2296432c62400783eb40f575b3115bc5
384
0
0
Texto completo
(2)
(3) Universitat Autònoma de Barcelona. CARACTERIZACIÓN Y VALIDACIÓN MORFOLÓGICA Y MOLECULAR DE LOS TUMORES MAMARIOS EXPERIMENTALES. MODULACIÓN POR LOS LÍPIDOS DE LA DIETA Memoria presentada por IRMGARD COSTA TRACHSEL para optar al grado de Doctor en Medicina.. Este trabajo se ha realizado bajo la dirección del Prof. Eduard Escrich Escriche, en el Departamento de Biología Celular, de Fisiología y de Inmunología de la Facultad de Medicina de la Universitat Autònoma de Barcelona, estando adscrita al programa de Doctorado de Biología Celular. .. Irmgard Costa Trachsel. Eduard Escrich Escriche. Barcelona, mayo 2014.
(4)
(5) Este trabajo ha sido cofinanciado, en parte, mediante ayudas del Plan Nacional de I+D+I del Ministerio de Educación y Ciencia/Economía y Competitividad (AGL2006-0769/AGL2011-24778), la Fundación Patrimonio Comunal Olivarero (FPCO2008-165.396), la Agencia para el Aceite de Oliva del Ministerio de Medio Ambiente y de Medio Rural y Marino (AAO2008-165.471), la Organización Interprofesional del Aceite de Oliva Español (OIP2009-165.646) y los Departamentos de Salud y de Agricultura, Alimentación y Acción Rural de la Generalitat de Catalunya (GC2010-165.000).. Los patrocinadores no han participado en el diseño del estudio, en la obtención de los datos, ni en el análisis o la interpretación de los resultados..
(6)
(7) Als meus pares. A l’Albert. A la Marina. "És, doncs, sols per l'amor que ens creixen roses als dits i se'ns revelen els misteris; i en l'amor tot és just i necessari" L’amor, Miquel Martí i Pol.
(8)
(9) Agradecimientos:. Una vez finalizada esta Tesis Doctoral, quisiera recordar y agradecer a todas las personas que me han acompañado durante este largo trayecto. Este trabajo ha estado presente durante muchos años de mi vida y, por ello, ha compartido muchas de mis vicisitudes. Debo decir que no ha sido una tarea fácil, y que he pasado momentos complicados, pero la ayuda de todos vosotros ha sido esencial para culminar la elaboración de este proyecto. En primer lugar, quisiera agradecer a mi Director de Tesis, el Dr. Eduard Escrich Escriche, Profesor Titular del Departamento de Biología Celular, de Fisiología y de Inmunología de la Universitat Autònoma de Barcelona i Director del Grup Multidisciplinari per a l’Estudi del Càncer de Mama, por haberme permitido colaborar en este proyecto de investigación, que es suyo, formando parte de un grupo de trabajo multidisciplinar ejemplar. A lo largo de todos estos años he podido constatar su clarividencia mental y vital, así como su espíritu luchador, exigente y positivo, que me han servido, en momentos concretos, como referencia en mi vida personal y profesional. Espero que esta relación, franca y productiva, perdure en el tiempo. Moltes gràcies, Eduard, per compartir amb mi els teus coneixements, per la teva confiança i el teu suport. Asimismo, quiero dar las gracias a mis compañeras y compañeros del “Grup Multidisciplinari per a l’Estudi del Càncer de Mama”, en especial a la Dra. Montserrat Solanas y a la Dra. Raquel Moral, por su impagable compañerismo y su ejemplo de seriedad, constancia y honradez; así como a todos los que han formado parte, en algún momento, de este proyecto: la Dra. Gemma García, la Dra. Laura Grau, la Dra. Elena Vela y el Dr. Toni Hurtado; a la Dra. Raquel Escrich, con la que he coincidido en el mismo período de tiempo en la defensa de nuestras Tesis Doctorales, por su ayuda y su soporte profesional y personal durante la fase final de este trabajo; y a los últimos en incorporarse al grupo: Cristina de Miguel, Txell Pelicano, Cristina Rodríguez, Isabel Sospedra, Xavi Franco y Miguel Ángel Manzanares. Ha sido un placer compartir con vosotros estos años y veros crecer. También, quiero agradecer a la Dra. María del Carmen Ruiz de Villa, del Departamento de Bioestadística de la Universitat de Barcelona y miembro del grupo, por su importante colaboración, disponibilidad y dedicación en el tratamiento estadístico de los datos obtenidos. Ahora ya llevo unos años en el Servicio de Patología del Hospital Parc Taulí, que se ha convertido en mi proyecto profesional. Mi más sincero agradecimiento a la Dra. Mercè Rey.
(10) por permitirme desarrollar y finalizar sin problemas este trabajo, facilitándome la conciliación con mis deberes profesionales, en todos los momentos en que lo he requerido, así como por los ánimos y la comprensión mostrada, especialmente, durante el trayecto final; a todos y a todas mis compañeros y compañeras del Servicio: a Carmen, por compartir despacho y mucho más (¡es un regalo tenerte tan cerca y haber podido disfrutar contigo este momento tan especial de tu vida!); a Neus, por escucharme y ayudarme cuando más lo he necesitado; a Javier, por su amistad y por transmitirme sus conocimientos en Patología Mamaria; a Tona, Empar, Mª Ángeles, Rut, Mariona, Mª Rosa y Àlex por permitirme aprender de ellos y por ser ejemplos de dedicación en lo que les apasiona; todos vosotros sois un innegable pilar del Servicio; a Olga, Mari Carmen, Jose, Dani, Tere, Rosa, Maribel, Luís, Cati, Anna Ferran, Anna Galcerán, Henar, Joana, Eva, Carla, Emi, María, Susana, Josep y Rubén, por vuestra capacidad en solucionar mis incidencias laborales y por los momentos deliciosos que me regaláis; y a todos los residentes con los que he coincidido, en especial a los últimos: Esther, Anna, Marta, Onica, Lina, Rodolfo y Rosa, por ser un aliciente indispensable para seguir progresando. Gracias a todos y a todas por vuestro compañerismo, vuestra profesionalidad y seriedad en el trabajo y en la vida. Quisiera expresaros en estas líneas mi más sincera amistad. Esta Tesis Doctoral empezó estando yo en el Hospital de Granollers. Allí tengo grandes amigos e inmejorables compañeros. Muchas gracias a todas y a todos: a la Dra. Mariana Bernet por ayudarme a iniciar este proyecto y creer en mí; y a todos los miembros del Servicio de Anatomía Patológica que compartieron conmigo aquellos años. No éramos muchos, así que puedo citaros a todos: Jordi, Ángel, Remei, Eulàlia, Josa, MªÀngels, Patricia, Asun, Núria, Clara, Eva, Sílvia y Toni. ¡Aún conservo la carpeta que me regalasteis para empezar a archivar artículos! Gracias a mis amigas y amigos, a los de siempre y a los más recientes. A mis amigas de la Facultad: Ana, Aurora, Elo, Lucila, Mar, María José, Mireia, Miriam, Iciar y Quima, a las que no puedo ver mucho pero que han marcado mi vida. A Caroline Le Bousse-Kerdiles por haber sido mi primera maestra del buen hacer profesional y por continuar siendo mi amiga. A Ana Chivite por soportarme durante los años de residencia y por su inquebrantable y sincera estima. A Vicky, Maite y Elena por sus locuras, por hacerme reír y por llorar conmigo. A Salva por estar siempre allí. A Paqui por su ayuda desinteresada en momentos difíciles; gracias por preocuparte tanto por mí. A Carlos y a María por formar parte de tantas épocas de mi vida, desde hace ya muchos años. A Francesca y a Jaume por saber que siempre puedo contar con vosotros. A Mireia Gimeno por su amistad y por las horas compartidas organizando los horarios escolares de nuestros hijos. A mis amigas y amigos del Club Esquí.
(11) Terrassa, que cada vez sois más, por ayudarme a desconectar de los problemas diarios y permitirme ser yo misma, especialmente a Carme Vizcaíno por demostrarme siempre su amistad. También quiero agradecer a Sally el sincero amor que brinda a mi familia. Por su generosidad, honradez y por cuidarnos tanto y tan bien. Moltes gràcies papà i mamà. Sempre he pogut comptar amb vosaltres i sentir la vostra estimació quan més ho he necessitat. Gràcies mamà per ser el pal de paller de la nostra família. Gràcies papà per conservar sempre els teus principis ètics i morals a pesar de les circumstàncies de la vida i per haver-me transmès la teva fe. Moltes gràcies a tots dos per impulsar i ajudar-me a desenvolupar la meva vocació en la Medicina. Gràcies Mònica per ser una germana excepcional i sempre disposada a donar-me un cop de mà; i Josep, Marc i Ferran que, tot i ser els meus germans petits, us preocupeu per mi. Vosaltres sou les meves arrels, d’on vinc. Junts hem viscut moltes coses i estic segura de que en viurem moltes més, d’alegres i felices. Hem plantat un arbre ben ferm que ha resistit moltes tempestes i molts vendavals. Gràcies Eduard i Montse per estimar tan a la Mònica i al Marc; i Marc, Helena, Marta i Anna per ser els millors cosins que la Marina podria tenir. Vull donar les gràcies al Quimo per la seva generositat i bondat. Has estat al nostre costat quan més ho hem necessitat. Gràcies, també, Xavier, Magda i Marta per saber que sempre esteu a mà; i als meus nebots i neboda més grans: Àlvar, Ignasi, Martí, Òscar i Susanna per ser els joves que heu arribat a ser, un extraordinari exemple per a la Marina. Finalment, vull agrair a l’Albert, el meu espòs, el seu amor, el seu suport i la seva incondicional confiança en mi. Fa ja molts anys que estem junts i hem viscut grans experiències plegats. La vida ens ha posat proves que hem sabut superar i estic segura que arribarem junts a la meta. I a la persona més important de la meva vida, el meu vertader tresor, la Marina, que va arribar del fred i m’ha donat tanta escalfor. Tu ets el meu motor i la meva gasolina. Amb tu he conegut i he sentit l’amor incondicional. Gràcies per entendre les meves absències i la meva poca paciència, per ser tan madura i responsable i per pensar que sóc la millor mama del món. Aquesta Tesi està especialment dedicada a tu. A la memòria de l’Enric i de la Pilar. Sempre us tenim presents..
(12)
(13) ÍNDICE. Abreviaturas ....................................................................................................................................1 Índice de Figuras .............................................................................................................................4 Índice de Tablas...............................................................................................................................6. I – INTRODUCCIÓN... ................................................................................................................7 A – GLÁNDULA MAMARIA NORMAL: DESARROLLO Y MORFOLOGÍA...................9 1. HUMANA ..............................................................................................................................9 2. RATA....................................................................................................................................14 B – CÁNCER DE MAMA ..........................................................................................................19 1. ETIOLOGÍA DEL CÁNCER DE MAMA...........................................................................20 a. Factores endocrinos..........................................................................................................21 b. Factores genéticos y epigenéticos ....................................................................................25 c. Factores ambientales ........................................................................................................31 2. ESTADIOS DE LA CARCINOGÉNESIS ..........................................................................33 3. PATOGÉNESIS DEL CÁNCER DE MAMA.....................................................................35 a. HUMANO ........................................................................................................................35 b. RATA...............................................................................................................................37 C – MODELO EXPERIMENTAL ............................................................................................39 1. CARACTERÍSTICAS DEL MODELO UTILIZADO.........................................................39 2. FACTORES QUE INFLUENCIAN LA RESPUESTA TUMORIGÉNICA .......................40 a. Especie seleccionada .......................................................................................................40 b. Dosis del carcinógeno .....................................................................................................40 c. Diferenciación glandular .................................................................................................41 D – LÍPIDOS DE LA DIETA Y CÁNCER DE MAMA ..........................................................43 1. INFLUENCIA SOBRE LAS ETAPAS DE LA CARCINOGÉNESIS................................45 2. INFLUENCIA SOBRE LOS NIVELES HORMONALES..................................................46 3. MODIFICACIONES DE LA MEMBRANA CELULAR....................................................47 4. EFECTOS SOBRE LOS MECANISMOS DE TRANSDUCCIÓN DE SEÑALES ...........49 5. EFECTOS SOBRE LA EXPRESIÓN GÉNICA……………………………………..........51 6. INFLUENCIA SOBRE LA PROLIFERACIÓN CELULAR……………………… ..........53 7. EFECTOS SOBRE EL SISTEMA INMUNITARIO………………………………. ..........54.
(14) ÍNDICE. E – CARACTERIZACIÓN MORFOLÓGICA Y MOLECULAR DEL CÁNCER DE MAMA: DIFERENCIACIÓN Y PROLIFERACIÓN...................................................... 56 1. HUMANA............................................................................................................................ 56 a. Características histopatológicas del cáncer de mama y factores pronósticos morfológicos........................................................................................................................ 57 b. Caracterización molecular. Clasificación pronóstica y predictiva del cáncer de mama……………………………………………………………………... ........................ 65 2. RATA ................................................................................................................................... 67 F – c-Ha-ras Y CÁNCER DE MAMA...................................................................................... 76 II – OBJETIVOS......................................................................................................................... 83 III – RESULTADOS................................................................................................................... 89 ARTÍCULOS ESPECÍFICOS DEL ESTUDIO MORFOLÓGICO............................. 91 ARTÍCULO 1: Histopathologic characterization of mammary neoplastic lesions induced with 7, 12 dimethylbenz(α)anthracene in the rat: A comparative analysis with human breast tumors. I.Costa, M. Solanas, E. Escrich. Archives of Pathology and Laboratory Medicine, 2002, 126: 915-927................................... 93 Antecedentes, resumen de los resultados y datos adicionales ................................................. 109 ARTÍCULO 2: High fat corn oil diet promotes the development of high histologic grade rat DMBA-induced mammary adenocarcinomas, while high olive oil diet does not. I. Costa, R. Moral, M. Solanas, E. Escrich. Breast Cancer Research and Treatment, 2004, 86: 225-235 .................................................. 119 Antecedentes, resumen de los resultados y datos adicionales ................................................. 133 ARTÍCULO 3: High corn oil and extra virgin olive oil diets and experimental mammary carcinogenesis: clinicopathological and immunohistochemical p21HaRas expression study. I Costa, R. Moral, M. Solanas, FJ. Andreu, MC. Ruiz de Villa, E. Escrich. Virchows Archiv, 2011, 458: 141-151 ............................................................................... 141 Antecedentes, resumen de los resultados y datos adicionales ................................................. 155.
(15) ÍNDICE. ARTÍCULOS DE APLICACIÓN E INTEGRACIÓN DE LOS RESULTADOS DEL ESTUDIO MORFOLÓGICO...................................................................................... 165 ARTÍCULO 4: Deregulated expression of the PCPH proto-oncogen in rat mammary tumors induced with 7,12-dimethylbenz(α)anthracene. M. Solanas, E. Escrich, A. Rouzaut, I. Costa, A. Martínez, V. Notario. Molecular Carcinogenesis 2002; 33: 219-227......................................................................... 169 Antecedentes, resumen de los resultados y datos adicionales ................................................. 181 ARTÍCULO 5: Effects of a high olive oil diet on the clinical behavior and histopathological features of rat DMBA-induced mammary tumors compared with a high corn oil diet. M. Solanas, A. Hurtado, I. Costa, R. Moral, J.A. Menéndez, R. Colomer, E. Escrich. International Journal of Oncology 2002; 21: 745-753............................................................ 183 Antecedentes, resumen de los resultados y datos adicionales ................................................. 195 ARTÍCULO 6: Identification of novel differentially expressed genes by the effect of a high-fat n-6 diet in experimental breast cancer. E. Escrich, R. Moral, G. Garcia, I. Costa, J.A. Sánchez, M. Solanas. Molecular Carcinogenesis 2004; 40: 73-78............................................................................. 199 Antecedentes, resumen de los resultados y datos adicionales ................................................. 207 ARTÍCULO 7: Are the olive oil and other dietary lipids related to cancer? Experimental evidence. E. Escrich, M. Solanas, R. Moral, I. Costa, L. Grau. Clinical and Translational Oncology 2006; 8: 868-883....................................................211 Antecedentes y resumen de los resultados ............................................................................... 229 ARTÍCULO 8: Molecular mechanisms of the effects of olive oil and other dietary lipids on cancer. E. Escrich, R. Moral, L. Grau, I. Costa, M. Solanas. Molecular Nutrition and Food Research 2007; 51: 1279-1292 ........................................231 Antecedentes y resumen de los resultados ............................................................................... 247 ARTÍCULO 9: Differential expression of H19 and vitamin D3 up-regulated protein 1 as a mechanism of the modulatory effects of high virgin olive oil and high corn oil diets on experimental mammary tumors. M. Solanas, R. Moral, G. García, L. Grau, E. Vela, R. Escrich, I. Costa, E. Escrich. European Journal of Cancer Prevention 2009; 18: 153-161 ..............................249 Antecedentes y resumen de los resultados ............................................................................... 261.
(16) ÍNDICE. ARTÍCULO 10: Diets high in corn oil or extra-virgin olive oil provided from weaning advance sexual maturation and differentially modify susceptibility to mammary carcinogenesis in female rats. R. Moral, R. Escrich, M. Solanas, E. Vela, I. Costa, MC. Ruíz de Villa, E. Escrich. Nutrition and Cancer 2011; 63: 410-420 ............................................................ 263 Antecedentes, resumen de los resultados y datos adicionales ................................................. 277 IV – DISCUSIÓN ...................................................................................................................... 285 A – ESTUDIO HISTOPATOLÓGICO DE LAS LESIONES MAMARIAS EXPERIMENTALES Y DEL EFECTO DE LOS LÍPIDOS DE LA DIETA SOBRE LOS CARCINOMAS DE MAMA EN EL MODELO EXPERIMENTAL INDUCIDO CON DMBA EN LA RATA .................................. 288 1. CARACTERIZACIÓN MORFOLÓGICA DEL MODELO EXPERIMENTAL (Objetivo “a”)................................................................................ 290 2. EFECTO DE LAS DIETAS RICAS EN PUFA N-6 (ACEITE DE MAÍZ) Y EN ACEITE DE OLIVA VIRGEN EXTRA EN LAS CARACTERÍSTICAS MORFOLÓGICAS DEL CÁNCER DE MAMA EXPERIMENTAL (Objetivo “b”) ............................................................................................................... 297 B – ESTUDIO DE LA INFLUENCIA DE LAS DIETAS HIPERLIPÍDICAS SOBRE LA EXPRESIÓN DE p21Ha-Ras EN LOS ADENOCARCINOMAS DE MAMA EXPERIMENTALES (Objetivo “c”) ..................................................... 302 C – APLICACIÓN DEL ESTUDIO MORFOLÓGICO PROPUESTO PARA EL ADENOCARCINOMA DE MAMA DE LA RATA INDUCIDO CON DMBA EN DIFERENTES TRABAJOS DE INVESTIGACIÓN REALIZADOS CON DICHO MODELO EXPERIMENTAL (Objetivo “d”)........ 310 1. ESTUDIOS CLÍNICOS............................................................................................ 310 2. ESTUDIOS MOLECULARES Y BIOLÓGICOS ................................................... 314 a. Alteración de la expresión de PCPH en la glándula mamaria virgen y lactante de la rata y en los tumores inducidos con DMBA ...................................... 314 b. Identificación de genes expresados diferencialmente por efecto de los lípidos de la dieta en el cáncer de mama experimental............................................ 317 c. Efecto diferencial de las dietas hiperlipídicas ricas en aceite de oliva y en aceite de maíz sobre la expresión de H19 y VDUP-1.............................................. 318.
(17) ÍNDICE. d. Modulación de la susceptibilidad de la glándula mamaria a la transformación maligna secundaria a los cambios en la maduración sexual inducidos por los lípidos de la dieta .........................................................................319 D – INTEGRACIÓN DE LOS RESULTADOS MORFOLÓGICOS EN EL ESTUDIO DE LÍPIDOS DE LA DIETA Y CÁNCER DE MAMA (Objetivo “e”) ..................................................................................................................324 V – CONCLUSIONES ..............................................................................................................329. VI – BIBLIOGRAFÍA...............................................................................................................335.
(18) ÍNDICE.
(19) ABREVIATURAS: GMECM IARC WHO/OMS. “Grup Multidisciplinari per a l’Estudi del Càncer de Mama” “International Agency for Research in Cancer” “World Health Organisation”/Organización Mundial de la Salud. C / LFCO / LF HFCOI. GRUPOS EXPERIMENTALES grupo experimental control (dieta normolipídica) grupo experimental de dieta hiperlipídica de aceite de maíz desde el destete. M / HFCOP HCO / HFCO. grupo experimental de dieta hiperlipídica de aceite de maíz desde la inducción con DMBA (HCO desde el destete en Artículo 10). O / HFOOP grupo experimental de dieta hiperlipídica de aceite de oliva virgen extra desde la inducción HOO / HFOO con DMBA (HOO desde el destete en Artículo 10) MO. grupo experimental de dieta hiperlipídica, con dieta rica en aceite de maíz desde la inducción hasta los 86 días de edad y luego la dieta rica en aceite de oliva. Dieta m3 Dieta M20 Dieta mO20. DIETAS dieta experimental normolipídica dieta experimental hiperlipídica de aceite de maíz dieta experimental hiperlipídica de aceite de oliva virgen. PND VTM VTT. PARÁMETROS CLÍNICOS día posnatal volumen tumoral medio volumen tumoral total. CGA H&E GH. ESTUDIO MORFOLÓGICO campos de gran aumento hematoxilina y eosina grado histológico. NGS SBS. “Nottingham Grading System” “Scarff Bloom Richardson”. ABs ADN ADN-LI ADNc AKT/PKB ARN ARNm ATPasa BRCA CD CDK CK CLA COX CYP. VARIOS yemas alveolares (“alveolar buds”) ácido deoxiribonucleico índice de marcaje de ADN ácido deoxiribonucleico complementario proteína quinasa B ácido ribonucleico ácido ribonucleico mensajero adenosin trifosfatasa “Breast Cancer” “cluster of differentiation” quinasa dependiente de ciclina citoqueratina ácido linoleico conjugado cicloxigenasa citocromo P450 1.
(20) DAG DBD DHA DHEA DMBA E2 EGF EGFR EPA ER ERK FFA FGF GAP GDI GDP GDS GLA GLUT GnRH GST GTP GTPasas hCG HMGCoA hPL HER2 HNF-4 IDC IDPs IGF IL IP3 KiSS1 LCIS LOX LXRs MAPK Mek MMPs MMR mSR MUFA NADPH N-CAM NER NK NFκB NOS OA 2. diacilglicerol dominio de unión al ADN ácido docosahexaenoico dehidroepiandrosterona 7,12-dimetilbenz-()antraceno 17β-estradiol factor de crecimiento epidérmico receptor del factor de crecimiento epidérmico ácido eicosapentaenoico receptor de estrógenos (también RE) “extracellular signal-regulated kinase” ácidos grasos libres factor de crecimiento fibroblástico “GPTase activating protein” “guanine nucleotide dissociation inhibitor” guanina difosfato “guanine nucleotide dissociation stimulator” ácido gamma linolénico “glucose transporter” hormona liberadora de gonadotrofinas glutatión-S-transferasa guanina trifosfato guanina trifosfatasas hormona gonadotropina coriónica 3-hidroxi-3-metilglutaril coenzimaA hormona del lactógeno placentario “Human Epidermal Growth Factor Receptor 2” receptor nuclear hepático 4 carcinoma ductal infiltrante (“invasive ductal carcinoma”) proliferaciones intraductales factor de crecimiento parecido a Insulina interleuquina inositol-trifosfato kisspeptina hipotalámica carcinoma lobulillar “in situ” (“lobular carcinoma in situ”) lipooxigenasa receptores X hepáticos “mitogen-activated protein kinase” “MAPK extracellular signal-regulated kinase” metaloproteinasas de la matriz “miss-match repair” receptores de estrógenos asociados a membrana ácido graso monoinsaturado nicotinamida adenina dinucleótido fosfato ·neural cell adhesión molecule” reparación por escisión de nucleótidos “natural killer” factor nuclear κB No especificado (“not otherwise specified”) ácido oleico.
(21) OEA OFA OMS PA PC PCR PDGF PDK1 PGE PhIP PIF PI3K PIP2 PKA PKC PLA PLC PLD PPARs PR pRb PRL PTPasas PUFA RA Rb RE ROS RSK RT-PCR RP RXRs SD SERMs SOS SRE SREBP SRU STAT3 TDs TDLU TEBs TGF TIMPs TNF TRX VEGF VER VDUP1 VNTR. oleiletanolamida “Oncins France Strain A” Organización Mundial de la Salud (WHO “World Health Organization”) ácido fosfatídico fosfatidilcolina reacción en cadena de la polimerasa factor de crecimiento derivado de las plaquetas quinasa 1 dependiente de 3-fosfoinositol prostaglandina E 2-amino-1-metil-6-fenilimidazo[4,5-b]piridina inhibidor de la secreción de prolactina fosfatidil inositol 3-quinasa fosfatidilinositol 4,5-difosfato proteína kinasa dependiente de AMP cíclico proteína kinasa C fosfolipasa A fosfolipasa C fosfolipasa D receptores activados por proliferadores peroxisomales receptor de progesterona (también RP) proteína Rb fosforilada prolactina tirosina fosfatasas ácidos grasos poliinsaturados receptor de andrógenos retinoblastoma receptor de estrógenos especies reactivas del oxígeno quinasa S6 ribosomal reacción en cadena de la polimerasa transcriptasa inversa receptor de progesterona receptores del ácido retinoico Sprague-Dawley “Selective Estrogen Receptor Modulators” “son of sevenless” “steroid response elements” “sterol-regulatory element-binding protein” “steroid response units” transductor de señal y activador de la transcripción 3 ductos terminales (“terminal ducts”) unidad ductulo-lobulillar terminal (“terminal ductal lobular unit”) yemas terminales (“terminal ends buds”) factor de crecimiento transformante inhibidores de las metaloproteinasas de la matriz factor de necrosis tumoral (“tumor necrosis factor”) tioredoxina factor de crecimiento del endotelio vascular “base-excision repair” proteína 1 suprarregulada por la vitamina D3 número de repeticiones variables en tándem 3.
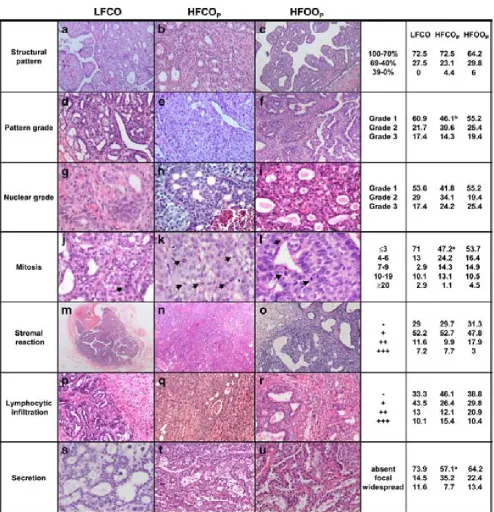
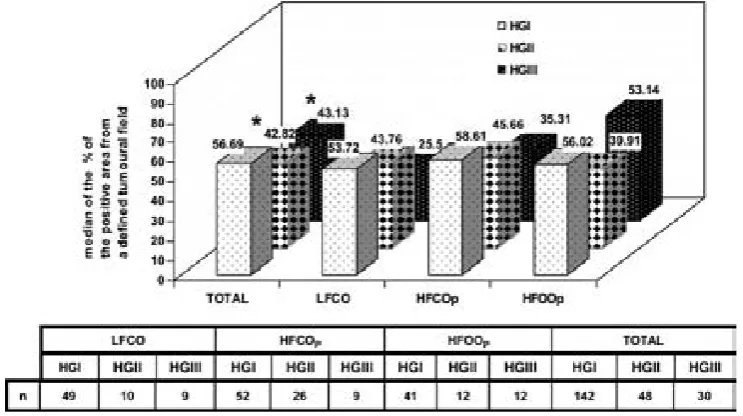
(22) ÍNDICE DE FIGURAS Figura 1: Desarrollo de la glándula mamaria humana ..................................................... 9 Figura 2: Desarrollo de la glándula mamaria de la rata virgen nulípara........................ 14 Figura 3: Estructura de una yema terminal (TEB)......................................................... 16 Figura 4: Topografía de la glándula mamaria de la rata ................................................ 17 Figura 5: Tasas de incidencia por países de cáncer de mama en mujeres .........................20 Figura 6: Carcinogénesis mamaria................................................................................. 34 Figura 7: Diferenciación de la glándula mamaria y susceptibilidad al carcinógeno ..... 43 Figura 8: Efecto de las dietas hiperlipídicas en la tumorigénesis mamaria ................... 56 Figura 9: Vías de señalización mediada por Ras ........................................................... 79 Figuras adicionales (A) a los artículos (descritas en el texto de los apartados correspondientes del capítulo de “Resultados”): Artículo 1: Figura A1: Áreas glandulares/cribiformes (H&E x40)................................................ 113 Figura A2: Grado nuclear I (H&E x200)..................................................................... 113 Figura A3: Grado arquitectural III (H&E x40)............................................................ 113 Figura A4: Pleomorfismo nuclear y nucleolos (H&E x200) ....................................... 113 Figura A5: Necrosis tipo comedo (H&E x100) .......................................................... 113 Figura A6: Infiltrado linfoplasmocitario denso (H&E x100) ...................................... 113 Figura A7: Infiltrado eosinofílico (H&E x200) ........................................................... 114 Figura A8: Patrón cribiforme predominante (H&E x40)............................................. 114 Figura A9: Patrón cribiforme y papilar mixto (H&E x40) .......................................... 114 Figura A10: Nodularidad tumoral (H&E x40)............................................................. 114 Figura A11: Nódulos tumorales expansivos (H&E x40)............................................. 114 Figura A12: Reacción estromal desmoplásica (H&E x200)........................................ 114 Figura A13: Patrón ductal y reacción estromal desmoplásica (H&E x200)................ 115 Figura A14: Dediferenciación (H&E x200) .............................................................. 115 Figura A15: Focos de carcinoma en lesiones benignas (H&E x100) .......................... 115 Figura A16: Nidos de carcinoma separados por tejido fibroso (H&E x100) .............. 115 Figura A17: Secreción citoplasmática en carcinoma (H&E x200).............................. 115 Figura A18: Patrón microcribiforme (H&E x100) ...................................................... 115 Figura A19: Infiltrado mastocitario en carcinoma de bajo grado arquitectural y nuclear (H&E x200)................................................................................................................... 116 Figura A20: Fibroadenoma pericanalicular (H&E x40) .............................................. 116 Figura A21: Fibroadenoma pericanalicular (H&E x100) ............................................ 116 Figura A22: Fibroadenoma intracanalicular (H&E x40) ............................................. 116 Figura A23: Fibroadenoma intracanalicular (H&E x100) ........................................... 116 Figura A24: Adenoma (H&E x100) ........................................................................... 116 Figura A25: Adenoma (H&E x200) ............................................................................ 117 Figura A26: Fibroma (H&E x100) ............................................................................. 117 Figura A27: Hiperplasia lobulillar (H&E x100) ......................................................... 117 Figura A28: Hendiduras foliáceas en tumor “phyllodes-like” (H&E x40).................. 117 Figura A29: Estroma celular en tumor “phyllodes-like” (H&E x200) ........................ 117 Figura A30: Figuras de mitosis en estroma de tumor “phyllodes-like” (H&E x200) . 117 Artículo 2: Figura A31: HFCOP, grados arquitectural y nuclear elevados (H&E x400) ............... 139 Figura A32: HFCOP, actividad mitótica (H&E x400) ................................................. 139 Figura A33: HFCOP, necrosis tumoral (H&E x40) ..................................................... 139 Figura A34: HFCOP, infiltrado inflamatorio (H&E x100) .......................................... 139 Figura A35: HFCOP, invasión de tejidos adyacentes (H&E x40) ............................... 139 4.
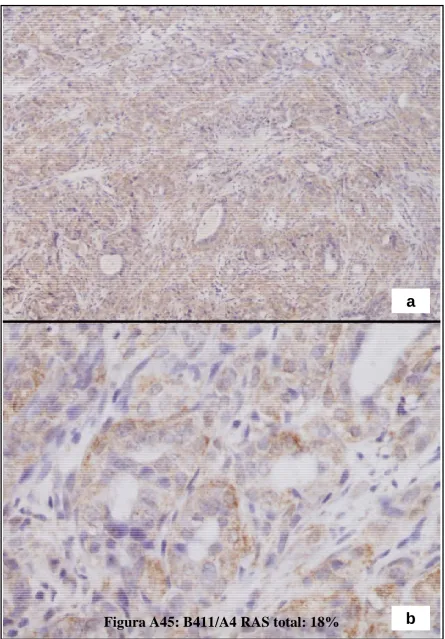
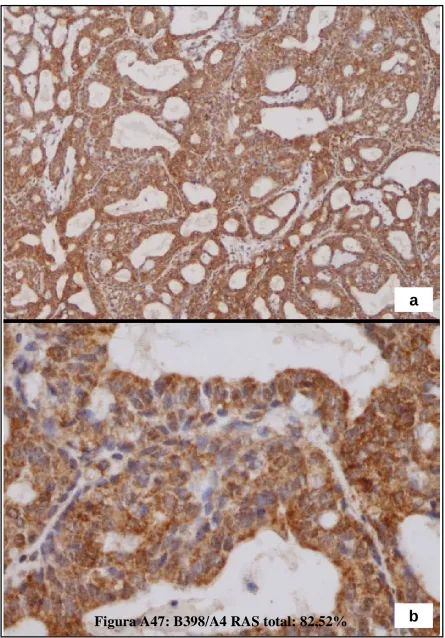
(23) Figura A36: HFCOP, reacción desmoplásica (H&E x200) ..........................................139 Figura A37: HFOOP, grados arquitectural y nuclear bajos (H&E x400) .....................140 Figura A38: HFOOP, actividad mitótica (H&E x400) .................................................140 Figura A39: HFOOP, patrón papilar (H&E x100)........................................................140 Figura A40: HFOOP, márgenes tumorales nodulares (H&E x40) ...............................140 Figura A41: HFOOP, márgenes tumorales nodulares (H&E x100) .............................140 Figura A42: HFOOP, cambio secretor (H&E x200).....................................................140 Artículo 3: Figura A43: Poca expresión de p21Ha-.Ras.................................................................161 Figura A44: Abundante expresión de p21Ha-Ras........................................................161 Figura A45: B411/A4 RAS total: 18%.........................................................................162 Figura A46: B435/A2 RAS total: 38,13%....................................................................163 Figura A47: B398/A4 RAS total: 82,52%....................................................................164 Artículo 4: Figura A48: Expresión del ARNm de PCPH en los tumores mamarios por efecto de una dieta rica en n-6 PUFA ..................................................................................................182 Artículo 5: Figura A49: Imágenes representativas de los tumores de cada grupo experimental ..198 Artículo 6: Figura A50: Imágenes grupo C y HCO........................................................................209 Artículo 10: Figura A51: 36 PND no ovulación...............................................................................281 Figura A52: 51 PND ovulación....................................................................................282 Figura A53: Estructuras mamarias en tejido montado en fresco..................................283. 5.
(24) ÍNDICE DE TABLAS: Tabla 1: Composición característica de los aceites de oliva virgen y de maíz .............. 45 Tabla 2: Grado histológico del carcinoma de mama (Elston y Ellis, 1991) .................. 58 Tabla 3: Clasificación de los tumores de glándula mamaria de la rata (J. Russo y I. Russo) ............................................................................................................................. 72 Tablas adicionales (A) a los artículos (especificadas en el texto de los apartados correspondientes del capítulo de “Resultados”): Artículo 2: Tabla A1: Incidencia tumoral y patología mamaria..................................................... 134 Tabla A2: Lesiones epiteliales proliferativas mamarias .............................................. 135 Tabla A3: Distribución de los grados histológicos de los carcinomas de mama experimentales entre diferentes grupos dietéticos, en cuanto a aporte de grasas, en una nueva serie experimental............................................................................................... 138 Artículo 3: Tabla A4: Estudio inmunohistoquímico de p21Ha-Ras (series A y B) ....................... 159 Tabla A5: Correlación expresión de p21Ha-Ras y grado histológico (serie A) .......... 159 Tabla A6: Correlación expresión de p21Ha-Ras y grado nuclear y arquitectural (serie B). 159 Tabla A7: Correlación expresión de p21Ha-Ras y mitosis (serie B) ........................... 160 Artículo 5: Tabla A8: Parámetros clínicos (I) ................................................................................ 195 Tabla A9: Parámetros clínicos (II)............................................................................... 196 Artículo 9: Tabla A10: Resultados del estudio de la expresión de H19, IGF2, VDUP1 y TRX .. 261 Artículo 10: Tabla A11: Resultados del estudio clínico y morfológico de la glándula mamaria..... 278. 6.
(25) I – INTRODUCCIÓN.
(26)
(27) INTRODUCCIÓN. A. GLÁNDULA MAMARIA NORMAL: DESARROLLO Y MORFOLOGÍA El desarrollo y la función de las glándulas mamarias, humana y de la rata, dependen de múltiples requerimientos hormonales, de componentes de la matriz extracelular y de factores de crecimiento. A pesar de ello, se reconocen algunas diferencias entre ambas especies. 1. HUMANA El desarrollo de la glándula mamaria humana se esquematiza en la Figura 1. Figura 1: Desarrollo de la glándula mamaria humana NACIMIENTO 7 SEMANAS 5º MES. PERIODO ENTRE MENARQUIA Y GESTACIÓN PRE-PUBERTAD 1ª OVULACIÓN. LINEA MAMARIA ESBOZOS SECUNDARIOS. 1ª GESTACIÓN. 1er TRIMESTRE. NULÍPARA PREMENOPÁUSICA. 3er TRIMESTRE Y LACTANCIA. POSMENOPÁUSICA. Yemas terminales (TEBs) Lobulillos 1. MENOPAUSIA. INVOLUCIÓN POSTLACTANCIA. Lobulillos 2 Lobulillos 3 Lobulillos 4. MULTÍPARA PREMENOPÁUSICA. Embriológicamente, la glándula mamaria deriva de un esbozo ectodérmico, conocido como línea o banda mamaria, que aparece hacia la 5ª semana de vida como un par de áreas de engrosamiento de la epidermis que se extienden desde las axilas a la región inguinal. Hacia la 7ª semana de vida, las líneas mamarias involucionan restando como dos masas epiteliales sólidas en la región torácica que empiezan a crecer hacia el mesénquima subyacente. En esta fase intervienen la testosterona, que determina el patrón de crecimiento mamario masculino, el estradiol y las hormonas lactógenas (583, 585, 1030). Hacia el quinto mes de vida fetal, se forman los esbozos secundarios de los que se originan los ductos galactóforos que desembocan en el pezón (585, 3303). Esta etapa parece estar controlada por el estradiol y la progesterona de la placenta y la prolactina fetal. En el recién nacido, la glándula mamaria consiste en un sistema rudimentario ramificado de ductos que permanece quiescente hasta la pubertad. El crecimiento de la mama será isométrico con la proliferación del tejido estromal y proporcional al crecimiento medio del cuerpo (583, 586, 1030, 2633). Poco antes de. 9.
(28) INTRODUCCIÓN. la menarquia, y con el inicio de la maduración folicular en el ovario, el parénquima mamario entra en una fase de crecimiento, con división de la estructura tubular y aparición de los ductos mamarios primarios y secundarios, que terminan en unas formaciones bulbosas o yemas terminales ([3503], [7671]). Con las primeras ovulaciones, estas formaciones inmaduras dan lugar a las yemas alveolares o ductulillos, que se disponen alrededor de un ducto, constituyendo los lobulillos tipo 1, que representan la unidad funcional básica de la mama humana o unidad ductulo-lobulillar terminal (“terminal ductal lobular unit” o TDLU). Cada lobulillo tipo 1 contiene de 6 a 11 ductulillos. Durante la etapa fértil de la mujer, se observan dos formas adicionales de lobulillos, el tipo 2 y el 3, cuya transición se asocia a formación de nuevas yemas alveolares (hasta 47 en el tipo 2 y hasta 80 en el tipo 3), con un incremento en el tamaño lobulillar. Los lobulillos tipo 1 y 2 se observan predominantemente en mujeres nulíparas, mientras que la progresión a lobulillo tipo 3 es estimulada por la gestación ([3165], [3503]). El desarrollo de los lobulillos está regulado por los cambios hormonales que acontecen durante cada ciclo menstrual y, en especial, durante el embarazo. Así, durante la fase folicular se produce un crecimiento activo del sistema glandular, que alcanza un máximo en la fase luteal y va seguido de una rápida involución. El balance entre proliferación e involución en edades reproductivas tempranas favorece la proliferación, con una progresión de la población celular en cada ciclo ovulatorio (2631, 2633, 2638, [3123], [3332]). Los cambios cíclicos en los niveles de estrógenos y progesterona ováricos durante cada ciclo menstrual y su acción sinérgica son necesarios para el crecimiento armónico de la glándula mamaria (2631, [4577]). Los estrógenos controlan la proliferación celular, induciendo la síntesis de receptores de progesterona en las células que contienen receptores de estrógenos y la secreción de prolactina hipofisaria y estimulando a células epiteliales del lobulillo que presentan sus receptores a producir factores de crecimiento que, a su vez, inducen la proliferación de las células vecinas ([2633], [3965], [6797]). En cuanto a la progesterona, el mecanismo subyacente no está claro y parece que actuaría mediando la regulación de genes del ciclo celular, de factores de crecimiento y de sus receptores ([3332]). En esta fase de la mamogénesis intervienen, además, prolactina, insulina, cortisol, tiroxina, hormona liberadora de gonadotrofinas (GnRH) y hormona de crecimiento, que interactúan entre ellas y cuyas acciones pueden ser directas y/o indirectas a través de factores de crecimiento (583, 586, 1030, [2631], 2632, 2639], [3332], [3334], [3372]). Entre ellos están el factor de crecimiento epidérmico (EGF), el factor de crecimiento transformante (TGF-), la familia del factor de crecimiento transformante (TGF-) y el factor de crecimiento parecido a Insulina I (IGF-I) ([2630], 2633, 2638, [3164]). Diversos factores 10.
(29) INTRODUCCIÓN. de crecimiento median sus efectos sobre el crecimiento a través de cambios en el “turnover” de los componentes de la matriz extracelular secretados por las células ([2633]), como las metaloproteinasas de la matriz (MMPs) y sus inhibidores (TIMPs) ([3372]). El tejido adiposo mamario también posee una influencia significativa sobre el crecimiento y el desarrollo del epitelio glandular, de modo que es necesaria la interacción entre ambos tejidos para que se produzca la morfogénesis mamaria normal (2633). La mama humana adulta normal consta de 15 a 25 ductos galactóforos que drenan los lóbulos mamarios que conforman la glándula mamaria. Los lóbulos mamarios están constituidos, a su vez, por numerosos lobulillos. Los ductos galactóforos nacen en el pezón, descienden una corta distancia, donde se expanden como ampollas (senos lactíferos), ramificándose en conductos más delgados que conducen a los lobulillos. En el pezón se observan, además, los tubérculos de Montgomery, que corresponden a glándulas sebáceas modificadas, asociados a la porción terminal de los ductos galactóforos. Éstos están revestidos, en la zona próxima a la apertura al pezón, por un epitelio escamoso queratinizante, extensión del epitelio escamoso de la piel. Entre las células epiteliales de las partes externas de los conductos galactóforos y de la piel del pezón y la areola se encuentran melanocitos dendríticos que confieren a estas zonas una pigmentación más oscura que la de la epidermis que los circunda. Los ductos mamarios distales a los galactóforos muestran dos capas de células: una interna de células epiteliales luminares y otra externa, mioepitelial. La capa de células mioepiteliales es discontinua en las zonas más distales y se apoya en una membrana basal que, a su vez, está rodeada por fibras elásticas, ausentes en los ductos y ductulillos intralobulillares ([3923]). Estudios de caracterización fenotípica de las células que revisten los lobulillos mamarios han demostrado una heterogeneidad celular considerable, con presencia de células que expresan citoqueratinas (CK) de alto (CK5/6/14) y bajo peso molecular (CK8/18/19) y marcadores de diferenciación a músculo liso, como la actina músculo liso específica (“smooth muscle” o sm-actina). Parece ser que las células que expresan CK5/6/14 serían precursoras comunes, diferenciándose progresivamente, por un lado, hacia una vía epitelial luminar (CK8/18/19 positiva) y, por otro lado, hacia células mioepiteliales que expresan sm-actina, y se localizan tanto en la capa luminar como en la capa basal del ductulillo mamario ([7266]). El tejido conectivo intralobulillar es un tejido especializado, hormono-sensible, de fibras de colágena distribuidas sobre una matriz mucopolisacárida ácida, con fibroblastos, ocasionales linfocitos y macrófagos, pero sin grasa, y muestra mayor vascularización que el tejido conectivo interlobulillar que es hipocelular y densamente colagenizado. A partir de los 18 años, el tejido conectivo interlobulillar 11.
(30) INTRODUCCIÓN. evoluciona progresivamente a fibroadiposo. Pueden observarse ganglios linfáticos intramamarios, normalmente menores de 5 mm de diámetro ([3923]). Durante el embarazo ocurre la expansión plena del sistema lobulilloalveolar, necesaria para la secreción de leche, alcanzándose el estado de máxima diferenciación de la glándula mamaria ([453], [583], [1030], [2631], [2632], [2636], [2637]). A lo largo del primer trimestre de gestación, se observa una proliferación y una ramificación de los elementos distales del árbol ductal, con aparición de numerosos lobulillos tipo 3 ([2633], [3503]). El epitelio glandular proliferante empieza a sustituir al tejido conectivo y adiposo y se produce un nuevo desarrollo de la vascularización. En el segundo trimestre, se observa diferenciación de los ductulillos a acinos o alvéolos, con capacidad de secreción láctea, constituyendo los lobulillos tipo 4 que aumentan progresivamente y se hacen más secretores ([3503]). Durante el último trimestre, se produce un rápido desarrollo lobulilloalveolar, el flujo sanguíneo mamario aumenta y las células mioepiteliales se hipertrofian. El crecimiento inicial del sistema ductal depende de los estrógenos, en sinergia con la hormona de crecimiento y el cortisol, mientras que la diferenciación del sistema lobulilloalveolar requiere de los estrógenos y la progesterona en presencia de la prolactina ([2632], [3332]). En las primeras fases del embarazo, el cuerpo lúteo materno es mantenido por la hormona gonadotropina coriónica (hCG), secretada por el trofoblasto primitivo, y produce progesterona, 17-hidroxiprogesterona, estradiol, inhibina y relaxina. A partir de la novena semana de gestación, el sincitiotrofoblasto placentario se convertirá en el principal productor de gran parte de estas hormonas, sintetizando progesterona, estriol, hCG y hormona del lactógeno placentario (hPL). La hCG se considera la principal responsable de la diferenciación glandular, al ejercer un efecto inhibidor directo sobre la proliferación del epitelio mamario humano de forma directa e, indirectamente, al inducir la síntesis de inhibina, al activar genes reguladores de la apoptosis y al incrementar la capacidad de reparación del ADN ([2406], [3165], [3370], [3400]). La hPL regula el metabolismo materno, aumentando la concentración de glucosa y ácidos grasos libres, y presenta propiedades lactógenas, además de un leve efecto tipo hormona de crecimiento ([2632], [2634]). La prolactina es la hormona clave que controla la producción de la leche. Al inicio del embarazo, la principal fuente de prolactina es la decidua, que alcanza un valor máximo en el segundo trimestre, mientras que la prolactina hipofisaria materna y fetal aumenta progresivamente hasta alcanzar, al término del embarazo, niveles diez veces superiores a los de la mujer no embarazada. Este incremento en la secreción de prolactina hipofisaria resulta de la inhibición de los elevados niveles de estrógenos y progesterona sobre el factor 12.
(31) INTRODUCCIÓN. hipotalámico inhibidor de la secreción de prolactina (PIF) o dopamina ([2636], [2637]). La prolactina induce proliferación y diferenciación de células mamarias indiferenciadas para formar células alveolares presecretoras y células mioepiteliales y regula la síntesis de las proteínas (caseína y -lactalbúmina) y las grasas de la leche. Además, se requieren otras hormonas, como glucocorticoides suprarrenales, hormona de crecimiento, hormona paratiroidea, tiroxina, insulina y los factores de crecimiento IGF-I e IGF-II, para proporcionar los aminoácidos, los ácidos grasos, la glucosa y los minerales imprescindibles para la formación de leche ([2634]). Durante el embarazo, la inhibición de los estrógenos y progesterona, a nivel periférico, de las acciones lactogénicas de la prolactina y de la hPL impiden la secreción láctea ([3332]). Las células epiteliales de los acinos y los ductos intralobulillares muestran vesículas y gotas de grasa intracitoplasmáticas y secreción en la luz, aumentando el tamaño de los lobulillos ([2633]). Las células adiposas pueden perder progresivamente sus reservas de grasa durante las últimas fases del embarazo y de la lactancia; por lo general no desaparecen aunque estén vacías y pueden volver a acumular grasa una vez que ha cesado la lactancia. Después del parto y tras la eliminación de la placenta se produce una caída brusca de los niveles de estrógenos y progesterona, permitiendo a la prolactina ejercer su papel lactogénico ([3332], [3503]). A los 3 ó 4 días después del parto, las mamas comienzan a secretar grandes cantidades de leche en lugar del escaso calostro de los primeros días. Durante la lactancia se para la maduración folicular y la ovulación. El estado hormonal que estimula y mantiene la lactancia está garantizado por un reflejo neurohormonal inducido por el efecto mecánico de la succión que permite la secreción de prolactina. En las primeras semanas posparto, los niveles plasmáticos basales de prolactina son elevados, disminuyendo hasta niveles normales a partir de la sexta semana, permaneciendo el efecto estimulador de la succión. La cantidad de prolactina liberada está determinada por la intensidad y la duración de esta estimulación y, su efecto, por un aumento progresivo del número de sus receptores en las células alveolares ([2631], [2634]). El mismo reflejo producido por la succión provoca la estimulación de la secreción de la oxitocina hipotalámica, que contrae a las células mioepiteliales que rodean los acinos, forzando la eyección láctea. Además, podría inducir indirectamente la relajación y la apertura de los conductos de mayor tamaño ([3302], [3335]). Cuando cesa la lactancia, la glándula mamaria interrumpe su actividad secretora y sufre una fase de regresión rápida, con una disminución de los elementos glandulares y reaparición del componente estromal interlobulillar y adiposo. Dentro del epitelio alveolar aparece una importante actividad autofágica y lisosomal que elimina los restos celulares. Se 13.
(32) INTRODUCCIÓN. ha observado sobreexpresión de genes proapoptóticos, como bcl-xs, p53, c-fos, c-jun o bax, regulada por factores de crecimiento y equilibrada por los TIMPs ([3104], [3105]). La mayoría de los lobulillos regresan, retornando a lobulillos tipo 3 y 2. El lobulillo tipo 3 es la estructura glandular predominante en las mamas de mujeres premenopáusicas multíparas [3165], ([3503]). Las gestaciones repetidas extienden la diferenciación lobulillar. Después de la menopausia, la caída de las hormonas ováricas causa la involución progresiva de los tejidos epitelial y conectivo. El componente ductulo-lobulillar se atrofia, la matriz fibrosa se hace más densa y el parénquima es sustituido por tejido adiposo y conectivo ([2632]). La mayoría de los lobulillos adquieren la apariencia morfológica de los tipos 1. La actividad proliferativa de estos lobulillos tipo 1 es menor en las glándulas mamarias de mujeres posmenopáusicas multíparas que en las nulíparas, lo que indica que la gestación puede imprimir cambios permanentes en las características biológicas de estas estructuras ([3165], [3923], [3924]). Como se comentará en el apartado B.3.a de la “Introducción”, el grado de diferenciación al cual llega la glándula mamaria durante el embarazo es un factor importante que se opone a la transformación neoplásica del órgano, lo que determina una diferencia fundamental entre las mujeres nulíparas y las que han tenido por lo menos un hijo, en cuanto a factores de riesgo de cáncer mamario ([603], [861], [2226], [7671]). 2. RATA El desarrollo de la glándula mamaria de la rata hembra virgen nulípara se esquematiza en la Figura 2. Figura 2: Desarrollo de la glándula mamaria de la rata virgen nulípara. 11 DIA. NACIMIENTO 14 1ª SEMANA 2ª SEMANA DIA. FENOTIPO SEXUAL. TEBs ABs Lobulillos TDs. 14. 30 A 55 DIAS EDAD ↑↑ TEBs A ABSs. 3ª SETMANA 21 DIAS. 84 DIAS. 180 DIAS. INICIO ACTIVIDAD OVÁRICA 25-42 DIAS EDAD. Nº TEBs máximo. Nº CONSTANTE ABs y lobulillos. ↓TEBs y ↑TDs.
(33) INTRODUCCIÓN. La rata posee 6 pares de glándulas mamarias distribuidas subcutáneamente a lo largo de dos líneas ventrolaterales, desde la región cervical a la inguinal: un par en la región cervical, dos en la región torácica, dos en la abdominal y uno en la inguinal (1141). La única porción externamente visible en la rata hembra es el pezón, mientras que el macho no posee pezones y, normalmente, presenta cuatro pares de glándulas rudimentarias. El primer esbozo de glándula mamaria aparece sobre el día 11 de gestación como dos estructuras de crecimiento alargadas de ectodermo que se extienden ventralmente desde la región cervical hasta la inguinal y que inducen, a su vez, el crecimiento del mesénquima subyacente. Hacia el día 14 de gestación, el fenotipo sexual de la glándula mamaria se halla determinado. En los machos, la testosterona secretada por el teste no permite el desarrollo de la glándula mamaria, que sí se desarrolla en la hembra con la aparición de los ductos galactóforos primordiales ([2633]). Al nacer, las glándulas mamarias de las ratas hembras constituyen parches de color claro en la piel, con pezones rudimentarios. En la época lactante, factores de crecimiento presentes en la leche materna, como el EGF, intervienen en el crecimiento y desarrollo de la mama de estos animales. En la rata Sprague-Dawley, la glándula mamaria evoluciona desde un ducto galactóforo mayor primordial que nace del pezón y que se ramifica en ductos secundarios, cuyas longitudes y nº de ramas secundarias y terciarias aumentarán con la edad del animal ([3122]). Así, entre el nacimiento y la primera semana de vida, se observa un ducto galactóforo con 3 a 5 ramificaciones a ductos secundarios, que hacia la segunda semana de vida mostrarán seis generaciones de ramificaciones que acaban en las yemas terminales (o “terminal end buds” – TEBs), considerados los centros de crecimiento activo y el elemento primitivo del parénquima mamario. El nº de TEBs es máximo cuando la rata tiene 21 días de edad. A partir de aquí, y coincidiendo con el período prepuberal y el inicio de la actividad ovárica, los TEBs inician la diferenciación y septación a 3 a 5 yemas alveolares (o “alveolar buds” - ABs). Esta diferenciación progresiva de los TEBs a ABs se acentuará en cada ciclo estral, que se inicia cuando el animal tiene de 25 a 42 días de edad, y éstos, a su vez y en los sucesivos ciclos estrales, iniciaran el desarrollo lobulillar, con aparición de lobulillos tipo 1 y 2 ([3165]). En las ratas vírgenes este proceso continúa durante la maduración sexual hasta que se consigue un nº constante de ABs y lobulillos, que acontece hacia los 84 días de edad, pudiendo observarse una disminución discreta a partir de los 180 días de edad del animal (943). Un nº elevado de TEBs sufrirá hipoplasia en la rata virgen, constituyendo los ductos terminales (TDs). Por lo tanto, la glándula mamaria de una rata virgen joven, en relación a una rata virgen adulta, contiene mayor número de TEBs, menor número de TDs y similar cantidad de ABs y lobulillos. Estos componentes estructurales de la 15.
(34) INTRODUCCIÓN. mama permanecerán inalterados mientras la rata siga virgen. En la hembra no gestante, el desarrollo de la glándula mamaria está controlado por el ovario que depende, a su vez, de las gonadotrofinas hipofisarias. Éstas interactuarán con la hormona de crecimiento (GH) y la prolactina para modular la esteroidogénesis ovárica, así como la secreción de glicoproteínas no esteroidales, como la inhibina. Parece ser que los estrógenos son los responsables del crecimiento de los ductos mamarios y la progesterona del desarrollo lobulilloalveolar. Los estrógenos actuarían a través de un efecto mitogénico directo e indirecto, a través de la estimulación de la secreción de factores de crecimiento (como EGF) de origen mamario o extramamario ([2633], [3966]). El desarrollo ductal requiere, además, de la insulina y la GH hipofisaria, que a su vez actúa directamente o a través de su mediador local, el IGF-I. Por otro lado, los estrógenos estimulan la secreción de la prolactina hipofisaria e inducen los receptores de progesterona, potenciando el desarrollo lobulilloalveolar. Desde el punto de vista histológico, los ductos mamarios están revestidos por dos capas de células, una interna epitelial y otra externa mioepitelial que descansa sobre la membrana basal. Los ductos mayores y galactóforos presentan un revestimiento epitelial pseudoestratificado, de células cuboideas. Los TEBs, tal como se muestra en la Figura 3, están constituidos por 4 o 6 capas de células, fenotípicamente diferentes. En la superficie basal de la parte más distal de los TEBs se hallan las células “cap”, que carecen de estigmas de diferenciación y que representan una población celular pluripotencial capaz de diferenciarse a células epiteliales y mioepiteliales ([3122]). Se observan, además, otros tipos de células como las intermedias que, a diferencia de las células oscuras o “dark” que son más numerosas en los lobulillos, no presentan estigmas de actividad secretora. Figura 3: Estructura de una yema terminal (TEB). Células “cap” Células “dark”. Células epiteliales. Células intermedias Membrana basal Células mioepiteliales. H&E x100. Se reconocen diferentes áreas topográficas en la mama de la rata que difieren en su morfología, características quinéticas, respuesta a estímulo hormonal y potencial 16.
(35) INTRODUCCIÓN. carcinogénico (3573). Estas áreas, esquematizadas en la Figura 4, resultan de dividir en tres tercios el parénquima mamario, a través del eje longitudinal, y ayudan a comprender el proceso de desarrollo y diferenciación mamario. Figura 4: Topografía de la glándula mamaria de la rata. LOBULILLO. LOBULILLOS. ABs. ABs. TEBs. TEBs. TDs. PEZÓN TDs. PIEL. AREA A. AREA B. AREA C. El área A es la más próxima al pezón, constituida por ductos galactóforos principales y secundarios, la B es la intermedia, con ductos de menor tamaño y la C es la más alejada del pezón, rica en estructuras ductales terminales. Durante el proceso de maduración lobulilloalveolar se describe un gradiente de desarrollo mamario. Así, el nº de TEBs disminuye marcadamente en las zonas A y B, en las que predominan los lobulillos y los ABs, respectivamente, pero no en la C, en la que predominan los TEBs y TDs, de crecimiento activo. Aunque todas las glándulas mamarias están constituidas por las tres zonas descritas, la cantidad y el tamaño de cada una de ellas difieren según la localización anatómica. Así, las glándulas torácicas, especialmente el tercer par, tienen más y mayores TEBs que las abdominales ya que muestran un enlentecimiento en la diferenciación que desaparece con la edad del animal ([2952], [3503]). Al igual que en la mama humana, se reconocen diferentes lobulillos que maduran de estructuras menos complejas, los lobulillos tipo 1 (con 6-10 ductulillos), a lobulillos tipo 2 (con 22-32 ductulillos) y a tipo 3 (con 53-90 ductulillos). El desarrollo glandular dependerá de diversos factores, como el nº de ciclos ocurridos, la dieta y las características genéticas específicas del animal, pero la diferenciación completa de la glándula mamaria se alcanza 17.
(36) INTRODUCCIÓN. con el embarazo y la lactancia. Durante la gestación, diferentes estímulos hormonales, entre ellos la prolactina, los estrógenos, la progesterona y la inhibina ováricas, así como la producción de hCG y hPL por el embrión que se desarrolla, contribuyen a desarrollar la glándula mamaria. Los lobulillos durante la gestación y la lactancia son de tipo 3 y 4, con numerosos acinos. Esta diferenciación es mayor en el área A y, en segundo lugar, en el área B del parénquima mamario. Aquellos animales que han completado la gestación muestran, además, extenso desarrollo lobulillar en la zona C (3573). La relaxina hipofisaria también estimula el desarrollo de los lobulillos y su capacidad de lactogénesis, y juega un papel importante en el desarrollo de los pezones durante la segunda mitad de la gestación. Al final de la gestación, la glándula mamaria está constituida por lobulillos con actividad secretora. La lactancia, que dura unas 3 semanas, retrasa la reiniciación del ciclo estral y la ovulación. Los niveles de prolactina y del ARNm de su receptor aumentan en las glándulas mamarias. Con el destete, los niveles de prolactina vuelven a la normalidad y el folículo ovárico se desarrolla hasta un estadio preovulatorio, secretando 17-estradiol en cantidades suficientes para inducir la secreción preovulatoria de gonadotropinas que reiniciarán el ciclo estral normal. La glándula mamaria disminuye de peso y tamaño y gran parte del parénquima es reemplazado por grasa, con colapso de los alvéolos secretores y eliminación activa de las células y secreciones por macrófagos. A pesar de que la involución de la glándula mamaria es aparentemente completa a los 10 días del destete, la arquitectura de la glándula mamaria restará permanentemente modificada. Las glándulas mamarias de un animal que ha completado totalmente la gestación y la lactancia retienen gran nº de ABs y lobulillos, resultado de la hipoplasia de los lobulillos de la gestación. Se ha descrito que a los 40-42 días posdestete, las glándulas mamarias de una rata multípara, en comparación con las de una rata nulípara, no tienen TEBs y sólo ocasionales TDs. El nº de ABs es similar, mientras que el nº de lobulillos es aproximadamente 4 veces mayor que el de una rata virgen adulta. Se reconocen diferencias biológicas entre las distintas estructuras anatómicas de la glándula mamaria de la rata. La determinación del índice de marcaje de ADN (ADN-LI) revela que el 100% de los TEBs presentes en una rata nulípara joven y adulta contienen células proliferando que incorporan el precursor marcado, siendo el porcentaje de células marcadas inferior en las ratas vírgenes adultas (14,8% versus 34.4%). El ADN-LI de los TDs se halla también reducido en las ratas vírgenes adultas, en relación con las jóvenes. En las glándulas en reposo de las ratas multíparas, sólo un 8% de los ocasionales TDs contienen células marcadas, con un ADN-LI muy bajo. En relación a los ABs, las ratas vírgenes jóvenes y adultas tienen el 50% y el 5% de sus ABs marcados, respectivamente, mientras que 18.
(37) INTRODUCCIÓN. en las glándulas en reposo de las ratas multíparas, sólo el 0.9% se marcan, con un ADN-LI bajo. Las estructuras lobulillares no incorporan el precursor de ADN (3122). Como se verá en el próximo apartado, el conocimiento de los múltiples factores implicados en el desarrollo y la función de la glándula mamaria normal resulta fundamental para comprender los mecanismos por los cuales se producen los procesos patológicos neoplásicos en este órgano.. B. CÁNCER DE MAMA El cáncer de mama es la neoplasia maligna más frecuente entre las mujeres en todo el mundo ([6691], [7519]) y el responsable de aproximadamente 1 de los 10 millones de neoplasias diagnosticadas cada año en ambos sexos y el 23% de todos los tipos de cáncer ([4647]). En España representa el 25%, y, en Cataluña, el 28%, de todo el conjunto de neoplasias malignas en mujeres ([3412], [4333]). En los países más desarrollados, se ha constatado un aumento acelerado de la incidencia en los últimos 20 años, que se ha atribuido a una mayor exposición a factores de riesgo y a la introducción de métodos diagnósticos que permiten detectar tumores incipientes ([4333], [4472], [7519]). Es, además, la primera causa de muerte por cáncer en este sexo, ocasionando 1 de cada 3 muertes por cáncer en el grupo de mujeres de edades entre 35 y 64 años ([3908], [4473], [4647], [7108]). Se ha detectado una tendencia al incremento de las tasas de mortalidad por cáncer de mama en el grupo de mujeres mayores de 50 años, con un descenso en las menores de 40 años, probablemente debido a la detección precoz y a los avances terapéuticos introducidos en las últimas décadas. Por otra parte, los estudios epidemiológicos muestran variaciones geográficas importantes de la incidencia del cáncer de mama, que es más elevada en países occidentales que en los subdesarrollados ([6093], [7108]) (Figura 5). Así, las tasa brutas de incidencia de cáncer de mama más altas a escala mundial corresponden a la población blanca de Estados Unidos. Los registros poblacionales de Israel, Canadá y diversos países europeos están en la lista de las áreas de tasas de incidencia más elevadas, mientras que las más bajas se registran en África y Asia y en la población no judía de Israel. En España la incidencia es más baja que en otros países de Europa, aunque se ha registrado un aumento del número de casos desde la década de 1980 hasta el año 2000 ([7672]). De hecho, según los últimos datos publicados, en España, el cáncer de mama es el que presenta mayor incidencia y mortalidad ([www.globocan.iarc.fr]). En relación a Cataluña, la incidencia de esta enfermedad ha aumentado en los últimos años, aunque en el contexto europeo sigue siendo de los más bajos ([7673], [7674]). 19.
(38) INTRODUCCIÓN. Varios estudios demuestran, además, que la población emigrada de un país de origen a otro de adopción asume las tasas de incidencia y mortalidad del último en un período de una a dos generaciones ([6093], [6862]). Estos datos sugieren que el grado de desarrollo y/o industrialización, los factores ambientales y el estilo de vida están claramente involucrados en estas diferencias, tal como se comentará en el siguiente subapartado. Figura 5. Tasas de incidencia por países de cáncer de mama en mujeres. Globocan, 2008 ([7108]). 1. ETIOLOGÍA DEL CÁNCER DE MAMA El cáncer representa un desorden del crecimiento celular, caracterizado por una proliferación incontrolada de las células de un determinado tejido, sin relación con las demandas fisiológicas del órgano del que forman parte, y que, además, muestran capacidad de invadir y destruir los tejidos adyacentes y diseminarse a distancia, con la consiguiente evolución fatal a corto, medio o largo plazo, según el tipo de cáncer. En la mama se reconocen diversas neoplasias que se originan de los distintos tejidos que la configuran y que se clasifican siguiendo criterios fundamentalmente histológicos. El cáncer que se origina del tejido epitelial o glandular es el carcinoma, constituye el grupo de neoplasias malignas más frecuente de la mama y es sobre el que se refieren la mayor parte de los estudios. El cáncer de mama es un proceso neoplásico de etiología multifactorial. Diversos factores, actuando de forma simultánea y/o secuencial, regulan las distintas etapas de la carcinogénesis mamaria. La naturaleza de estos factores es variada y se pueden clasificar en tres grupos: endocrinos, genéticos y ambientales. Estos factores no actúan aisladamente, sino que muestran una clara interdependencia entre ellos. 20.
Figure


+7

Documento similar
Analysis of breast cancers from The Cancer Genome Atlas (TCGA) showed that HSD17B14 expression is increased significantly in cancer compared with normal breast tissue and that it
The sst5 truncated isoform, sst5TMD4, is expressed in primary breast cancer samples and increases the malignancy of breast MCF-7 cellsS. Expression of the ghrelin and
a) To systematically review the indicators of PPF in breast cancer. i) To study the associations between the main constructs from positive psychology and breast cancer. ii)
All tumors were assigned an intrinsic molecular subtypes of breast cancer (luminal A, luminal B, HER2-enriched, and basal-like) and the normal-like group using the PAM50
SFKs participate in the development and progression of several human cancers, including breast cancer. To investigate the role of c-Src in the first steps that lead to metastasis,
An ER-true/TN-like classifier, including 14 proteins, was used to assign new samples to ER-true or TN-like (sup. We used gene expression data from 1296 breast cancer tumors,
Triple-negative breast cancer (TNBC) is am immunohistochemically-defined breast cancer subtype negative for ER, PR and HER2 expression, with dismal prognosis that
The two major breast cancer susceptibility genes BRCA1 and BRCA2 are involved in 30% of hereditary breast cancer cases, but the discovery of additional breast cancer
