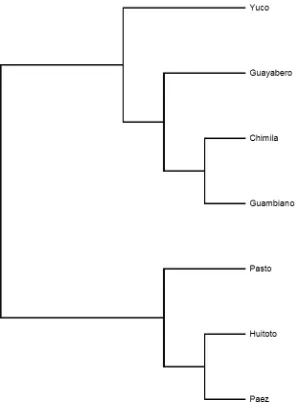
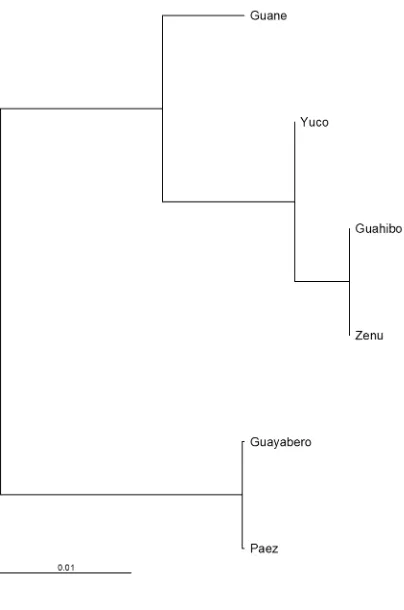
Figura 19 Dendrograma por UPGMA de las poblaciones en estudio. ... 110
Figura 20 Cladograma por UPGMA de las poblaciones en estudio. ... 111
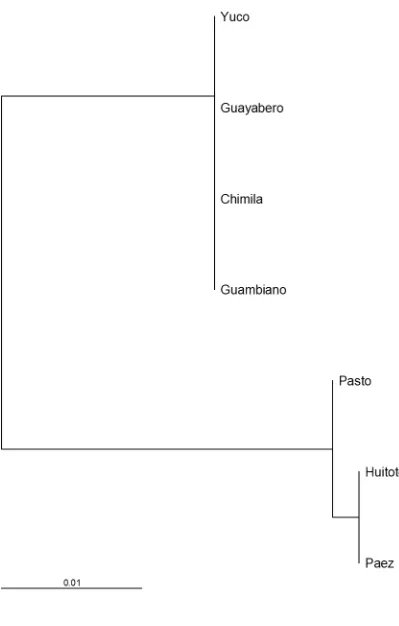
Figura 21 Filograma por UPGMA de las poblaciones en estudio. ... 112
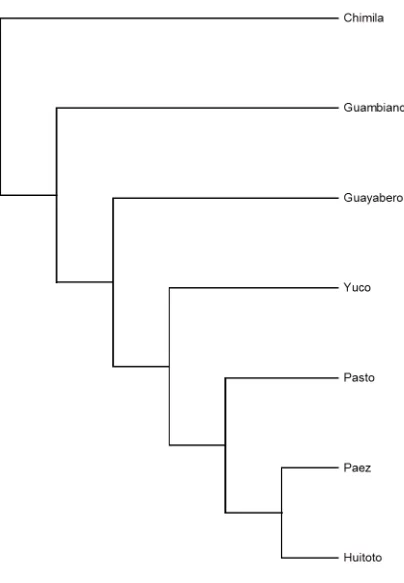
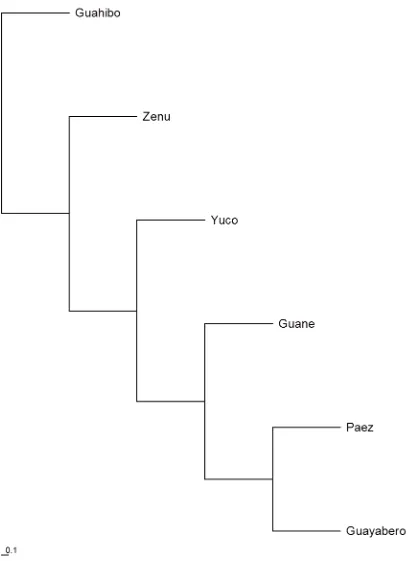
Figura 22 Cladograma por UPGMA + majority-rule consensus tree de las poblaciones
en estudio. ... 113
Figura 23 Filograma por UPGMA + majority-rule consensus tree de las poblaciones en estudio. ... 114
Figura 24 Geles de agarosa por haplogrupos ... 118
Figura 25 Network de las poblaciones de estudio. ... 127
Figura 26 Árbol de correspondencia Cromosoma Y ... 131
Figura 27 Árbol filogenético Y ... 133
Figura 28 Expansión 1 por Underhill. [157] ... 138
Figura 29 Expansión 2 por Underhill ... 139
Figura 30 Componente frecuencias haplogrupos en cromosoma Y en el presente estudio. ... 140
Figura 31 Ocurrencia de haplotipos Y por SNaPShot en las poblaciones afrocolombianas ... 141
Figura 32 Ocurrencia de haplotipos Y por SNaPShot en Palenque de San Basilio ... 142
Figura 33 Ocurrencia de haplotipos Y por SNaPShot en Cartagena ... 143
Figura 34 Distribución De las frecuencias alélicas en la Costa Caribe para el locus DYS19 ... 147
Figura 35 Distribución De las frecuencias alélicas en la Costa Caribe para el locus DYS389I ... 148
Figura 36 Distribución De las frecuencias alélicas en la Costa Caribe para el locus DYS389II ... 149
Figura 37 Distribución De las frecuencias alélicas en la Costa Caribe para el locus DYS390 ... 150
Figura 38 .Distribución De las frecuencias alélicas en la Costa Caribe para el locus DYS391 ... 150
Figura 39 Distribución De las frecuencias alélicas en la Costa Caribe para el locus
DYS392 ... 151
Figura 40 Distribución De las frecuencias alélicas en la Costa Caribe para el locus DYS393 ... 151
Figura 41 . Distribución De las frecuencias alélicas en la Costa Caribe para el locus DYS385 ... 152
Figura 42 Distribución De las frecuencias alélicas en la Costa Caribe para el locus DYS437 ... 153
Figura 43 Distribución De las frecuencias alélicas en la Costa Caribe para el locus DYS438 ... 154
Figura 44 Distribución De las frecuencias alélicas en la Costa Caribe para el locus DYS439 ... 154
Figura 45 Distribución De las frecuencias alélicas en la Costa Caribe para el locus DYS448 ... 154
Figura 46 Distribución De las frecuencias alélicas en la Costa Caribe para el locus DYS456 ... 155
Figura 47 Distribución De las frecuencias alélicas en la Costa Caribe para el locus DYS458 ... 155
Figura 48 Distribución De las frecuencias alélicas en la Costa Caribe para el locus DYS635 ... 156
Figura 49 Distribución De las frecuencias alélicas en la Costa Caribe para el locus YGATA H4 ... 156
Figura 50 UPGMA poblaciones Afrocolombianas ... 166
Figura 51 Median Joining Haplogrupo E CR-Y ... 170
Figura 52 Reduced Median Haplogrupo E CR-Y ... 170
Figura 53 Reduced Median Haplogrupo R CR-Y ... 171
Figura 54 Median Joining Haplogrupo R CR-Y ... 172
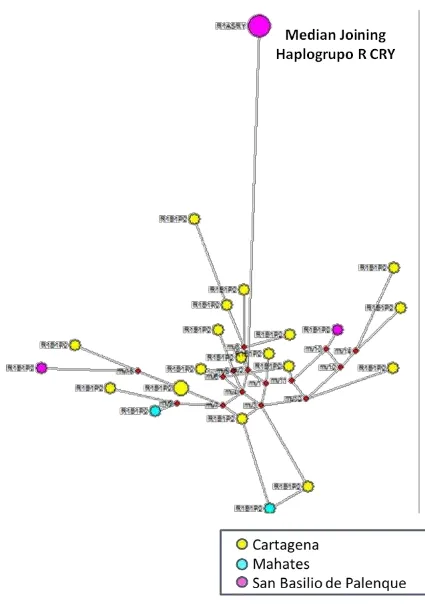
Figura 55 Reduced Median Haplogrupo E Presente Estudio + Gomes 2010 [3] ... 173
Figura 56 Cladograma método UPGMA SISTEMA TRANSFERRINA ... 190
Figura 57 Filograma método UPGMA SISTEMA TRANSFERRINA ... 191
Figura 58 Cladograma método UPGMA + majority-rule consensus tree SISTEMA TRANSFERRINA ... 192
Figura 59 Filograma método UPGMA + majority-rule consensus tree SISTEMA TRANSFERRINA ... 193
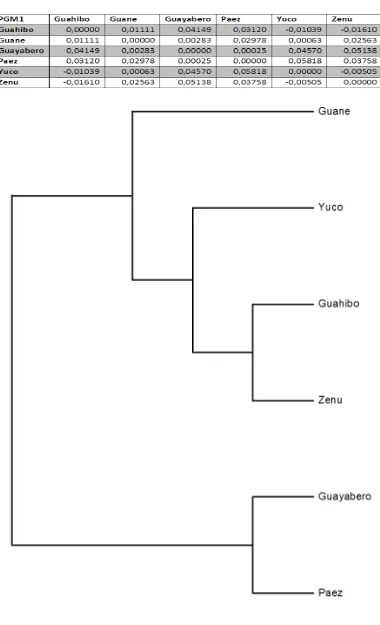
Figura 60 Cladograma método UPGMA SISTEMA PGM1 ... 194
Figura 61 Filograma método UPGMA SISTEMA PGM1 ... 195
Figura 62 Cladograma método UPGMA + majority-rule consensus tree SISTEMA PGM1 ... 196
Figura 63 Cladograma método UPGMA + majority-rule consensus tree SISTEMA PGM1 ... 197
Figura 64 Filograma método UPGMA + majority-rule consensus tree SISTEMA PGM1 ... 198
Figura 65 Cladograma método UPGMA SISTEMA MITOCONDRIAL ... 199
Figura 66 Filograma método UPGMA SISTEMA MITOCONDRIAL ... 200
Figura 67 Cladograma método UPGMA + majority-rule consensus tree SISTEMA MITOCONDRIA ... 201
Figura 68 Filograma método UPGMA + majority-rule consensus tree SISTEMA MITOCONDRIA ... 202
Figura 69 Cladograma método UPGMA SISTEMA LUTHERANS ... 203
Figura 70 Filograma método UPGMA SISTEMA LUTHERANS ... 204
Figura 71 Cladograma método UPGMA + majority-rule consensus tree SISTEMA LUTHERANS ... 205
Figura 72 Filograma método UPGMA + majority-rule consensus tree SISTEMA LUTHERANS ... 206
Figura 73 Cladograma método UPGMA SISTEMA LEWIS ... 207
Figura 74 Filograma método UPGMA SISTEMA LEWIS ... 208
Figura 75 Cladograma método UPGMA + majority-rule consensus tree SISTEMA LEWIS ... 209
Figura 76 Filograma método UPGMA + majority-rule consensus tree SISTEMA LEWIS ... 210
Figura 77 Cladograma método UPGMA SISTEMA KPA-B ... 211
Figura 78 Filograma método UPGMA SISTEMA KPA-B ... 212
Figura 79 Cladograma método UPGMA + majority-rule consensus tree SISTEMA KPA- B ... 213
Figura 80 Filograma método UPGMA + majority-rule consensus tree SISTEMA KPA-B ... 214
Figura 81 Cladograma método UPGMA SISTEMA KIDD ... 215
Figura 82 Filograma método UPGMA SISTEMA KIDD ... 216
Figura 83 Cladograma método UPGMA + majority-rule consensus tree SISTEMA KIDD ... 217
Figura 84 Filograma método UPGMA + majority-rule consensus tree SISTEMA KIDD ... 218
Figura 85 Cladograma y Filograma método UPGMA SISTEMA IgSA2 ... 219
Figura 86 Cladograma y Filograma método UPGMA + majority-rule consensus tree SISTEMA IgSA2 ... 219
Figura 87 Cladograma y Filograma método UPGMA SISTEMA IgSA1 ... 220
Figura 88 Cladograma y Filograma método UPGMA + majority-rule consensus tree SISTEMA IgSA1 ... 220
Figura 89 Cladograma método UPGMA SISTEMA HLA DR ... 221
Figura 90 Filograma método UPGMA SISTEMA HLA DR ... 222
Figura 91 Cladograma método UPGMA + majority-rule consensus tree SISTEMA HLA DR ... 223
Figura 92 Filograma método UPGMA + majority-rule consensus tree SISTEMA HLA DR ... 224
Figura 93 Cladograma método UPGMA SISTEMA HLA DQ ... 225
Figura 94 Filograma método UPGMA SISTEMA HLA DQ ... 226
Figura 95 Cladograma método UPGMA + majority-rule consensus tree SISTEMA HLA DQ ... 227
Figura 96 Filograma método UPGMA + majority-rule consensus tree SISTEMA HLA DQ ... 228
Figura 97 Cladograma método UPGMA SISTEMA HLA C ... 230
Figura 98 Filograma método UPGMA SISTEMA HLA C ... 230
Figura 99 Cladograma método UPGMA + majority-rule consensus tree SISTEMA HLA C ... 231
Figura 100 Filograma método UPGMA + majority-rule consensus tree SISTEMA HLA C ... 233
Figura 101 Cladograma método UPGMA SISTEMA HLA A ... 234
Figura 102 Filograma método UPGMA SISTEMA HLA A ... 234
Figura 103 Cladograma método UPGMA + majority-rule consensus tree SISTEMA HLA A ... 235
Figura 104 Filograma método UPGMA + majority-rule consensus tree SISTEMA HLA A ... 236
Figura 105 Cladograma método UPGMA SISTEMA HLA B ... 237
Figura 106 Filograma método UPGMA SISTEMA HLA B ... 238
Figura 107 Cladograma método UPGMA + majority-rule consensus tree SISTEMA HLA B ... 239
Figura 108 Filograma método UPGMA + majority-rule consensus tree SISTEMA HLA B ... 240
Figura 109 Cladograma método UPGMA SISTEMA HAPTOGLOBINAS ... 241
Figura 110 Filograma método UPGMA SISTEMA HAPTOGLOBINAS ... 242
Figura 111 Cladograma método UPGMA + majority-rule consensus tree SISTEMA HAPTOGLOBINAS ... 243
Figura 112 Filograma método UPGMA + majority-rule consensus tree SISTEMA HAPTOGLOBINAS ... 244
Figura 113 Cladograma método UPGMA SISTEMA DUFFY ... 245
Figura 114 Filograma método UPGMA SISTEMA DUFFY ... 246
Figura 115 Cladograma método UPGMA + majority-rule consensus tree SISTEMA DUFFY ... 247
Figura 116 Filograma método UPGMA + majority-rule consensus tree SISTEMA DUFFY ... 248
Figura 117 Cladograma método UPGMA SISTEMA C3 ... 249
Figura 118 Filograma método UPGMA SISTEMA C3 ... 250
Figura 119 Cladograma método UPGMA + majority-rule consensus tree SISTEMA C3 ... 251
Figura 120 Filograma método UPGMA + majority-rule consensus tree SISTEMA C3 ... 252
Figura 121 Cladograma método UPGMA SISTEMA ABO ... 253
Figura 122 Filograma método UPGMA SISTEMA ABO ... 254
Figura 123 Cladograma método UPGMA + majority-rule consensus tree SISTEMA ABO ... 255
Figura 124 Filograma método UPGMA + majority-rule consensus tree SISTEMA ABO ... 256
Figura 125 Cladograma y Filograma método UPGMA SISTEMA ALFA 1
ANTITRIPSINA ... 257
Figura 126 Cladograma y Filograma método UPGMA + majority-rule consensus tree SISTEMA ALFA 1 ANTITRIPSINA ... 257
RESUMEN
El estudio de la variabilidad genética de las poblaciones humanas no solo ha ampliado el panorama de la evolución de la especie y su prehistoria, sino que ha facilitado la identificación de las variantes patológicas dentro de la diversidad genética natural [1]. La aproximación más destacada en Colombia para el estudio de dicha variabilidad fue, sin duda, la Expedición Humana, proyecto adelantado por el Instituto de Genética Humana (IGH), de la Pontificia Universidad Javeriana, en Bogotá, durante la década de los 90s. Muchos estudios se han realizado desde ese entonces en el país, sin que se haya hecho un esfuerzo claro por consolidar resultados y sistematizar la información existente. El presente proyecto busca, como un complemento de la Expedición Humana, ampliar con técnicas moleculares actuales el estudio de la variabilidad genética humana en Colombia. Un gran obstáculo en la consolidación de un panorama global de dicha variabilidad es la heterogeneidad en los marcadores estudiados y las diferentes técnicas que los distintos investigadores en todo el país han reportado, por lo cual un estudio que incluya una extensa muestra de distintas etnias a lo largo de la geografía colombiana, empleando una amplia gama de técnicas (ADN mitocondrial (ADNmt), Polimorfismos de repeticiones cortas en tándem (STRs) autosómicos y minisecuenciación por SNaPShot, permitirá fundamentar una visión integral a nivel genético del país que, por su posición geográfica, es asimismo la entrada norte hacia Suramérica.
Con objeto de dilucidar los procesos de migración en poblaciones indígenas y Afrocolombianas, así como la influencia, miscegenación y aporte genético de diversos grupos humanos de antes y después de la conquista, este trabajo ha estudiado la estructura genética mediante la caracterización de los haplogrupos y haplotipos del ADN mitocondrial y el establecimiento de relaciones filogenéticas de los grupos amerindios, utilizando parámetros genéticos de distancia, subestructura y diversidad, análisis de polimorfismos STRs y caracterización y determinación de haplogrupos de cromosoma Y mediante análisis de secuenciación y extensión de una sola base (SBE) por SNaPShot.
Los estudios mitocondriales se llevaron a cabo en 14 poblaciones indígenas mediante Polimorfismos de longitud de fragmentos de restricción (RFLPs), definiendo los
haplogrupos existentes (cuatro haplogrupos fundadores), en función de tres sitos de restricción [2], y de una deleción en la región intergénica V, secuenciación de la Región Hipervariable I (HVR-I), secuencias que fueron alineadas de acuerdo con la secuencia de referencia Cambridge (rCRS). Los resultados confirman la variabilidad existente en la población colombiana, el 97.87% de los individuos, presentó como se esperaba, los linajes fundadores en América (A, B, C y D); el 2,13% no fué determinado por los RFLPs. El análisis de Región Hipervariable I. (HVR-I), en la región control del ADN mitocondrial (ADNmt), mostró coincidencia con los resultados obtenidos en el análisis mediante RFLPs. Adicionalmente, en la población Palenque de San Basilio el linaje materno africano, presentando los haplotipos L0a, L1b, L1c y Le31a, se observó en un 100%,
Por otro lado se estudiaron 6 poblaciones de la región Caribe colombiana mediante Y-Plex (17 STRs de cromosoma Y) [3, 4] con los microsatélites: DYS19, DYS389I, DYS389II, DYS390, DYS391, DYS392, DYS393, DYS438, DYS439, DYS437, DYS448, DYS456, DYS458, DYS635, YGATAH4 y DYS385 y por la genotipificación de un largo número de SNPs de cromosoma Y, pertenecientes a los principales haplogrupos representados en el mundo. Se hizo el análisis SNaPShot, con el estudio de 41 Polimorfismos de nucleótido simple (SNPs), específicos de cromosoma Y, y determinación de sus haplogrupos, permitiendo la aproximación hacia el estudio del origen de las poblaciones Afrocolombianas y la medida de variación entre ellas [3]. Los resultados evidencian un alto componente europeo, 67% Haplogrupo R, 26% africano haplogrupo E y 7% Amerindio, Haplogrupo Q. Un solo individuo presentó el haplogrupo B exclusivo y restrictivo de África subsahariana; los datos se compararon con los obtenidos en un estudio de poblaciones africanas [3]. Nuestros resultados permitieron hacer inferencias acerca del origen de la población estudiada; los datos fueron consistentes con el consorcio de árbol de cromosoma Y [5], encontrando sobretodo en la población Palenque de San Basilio un elevado componente africano, con el haplogrupo E1b1a-M2, perteneciente a África oriental, pudiendo aseverar que las poblaciones afrocolombianas son diferentes por evolución divergente.
ABSTRACT
The study of genetic variability of human populations has not only expanded the overview of the evolution of the species and it prehistory, it has facilitated the identification of the pathological variants within the natural genetic diversity (1). The most prominent approach in Colombia to study this variability was, without doubt, the Human Expedition, a project advanced by the Institute of Human Genetics (IGH) in the Pontifical Javeriana University in Bogotá during the 90’s. Many studies have been conducted since then in the country, in a clear effort to consolidate results and systematize the existing information.
Mitochondrial studies were conducted in 14 indigenous groups, through restriction fragment length polymorphism (RFLPs), defining existing haplogroups (four founding haplogroups) based of three restriction sites (2) and a deletion in the intergenic V region (HVR-I). Sequences were aligned according to the Anderson reference sequence (rCRS). The results confirm the variability in Colombian population; The 97,87% of individuals presented as expected the American founding lineages (A, B, C and D). The analyses on hipervariable region I (HVR-I) in the control region of mitochondrial DNA (mtDNA) showed agreement with the results obtained in the RFLP analyses and evidence 100% of African maternal lineages presenting haplotypes L0a, L1, L1b, L1c, and Le31a at Palenque San Basilio population.
On the other hand, six populations were studied in the Colombian carribean region with microsatellites (Y-PLEX, 17 Y-Chromosome STR) (3-4): DYS19, DYS389I, DYS389II, DYS390, DYS391, DYS392, DYS393, DYS438, DYS439 and YGATAH4. A large number of Y-Cromosome SNPs (single nucleotide polymorphisms) were genotyped by SNaPShot analysis, performed with a study of 40 Y chromosome specific SNPs, to determine african origin and the extent of variation between them (3). The results show high european component: Haplogroup R (67%), african Haplogroup E(26%), and amerindian haplogroup Q (7%). A single individual had Haplogroup B restrictive to Áfrican sub- Saharan region. The data were consistent with the consortium of Y- Chromosome tree and Karafet, 2008 (5), A large east Áfrican component with subhaplogroup E1b1a*-M2 was found in the Palenque San Basilio population.
1. INTRODUCCIÓN
Una de las características de Colombia, es su diversidad sociocultural debido a los grupos étnicos existentes: Indígenas, afrocolombianos, raizales y gitanos entre otros y que se ubican en diferentes partes del territorio nacional tanto en la parte rural como la urbana. Los análisis de diversidad nucleotídica de los haplogrupos mitocondriales sugieren el paso de éstos por un cuello de botella con una posterior recuperación poblacional, refiriéndose un cuello de botella cuando se ha experimentado un drástico descenso en el número de miembros en algún momento del pasado, llegando en algunos casos a estar al borde de la desaparición [6]. Como consecuencia, los ejemplares de las generaciones posteriores al cuello de botella presentan una escasa variabilidad genética y la antigua proporción de alelos en el conjunto de la población ha cambiado considerablemente. Por otro lado, también se ha observado en algunas zonas como el noroeste colombiano un efecto fundador. [7].
Estos estudios sugieren, que el comienzo de la diferenciación de la población ancestral del hombre moderno ocurrió hace 30.000 a 40.000 años, valores que se correlacionan con el modelo de colonización del Nuevo Mundo que se dio hace unos 14.000 años. Aún sigue siendo tema de debate el origen de los indígenas americanos [8-10]. Muchos estudios previos han descrito a la perfección que se produjo desde Asia; debido que en épocas pre-hispánicas (S. XV AC), los medios de transportes marítimos eran limitados y no permitían grandes trayectos por el mar, era posible pensar que las migraciones hacia América desde África fueron por Asia menor, Europa y Asia oriental y posteriormente atravesando el estrecho de Bering para llegar a las Américas [11, 12].
El estudio de ADN mitocondrial (ADNmt), [13, 14], se ha utilizado en investigaciones que buscan probar hipótesis relacionadas con el poblamiento humano, debido a sus importantes características que permiten discriminar y cuantificar la relación genética entre individuos y poblaciones relacionadas cercanamente [15-20]. La divergencia que se presenta en el ADN de mitocondria, ha permitido observar en términos moleculares y de tiempo la escisión de los grupos humanos desde África hace 50.000 - 60.000 años aproximadamente según algunos investigadores y que los orígenes de la población europea estuvieron en un grupo escindido que salió de África [21]. Todas las
diferencias encontradas en el ADN mitocondrial son debidas a mutaciones, las cuales con el tiempo se acumulan y dan lugar a los linajes mitocondriales [22]. Estos lazos existentes entre los linajes pueden ser evaluados a través de redes filogenéticas donde las mutaciones pueden clasificarse según una estructura jerárquica y de manera importante nos dan información sobre las migraciones, incluyendo las que se iniciaron en África [23, 24].
El Caribe colombiano, zona de impregnación africana en Colombia como lo refiere Nina Friedemann [25], guarda su historia cultural, y también, importantes elementos para el entendimiento de los procesos que dieron forma a su actual estructura. Los estudios realizados en la costa Caribe colombiana han sido escasos entre los cuales se encuentran los publicados por Builes [26, 27]. En cuanto a los estudios realizados con base en estudios de cromosoma-Y, solamente se han presentado algunos de carácter específico en poblaciones europeas, asiáticas y amerindias [28-30] [31]. Los resultados de Cromosoma Y nos permiten investigar el origen, estructura y dinámica de las poblaciones; el análisis genético – poblacional proporciona datos sobre aquellas características que estarían en la población original y que siguen estando representadas en la población actual lo cual podrá ser usado para ser inferencias sobre el origen y composición de la población.
Los orígenes de la diversidad poblacional, se relacionan no solamente con el flujo génico (migración), sino también con la deriva genética y con el proceso de adaptación a diferentes ecosistemas que debieron generar una relación entre las mutaciones y la selección natural.
2. JUSTIFICACIÓN
Los resultados de las investigaciones realizadas a través de los años han puesto en consideración dos preguntas sobre el origen del hombre (Número y fecha de las migraciones). Desde Norteamérica cruzaron a través de Centroamérica hacia lo que hoy és Colombia, que se convirtió en el punto de dispersión de las diversas poblaciones amerindias suramericanas [6, 16, 18, 19, 32, 33].
Debido a la alta variabilidad encontrada en el país, surge la necesidad de profundizar en el estudio de la estructura genética de la población colombiana. Desde hace varios años se viene estudiando la estructura genética de la población colombiana [10, 20, 34-47], evaluando de manera directa la composición genética de las poblaciones, lo que permite inferir los procesos evolutivos sucedidos en la historia (Schurr et al. 2004). Los marcadores mitocondriales permiten en este estudio evaluar claramente las poblaciones, identificando el origen de los individuos por medio de haplotipos característicos y la secuenciación de la Región Hipervariable I (HVR-I) en la Región Control [48, 49]. Además de los estudios mitocondriales, los análisis de Cromosoma Y, consisten en el estudio de la variabilidad de una parte del ADN nuclear que se transmite por vía paterna y que no experimenta recombinación, arrojan datos que proporcionan información muy valiosa sobre otros aspectos de nuestra historia evolutiva; mediante análisis microsatélites y por SNaPShot permitieron evaluar la diversidad génica y patrones de variación genética del Cromosoma Y [47] junto con los elementos Alu que son fuente importante de indicadores genéticos y que generan un gran interés para el estudio de la estructura genética de las poblaciones y origen geográfico individual [1, 50].
Este proyecto pretende ser un complemento de la Expedición Humana, ampliando con la utilización de técnicas moleculares el estudio de la variabilidad genética humana en Colombia.
3. OBJETIVOS
3.1. Objetivo general:
Estudiar la estructura genética de poblaciones amerindias y afrocolombianas mediante marcadores moleculares nucleares y mitocondriales que permiten evaluar diversidad y establecer relaciones filogenéticas entre los grupos humanos.
3.2. Objetivos específicos:
Establecer la variabilidad genética de poblaciones indígenas contemporáneas utilizando parámetros genéticos de distancia, sub-estructura y diversidad.
Determinar polimorfismos mitocondriales mediante análisis con enzimas de restricción (RFLP’s) y secuenciación de la región hipervariable I (HVR-I) de una muestra de poblaciones indígenas y afrocolombianas.
Determinar la estructura genética de poblaciones afrocolombianas con el análisis de polimorfismos microsatélites STRs
Identificar los haplogrupos de cromosoma Y, mediante análisis SNaPShot.
Establecer las relaciones filogenéticas de los grupos indígenas y afrocolombianos contemporáneos estudiados.
4. MARCO TEÓRICO
Tanto por su posición privilegiada a la salida del istmo centroamericano, como por su extensión y la variedad de ambientes, desde épocas remotas, Colombia debió ser un lugar de paso y asentamiento para numerosas poblaciones de lenguas diferentes; lugar de paso para pueblos mesoamericanos en movimiento hacia Suramérica; lugar de paso o de asentamiento para pueblos suramericanos en movimiento hacia Centroamérica o Norteamérica [51]. La dividida geografía e intrincada historia en gran parte imposible de reconstruir, produjeron una notable diversidad lingüística y una variedad perceptible tanto en lo tipológico como en lo genético, únicas a escala en todo el continente americano [51]. El conocimiento de las lenguas indígenas ha crecido, con niveles de trabajos muy variables y un trabajo de recopilación y de elaboración de datos considerable. Desde hace varios años se ha estudiado la estructura genética de la población colombiana [6, 10, 20, 34, 35, 37, 41, 43, 44, 46, 47, 52-55]., siendo hoy importante continuar el estudio de la estructura genética colombiana con el fin de aportar nuevos elementos que permitan avances en el entendimiento de la estructura genética de la población colombiana. Además de esto, complementar los hallazgos y fortalecer los resultados mediante el uso de técnicas moleculares actuales y nuevos marcadores que nos permitirán trazar eventos únicos ocurridos en la historia.
4.1. Origen y poblamiento
Hace 600.000 años, la tierra entró en una serie de eras glaciales, las cuales produjeron enormes capas de hielo que llegaron a cubrir el norte de Europa, América y Asia; el nivel del mar descendió hasta 90 metros debido a la acumulación de agua en los grandes glaciares que se formaron. En ese entonces habitaba el Homo erectus,
sucesor del Homo habilis y Homo ergaster el cual aprendió a dominar el fuego ya
hace 500.000 años y fue el primero en cocer los alimentos; también utilizaba la industria lítica tanto para el combate como para el trabajo de la madera, hueso y otros materiales [56].
Hace 200.000 años, los primeros homínidos con un cerebro tan grande como el nuestro, evolucionaron a partir del Homo erectus, el denominado hombre de
Neanderthal q u e por su desarrollo cerebral, su forma de vida y capacidad inventiva,
se encuentra más próximo del Homo sapiens que del pitecántropo. El Neanderthal es
el tercer peldaño de la evolución humana, encontrándose en 1856 restos de este en una cueva del valle por el cual lleva su nombre, valle de Neanderthal. El hombre de Neanderthal, surgió y evolucionó en Europa y medio oriente hace unos 230.000 años, hasta el pleistoceno medio y superior y culturalmente integrado en el paleolítico medio. Su cráneo era alargado y amplio, con baja estatura y complexión rasgos que denotan adaptación a climas fríos, como se puede observar actualmente en las poblaciones del Ártico [57]. Su cerebro era igual o incluso más grande que el de los hombres modernos. Un Neanderthal promedio podría llegar a tener una altura de 1,65 m, de contextura pesada, y musculatura robusta. En un periodo de aproximadamente 5.000 años, se cree convivió paralelamente en los mismos territorios europeos con el hombre de Cromagnón. Esta convivencia se ha demostrado por fósiles hallados en las cuevas de Châtelperron [58].
El análisis de restos de un Neanderthal, a partir de restos encontrados en una cueva en Feldhofer, Alemania, mostró que éste, se encuentra fuera de los límites del humano moderno; con estudios realizados en Rusia en 1999 y en Croacia en el año de 2000, se corroboraron tales hipótesis [57, 59].
El ser humano, apareció en África hace más o menos 200 mil años, el denominado
Homo sapiens arcaico u hombre de Cromañón hace 100.000 años, conviviendo con el Homo neanderthalensis casi al final de la existencia de éste. En 1868, se encontraron restos en un abrigo rocoso europeo llamado Cromagnón, de los cuales se determinó tenían más de 40.000 años, éstos tomaron el nombre del lugar. En el lugar también se han encontrado restos con 10.000 años de antigüedad, estos periodos corresponden al comienzo del paleolítico superior desde el punto de vista antropológico. El hombre de Cro-Magnon se considera el poblador de la Europa occidental de la actual especie humana el ancestro más reciente. Según la historia el ser humano llegó al próximo oriente hace 90.000 años, donde se encuentra con el hombre de Neanderthal, huyendo de la glaciación en Europa en ese momento; el homo sapiens continuó su expansión llegando hace 45.000 años a Europa occidental (Francia) y paralelamente el Homo neanderthalensis se iba retirando, empujado por el
homo sapiens hacia Portugal, Gibraltar y las mesetas altas de Croacia. Se ha postulado que el homo Neanderthalenesis no es ancestro del homo sapiens sino una especie o línea evolutiva que coexistió con el homo sapiens en la Europa occidental [57, 59].
Posteriormente y hace entre 35.000 años Ap, comienza en Europa el Paleolítico Superior, apareciendo el hombre moderno, con una estructura social avanzada, e innovación en la fabricación de útiles, evidenciándose las primeras huellas de ocupación humana en Norteamérica; al final de ese periodo y con la ocurrencia de la última glaciación, comienza la etapa paleoindia [57, 59]. Dicha datación es proveniente del reloj molecular, pero las evidencias óseas, culturales y las realizadas mediante radiocarbono se remontan a hace 40.000 años Ap.
Con respecto al poblamiento del continente americano, existen diversas teorías, entre ellas, la teoría de Ales Hrdlika, investigador norteamericano quien afirmó que América se pobló con seres humanos llegados desde Asia pasando por el Estrecho de Bering
y, desde allí, hacia todo el continente (Teoría Asiática). El antropólogo francés Paul Rivet (teoría polinésica y melanésica) [60], afirmó que al continente también llegaron seres humanos por el océano pacífico; otra teoría es la Teoría Australiana, apoyada por Mnedez Correia con respecto al poblamiento de Amé rica por los hombres a través de la ruta de la Antártida [61].
4.2. Aspectos históricos y Arqueológicos
La antigüedad de la ocupación americana de los pueblos originarios es todavía tema de estudio e investigación, sin embargo se ha aceptado para América un poblamiento cuyas evidencias arqueológicas se remontan a 14.000 años antes de la era presente [62].
Para entender de dónde provino el hombre americano y hacer posible la reconstrucción de la historia del poblamiento de América, a continuación se explicará brevemente a partir de la ocurrencia de las últimas glaciaciones y posteriores emigraciones en pequeñas oleadas sucesivas por el estrecho de Bering, describiendo desde el momento de su llegada el desarrollo evolutivo que se llevó a cabo a través de importantes periodos como lo son el Paleolítico, Arcaico y Formativo.
Los pueblos originarios llegaron al territorio americano del norte del continente, de migraciones promovidas por los cambios climáticos de la última glaciación.
Con la ocurrencia de la última glaciación, casi todo lo que ocupa América del Norte estaba cubierto por glaciares que llegaban hasta la actual frontera entre Canadá y Estados Unidos. La disminución del nivel del mar en cerca de 90 - 120 metros en el
final del Pleistoceno hace 18.000 años como ya se había mencionado anteriormente, permitió entre 18.000-14.000 años, la masa terrestre llamada Beringia, que unió el noreste de Asia con el Noroeste de Norteamérica Sin embargo el paso transberíngeo continuó siendo imposible debido a la obstrucción glaciar que cubría esa parte en ambos continentes [63]. Esta situación se mantuvo hasta hace unos 12000 – 10.000 años, momento en el cual hubo un aumento de temperatura que permitió la formación del corredor de Alberta, ésta era la vía de tránsito transcontinental. Muchos de ellos se mezclaron con otros pueblos o desaparecieron dejando huellas que hoy se conservan como un patrimonio arqueológico de los pueblos indígenas y de la Nación [11, 12].
En el territorio de los que hoy es Colombia estos primeros habitantes desarrollaron hacia los 14.000-12.000 AC sistemas de vida móvil basados en la caza y la recolección, sistemas que miles de años después ante la extinción de la megafauna, se transformarían a la explotación de medio ambientes costeros, a la adopción de la agricultura y de la residencia sedentaria. A finales del último milenio AC la vida de los antiguos pobladores contaba con un marcado desarrollo cerámico y agrícola orientado hacia los cultivos de maíz y yuca, mostrando nuevas tendencias de poblamiento (hacia las laderas de las cordilleras), así como profundas transformaciones en su sistema económico, político y social [64].
Figura 1. Áreas Arqueológicas y su cronología [64]
Colombia fue escenario de intercambio de bienes (Tairona y Muisca), de las cuales hoy en día aún se encuentran piezas de alfarería, orfebrería, recintos ceremoniales y obras hidráulicas y que lograron una avanzada organización política; algunas de éstas sociedades se encontraban desde la costa atlántica a los Andes colombianos pero también se encontraron comunidades con diferentes estilos de vida y subsistencia de la pesca y la caza [64].
Los aborígenes provienen de tres periodos que son el periodo Paleoindio, el periodo Arcaico y el periodo Formativo, periodos muy importantes, debido que, marcaron lo que serían hoy los indígenas y lo que hoy es la geografía de nuestro país [64].
28
Figura 2 Mapa de poblamiento de América
4.3. El tráfico de esclavos africanos
El tráfico de esclavos por parte de los portugueses, tuvo inicio en 1441, aunque se realizó en diferentes momentos, por parte éstos. Eran capturados para ser exportados para Portugal, España e Italia y otras partes de Europa. El tráfico de esclavos en la costa occidental africana fue por algún tiempo, monopolio de los portugueses pero posteriormente España e Inglaterra también tomaron parte en África; en África oriental el tráfico comenzó hacia 1645, especialmente en Mozambique, intensificándose hacia la segunda mitad del siglo XVIII, las relaciones comerciales dependieran de la exportación de esclavos hacia América y Golfo Pérsico. Después de la conquista de América el tráfico de esclavo aumentó extraordinariamente y entre los siglos XVI y XVIII, millones de africanos fueron arrancados violentamente de sus tierras y aldeas con destino a América e Islas del Océano Índico. El negro africano hizo su aparición en el Nuevo Mundo recién iniciada la conquista, sin embargo sólo se hizo de manera sistemática hasta cuando la explotación económica y la disminución demográfica de la población indígena fue considerable. Debido a que la mano de obra en América era insuficiente y no contaba con las condiciones deseadas, los europeos recurrieron a la mano de obra africana; con el tráfico de esclavos el hombre pasó a ser un objeto de cambio [65].
3
los años de 1640 y 1740, llegaron al puerto de Cartagena esclavos provenientes adicionalmente de Mozambique, lo cual fue confirmado por trabajos lingüísticos [67- 71], y que además, suponen la presencia de las lenguas ki-kongo y ki-mbundo habladas y lenguas bantúes de la región Congo – Angoleña sobre la lengua de Palenque de San Basilio, cerca de Cartagena de Indias [25].
4.4. Etnografía de las poblaciones estudiadas
En Latinoamérica existe gran variedad de lenguas, siendo la mayoría de estas, lenguas indígenas y muchas de ellas en vía de extinción o ya extintos, a causa de la conquista europea en el siglo XVI a su vez, con su extinción también desaparece parte de su cultura e historia regional [51]. Es importante mantener y preservar su estudio así como la lengua en sí, preservando la influencia que ejercen o han ejercido para algunas de nuestras poblaciones actuales, así como la historia de nuestros orígenes.
En las poblaciones indígenas, el término amerindio, término ambiguo derivado del término Indoamericano, es dado a los descendientes de cualquiera de los pueblos originales del continente americano (exceptuando los esquimales, cuya llegada a América fue posterior), para distinguirlos de los nuevos inmigrantes (europeos, africanos...etc.), así como de los mestizos y criollos de todas las etnias [72], Pueblos indígenas de Colombia. Los indígenas colombianos están distribuidos en más de 80 etnias.
Algunos de los pueblos indíegnas colombianos aún mantienen su propi idioma,
existiendo lenguas de las familias: Chibcha (kogui, wiwa, arhuaco, tule, chimila y u’wa),
3
4.4.1. Chimila ó Ette ennaka
Se ubica en las sabanas de San Ángel, en el departamento del Magdalena, es un pueblo amerindio que habla el idioma ette taara (lengua de la gente), familia lingüística chibcha, La producción económica de los Chimila se sustenta en la horticultura, la cacería y la pesca, complementadas con la cría de animales y aves domésticas. La actividad agrícola es complementada con la pesca, la caza y la elaboración de productos artesanales como mochilas y hamacas. [39, 73].
4.4.2. Guahibo
Pueblo indígena que habita en los llanos del Orinoco, entre los Ríos Guaviare y meta en los departamentos con presencia en el municipio de Puerto Gaitán en el departamento del meta y en Venezuela al occidente del Amazonas, Bolivia, y al sur de Apure. Forman el grupo más numeroso en la zona de la influencia del Orinoco. Algunos de los autores los clasifican dentro de la familia arawak, como emigrantes de las Guyanas, se Caracterizaban por un tránsito a la sedentarización desde la vida seminómada más tradicional. Practicando la agricultura, caza y pesca [39, 51, 72- 74].
4.4.3. Guane
Grupo indígena chibcha, habitó lo que hoy es el municipio de Los Santos, Jordán Sube, Guane y Cabrera, hasta la desembocadura del Río Fonce en el Saravita o Suarez, pertenecientes actualmente al departamento de Santander. Creaban sus propias armas, incluyendo flechas, tiraderas, garrotes y lanzas en macana; cultivaron el algodón, piña y maíz, también hábiles artesanos y eran expertos en el trabajo en hilos de algodón que ellos mismos fabricaban. Estudios anteriores como el llevado a cabo por el antropólogo Gonzalo Correal y el médico Iván Flórez, confirmaron que los restos humanos encontrados datan de 600 d.c, según datos entregados por el laboratorio Groningen de Holanda. [39, 72].
4.4.4. Guayabero
31
en riberas del Río Guaviare. Son aproximadamente 1.100 personas, que hablan una lengua de la familia guahibo. Se dedican a la caza y recolección de frutos y semillas por el bosque, pero la agricultura es fundamental para su subsistencia al lado de la pesca y la artesanía. Su cultivo principal la yuca amarga, de la cual obtienen el casabe, cultivan también el maíz, ñame, aguacate, cacao y chontaduro entre otros. Actualmente viven en casas unifamiliares, con techo de paja de hojas de palma y subsisten de la pesca en la playa [39, 73, 75].
4.4.5. Inga ó Ingano.
Es el grupo Quechua, cuyos territorios se encuentran más al norte, situados en Colombia, en el actual departamento del Putumayo, norte de Nariño, Sur de Caquetá y la bota Caucana. En cuanto a su origen, descienden de la población instalada por el Inca Huayna Cápac en Mocoa y el Valle de Simundoy (Putumayo). Se encuentra que los principales asentamientos Inga son los asentamientos Andinos y amazónicos, siendo en primer lugar agricultores y otros en menor escala ganaderos y avicultores (pavos) también practicando para complementar su alimentación la caza y pesca [39, 76].
4.4.6. Pastos
Los Pastos habitan en su mayoría en los resguardos coloniales de: Aldana, Carlosama, Córdoba, Colimba, Chiles, Mayasquer, Panam, Cumbal, Chiles, Potosí, Guachucal, Mueyamués, Ipiales, San Juan, Yaramal, Mallama, Guachavez, Túquerres y Yascual. Están ubicados en el Altiplano de Túquerrese Ipiales, en el departamento de Nariño, viven principalmente de la agricultura y de las actividades pecuarias orientadas a la producción de leche y ganado lanar. [73, 76].
4.4.7. Tukano.
Tucano, Tukano o Dasea, son un grupo étnico indígena nativo de las selvas del departamento colombiano del Vaupés y el estado del Amazonas (Brasil), frontera Colombo-Brasileña. El idioma Tucano hace parte del grupo oriental de la familia Tucano [76, 77].
4.4.8. Uitoto
33
varias regiones del Amazonas, habitan la región conocida como el medio Amazonas colombiano así como un sector importante de Perú en la frontera con Colombia. Sobreviven del procesamiento de algunos alimentos (Casabe), arepa hecha de harina de yuca Brava, también se dedican a la caza y pesca. Preuss [39, 78].
4.4.9. Yuco
Los Yuco ó yukpa. Pertenece a la familia lingüística Arawak. Viven en el Departamento del Cesar, cerca de la frontera con Venezuela, en la serranía del Perijá. Los yuko practican una agricultura de tala, quema, rotación de cultivos, junto con la caza y recolección. Se dedican a recolección de frutas, semillas, gusanos y caracoles, y caza menor de ardillas y aves [39, 79].
4.4.10. Zenú
Los Zenúes, Zenú o sinú, son un pueblo amerindio cuyo territorio ancestral está Constituido por los valles del río sinú y el San Jorge y el litoral Caribe en los alrededores del Golfo de Morrosquillo, en los actuales departamentos colombianos de Córdoba y Sucre. Sus principales actividades son la agricultura y la artesanía; cultivan el maíz, yuca, fríjol, frutales, etc. Como artesanos se destacan en el trenzado de La fibra vegetal para fabricar sombreros y otros objetos [76].
4.4.11. Paez.
Los Páez están localizados en su mayoría al sur de los Andes, en la región de Tierradentro entre los departamento de Cauca y Huila. Habitan además, en otras regiones de la vertiente oriental de la cordillera Central y del piedemonte amazónico. Están organizados políticamente en cabildos y su economía se basa en la agricultura y en la posesión de rebaños de ovejas [39, 80].
4.4.12. Guambiano.
33
4.5. Clasificación de Lenguas Indígenas de Colombia
Se encuentra un gran número de clasificaciones lingüísticas de las poblaciones indígenas colombianas, con diferentes puntos de vista y criterios. Paul Rivet [60], un importante antropólogo francés que buscó comparar la lengua que quería clasificar con familias enteras; W. Lehmann [82], M. Swadesh [83], etc. y posteriormente el norteamericano Daniel Brinton [84], quien a pesar de algunas dificultades, recopiló y examinó todos los datos disponibles en la literatura de su época, buscando clasificar a partir de datos lingüísticos. Recogía así la mejor herencia de Gilij, un padre Jesuita italiano quien realizó estudios lingüísticos y étnicos de las diversas tribus indígenas en particular en las orillas del Orinoco; discutió características de área entre idiomas no relacionados, préstamos (entre idiomas americanos y de idiomas americanos en idiomas europeos), orden de las palabras [85]. Otro aporte importante fue el del alemán Theodor Koch-Grünberg, quién reveló la existencia de muchas lenguas del oriente amazónico colombiano [86]. La precisión de su documentación y el rigor de sus comparaciones lexicales lo llevaron a constituir o a definir familias lingüísticas vigentes aún hoy en día (witoto, bora, macú, tucano) [86]. De igual forma encontramos al lingüista checo Chestmir Loukotka (1895-1958), que analizó amplia información y que le permitió posteriormente elaborar una clasificación general de las lenguas de América del sur, la más completa y detallada de todas las que se tienen a disposición. Presenta 73 troncos o cepas de lenguas ("stocks") y 44 lenguas aisladas, para América del sur y Central [87].
Algunos otros lingüistas de Norteamérica, hicieron esfuerzo por reducir las lenguas americanas mediante el uso de macro-clasificaciones, aunque con base en clasificaciones anteriores [51].
Greenberg en 1986, postuló una unidad originaria de todas las lenguas del continente americano, menos la del grupo Na – Dene y la del grupo esquimal. (Greenberg [88].
Todas estas lenguas procederían de una cepa única llamada amerindio y que posteriormente se habría fragmentado en 11 ramas:
Macro – Ge, Macro – pano, Macro – Caribe, Ecuatorial, Macro – Tucano, Andino, Chibcha – Paez, Amerindio Central, Hoka, Penutiano y Almosán – Keresiouan.
Bora, Huitoto, Yagua; Ecuatorial: Familias Macro – Arawal (Otomano, Tinigua, Guiahibo) Ketembri; Arawak (que incluye Maipure), Jíbaro – Kandoshi (que incluye Cofán y Yaruro); Macro – Tukano: Ticuna, Tucayo, Yuri; Chibchas – Paez: Familias Chibchas (Antioquia, Chibcha, Cuna, Malibú (=Chimila), Motilón (=Bari). A continuación se muestran los grupos indígenas encontrados en Colombia, incluyendo los contemplados en este estudio [89].
Jon Landaburu presenta una propuesta de clasificación de lenguas basad en criterios topológicos en busca de la reflexión acerca de la lengua, anotando que esta debe tomarse no como individuo biológico, sino como sistema simbólico e institución social [51]. Esta clasificación se presenta de la siguiente manera:
4.5.1. Lenguas aisladas:
El conglomerado Chibcha: Loukotka (1968) [87], reagrupó dentro de un tronco lingüístico Chibcha, cuyo nombre fue en honor a la lengua de más alta civilización, enumera 21 subgrupos que son: Paleochibcha, Rama, Guatuso, Dorasque, Guaymi, Cuna (que no incluye cuna), Antioquia (lenguas todas extintas), Chibcha propio, Moltilón (incluye bari), Betoi (Lenguas de Arauca y Casanare extintas), Arhuaco (Incluye , Ika o binkutua wiwa ó damana, kankuama).Malibú (incluye chimila),Andaqui (incluye andaqui extinto), Paez (incluye paez), Coconuco (incluye guambiano y totoró), Barbacoas (incluye awa o quaiquer), Sibundoy (incluye kamsá), Misquito, Matagalpa y Paya. Esta clasificación es débil en su base evidencial y es afín a las macro clasificaciones de Greenberg. Se pone en duda la inclusión de algunas lenguas como paez, grupos Coconuco, Barbacoa y Andaqui. Posteriormente se admitió la relación entre algunas lenguas y más adelante el costarricense Adolfo Constela [90] pensando que hay lenguas emparentadas crea una agrupación denominada “Paya Chibcha” un micrófilo que incluye la familia Chibcha y quedaría por consolida entonces el páez con sus dialectos, dentro de los cuales se encuentra el paniquita.
4.5.2. Tronco Lingüístico Arawak
La familia más extensa y la más difundida en el continente dentro de las cuales se incluyen: El guajiro de la península de la Guajira, el achagua, el piapoco y el baniva de la inmensa región comprendida entre el alto río negro y el Meta; el Jucuna (yucuna) del medio Caquetá [85].
4.5.3. Familia Caribe
Tercera en extensión en América del sur después de la Arawak y de la Tupi – Guraní; por la cercanía lingüística que presentan todas las lenguas Caribe, se supone que esta dispersión debió ser más antigua que al Arawak.
4.5.4. Familia Quechua
Parece que su presencia es reciente, hablándose actualmente el Inga o Ingano en el departamento de Nariño (Aponte), en el valle de Siiibundoy (Putumayo) y el departamento del Caquetá (alto río Caquetá, río Fraguas, río Yuruyacu y río Orteguita.
Otro importante lingüista es Merritt Ruhlen, un norteamericano, precursor del uso del método interdisciplinario para el conocimiento de la lingüística histórica, combinando la genética humana y la arqueología. Este lingüista colaboró con el genetista Luigi Luca Cavalli – Sforza [8].
Es uno de los más conocidos sostenedores de la hipótesis de la súperfamilia lingüística del Amerindio, como discípulo principal de la lingüística taxonomista de Joseph Greenberg y se centra en la defensa de su técnica de clasificación llamada
“Comparación Total” ó la “Comparación Multilateral”, que implica el comparar el vocabulario de los idiomas que son investigados, examinándolos por las semejanzas de sonido y significado y formulando una hipótesis de clasificación. Ruhlen mantiene que la clasificación (por medio de la comparación multilateral), es el primer paso del método comparativo; según él, toda lengua viene de alguna otra y así hasta el origen del lenguaje. Los lingüistas sistemáticos tratan de desentrañar, las relaciones entre las diferentes lenguas existentes hoy en el mundo, para determinar sus raíces comunes y poder agruparlas mejor [91].
Figura 3 Familias lingüísticas del mundo antes de Greenberg.
4.6. Población Afrocolombiana del Caribe
La primera documentación autorizando la entrada de negros a nuestro territorio, fue la instrucción dirigida por los reyes en 1501 a Don Nicolás de Oviedo, gobernador de las indias. En la conquista, Cartagena era en el siglo XVII, la más importante del Nuevo Reino. Sirvió de nexo con el Darién, y fue la puerta de entrada y salida para el intercambio con el resto del mundo. Limitaba al oriente con Santa Marta, separada por el rio grande de la magdalena, al oriente con los territorios del Darién hasta el Golfo de Urabá y el Rio Atrato, y al sur con la provincia de Antioquia, a través de límites poco definidos. Hubo conflictos en la frontera sur con otras provincias meridionales.
El relieve de Cartagena se formaba por valles y alturas poco elevadas. Los valles, llamados sabanas, comprendían las de Zamba, Zenú, Tolú, Mompox, Barranca y otros. En ellas había poblaciones grandes y pueblos pequeños habitados por europeos, criollos, españoles y gente de castas. Las sierras de Luruaca, las de María y más al sur, la serranía de San Lucas, se convirtieron en reducto de negros cimarrones (Huellas). Además de las villas y pueblos oficialmente constituidos existían en la provincia, un gran número de palenques formados por negros cimarrones, desertores de las estancias, las minas, el servicio doméstico y las galeras; la gente que constituía
los palenques propiamente dichos, estaba apartada y como se trataba de fugitivos de sus amos, vivía en son de guerra.
Figura 4 Localización Geográfica de la población del Caribe.
Abreviaturas: 1 - Barú; 2 - Tabacal; 3 - Turbaco; 4 - Canal del Dique; 5 - Mahates; 6 - Palenque de San Basilio; 7 - María; 8 - Arenal; 9 - Sierra de María; 10 - Barranca Vieja; 11 - San Miguel; 12 - Tenerife; 13 - Islas de San Bernardo; 14 - San Antero; 15 - Bahía de Cispatá; 16 - Tolú; 17 - Tolú Viejo; 18 - Lorica; 19 - Cereté; 20 - Momil; 21 - Sincelejo; 22- Chimú; 23 - San Benito Abad; 24 - Magangué; 25 - Mompox; 26 - Barranco de Loba; 27 - Caño de Loba; 28 - Cabo de La Vela; 29 - Riohacha; 30 - Gaira; 31 - Santa Marta; 32 - Villa de Ayapel; 33 - Río San Jorge; 34 - Río Nechí; 35 - Río Cauca; 36 - Cáceres; 37 - Zaragoza; 38 - Río Sinú; 39 - Río Grande de la Magdalena; 40 - Guamocó; 41 - Serranía de San Lucas; 42 -Segovia; 43 - Remedios; 44 - Río Atrato; 45 - Golfo del Darién; 46 - Panamá; 47 - Chagres; 48 - Portobelo.
Basado de [92].
Debido al conflicto de intereses de la corona española, se generó un gran interés por Cartagena ya que por su condición de puerto, por el que salían todas las mercancías del Nuevo Reino, principalmente oro, fue una de las presas más codiciadas durante toda su existencia como posesión española [93].
Nexo de unión entre el Darién, el Istmo, y el Nuevo Reino de Granada, Cartagena fue la provincia de más importancia del territorio neogranadino, no solo por ser la única salida con que contaba para el intercambio de sus productos y mercancías, sino porque ejerció desde un principio, y a través del puerto de Cartagena, el papel de flanco protector del resto del territorio. Muy hábiles en el manejo de las armas y de extraordinaria fortaleza física [65].
El indio, existía en la provincia de Cartagena, pero no abundaba en la ciudad, ya que su sitio lo ocupaba el esclavo doméstico negro. Cercanos a las riberas del rio magdalena, donde su cauce era más elevado, abundaban los indios encomendados, los cuales durante todo el siglo XVI, habían sido explotados por los encomenderos, pues además de pagar su tributo en especie, les obligaban a bogar por el Magdalena para llevar y traer a la ciudad de Cartagena los productos que dichos encomenderos deseaban. El indio no tenía trato con el negro, y se consideraba superior a éste racial y socialmente. Es de destacar igualmente en la lucha contra los cimarrones, el gran papel que desempeñaron los indios Chimilás, habitantes del pueblo de San Ángelo en la provincia de Santa Marta, muy hábiles en el manejo de las armas y de extraordinaria fortaleza física [25, 94].
El negro fue llegando al puerto cartagenero en grandes cantidades y desde allí distribuido a todo el interior. La procedencia de los negros bozales del siglo XVII, quienes recibían este nombre que se designaba a los recién llegados desde África, que no comprendían nada en referencia al idioma; éstos africanos se caracterizaban por ser más fuertes y resistentes al trabajo), les hacía dividirse principalmente en las castas Arará (actual Dahomey), mina (de la factoría de San Jorge de Mina), Popo (tribu situada en la frontera de Togo y Dahomey. Los primeros esclavos negros integraron el séquito de los conquistadores e incluso participaron en las batallas [95]. Eran negros cristianizados, "ladinos", que hablaban castellano y que incluso podían haber nacido en la Península, hijos de otros esclavos; ellos pertenecían a todas las etnias africanas utilizadas en las Américas como esclavos.
Estos fueron los esclavos de los primeros siglos y cuyas condiciones especiales facilitaron el comercio esclavista. En primer lugar, se aclimataban fácilmente a las regiones de clima templado; además, tenían fama de ser dóciles y serviles y trabajadores fuertes; por último, la presión de la Iglesia y la Corona en contra de la explotación de los indios, a quienes se consideraba súbditos que debían ser
![Figura 55 Reduced Median Haplogrupo E Presente Estudio + Gomes 2010 [3]](https://thumb-us.123doks.com/thumbv2/123dok_es/4088801.23628/180.595.66.479.61.669/figura-reduced-median-haplogrupo-e-presente-estudio-gomes.webp)