Bogotá, Marzo de 2006
Señores
PONTIFICIA UNIVERSIDAD JAVERIANA Ciudad
Estimados Señores:
Yo Estefanía Guevara Vega, identificada con CC. No 46380317 autor del trabajo de grado
Titulado EFECTO DEL CLOROFILIN SOBRE LA PROLIFERACION DE CÉLULAS DE FIBROSARCOMA HUMANO HT1080, presentado como requisito para optar al título de Bióloga en el año de 2006 autorizo a la Universidad Javeriana a:
a) Reproducir el trabajo en medio digital o electrónico con el fin de ofrecerlo para la consulta en la Biblioteca General. Si
b) Poner a disposición para la consulta con fines académicos, en la página web de la Facultad, de la Biblioteca General y en redes de información con las cuales tenga convenio la Universidad Javeriana. Si
c) Enviar el trabajo en formato impreso o digital, en caso de que sea seleccionado para participar en concursos de trabajos de grado. Si
d) Distribuir ejemplares de la obra, para la consulta entre las entidades educativas con las que la facultad tenga convenio de intercambio de información, para que este sea consultado en las bibliotecas y centros de documentación de las respectivas entidades. No
e) Todos los usos, que tengan finalidad académica. Si
Los derechos morales sobre el trabajo son de los autores de conformidad con lo establecido en el artículo 30 de la Ley 23 de 1982 y el artículo 11 de la Decisión Andina 351 de 1993, los cuales son irrenunciables, imprescriptibles, inembargables e inalienables. Atendiendo lo anterior, siempre que se consulte la obra, mediante cita bibliográfica se debe dar crédito al trabajo y a su(s) autor(es). Este documento se firma, sin perjuicio de los acuerdos que el autor(es) pacte con la Unidad Académica referentes al uso de la obra o a los derechos de propiedad industrial que puedan surgir de la actividad académica.
___________________________ Estefanía Guevara Vega
AUTOR
Apellidos Nombres
Guevara Vega Estefanía
DIRECTOR
Apellidos Nombres
Díaz Germán Darío
TRABAJO PARA OPTAR POR EL TÍTULO DE: Bióloga
TÍTULO COMPLETO DEL TRABAJO: Efecto del clorofilin sobre la proliferación de células de fibrosarcoma humano HT1080
SUBTÍTULO DEL TRABAJO: Efecto del Clorofilin sobre células HT1080 FACULTAD: Ciencias
PROGRAMA: Carrera X Especialización ____ Maestría ____ Doctorado ____ NOMBRE DEL PROGRAMA: Biología
CIUDAD: BOGOTA AÑO DE PRESENTACIÓN DEL TRABAJO: 2006 NÚMERO DE PÁGINAS 83
TIPO DE ILUSTRACIONES: __ Ilustraciones
__ Mapas __ Retratos
X Tablas, gráficos y diagramas __ Planos
__ Láminas __ Fotografías
DESCRIPTORES O PALABRAS CLAVES: Apoptosis, células de fibrosarcoma humano HT1080, ciclo celular, clorofilin, proliferación celular.
EFECTO DEL CLOROFILIN SOBRE LA PROLIFERACION DE CÉLULAS DE FIBROSARCOMA HUMANO HT1080
ESTEFANÍA GUEVARA-VEGA
TRABAJO DE GRADO Presentado como requisito parcial
Para optar al título de
Bióloga
PONTIFICIA UNIVERSIDAD JAVERIANA FACULTAD DE CIENCIAS
CARRERA DE BIOLOGIA Bogotá, D. C.
NOTA DE ADVERTENCIA
Artículo 23 de la Resolución N° 13 de Julio de 1946
EFECTO DEL CLOROFILIN SOBRE LA PROLIFERACION DE CÉLULAS DE FIBROSARCOMA HUMANO HT1080
ESTEFANÍA GUEVARA-VEGA
APROBADO
________________________ Germán Darío Díaz, M.D., Ph.D.
Director
________________________ ________________________ Jorge Robles, Ph.D Adriana García, M. Sc.
EFECTO DEL CLOROFILIN SOBRE LA PROLIFERACION DE CÉLULAS DE FIBROSARCOMA HUMANO HT1080
ESTEFANÍA GUEVARA-VEGA
APROBADO
DEDICATORIA
A mi Papá por sus bendiciones y fidelidad hacia mí siempre.
AGRADECIMIENTOS
A Dios, quien me infundió fuerzas a lo largo de la realización del proyecto y hoy me permite terminarlo.
Al Dr. Germán Darío Díaz, director del trabajo, por su paciencia, sus enseñanzas y su valioso apoyo en la realización de esta investigación.
Al Dr. Octavio Alberto González, miembro del Centro de Investigaciones Odontológicas (CIO) de la Facultad de Odontología de la Pontificia Universidad Javeriana, por su apoyo y por compartir conmigo su conocimiento sobre cultivos celulares.
A todo el personal del CIO por permitirme llevar a cabo la parte experimental del proyecto en su laboratorio.
Al grupo de Inmunología de la Facultad de Ciencias de la Pontificia Universidad Javeriana por su colaboración.
TABLA DE CONTENIDO
1. Introducción 1
2. Marco Teórico 2
2.1Ciclo Celular 2
2.1.1 Control del Ciclo Celular 4
2.1.2 Muerte celular 8
2.1.2.1 Apoptosis o Muerte Celular Programada 9
2.2 Biología del Cáncer 15
2.2.1 Iniciación, Promoción y Progresión del Cáncer 16
2.3 Sarcomas de Tejidos Blandos 21
2.3.1 Fibrosarcoma 21
2.4 Clorofilin 24
3. Formulación del problema y Justificación 28
3.1 Formulación del problema 28
3.2 Pregunta de Investigación 29
3.3 Justificación de la Investigación 30
4. Hipótesis 30
5. Objetivos 31
5.1 Objetivo General 31
5.2 Objetivos Específicos 31
6. Materiales y Métodos 31
6.1.1 Población de estudio y muestra 31
6.1.2 Variables del estudio 32
6.2 Metodología 32
6.2.1 Cultivo Celular 32
6.2.2 Tratamiento de las Células con Clorofilin 33
6.2.3 Determinación de la Viabilidad Celular por MTS 34
6.2.4 Células Adheridas y Células Flotantes 34
6.2.5 Análisis de Morfología Celular por Microscopía de Luz 35 6.2.6 Análisis de Ciclo Celular por Citometría de Flujo 36
6.3 Recolección de la Información 37 6.4 Análisis de la Información 37 7. Resultados 38
7.1 Análisis de la Viabilidad celular por MTS 38 7.2 Células Adheridas y Células Flotantes 41
7.3 Análisis de Morfología Celular por Microscopía de Luz 44
7.4 Análisis de Ciclo Celular por Citometría de Flujo 45
8. Discusión 50
9. Conclusiones 60
10.Recomendaciones 60
INDICE DE TABLAS
1. Diferencias entre Apoptosis y Necrosis 9 2. Valores promedio de absorbancia de células HT1080 expuestas
a diferentes concentraciones de CHL durante 24, 48 y 72 horas 39 3. Porcentaje de células adheridas y células flotantes a diferentes
INDICE DE FIGURAS
1. Fases del Ciclo Celular 3
2. Carcinogénesis 20
3. Estructura de la Clorofila 24
4. Estructura del Clorofilin 25
5. Viabilidad celular en Células HT1080 expuestas a CHL durante
24, 48 y 72 horas 40
6. Porcentaje de Células flotantes y Células adheridas expuestas
a CHL durante 24, 48 y 72 horas 43 7. Efecto del CHL sobre el Ciclo Celular de Células HT1080 hasta
72 horas de Tratamiento 49
8. Modelo propuesto de Acción del Clorofilin sobre Células
Resumen
Abstract
1. Introducción
La tasa de incidencia y mortalidad del cáncer se ha incrementado en las últimas décadas en nuestro país, constituyéndose en la segunda causa de muerte de la población después de las enfermedades cardiovasculares. Los tratamientos actuales contra el cáncer no son del todo satisfactorios y en muchas ocasiones alteran la calidad de vida del paciente. Si bien, el tratamiento de los sarcomas de tejidos blandos ha evolucionado de forma considerable en los últimos años, siendo la cirugía la modalidad terapéutica más efectiva, la quimioprevención o utilización de agentes químicos naturales o sintéticos para revertir, suprimir o impedir el proceso de la carcinogénesis, es una alternativa muy válida. El clorofilin, un derivado hidrosoluble de la clorofila, es una mezcla de sales de sodio y cobre que exhibe una potente actividad antimutagénica in vitro y ha sido reportado como anticarcinógeno en varios modelos animales. Con el uso de este agente han sido demostrados efectos benéficos sobre diferentes tipos de células tumorales, por lo cual se ha extendido su uso.
2. Marco Teórico
2.1 Ciclo Celular
El ciclo de división celular constituye un proceso básico en la génesis de nuevas células (Junqueira y Carneiro 1997). Éste es un conjunto ordenado de eventos que culmina con el crecimiento de la célula y la división en dos células hijas (Stein et.al, 2002). El ciclo celular se divide en dos periodos, uno denominado mitosis, en el que los cromosomas se condensan haciéndose visibles al microscopio óptico y que finaliza con los procesos de segregación y división celular (Nigg 2001) y otro, que comprende la mayor parte del ciclo celular denominado interfase por transcurrir entre dos mitosis. La interfase se caracteriza por ser un periodo de gran actividad, en el que se llevan a cabo y en forma ordenada los eventos que conducen a la división celular. La interfase comprende a un periodo conocido como G1, que transcurre entre el final de la mitosis del ciclo anterior y el comienzo de la síntesis de DNA, a un periodo conocido como fase S, en el cual ocurre la síntesis del DNA y a una fase G2, intervalo entre el fin de la fase S y el comienzo de la mitosis (Stein et.al, 2002) (Figura 1).
Figura 1. Fases del Ciclo Celular. En esta grafica se observan las diferentes fases del ciclo celular, G1, S, G2 y M. El estado G1 quiere decir "GAP 1"(Intervalo 1). El estado S
representa "Síntesis". Este es el estado cuando ocurre la replicación del DNA. El estado G2
representa "GAP 2"(Intervalo 2). El estado M representa "mitosis", y es cuando ocurre la
división nuclear (los cromosomas se separan) y citoplasmática (citocinesis).
2.1.1 Control del Ciclo Celular
El ciclo celular es un proceso ordenado que se encuentra regulado por una serie de proteínas denominadas ciclinas, Cinasas Dependientes de Ciclinas (CDKs) y por los complejos que se forman entre ambas, CDK-ciclina (Stein et.al, 2002). Las ciclinas controlan los cambios que conducen a la división celular formando complejos cinasa-ciclina los cuales fosforilan una serie de proteínas involucradas en la replicación del DNA, como la proteína supresora de tumores del Retinoblastoma (Rb), y en otros procesos del ciclo celular como en la formación de husos mitóticos (Nahle et.al., 2003). Hay dos clases principales de ciclinas: las ciclinas G1 (D y E), que se unen a CDKs durante la fase G1 del ciclo celular y son necesarias para el inicio de la fase S, y las ciclinas mitóticas (A y B) las cuales se acumulan gradualmente durante la fase G2 y se unen a CDKs formando el complejo cdk-ciclina también llamado Factor Promotor de la Mitosis (MPF) el cual es necesario para la entrada en mitosis (Stein et.al, 2002).
específicos de serina y treonina y por la unión de los denominados inhibidores de cinasas dependientes de ciclinas (CKIs). Las CDKs actúan cuando son activadas por la ciclinas fosforilando moléculas importantes para la división celular como la proteína del Retinoblastoma (Rb) (Sears y Nevins, 2002; Stein et.al, 2002).
Estos complejos CDK-ciclina pueden inducir la interrupción del ciclo celular cuando existe algún daño genético. En este caso, los mecanismos de control transcripcional de los complejos CDK-ciclina inducen la interrupción del ciclo celular hasta que el daño se corrige. Se ha identificado que esta interrupción de la proliferación es mediada en mamíferos por proteínas como p27 que evita el ensamblaje y activación del complejo CDK2-E, p16 y p21 (Koff et.al, 1993; Poliak et.al, 1994).
La interrupción del ciclo celular se da en puntos de regulación específicos muy importantes que se encuentran en las diferentes fases de éste y que garantizan que las células se dividan cuando las condiciones son óptimas (Kastan y Bartek 2004). La existencia de éstos puntos control del ciclo celular es clave, pues permiten que todo el proceso tenga lugar cuando la célula está correctamente preparada mediante la comprobación de que las condiciones necesarias para iniciar una nueva fase se han dado y sirven de freno durante el ciclo, asegurando que una fase no se inicia antes de que la anterior haya finalizado (Kastan y Bartek 2004).
supresor de tumores Rb que regula la proliferación y supervivencia celular (Hanahan y Weinberg 2000; Sears y Nevins 2002). La proteína Rb, de peso molecular 110 kDa, regula la proliferación celular, en parte, modulando la actividad de los factores de transcripción E2F y DP1 (Hanahan y Weinberg 2000). Cuando está hipofosforilada, Rb bloquea la progresión del ciclo celular. Ello se debe a que se une e inhibe a varias proteínas, entre ellas la familia de factores de transcripción E2F, los cuales controlan la expresión de genes esenciales para la síntesis de DNA (Nahle et.al., 2003). Tras su fosforilación, Rb ya no es capaz de unirse y bloquear a los factores de trascripción E2F los cuales se expresan, permitiendo la progresión del ciclo celular (Sears y Nevins 2002). Así, cuando hay presencia de factores de crecimiento apropiados, las células pasan el punto de restricción y entran en la fase S del ciclo celular, donde tiene lugar la síntesis de DNA y duplicación de los cromosomas (Stein et.al, 2002). Durante la fase S se regulan los eventos asociados con la replicación del DNA, la biosíntesis de histonas y la fidelidad del proceso de ensamblaje de la cromatina (Stein et.al, 2002).
Por otra parte, si no hay factores de crecimiento apropiados disponibles en G1, se frena la progresión a través del ciclo celular en el punto de restricción y las células entran en un estado quiescente llamado G0. Las células G0 son metabólicamente activas, aunque ellas cesan el crecimiento, la división y reducen la tasa de síntesis de proteínas (Kastan y Bartek 2004).
las decisiones celulares de detener el ciclo celular permitiendo la reparación del DNA dañado o conduciendo a la muerte celular (Oren 1999; Vogelstein et.al., 2000). Cuando hay daños en el DNA, por ejemplo por estrés celular, radiación gamma y/o fármacos genotóxicos, los niveles de la proteína p53 se elevan y las células en proliferación se detienen en la fase G0/G1 del ciclo celular proporcionando un lapso de tiempo para que se verifique la reparación del DNA antes de que éste sea replicado (revisado en Lutz y Nowakowska 2002). De mantenerse las concentraciones elevadas de p53 se desencadena la muerte celular programada o apoptosis por inducción de la expresión de Bax, miembro pro-apoptótico de la familia de proteínas Bcl-2. Sin embargo, se conoce que la sobreexpresión de miembros anti-apoptóticos de esta familia de proteínas, contrarresta el efecto apoptótico de p53 (Blagosklonny 2000).
2.1.2 Muerte Celular
La muerte celular es un mecanismo importante para mantener la homeostasis celular. Los estudios de Kerr y colaboradores (1972) proveen evidencia de que existen al menos dos formas diferentes de muerte celular. Una de estas, es la apoptosis, que, se caracteriza por una reducción del tamaño celular, formación de
espacio extracelular y la remoción de estas células no causa respuesta inflamatoria (Revisado en Guimaraes y Linden, 2004). La otra forma de muerte celular es la necrosis, una forma de degeneración rápida y algunas veces abrupta que se
[image:24.612.113.542.412.550.2]caracteriza por un abultamiento del citoplasma, la destrucción de los organelos celulares y el rompimiento de la membrana plasmática, que conduce a la liberación de los contenidos intracelulares generando una respuesta inflamatoria (Revisado en Guimaraes y Linden, 2004). A diferencia de la necrosis, la apoptosis usualmente requiere energía de la célula en forma de ATP y es regulada genéticamente (Tabla 1) (Kerr et.al., 1972). La necrosis puede ser causada por toxinas, radiación, trauma e hipoxia entre otras (Revisado en Guimaraes y Linden, 2004).
Tabla 1. Diferencias entre Apoptosis y Necrosis (Kerr et.al., 1972).
APOPTOSIS NECROSIS
• Afecta a células individuales • Afecta extensas poblaciones celulares
• No induce liberación de enzimas proteolíticas
• Induce liberación de enzimas proteolíticas
• No induce daño tisular • Induce daño tisular
• No causa respuesta inflamatoria • Causa respuesta inflamatoria
• Requiere energía (ATP) • No requiere energía
• Regulada genéticamente • No es regulada genéticamente
2.1.2.1 Apoptosis o Muerte Celular Programada
Este termino resalta el carácter fisiológico de la apoptosis, pues implica que para que un organismo funcione adecuadamente, no solo debe tener la capacidad de producir nuevas células sino también la habilidad de eliminar las que ya no son necesarias (revisado en Sánchez-Torres y Vargas 2003). La muerte celular programada juega un papel importante en el desarrollo y en la homeostasis celular. En organismos multicelulares es necesario que el número de células de un determinado órgano se mantenga constante para que conserve su morfología y correcto funcionamiento.
Enfermedades degenerativas humanas (e.j Alzheimer y Parkinson), autoinmunes (e.j artritis reumatoide), virales (e.j infección por VIH) y cáncer, se caracterizan por presentar alteraciones en la ejecución del programa de muerte celular programada constituyéndolo como un componente importante del desarrollo normal de un organismo (Guimaraes y Linden 2004).
Durante el proceso de la apoptosis las células sufren una serie de cambios morfológicos. El primer cambio se observa a nivel de la membrana celular, donde se
definidos. La reorganización y condensación de la cromatina está acompañada por el clivaje del DNA (Kroemer et.al., 1997; Chang y Yang 2000). Después, el núcleo se fragmenta, y las protuberancias en la superficie celular se desprenden envueltas en su membrana, formando cuerpos apoptóticos de forma esférica u ovalada. El número, tamaño y composición de los cuerpos apoptóticos derivados de una célula puede variar ampliamente. Algunos contienen uno o más fragmentos nucleares y el citoplasma condensado, mientras que otros contienen elementos citoplasmáticos (Revisado en Sánchez-Torres y Vargas 2003). Una característica importante de la apoptosis es la remoción de las células apoptóticas que se producen durante el proceso. Los responsables de retirar la mayoría de las células apoptóticas son los fagocitos profesionales, pero existen evidencias de que fagocitos no profesionales, como las células dendríticas, epiteliales y fibroblastos, también participan en la remoción de células apoptóticas (Gregory 2000).
2000). Una vez activas, las caspasas producen la hidrólisis a partir de residuos de ácido aspártico en la proteína sustrato. Así, la activación inicial de una caspasa provoca una reacción en cadena que conduce a la activación de otras caspasas y a la muerte de la célula. Por lo tanto, la regulación de la activación de las caspasas es fundamental para determinar la supervivencia celular (Thornberry y Lazebnik 1998).
Según su función, las caspasas se dividen en 2 grupos. El primer grupo consiste en las caspasas iniciadoras (caspasa-2, -8 y -9), cuya función es la de activar a otras caspasas y el segundo grupo consiste en las caspasas ejecutoras (caspasa-3, -6 y -7), que son responsables de la interacción con otras moléculas que desencadenan la apoptosis (Mehmet 2000; Chang y Yang 2000; Elinos-Baez et.al, 2003).
En humanos, la caspasa-1, conocida como la enzima convertidora de interleuquina (ICE), fue el primer miembro descrito de esta familia de cisteina proteasas, que de forma casi exclusiva degradan sus sustratos en residuos de ácido aspártico (Asp). Desde el descubrimiento inicial de ICE, se han descrito hasta 13 moléculas homólogas conocidas con el nombre genérico de caspasas (Thornberrry y Lazebnik 1998). Las caspasas funcionan en forma de cascada, en la cual una caspasa iniciadora es activada por su interacción con el adaptador de caspasa. Una vez activas, las caspasas iniciadoras activan a una o más caspasas efectoras o ejecutoras las cuales, a su vez, clivan varias proteínas celulares conduciendo a la muerte celular (Chang y Yang 2000; Nahle et.al., 2003).
inicia en la mitocondria y es mediada por estrés celular o por lesión en el DNA. Estas dos vías son denominadas extrínseca e intrínseca, respectivamente (Mehmet 2000).
La vía extrínseca ocurre a nivel de la membrana celular y se inicia por la unión de un ligando con su receptor transmembranal, llamado receptor de muerte (DR) como Fas, receptor del factor de necrosis tumoral (TNFR) y el ligando inductor de la apoptosis (TRAIL) entre otros (Reed 2000). Los DR, que se localizan sobre la membrana celular, están compuestos por tres proteínas homólogas, con una estructura extracelular, una transmembranal y una intracelular en la que se encuentran los dominios de muerte (DD) los cuales se agrupan en trímeros (Roseto et.al., 1999). La unión del ligando con su DR, activa a las caspasas iniciadoras, que a su vez, activan por proteolísis a las caspasas ejecutoras o efectoras, conduciendo a la apoptosis (Chang y Yang 2000). Por tanto, los DR juegan un papel importante en la apoptosis ya que pueden activar la cascada de caspasas en pocos segundos después de la unión del ligando y el receptor (Reed 2000).
anti-apoptóticos como Bcl-2 y Bcl-xL la previenen (Korsmeyer et. al., 2000 Hengartner 2000). Entre las proteínas mitocondriales liberadas al citosol tras un estímulo apoptótico, se encuentran el citocromo c, Smac/Diablo y el Factor Inductor de la Apoptosis (AIF) (Adrain y Martin 2001). Una vez en el citosol, el citocromo c puede unirse al factor activador de las proteínas pro-apoptóticas (Apaf-1) y una vez unido y en presencia de dATP o ATP se forma el complejo conocido como apoptosoma, el cual recluta y activa a la procaspasa-9, la cual a su vez puede activar a las caspasas 3, 6 y 7 (Kroemer et.al., 1998).
El citocromo c es una proteína de 15-17 kDa que en condiciones normales se encuentra localizada en el espacio intermembranal de la mitocondria. Su liberación al citosol es activada por varios tipos de estrés celular, entre los que se incluyen drogas citotóxicas, deprivación de factores de crecimiento y lesión en el DNA (Chang y Yang 2000) y es regulada por proteinas miembros de la familia Bcl-2 (Adams y Cory 1998). Las proteínas anti-apoptóticas como Bcl-2 y Bcl-xL la previenen mientras que proteínas pro-apoptóticas como Bax y Bid la promueven (Gross et.al., 1999). Al ser liberado al citosol, el citocromo c, inicia la activacion de las caspasas (Liu et.al., 1996).
Otra de las proteínas liberadas al citosol, es el Factor Inductor de la Apoptosis (AIF). Después de que la célula recibe una señal o un estimulo apoptótico, la membrana mitocondrial externa se permeabiliza y el AIF se transloca al citosol y al núcleo en donde induce la condensación de la cromatina y la fragmentación del DNA (Candé et.al., 2002). El AIF y las caspasas cooperan en la cascada de la muerte celular, y su contribución puede depender del estimulo inductor de la apoptosis y tal vez del tipo celular. En muchos casos se requiere que se de la neutralización simultanea de las caspasas y el AIF para prevenir eventos de la apoptosis como la condensación de la cromatina (Candé et.al., 2002).
2.2 Biología del Cáncer
constituido por tejido conectivo y vasos sanguíneos (Cruz-Coke 1997; revisado en Robbins 2004).
Las células tumorales se caracterizan por tener una morfología alterada que refleja modificaciones en su proceso de diferenciación (Matilla 2000). La diferenciación celular en un tumor o neoplasia hace referencia al grado de semejanza de las células que lo componen a las células normales de donde proceden, tanto morfológica como funcionalmente. Por ejemplo, los tumores bien diferenciados, son los compuestos por células que recuerdan a las células maduras normales del tejido del que proceden y los tumores poco diferenciados o indiferenciados están compuestos por células de aspecto primitivo, no especializadas. Generalmente los tumores benignos son bien diferenciados y los malignos, por el contrario, varían desde bien diferenciados a indiferenciados (revisado en Robbins 2004).
2.2.1 Iniciación, Promoción y Progresión del Cáncer
La carcinogénesis, es un proceso que lleva a que una célula normal adquiera alteraciones en su DNA, aumento en su proliferación y pérdida en su diferenciación, generando un tumor, y que se caracteriza por estar dividido en 3 etapas: iniciación tumoral, promoción tumoral, y progresión tumoral (Balmain y Harris 2000; Yuspa 2000).
consecuentemente a la iniciación, se refiere a la expansión clonal de células iniciadas y precisa la exposición crónica a diferentes factores promotores, es decir, factores que no tengan propiedades mutagénicas y, que por sí solos, sean incapaces de inducir neoplasias (revisado en Balmain y Harris 2000; Yuspa 2000). Los hidrocarburos policíclicos aromáticos desprendidos en la combustión de cigarrillos y tabacos representan un ejemplo de elementos exógenos muy ricos tanto en factores iniciadores como en promotores, razón por la cual han sido empleados en modelos experimentales de carcinogénesis (Bernal Balaez 1996). El efecto de los factores promotores es estimular la división celular de una célula que sufrió previamente una mutación por la acción de un agente iniciador. La célula iniciada responde ante la acción del promotor con un crecimiento distinto a las células normales (Cotran et.al, 1990; Balmain y Harris 2000).
actúan directamente sobre el DNA de la célula (Loeb 1994; revisado en Balmain y Harris 2000).
Figura 2. Carcinogénesis. La carcinogénesis es un proceso que lleva a que una célula normal adquiera alteraciones en su DNA, aumento en su proliferación y pérdida en su diferenciación, generando un tumor, y que se caracteriza por estar dividido en 3 etapas: iniciación tumoral, promoción tumoral y progresión tumoral. La activación de los oncogenes y la inactivación de los genes supresores de tumores resultan
2.3 Sarcomas de Tejidos Blandos
Los sarcomas de tejido blando son un grupo heterogéneo de neoplasias de tejido conectivo que se originan en los tejidos blandos por todo el cuerpo (Weiss y Goldblum 2001). En Colombia, representan el 15% aproximadamente de los tumores en pacientes pediátricos y en adultos corresponden al 1% de las neoplasias (Ministerio de Salud 2001). Los sarcomas se clasifican según su tejido de origen; los rabdomiosarcomas, tumores de músculo estriado y los sarcomas no diferenciados, representan más de la mitad de todos los sarcomas de tejido blando infantiles. El resto de los sarcomas de tejido blando no rabdomiosarcomatosos representan aproximadamente el 3% de todos los tumores en pacientes pediátricos (Miser et.al 1997). Este grupo heterogéneo de tumores incluye las neoplasias de músculo liso (leiomiosarcoma), de tejido conectivo (fibroso y adiposo), de tejido vascular (vasos sanguíneos y linfáticos) y del sistema nervioso periférico (Weiss y Goldblum 2001). Los sarcomas sinoviales, los fibrosarcomas y los neurofibrosarcomas predominan en los pacientes pediátricos (Weiss y Goldblum 2001).
2.3.1 Fibrosarcoma
Dentro de los sarcomas de tejidos blandos, se encuentra el fibrosarcoma, tumor maligno de origen mesenquimal caracterizado histológicamente por una población predominante de células epiteliales dispuestas en fibras incrustadas en un estroma fibrótico extensamente hialinizado (Chow et.al, 2004). El fibrosarcoma exhibe diferenciación fibroblástica y muestra una fibrosis extendida evidente. El
tumor se presenta principalmente en musculatura profunda y está frecuentemente asociado con la banda adyacente al periostio. El fibrosarcoma se caracteriza por una proliferación de las células epiteliales con escaso o sin citoplasma dispuesto en fibras y asociado con una esclerosis hialina prominente del estroma (Chow et.al, 2004).
El fibrosarcoma fue diagnosticado con más frecuencia en el pasado pero ahora se distingue histológicamente de lesiones similares como tumores desmoides, histiocitoma fibroso maligno, schwannoma maligno y osteosarcoma de alto grado (Antonescu et.al, 2000).
Según características histológicas que permiten su identificación, los fibrosarcomas se dividen en fibrosarcoma mixoide (mixofibrosarcoma), sarcoma fibromixoide de bajo grado y fibrosarcoma epitelial escleroso (Weiss y Goldblum 2001). Histológicamente, la mayoría de los fibrosarcomas tiene en común un patrón de crecimiento fasciculado consistente en células fusiformes o delgadas en forma de espiga con escaso citoplasma y que están separadas por fibras de colágeno dispuestas de manera paralela (Weiss y Goldblum 2001). El grado histológico de los fibrosarcomas se basa principalmente en el grado de celularidad, grado de diferenciación celular, número de mitosis, cantidad de colágeno producido por las células tumorales y el grado de necrosis (Inwards y Unni 1995; Weiss y Goldblum 2001).
El tumor puede ocurrir en cualquier tejido blando del cuerpo pero es más común en los tejidos blandos profundos de las extremidades inferiores, particularmente en el muslo y las rodillas, seguido por las extremidades superiores y
el tronco (Weiss y Goldblum 2001). Generalmente la piel que rodea el tumor está intacta aunque la mayoría de neoplasias localizadas superficialmente que crecen rápidamente o han sufrido trauma pueden resultar en ulceración de la piel (Weiss y Goldblum 2001). El fibrosarcoma puede ocurrir a cualquier edad pero es mas frecuente entre los 30 y los 55 años (Weiss y Goldblum 2001). La edad media en la literatura está entre los 39.4 y los 47.7 años (Pack y Ariel 1952; Pritchard et.al, 1977). La mayoría de estudios han reportado una incidencia ligeramente mayor en hombres que en mujeres, por ejemplo, Scott et.al (1989) en un estudio de 132 casos encontró que el 61% de los pacientes eran hombres. La mayoría de los pacientes consulta por presentar masas palpables entre los 3 y los 8 centímetros que durante la fase inicial crecen lentamente y causan dolor únicamente en la tercera parte de los casos (Weiss y Goldblum 2001). Además, la mayoría de los pacientes carece de manifestaciones sistémicas exceptuando la perdida de peso en aquellos en los que los tumores son de gran tamaño y han hecho metástasis (Weiss y Goldblum 2001).
El tratamiento del fibrosarcoma más utilizado es la cirugía radical del tumor, que a veces requiere la amputación o desarticulación del miembro donde se localiza, debido a los altos índices de recidiva y a su capacidad metastásica por lo que la radioterapia y la quimioterapia son utilizadas de forma adyuvante para la cirugía (Weiss y Goldblum 2001).
2.4 Clorofilin
La clorofila y sus derivados hidrosolubles son constituyentes de la dieta humana y son efectivos anticarcinógenos en varios modelos animales (Dashwood et.al, 1998). Los derivados sintéticos o semisintéticos de los tetrapirroles son ampliamente usados en fototerapia contra el cáncer, la psoriasis y degeneración macular causada por la edad (Lane 2003). El clorofilin (CHL) (C34H31CuN4Na3O6) es un derivado hidrosoluble de la clorofila, básicamente, es una mezcla de sales de sodio y cobre que al igual que la clorofila exhibe una potente actividad antimutagénica in vitro y ha sido reportado como anticarcinógeno en varios modelos animales (Dashwood, 1997; Negishi et.al, 1997; Dashwood et.al, 1998). Al igual que la clorofila, la molécula de clorofilin también está compuesta por anillos tetrapirrólicos y se diferencia de la clorofila en que su átomo central es cobre y en que no posee el extremo fitol (Fahey et.al, 2005) (Figura 3 y 4).
Figura 3. Estructura de la Clorofila. La molécula de clorofila es grande y está formada en su mayor parte por carbono e hidrógeno; posee un átomo de magnesio en el centro, rodeado
por un grupo de átomos que contienen nitrógeno llamado anillo de porfirinas. El anillo de
porfirina es un tetrapirrol, con cuatro anillos pentagonales de pirrol enlazados para formar un
anillo mayor que es la porfirina. La clorofila posee una cadena larga llamada fitol que es una
cadena hidrocarbonada con restos de metilo (-CH3) a lo largo y que como todas las cadenas
[image:41.612.233.404.264.395.2]orgánicas basadas sólo en C e H, tiene un carácter hidrofóbico.
Figura 4. Estructura del Clorofilin. La molécula de clorofilin está formada en su mayor parte por carbono e hidrógeno y está compuesta por anillos tetrapirrólicos su átomo central es
cobre es grande y a diferencia de la clorofila no posee el extremo fitol, dándole un carácter
hidrofílico.
Los efectos de la clorofila y sus derivados han sido probados en laboratorios de cultivos celulares y en animales (Sarkar et.al, 1994; Chernomorsky et.al, 1999). Un estudio realizado en Qidong, China provee evidencia de los efectos
anticarcinógenos del CHL ya que se observa que el CHL puede reducir la cantidad de casos de cáncer de hígado, el cual proviene de la exposición a aflatoxinas en las comidas (maíz, nueces, soya y salsa de soya) (Egner et.al, 2001; Egner et.al, 2003). Previamente, se ha demostrado que el clorofilin inhibe significativamente la actividad mutagénica de gran variedad de xenobióticos actuando directamente como antioxidante y formando complejos con los carcinógenos facilitando de este modo su excresión (Dashwood et.al, 1998). El CHL puede disminuir la biodisponibilidad de los carcinógenos presentes en la dieta impidiendo su absorción y transportándolos a través de la materia fecal (Kensler et.al, 1998; Breinholt et.al, 1999).
Varios estudios sugieren que el CHL actúa como una “molécula interceptora” entre la formación de complejos moleculares con carcinógenos como la aflatoxina B1 (Negishi et.al, 1997). Como un “agente bloqueador”, el CHL forma complejos moleculares con compuestos aromáticos carcinógenos como las aminas heterocíclicas, carbonos aromáticos policíclicos y aflatoxinas y reduce su actividad carcinógena (Dashwood 1997; Dashwood 2002). Se ha demostrado que el CHL inhibe la actividad genotóxica y carcinógena de la aflatoxina B1 (AFB1) en trucha arcoiris (Breinholt et.al, 1995).
El CHL inhibe la mutagenicidad de 2-amino-3-metilimidazol[4,5-f]quinolina (IQ), 3-amino-1-metil-5H- pirido[4,3- b]indol (Trp-P-2), aflatoxina B1 y benzo[a]pireno (B[a]P). Recientes estudios in vitro e in vivo han mostrado que el complejo molecular es formado entre IQ y el CHL, sugiriendo que la formación de este complejo puede ser la responsable de los efectos antigenotóxicos del CHL. La
enzima Citocromo P450 es uno de los principales catalizadores de la bioactivación de estos carcinógenos (Yun et.al, 1995). Se ha sugerido que la actividad antimutagénica del CHL proviene de su fuerte asociación con los mutágenos o sus intermediarios, de la recolección de los radicales libres y de la supresión o la interferencia con la activación metabólica por citocromos específicos (P-450) y otras enzimas metabólicas (Newmark, 1984; Arimoto et.al, 1980; Dashwood et.al, 1991; Romert et.al, 1992).
El CHL también inhibe la carcinogenicidad de 2-amino-1-metil-6-fenilimidazol [4,5-b] piridina (PhIP). El CHL reduce la carcinogénesis mamaria PhIP en ratas lo cual sugiere que el CHL es un quimiopreventor efectivo cuando es ingerido simultáneamente con carcinógenos (Hasegawa et.al, 1995).
Trabajos recientes también muestran que el CHL altera el espectro de mutaciones de β-catenina en tumores de colon inducidos en rata (Blum et.al, 2001) y que genera cambios en las tasas de proliferación celular y apoptosis en la cripta del colon (Dashwood et.al, 2001).
El CHL induce apoptosis en células HCT116 sin la liberación de Citocromo c de la mitocondria ni la activación de la vía Apaf-1/Caspasa-9. Como un mecanismo alternativo, el CHL activa la vía que envuelve a la caspasa-6 y la liberación de AIF de la mitocondria, resultando en el clivaje de las laminas nucleares (Díaz et.al, 2003). El CHL también induce cambios en la expresión de proteínas de la familia Bcl-2 incluyendo a tBid y Bak conduciendo a la apoptosis (Díaz et.al, 2003). El CHL
también actúa como un antioxidante para inhibir la peroxidación de los lípidos (Kammat et.al, 2000).
El CHL ha sido usado en el tratamiento de varias condiciones humanas sin la evidencia de toxicidad (Harrison et.al, 1954; Young and Beregi, 1980; Ong et.al, 1986) y recientemente ha sido usado como un inhibidor potente de la hepatocarcinogénesis causada por aflatoxina B1 en trucha (Breinholt et.al, 1995). Comercialmente, el clorofilin es ampliamente usado como desodorante de la materia fecal y orina en pacientes geriátricos, para la cicatrización de heridas y como colorante para comidas (PDR 2001). Ademas, se ha demostrado que la administración oral a corto tiempo de CHL disminuye la genotoxicidad inducida por aminas heterocíclicas mediante la inducción de enzimas de fase 2 (Dingley et.al, 2003). Actualmente, las guías de la FDA (Administración de Alimentos y Drogas) permiten la ingestión de 3 tabletas de 100 mg por día (Ikeda 2001).
3. Formulación del problema y Justificación
3.1 Formulación del problema
El cáncer es una enfermedad de origen multifactorial que se caracteriza por la proliferación de células que presentan alteraciones en su DNA y que escapan a los mecanismos de control del ciclo celular y muerte celular programada permitiendo una división rápida e incontrolada. El cáncer ocasiona cerca del 20% de todas las muertes
a nivel mundial y constituye la segunda causa de muerte en los países desarrollados. Por ejemplo, en Estados Unidos, más de un millón de personas son diagnosticadas cada año para algún tipo de cáncer. En Colombia, las tasas de incidencia y de mortalidad de cáncer se han venido incrementando en las últimas décadas; cerca de 28 mil hombres y mujeres mueren anualmente por esta causa.
Existen diferentes tipos de tumores los cuales se clasifican por su tejido de origen, por ejemplo, el sarcoma es un tumor maligno que se origina en los tejidos conectivos. Entre los sarcomas, el fibrosarcoma, es un tumor maligno de origen mesenquimal que se caracteriza histológicamente por presentar una población predominante de células epiteliales y se localiza con mayor frecuencia en las extremidades inferiores. En Colombia, el fibrosarcoma tiene una incidencia promedio de 2 casos por cada 100.000 habitantes, correspondiendo al 1% de las neoplasias en adultos y al 15% en niños. Según la American Cancer Society, en Estados Unidos se diagnostican 5000 casos nuevos al año y se estiman en 2000 las muertes anuales.
Si bien, la incidencia del fibrosarcoma es baja, en general, es un tumor con un índice de agresividad alto, por lo cual, su pronostico en la actualidad es muy malo debido a la ausencia de un tratamiento eficiente. Dentro de de las formas de tratamiento para el fibrosarcoma, el quirúrgico continúa siendo el método más efectivo, pero sus resultados no son del todo satisfactorios debido a los índices de recidiva y a su capacidad metastatizante. La radioterapia y la quimioterapia son alternativas válidas como coadyuvantes de la cirugía, pero tienen un alto costo y
efectos secundarios adversos reflejando la necesidad de implementar nuevas estrategias terapéuticas para el tratamiento de la enfermedad.
3.2 Pregunta de Investigación
¿Es el clorofilin un regulador de la proliferación celular en células de fibrosarcoma humano HT1080?
3.3 Justificación de la Investigación
Ya que el objetivo principal de la mayoría de las terapias contra el cáncer es reducir el número de células tumorales y/o inducir su diferenciación, se plantea la necesidad de realizar investigaciones con moléculas que tengan efectos sobre el control de la proliferación y diferenciación celular. El clorofilin, un derivado hidrosoluble de la clorofila, se perfila como una molécula con efectos importantes sobre el control de la proliferación y diferenciación celular, tal como ha sido reportado, no solo por sus propiedades antimutagénicas, sino también por la modulación que ejerce sobre la apoptosis y diferenciación celular. Específicamente, se ha observado que el clorofilin regula mecanismos de proliferación y diferenciación en células de carcinoma de colon, mostrando una relación entre los efectos de esta molécula con los mecanismos de control de muerte celular programada, ciclo y diferenciación celular. A la fecha, la literatura no reporta estudios sobre los efectos del clorofilin en células de fibrosarcoma. Con este trabajo se pretende conocer el efecto del clorofilin sobre el ciclo y muerte celular en células de fibrosarcoma.
4. Hipótesis
Ho: El clorofilin regula la proliferación celular en células de fibrosarcoma humano
HT1080.
Ha: El clorofilin no regula la proliferación celular en células de fibrosarcoma humano
HT1080. 5. Objetivos
5.1 Objetivo General
Evaluar el efecto del clorofilin sobre la proliferación celular en células de fibrosarcoma humano HT1080.
5.2 Objetivos Específicos
1. Determinar el efecto del clorofilin a concentraciones de 0.25 mM, 0.50 mM y
0.75 mM sobre la proliferación y muerte de células de fibrosarcoma humano HT1080.
2. Determinar la viabilidad de células de fibrosarcoma humano HT1080 en respuesta al clorofilin in vitro.
3. Determinar el efecto del clorofilin sobre el contenido de DNA en células
HT1080.
6. Materiales y Métodos
6.1 Diseño de la Investigación
Diseño experimental in vitro
6.1.1 Población de Estudio y Muestra
La población correspondió a la línea celular de fibrosarcoma humano HT1080 obtenida de la American Type Culture Collection (ATCC) (Rockville, MD).
Se hicieron dos pruebas por triplicado para la determinación de la viabilidad celular por MTS. Así mismo, se hicieron dos pruebas por triplicado para el conteo de células adheridas y células flotantes, para el análisis de morfología celular por microscopia de luz y para el análisis de ciclo celular por citometría de luz.
6.1.2 Variables del Estudio
En este estudio la variable independiente (X) fueron las distintas concentraciones de CHL (0.25, 0.50 y 0.75 mM) y el tiempo de exposición a éste. La variable dependiente (Y) fue el efecto del CHL sobre la proliferación y ciclo celular de las células de fibrosarcoma humano HT1080.
6.2 Metodología
6.2.1 Cultivo Celular
Las células de fibrosarcoma humano HT1080 fueron sembradas en cajas de cultivo T25 cm2 a una densidad de 1 x 105 en 5 ml de Medio Esencial Mínimo (MEM) suplementado con 10% de Suero Fetal Bovino, 2.2 g/l de bicarbonato de sodio, 100 u/ml de penicilina y 100 µg/ml de estreptomicina al que se denominó medio completo.
Los cultivos se mantuvieron a 37ºC en atmósfera húmeda al 5% de CO2. Se hicieron pases sucesivos por desprendimiento de las células con Tripsina 0.25%-EDTA [1mM] durante cinco minutos a 37ºC y posteriormente fueron lavadas con MEM suplementado con SFB al 1% (medio de lavado) y centrifugadas a 2000 rpm durante cinco minutos a 4ºC con el objetivo de separar el medio de lavado y la tripsina de las células. Posteriormente se descartó el sobrenadante teniendo precaución de no disgregar el pellet. Este, fue resuspendido en 5 ml de medio completo y la suspensión celular fue sembrada en nuevas cajas de cultivo T25 cm2 con el fin de expandir el cultivo celular. Se efectuaron cambios de medio cada dos o tres días cuando se observaba el medio metabolizado por cambio de pH. El seguimiento del cultivo se llevó a cabo en el microscopio invertido hasta que se observó proliferación celular. Todas las pruebas fueron realizadas en cultivos que se encontraban entre el 5º y el 12º pase.
6.2.2 Tratamiento de las Células con Clorofilin
Se preparó una solución stock de clorofilin de 20 mM en agua destilada estéril, la cual fue protegida de la luz y diluciones en serie de 0.25mM, 0.50mM, y 0.75mM fueron preparadas justo antes de su adición a las células. Las células fueron sembradas en placas de 6, 12 y 96 pozos a una densidad de 400 000 células en tres ml de medio completo, 160.000 células en dos ml de medio completo y 30 000 células en 200 µl de medio completo respectivamente, y después de 24 horas de incubación a 37ºC en atmósfera húmeda al 5% de CO2 para lograr adherencia celular, se inició el tratamiento con CHL durante 24, 48 y 72 horas. Se tuvo un grupo control células de fibrosarcoma humano HT1080 únicamente con medio completo.
6.2.3 Determinación de la Viabilidad Celular por MTS
Se tomaron células de fibrosarcoma humano HT1080 de las cajas de cultivo T25 cm2 y fueron sembradas en placas de 96 pozos a una densidad de 30 000 células en 200 µl de medio completo y después de ser incubadas a 37ºC en atmósfera húmeda al 5% de CO2 con concentraciones de 0.25 mM, 0.50 mM y 0.75 mM de CHL y el respectivo grupo control, durante 24, 48 y 72 horas respectivamente, se aspiró el medio, se agregaron 50 µl de medio completo y 10 µl de CellTiter 96® AQueous One Solution Assay (Promega) y se incubaron a 37ºC durante dos horas. Posteriormente se hizo la lectura de absorbancia a 480 nm en un lector de ELISA HUMAREADER para determinar la viabilidad celular frente a las diferentes concentraciones de clorofilin. El CellTiter 96® AQueous One Solution Assay es un método
colorimétrico para determinar la viabilidad celular en ensayos de proliferación o citotoxicidad. Este ensayo contiene un novedoso componente de tetrazolio llamado MTS [3- (4,5-dimetiltiazol-2-il) -5- (3-carboximetoxifenil) -2- (4-sulfofenil) -2H- tetrazolio] y un reactivo llamado fenasina etosulfato (PES) el cual aumenta la
estabilidad quimica. El MTS es reducido por las células metabólicamente activas en
un producto de formazan que es soluble en el medio de cultivo (Barltrop et.al, 1991)
6.2.4 Células Adheridas y Células Flotantes
Se tomaron células de fibrosarcoma humano HT1080 de las cajas de cultivo T25 cm2 y fueron sembradas en placas de doce pozos a una densidad de 160 000 células en 2 ml de medio completo y después de ser incubadas a 37ºC en atmósfera húmeda al 5% de CO2 con concentraciones de 0.25 mM, 0.50 mM y 0.75 mM de CHL y el respectivo grupo control, durante 24, 48 y 72 horas respectivamente, se tomó el medio contenido en cada pozo y se llevó a un tubo falcon previamente marcado de donde se tomaron 10 µl para ser montados en cámara de NeuBauer y hacer el conteo de células flotantes.
Posteriormente, cada pozo fue lavado con 1 ml de PBS 1X pH 7.4 durante un minuto y después se le agregaron 200 µl de Tripsina 0.25%-EDTA [1mM] y se incubó durante cinco minutos a 37ºC en atmósfera húmeda al 5% de CO2 para desprender las células. El monitoreo del desprendimiento celular se hizo en microscopio de luz invertida. Una vez desprendidas las células, se inactivó la tripsina añadiendo 1 ml de medio de lavado y se llevó la suspensión al tubo falcon
previamente marcado correspondiente a cada tratamiento para ser centrifugado a 2000 rpm durante cinco minutos a 4ºC. El pellet fue resuspendido en un volumen final de 2 ml de medio completo de donde se tomaron 10 µl de la suspensión celular para ser montados en cámara de NeuBauer y hacer el conteo de células adheridas.
6.2.5 Análisis de Morfología Celular por Microscopía de Luz
Se tomaron células de fibrosarcoma humano HT1080 de las cajas de cultivo T25 cm2 y fueron sembradas en placas de doce pozos a una densidad de 160.000 células en 2 ml de medio completo. Después de ser incubadas a 37ºC en atmósfera húmeda al 5% de CO2 con concentraciones de 0.25 mM, 0.50 mM y 0.75 mM de CHL y el respectivo grupo control, durante 24, 48 y 72 horas respectivamente, se observaron bajo microscopio de luz para determinar si tenían características morfológicas típicas de muerte celular programada (Alfaro et.al, 2000; Barrio et.al, 2003).
6.2.6 Análisis de Ciclo Celular por Citometría de Flujo
Para el análisis de ciclo celular, se tomaron células de fibrosarcoma humano HT1080 de las cajas de cultivo T25 cm2 y se sembraron en placas de doce pozos a una densidad de 200 000 células en 2 ml de medio completo y después de ser incubadas a 37ºC en atmósfera húmeda al 5% de CO2 con concentraciones de 0.25 mM, 0.50 mM y 0.75 mM de CHL y el respectivo grupo control, durante 24, 48 y 72 horas respectivamente, se descartó el medio y los pozos fueron lavados con 1 ml de
PBS 1X pH 7.4 durante 1 minuto y desprendidos con 200 µl de Tripsina 0.25%-EDTA [1mM] durante cinco minutos a 37ºC en atmósfera húmeda al 5% de CO2. El monitoreo del desprendimiento celular se hizo en microscopio de luz invertida. Una vez desprendidas las células, se agregó 1 ml de medio de lavado por pozo con el objetivo de inactivar la tripsina y se llevó la suspensión celular a los tubos de citometría de flujo previamente marcados y se centrifugaron a 2000 rpm durante cinco minutos a 4ºC con el objetivo de separar el medio de lavado y la tripsina de las células. Posteriormente se descartó el sobrenadante teniendo precaución de no disgregar el pellet. A cada tubo se agregaron 300 µl de reactivo de Vindelov (100 ml de buffer salino Tris pH 7.6, 1 mg (350 unidades) de Ribonucleasa A, la cual degrada el RNA para permitir únicamente la lectura del DNA, 7.5 mg de Ioduro de Propidio que se une tanto a DNA como a RNA, 0.1 ml de Igepal-C 240 el cual es un detergente que rompe el citoplasma permitiendo la exposición del núcleo) (Vindelov 1997) y los tubos fueron incubados durante una hora a 4ºC en oscuridad. Posteriormente, las células fueron llevadas a estudio de citometría por medio de un citómetro de flujo FACScalibur donde se adquirieron 10 000 eventos y se obtuvieron los porcentajes relativos del número de células que se encontraban en cada fase del ciclo celular (G0/G1, S, G2/M) a través del análisis con el software ModFit.
6.3 Recolección de la Información
Los datos del análisis de la viabilidad celular por MTS se obtuvieron mediante la lectura de absorbancia a 480 nm en un lector de ELISA HUMAREADER. Así
mismo, los datos del ciclo celular fueron los porcentajes relativos del número de células en cada fase del ciclo celular (G0/G1, S, G2/M) los cuales se obtuvieron del análisis por citometría de flujo a través del software ModFit.
6.4 Análisis de la Información
El análisis de la información se hizo mediante la prueba estadística t de student.
7. Resultados
7.1 Análisis de la Viabilidad Celular por MTS
El ensayo de viabilidad celular MTS mostró el efecto del CHL a diferentes concentraciones durante 24, 48 y 72 horas de tratamiento sobre células de fibrosarcoma humano HT1080. Para el análisis estadístico, se realizó una prueba t de student en la que un valor de p<0.05 fue considerado estadísticamente significativo.
A las 24 horas de tratamiento las células del grupo control mostraron un valor promedio de absorbancia de 0.968 ± 0.066; en las células expuestas a una concentración de 0.25 mM de CHL, este valor disminuyó a 0.815 ± 0.036 siendo el valor de p de 0.013 es decir estadísticamente significativo; para la concentración de 0.50 mM el valor de absorbancia fue de 0.899 ± 0.043 con un valor de p de 0.1821, y para 0.75 mM de 0.829 ± 0.051 con un valor de p de 0.0284. A las 48 horas de tratamiento con CHL se encontró para las células control un valor promedio de
absorbancia de 1.325 ± 0.034; el cual disminuyó a 1.282 ± 0.057 con un valor de p de 0.3017 en las células expuestas a una concentración de 0.25 mM de CHL, a 1.127 ± 0.037 con un valor de p de 0.0005 en las células expuestas a una concentración de 0.50 mM de y a 0.821 ± 0.090 con un valor de p de 0.0001 en las células expuestas a 0.75 mM de CHL. A las 72 horas de tratamiento se observó para el grupo control un valor promedio de absorbancia de 1.400 ± 0.057, que disminuyó a 1.323 ± 0.057 con un valor de p de 0.1467 en las células expuestas a 0.25 mM de CHL, a 0.746 ± 0.089 con un valor de p menor de 0.0001 para la concentración de 0.50 mM de CHL y a 0.437 ± 0.061 con un valor de p de 0.0001 para la concentración de 0.75 mM de CHL (ver Tabla 2, Figura 5)
Tabla 2. Valores promedio de absorbancia de células de fibrosarcoma humano HT1080 expuestas a diferentes concentraciones de CHL durante 24, 48 y 72 horas. Se muestran los valores promedio de absorbancia y ± desviación éstandar de dos experimentos por
cuadriplicado, del grupo control y de las concentraciones de 0.25 mM, 0.50 mM y 0.75 mM
de CHL. Un valor de p<0.05 fue considerado estadísticamente significativo *.
Tratamiento
24 horas 48 horas 72 horas
Promedio ± D.E Promedio ± D.E Promedio ± D.E
Control 0,968 0,066 1,325 0,034 1,400 0,057
0.25 mM 0,815* 0,036 1,282 0,057 1,323 0,057
0,5 mM 0,899 0,043 1,127* 0,037 0,746* 0,089
0,75 mM 0,829* 0,051 0,821* 0,090 0,437* 0,061
Viabilidad Celular en Células HT1080 tratadas con CHL 0,4 0,5 0,6 0,7 0,8 0,9 1,0 1,1 1,2 1,3 1,4 1,5
24 horas 48 horas 72 horas
Figura 5. Viabilidad Celular en células de fibrosarcoma humano HT1080 expuestas a CHL durante 24, 48 y 72 horas. Se muestran los valores promedio de absorbancia y las ± desviaciones estándar de dos experimentos por cuadriplicado para cada una de las
condiciones experimentales
Como se observa en la figura cinco y según los resultados de la prueba t de student, la viabilidad celular de las células expuestas a las concentraciones de 0.25 mM y 0.75 mM de CHL durante 24 horas de tratamiento, disminuyó cerca del 15% siendo estadísticamente significativa con respecto a las células del grupo control mientras que la disminución de la viabilidad celular en las células expuestas a 0.50 mM fue del 8%, no estadísticamente significativa. Según esto, las concentraciones de 0.25 y 0.75 mM de CHL sobre las células de fibrosarcoma humano HT1080 durante 24 horas afectan la proliferación celular. A las 48 horas de tratamiento con CHL, la viabilidad celular de las células de fibrosarcoma humano HT1080 disminuyó de manera dosis-dependiente con respecto al control siendo para la concentración de 0.25 mM del 5% aproximadamente, no estadísticamente significativa mientras que para 0.50 mM la disminución en la viabilidad celular fue del 15% aproximadamente y para 0.75 mM del 38% aproximadamente, estadísticamente significativa para ambas concentraciones. A las 72 horas de tratamiento con CHL es donde se observa la mayor disminución de la viabilidad celular en las concentraciones más altas de CHL utilizadas siendo para 0.50 mM del 47% aproximadamente y para 0.75 mM del 70% aproximadamente. De acuerdo con estos resultados el CHL evidencia un efecto fuerte sobre la proliferación y viabilidad de células de fibrosarcoma humano HT1080.
7.2 Células Adheridas y Células Flotantes
La actividad del CHL sobre células de fibrosarcoma humano HT1080 se evaluó mediante el conteo de células adheridas y células flotantes en el que se observó un incremento estadísticamente significativo en el porcentaje de células flotantes a medida que aumentó la concentración de CHL y el tiempo de exposición a éste al igual que una disminución estadísticamente significativa en el porcentaje de células adheridas. A las 24 horas de tratamiento en las células del grupo control, el porcentaje de células flotantes fue de 0.77% ± 0.19; en las células expuestas a una concentración de 0.25 mM de CHL, este valor aumentó a 11.50% ± 1.17; para la concentración de 0.50 mM continuó aumentando a 32.32% ± 1.44 y para 0.75 mM aumentó a 69.21% ± 1.10.
A las 48 horas de tratamiento con CHL, el porcentaje de células flotantes para el grupo control fue de 4.57% ± 1.15; el cual aumentó en las células expuestas a una concentración de 0.25 mM de CHL a 25.65% ± 0.66; para la concentración de 0.50 mM continuó aumentando a 65.77% ± 3.28 y para 0.75 mM aumentó a 87.96% ± 0.36. A las 72 horas de tratamiento, se observó también un aumento en el porcentaje de células flotantes y una disminución en el porcentaje de células adheridas a medida que aumentaba la concentración de CHL, aunque de una manera más evidente pues para el grupo control el porcentaje de células flotantes fue de 6.84% ± 0.97 mientras que para las células expuestas a una concentración de 0.25 mM de CHL fue de 40.47% ± 0.56; para la concentración de 0.50 mM fue de 86.56% ± 0.83 y para 0.75 mM fue de 94.39% ± 0.53 (ver Tabla 3, Figura 6)
Tabla 3. Porcentaje de células adheridas y células flotantes a diferentes concentraciones de CHL durante 24, 48 y 72 horas. Se muestran los porcentajes promedio de células adheridas y células flotantes de dos experimentos por triplicado para cada una de las
condiciones experimentales. Un valor de p<0.05 fue considerado estadísticamente
significativo *.
Tratamiento
% Células Adheridas % Células Flotantes 24 Horas 48 Horas 72 Horas 24 Horas 48 Horas 72 Horas
Control 99,23 95,44 93,16 0,77 4,57 6,84
0,25 mM 88,51* 74,35* 59,53* 11,50* 25,65* 40,47*
0,50 mM 67,69* 34,24* 13,44* 32,32* 65,77* 86,56*
0,75 mM 30,79* 12,05* 5,62* 69,21* 87,96* 94,39*
A.
Efe cto de l CHL sobre cé lulas de Fibrosarcoma Humano HT1080 0 10 20 30 40 50 60 70 80 90 100
24 Horas 48 Horas 72 Horas
B.
Efe cto de l CHL sobre cé lulas de Fibrosarcoma Humano HT1080 0 10 20 30 40 50 60 70 80 90 100
24 Horas 48 Horas 72 Horas
Tie mpo % C é lu la s A d h e r id a s control 0,25 mM 0,50 mM 0,75 mM * * * * * * * * *
Figura 6. Porcentaje de Células Flotantes (A) y Adheridas (B) de células de fibrosarcoma humano HT1080 expuestas a CHL durante 24, 48 y 72 horas. Se muestran los promedios y las desviaciones estándar de cada una de las condiciones experimentales
hasta 72 horas de tratamiento. Un valor de p<0.05 fue considerado estadísticamente
significativo *.
[image:60.612.150.493.96.363.2]Como se observa en la Figura 6A, el porcentaje de células flotantes aumenta con respecto al del grupo control de manera dosis y tiempo-dependiente. Así mismo, en la Figura 6B lo que se observa es la disminución del porcentaje de células adheridas con respecto al grupo control de manera dosis y tiempo-dependiente.
7.3 Análisis de Morfología Celular por Microscopía de Luz
Para analizar los cambios en la morfología celular después del tratamiento con las diferentes concentraciones de CHL, se utilizó la microscopia de luz (Alfaro et.al, 2000; Barrio et.al, 2003). Las células del grupo control presentaron una morfología ovoide con núcleo pequeño, citoplasma claro rodeado de fibras de colágeno hialinas y proyecciones hacia los lados. En estado de confluencia estas células crecieron en múltiples capas y formaron microtumores a través de las placas de cultivo (Rasheed et.al, 1974). En las células expuestas a las concentraciones de 0.25 mM, 0.50 mM y 0.75 mM se pudieron apreciar cambios en la organización celular, característicos de la apoptosis tales como la condensación citoplasmática y de la cromatina, desarrollo de vacuolas citoplasmáticas y aparición de fragmentos celulares o cuerpos apoptóticos, sugiriendo que el CHL desencadenó la muerte celular programada en células de fibrosarcoma humano HT1080.
7.4 Análisis de Ciclo Celular por Citometría de Flujo
Para el análisis de ciclo celular en células de fibrosarcoma humano HT1080 in vitro, se realizó una marcación de las mismas con el reactivo de Vindelov y posteriormente se llevaron a estudio de citometría de flujo donde se adquirieron 10.000 eventos. Para el análisis de esta información, se cuantificó el DNA, mediante la obtención de histogramas que mostraron una distribución porcentual de cada una de las fases del ciclo celular (G0/G1, S, G2/M).
En las células del grupo control a las 24 horas se observó un porcentaje de células en G0/G1 de 38.33% ± 1.70; que a las 48 horas aumentó a 41.50% ± 0.71 y a las 72 horas a 59.33% ± 0.47. De igual manera, el porcentaje de células en fase S disminuyó en el tiempo para los diferentes intervalos de tiempo siendo menor a las 72 horas de tratamiento (24h: 57.67% ± 1.70; 48h: 43.63% ± 0.26; 72h: 33% ± 0). El valor de porcentaje de células en fase G2/M incrementó entre las 24 y las 48 horas de tratamiento siendo de 4.00% ± 0.82 y de 14.87% ± 0.90 respectivamente, mientras que a las 72 horas de tratamiento el porcentaje de células en esta fase disminuyó a 7.67 % ± 0.47.
En las células expuestas a una concentración de 0.25 mM de CHL, se observó a las 24 horas un porcentaje de células en G0 /G1 de 42.40% ± 1.23 que aumentó tanto a las 48 como a las 72 horas de tratamiento a 54.80% ± 1.07 y a 67.40% ± 3.11 respectivamente. El porcentaje de celulas en fase S a las 24 horas de tratamiento fue de 49.00% ± 2.45; que disminuyó a 30.93% ± 0.90 a las 48 horas y a 18.33% ± 0.68 a las 72 horas. Para G2 /M, el porcentaje de células a las 24 horas fue
de 8.60% ± 1.98, que aumentó a 13.93% ± 1.55 a las 48 horas y a 14.27% ± 2.68 a las 72 horas.
En el análisis de las células expuestas a una concentración de 0.50 mM de CHL se encontró un porcentaje de 46.67% ± 1.25 de células en G0/G1 a las 24 horas el cual aumentó a 59.40% ± 5.94 a las 48 horas y a 60.53% ± 1.05 a las 72 horas. Para la fase S se observó un valor de 44.50% ± 1.47 a las 24 horas, que disminuyó a 25.33% ± 4.43 a las 48 horas y a 13.93% ± 3.65 a las 72 horas de tratamiento. El valor de porcentaje de células en fase G2/M a las 24 horas fue de 8.83 % ± 2.32, el cual aumentó a 15.27% ± 1.52 a las 48 horas y a 25.53% ± 3.45 a las 72 horas.
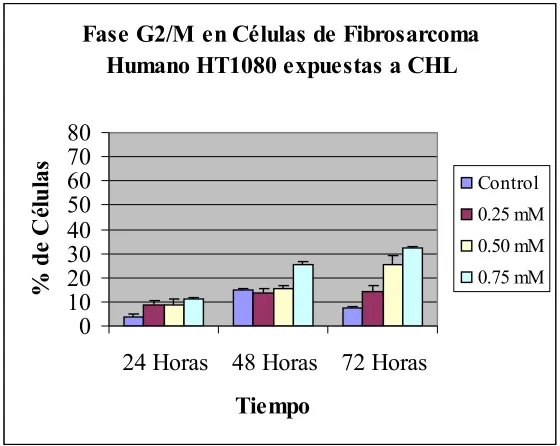
En la concentración de 0.75 mM de CHL se observó un valor de 52.67% ± 2.36 de células en G0/G1 a las 24 horas, que aumentó a 68.04% ± 3.47 a las 48 horas y disminuyó a 38.87% ± 5.08 a las 72 horas. En el porcentaje de células en fase S se encontró un valor de 36.13% ± 2.99 a las 24 horas, que disminuyó a 11.87% ± 1.98 a las 48 horas y aumentó a 29.07% ± 4.08 a las 72 horas. Para la fase G2 /M se observó un valor de 11.20% ± 0.86 a las 24 horas, el cual aumentó a 25.54% ± 1.04 y a 32.07% ± 1.05 a las 48 horas y a las 72 horas respectivamente (ver Tabla 4, Figura 7).
Tabla 4. Resultados del análisis de ciclo celular por citometría de flujo. Se muestran los porcentajes de células HT1080 del grupo control y de las células expuestas a las diferentes
concentraciones de CHL así como los valores de desviación estándar hasta 72 horas de
tratamiento. Un valor de p<0.05 fue considerado estadísticamente significativo *.
Control Fase G0-G1 Fase S Fase G2-M
Tiempo Promedio D.E Promedio D.E Promedio D.E
24 Horas 38,33 1,7 57,67 1,7 4 0,82
48 Horas 41,5 0,71 43,63 0,26 14,87 0,9
72 Horas 59,33 0,47 33 0 7,67 0,47
0.25 mM Fase G0-G1 Fase S Fase G2-M
Tiempo Promedio D.E Promedio D.E Promedio D.E
24 Horas 42,40 1,23 49,00* 2,45 8,60* 1,98
48 Horas 54,80* 1,07 30,93* 0,90 13,93 1,55
72 Horas 67,40* 3,11 18,33* 0,68 14,27* 2,68
0.50 mM Fase G0-G1 Fase S Fase G2-M
Tiempo Promedio D.E Promedio D.E Promedio D.E
24 Horas 46,67* 1,25 44,50* 1,47 8,83* 2,32
48 Horas 59,40* 5,94 25,33* 4,43 15,27 1,52
72 Horas 60,53 1,05 13,93* 3,65 25,53* 3,45
0.75 mM Fase G0-G1 Fase S Fase G2-M
Tiempo Promedio D.E Promedio D.E Promedio D.E
24 Horas 52,67* 2,36 36,13* 2,99 11,20* 0,86
48 Horas 62,59* 1,77 11,87* 1,98 25,54* 1,04
72 Horas 38,87* 5,08 29,07* 4,08 32,07* 1,05
A.
Fase G0/G1 en Células de Fibrosarcoma Humano HT1080 expuestas a CHL
0 10 20 30 40 50 60 70 80
24 Horas 48 Horas 72 Horas Tiempo
%
d
e C
él
u
las Control
0.25 mM
0.50 mM
0.75 mM
B.
Fase S en Células de Fibrosarcoma Humano HT1080 expuestas a CHL
0 10 20 30 40 50 60 70 80
24 Horas 48 Horas 72 Horas Tiempo
%
d
e C
él
u
las Control
0.25 mM
0.50 mM
0.75 mM
C.