Determinación del crosstalk estructural entre los receptores de membrana 5 HT3a y P2X4 mediante microscopía de fuerza atómica
65
0
0
Texto completo
(2) ii. DEDICATORIA. Esta tesis doctoral está dedicada a mi padre, Juan Bernardo Soto (1940-2018) y a mi madre, María Elena Sepúlveda. Me motivaron y apoyaron constantemente para lograr mis sueños. Muchos de mis logros se los debo a ustedes, el doctorado es uno de ellos..
(3) iii. AGRADECIMIENTOS. Quiero agradecer a todas las personas que me ayudaron y participaron de alguna manera en la realización de esta tesis doctoral. Quiero agradecer al Dr. Nelson Barrera por darme la oportunidad de ser parte de su equipo, trabajar en su laboratorio y apoyarme en el diseño, supervisión y discusión de la tesis. No sólo contribuyó en mi formación científica, lo hizo también en lo personal con sus consejos y compañía en difíciles momentos. Quiero agradecer al Dr. Michael Edwardson (Departamento de Farmacología, Universidad de Cambridge, Reino Unido), por permitirme hacer una pasantía en su laboratorio, lo que se convirtió en una enriquecedora experiencia. Además, por proporcionar los constructos de receptores 5-HT3a y P2X4, y por sus comentarios científicos sobre el diseño experimental, el análisis y procedimiento. Quiero agradecer al Dr. Gareth Owen, Dr. Alejandro Godoy, Dr. Manuel Villalón y Dr. Ricardo Moreno, por el préstamo de equipos y reactivos cuando lo necesité. Quiero agradecer a cada uno de mis compañeros de laboratorio, en especial a Jorge Cantero, por su ayuda técnica, comentarios y consejos que fueron útiles en este proceso. Quiero agradecer a todo el quinto piso del Departamento de Fisiología, fueron años de compartir momentos ligados a la ciencia y a nuestra vida personal, puedo decir que hice grandes amigos, no sólo una tesis. Quiero agradecer a mis amigos, fueron muy importantes en este largo proceso, me apoyaron cuando el doctorado mostró su lado poco amable, estuvieron conmigo en los buenos y en los malos momentos, no me dejaron caer..
(4) iv. De manera especial, agradezco a mis padres por su apoyo incondicional en todas las decisiones que he tomado, por la paciencia, por confiar plenamente en mí, por alentarme a ser mejor cada día. Finalmente, agradezco a las fuentes de financiamiento que hicieron posible la realización de esta tesis doctoral: Beca de Ayudante Becario VRI-CPD (2013-2014) Beca de Instructor Becario VRI (2015-2016) Beca de Estadía en el Extranjero para Tesistas de Doctorado VRI-CPD (2016) Beca de Residencia VRI-CPD (primer semestre 2017) Proyecto DPI20140080 (2015-2016)..
(5) v. ÍNDICE DE MATERIAS. DEDICATORIA ......................................................................................................................... ii AGRADECIMIENTOS ............................................................................................................. iii ÍNDICE DE MATERIAS .......................................................................................................... ix ÍNDICE DE FIGURAS ............................................................................................................. ix LISTA DE ABREVIATURAS ................................................................................................ ixx RESUMEN ................................................................................................................................. 1 ABSTRACT................................................................................................................................ 3 INTRODUCCIÓN ...................................................................................................................... 5 Crosstalk entre proteínas de membrana ..................................................................................... 5 Crosstalk funcional entre receptores de membrana de la familia Cys-loop y P2X .................... 6 Crosstalk e interacción física entre receptores de membrana 5-HT3a y P2X4 .......................... 8 Los receptores 5-HT3a y P2X4 y su implicancia fisiológica ..................................................... 9 Estructura del receptor 5-HT3a................................................................................................. 11 Estructura del receptor P2X4 ................................................................................................... 13 Estudios topográficos mediante AFM como herramienta de Biología Estructural ................. 15 HIPÓTESIS .............................................................................................................................. 18 OBJETIVO GENERAL ............................................................................................................ 18 OBJETIVOS ESPECIFÍCOS ................................................................................................... 18.
(6) vi. MATERIALES Y MÉTODOS ................................................................................................. 19 Transformación de células competentes con pDNA del receptor 5-HT3a y P2X4 .................. 19 Purificación de pDNA del receptor 5-HT3a y P2X4 ................................................................ 19 Cultivo celular y transfección de células tsA 201 con pDNA del receptor 5-HT3a y P2X4.... 20 Solubilización y purificación del receptor 5-HT3a conteniendo el tag Myc/His-6 .................. 21 Solubilización y purificación del receptor P2X4 conteniendo el tag HA................................. 22 Solubilización y purificación del complejo de receptores 5-HT3a-P2X4 conteniendo el tag HA ............................................................................................................................................. 23 Análisis de inmunofluorescencia del receptor 5-HT3a y P2X4 ............................................... 24 Análisis de western blot del receptor 5-HT3a y P2X4 ............................................................. 24 Formación de liposomas e inserción de los complejos de receptores 5-HT3a-P2X4 ............... 25 Imágenes de los complejos de receptores 5-HT3a-P2X4 por AFM ......................................... 26 Medición de la concentración intracelular de calcio ................................................................ 28 RESULTADOS ........................................................................................................................ 29 Co-localización de los receptores 5-HT3a y P2X4 expresados en células tsA 201 mediante inmunofluorescencia. ................................................................................................................ 29 Detección de los receptores 5-HT3a y P2X4 mediante análisis de western blot. ..................... 31 Análisis topográfico del complejo 5-HT3a-P2X4 mediante AFM. .......................................... 33 Distribución de frecuencias de los volúmenes moleculares del complejo 5-HT3a-P2X4. ....... 34.
(7) vii. Cambios dinámicos del complejo de receptores 5-HT3a-P2X4 en presencia de ATP mediante HS-AFM. .................................................................................................................................. 38 El receptor 5-HT3a regula la señalización de calcio intracelular a través del receptor P2X4 en presencia de ATP ...................................................................................................................... 40 DISCUSIÓN ............................................................................................................................. 42 CONCLUSIÓN ......................................................................................................................... 47 BIBLIOGRAFÍA ...................................................................................................................... 49.
(8) viii. ÍNDICE DE FIGURAS. FIGURA 1. Crosstalk estructural entre los receptores de membrana 5-HT3a y P2X4 ........... 17 FIGURA 2. Esquema representativo de una muestra de receptores 5-HT3a y P2X4 insertos en liposoma y analisados mediante HS-AFM ............................................................................... 26 FIGURA 3. Esquema representativo de una muestra de receptores 5-HT3a y P2X4 en una micela de detergente (CHAPS) analizados mediante AFM ..................................................... 28 FIGURA 4. Co-localización de los receptores 5-HT3a y P2X4 en células tsA 201 ............... 30 FIGURA 5. Análisis de western blot de los receptores 5-HT3a y P2X4 ................................. 32 FIGURA 6. Imagen mediante AFM del complejo 5-HT3a-P2X4 ........................................... 34 FIGURA 7. Histograma de los volúmenes moleculares de los receptores 5-HT3a y P2X4. ....35 FIGURA 8. Histograma de los volúmenes moleculares del complejo 5-HT3a-P2X4...............36 FIGURA 9. Histograma de los volúmenes moleculares del complejo 5-HT3a-P2X4 a distintas concentraciones de ATP ......................................................................................................................37 FIGURA 10. Imagen mediante HS-AFM del complejo 5-HT3a-P2X4. .....................................39 FIGURA 11. Mediciones de calcio en células tsA 201 que expresan el complejo de receptores 5-HT3a-P2X4........................................................................................................................................41 FIGURA 12. Modelo planteado de Crosstalk estructural entre los receptores de membrana 5-HT3a y P2X4 .....................................................................................................................................48.
(9) ix. LISTA DE ABREVIATURAS. ATP: adenosina 5´-trifosfato AFM: microscopía de fuerza atómica CHAPS: 3-[(3-Colamidopropil)dimetilamonio]-1-propanesulfonato DMSO: dimetilsulfóxido DOPC: 1,2-dioleoil-sn-glicero-3-fosfocolina DOPS: 1,2-dioleoil-sn-glicero-3-fosfoserina DTT: ditiotreitol EDTA: ácido etilendiaminotetraacético FRET: transferencia de energía de resonancia de Förster GABA: ácido γ-aminobutírico HS-AFM: microscopía de fuerza atómica de alta velocidad nACh: receptor nicotínico de acetilcolina PEI: polietilenimina PPADS: ácido piridoxalfosfato-6-azofenil-2,4-disulfónico SNC: sistema nervioso central SNP: sistema nervioso periférico tsA 201: células humanas embrionales de riñón.
(10) 1. RESUMEN. Los receptores ionotrópicos 5-HT3 y P2X representan dos tipos diferentes de canales iónicos activados por ligando, tanto estructural como funcionalmente. Desde el punto de vista fisiológico, cabe señalar que ambos receptores se expresan en el sistema nervioso central y periférico, pero es en el sistema nervioso central donde la participación de ambos receptores es importante, ya que participan en una serie de patologías como enfermedades neurodegenerativas y dolor neuropático. Aunque se ha demostrado el crosstalk funcional entre receptores ionotrópicos; aún se desconoce cómo los receptores 5-HT3a pueden interactuar directamente con los receptores P2X, específicamente con P2X4 y si esto depende de la presencia de agonistas como ATP. Se transfectó las células tsA 201 con pDNA de los receptores 5-HT3a y P2X4, marcados con los tags Myc/His-6 y HA respectivamente.. La. presencia. del. complejo. 5-HT3a-P2X4. se. analizó. por. inmunofluorescencia y western blot. Se obtuvo imágenes topográficas de los complejos 5HT3a-P2X4 por microscopía de fuerza atómica (AFM) para determinar su estequiometría. Se observó una distribución de partículas polidispersa que sugiere dos poblaciones, una de ellas, la de menor volumen molecular sugiere la presencia de proteínas aisladas, en cambio la población de mayor volumen molecular sugiere la presencia de un complejo de proteínas con una estequiometría 1:1, cuya abundancia aumenta en presencia de ATP. Se determinó que la activación de P2X4 mediante ATP aumenta el nivel de calcio intracelular que es inhibido por el receptor 5-HT3a. Pre-incubación con PPADS, antagonista del receptor P2X4, inhibe el aumento de calcio del receptor P2X4 cuando está aislado o formando complejo..
(11) 2. Mediante AFM de alta velocidad (HS-AFM) se observó en tiempo real la interacción física, probablemente entre los receptores 5-HT3a y P2X4 insertos en liposomas. Estos hallazgos proporcionan información sobre la arquitectura molecular del complejo de receptores 5-HT3a-P2X4 y nuevos antecedentes biofísicos sobre la regulación del crosstalk estructural entre receptores ionotrópicos..
(12) 3. ABSTRACT. The 5-HT3 and P2X ionotropic receptors represent two different types of ligand activated ion channels, structurally and functionally. From the physiological point of view, it should be noted that both receptors are expressed in the central and peripheral nervous system, but it is in the central nervous system where the participation of both receptors is important, since they are involved in a series of pathologies such as neurodegenerative diseases and neuropathic pain. Although the functional crosstalk between ionotropic receptors has been demonstrated; it is still unknown how 5-HT3a receptors can interact directly with P2X receptors, specifically with P2X4 and whether this depends on the presence of agonists such as ATP. tsA 201 cells were transfected with pDNA of the 5-HT3a and P2X4 receptors, labeled with the Myc/His-6 and HA tags respectively. The presence of the 5-HT3a-P2X4 complex was analyzed by immunofluorescence and western blot. Topographic images of the 5-HT3a-P2X4 complexes were obtained by atomic force microscopy (AFM) to determine its stoichiometry. A distribution of polydisperse particles was observed that suggests two populations, one of them, the one with the lowest molecular volume suggests the presence of isolated proteins, while the population with the highest molecular volume suggests the presence of a protein complex with a 1: 1 stoichiometry, whose abundance increases in the presence of ATP. It was determined that activation of P2X4 by ATP increases the level of intracellular calcium that is inhibited by the 5-HT3a receptor. Pre-incubation with PPADS, P2X4 receptor antagonist, inhibits the increase in calcium from the P2X4 receptor when it is isolated or forming complex. Using high-speed AFM (HS-AFM), the physical interaction.
(13) 4. was observed in real time, probably between the 5-HT3a and P2X4 receptors inserted in liposomes. These findings provide information on the molecular architecture of the 5-HT3a-P2X4 receptor complex and new biophysical background on the regulation of structural crosstalk between ionotropic receptors..
(14) 5. INTRODUCCIÓN. La actividad neuronal es regulada por distintos neurotransmisores que actúan sobre diferentes tipos de receptores que podrían ser independientes funcional y físicamente. La rápida neurotransmisión es mediada por canales iónicos activados por ligando, que incluyen a los receptores 5-HT3 y a los receptores P2X. Los receptores 5-HT3 y P2X, son canales iónicos que permiten el rápido flujo de iones (sodio, potasio, calcio) a través de la membrana. El receptor 5-HT3 se abre en respuesta a serotonina, mientras que el receptor P2X lo hace en respuesta a su ligando ATP. Estos receptores forman parte de las tres familias de receptores ionotrópicos: receptores Cys-loop, los cuales incluyen al receptor nicotínico de acetilcolina (nACh), el receptor 5-HT3, el receptor GABAA y el receptor de glicina (Karlin, 2002); receptores de glutamato que comprenden el AMPA, NMDA y kainato (Mayer, 2006); y los receptores de la familia P2X (North, 2002). Estos receptores son oligómeros y están formados por múltiples tipos de subunidades. Al estar formados por diferentes combinaciones de subunidades tienen distintas propiedades funcionales, tales como conductancias de canal único, desensibilización, sensibilidad a agonistas y antagonistas, además de diferente ubicación intracelular (E. Boué-Grabot et al., 2003). Estos receptores se encuentran localizados en una diversidad de tejidos, en particular en el sistema nervioso central (SNC) (E. Boué-Grabot et al., 2003). Crosstalk entre proteínas de membrana Las interacciones receptor-receptor se han convertido en un área de interés que ha revelado un nuevo nivel de complejidad en la regulación de los receptores. Sin embargo, estudios.
(15) 6. sugieren que tales interacciones podrían ir más allá de los mecanismos clásicos funcionales de crosstalk y se extienden a la modulación del receptor, tráfico, expresión en la superficie y posicionamiento en la membrana plasmática (Jo et al., 2011) (Shrivastava, Triller, Sieghart & Sarto-Jackson, 2011). En un ambiente fisiológico, las células están continuamente respondiendo a más de un estímulo, donde el crosstalk entre distintos receptores de membrana y sus vías de transducción de señal generan un impacto en la función celular (Barrera, Morales, Torres, & Villalón, 2005). La transmisión sináptica es realizada a través de la liberación de uno o más neurotransmisores desde el terminal presináptico, resultando en la activación de diferentes tipos de receptores co-localizados en el mismo sitio postsináptico. Recientes investigaciones han demostrado la co-activación de distintos receptores postsinápticos por sus respectivos neurotransmisores induciendo crosstalk de sus propiedades funcionales (Jo et al., 2011) (Emerit et al., 2016) Crosstalk funcional entre receptores de membrana de la familia Cys-loop y P2X La co-transfección de los receptores GABAA y P2X2 muestra un agrupamiento de estos receptores en membrana plasmática de neuronas hipocampales y que los receptores P2X2 modifican el direccionamiento de los receptores GABAA en una vía que induce su translocación desde un compartimento vesicular interno hacia la superficie compartida por ambos receptores (Khakh & Egan, 2005). Por otro lado, la co-activación de los receptores P2X2 y GABAA expresados en ovocitos de Xenopus, generan un crosstalk inhibitorio dependiente de las subunidades del receptor GABAA. Este crosstalk funcional es.
(16) 7. independiente del potencial de membrana, cambios en la dirección de las corrientes y la concentración de calcio (É. Boué-Grabot, Toulmé, Émerit, & Garret, 2004). La activación de los receptores nicotínicos (α3β4) y P2X2 genera una respuesta no aditiva debido a la inhibición de ambos tipos de receptores. Además, una disminución de los niveles de expresión de ambos receptores resulta en respuestas aditivas, esto sugiere que cierta proximidad de los receptores es necesaria para la inhibición (Khakh, Zhou, Sydes, Galligan, & Lester, 2000). Otro estudio en neuronas de la submucosa caracteriza funcionalmente la interacción entre 5-HT3 y P2X; los hallazgos indican que la activación de estos receptores abre dos poblaciones de canales diferentes que se modulan el uno al otro negativamente cuando son activados de manera simultánea. (Barajas-López, Montaño, & Espinosa-Luna, 2002). Las corrientes de sodio inducidas por ATP y serotonina tienen diferentes cinéticas y son mediadas por la activación farmacológica de los distintos receptores. Similares interacciones se han descrito entre receptores nACh y P2X (Barajas-López, Espinosa-Luna, & Zhu, 1998). Una investigación del año 2011 mostró que el tráfico del receptor P2X4 juega una función importante en el control neuronal, así como en la excitabilidad sináptica. La evidencia fisiológica dice que el receptor P2X4 interactúa directamente con el receptor GABAA, estos dos receptores se encuentran co-expresados en neuronas del núcleo ventromedial del hipotálamo y los experimentos de electrofisiología revelan que existe una interacción negativa entre estos receptores en sistemas de expresión recombinante y en preparaciones de corte de cerebro. Por otro lado, el extremo C-terminal de P2X4 es clave en la interacción.
(17) 8. física con GABAA y las subunidades moleculares específicas de P2X son determinantes en las interacciones funcionales con otros canales activados por ligando (Jo et al., 2011). Crosstalk e interacción física entre los receptores de membrana 5-HT3a y P2X4 Un crosstalk inhibitorio similar entre los receptores P2X2 y 5-HT3a, se ha descrito a través de estudios de co-expresión en neuronas mientéricas y en sistemas heterólogos, donde la activación de P2X2 y 5-HT3a favorece una interacción funcional y corrientes no aditivas en neuronas mientéricas en presencia de ATP y serotonina (E. Boué-Grabot et al., 2003). Registros de electrofisiología en ovocitos de Xenopus así como en sistema heterólogo de células HEK293 muestran crosstalk funcional entre estos receptores, donde la interacción del loop intracelular de la subunidad 5-HT3a y el dominio C-terminal de P2X2 son importantes para la generación del crosstalk, ya que al eliminar el dominio C-terminal del receptor P2X2 y cuando minigenes insertados que codifican para los dominios citoplasmáticos de los receptores 5-HT3a y P2X2 son sobreexpresados, se interrumpe la interacción entre ambos receptores (E. Boué-Grabot et al., 2003). Cultivos de neuronas hipocampales de rata mediante inmunofluorescencia y análisis de colocalización muestran que existe interacción entre el receptor 5-HT3a y P2X2. Estos receptores se co-expresan en las mismas neuronas, pero en este modelo, el receptor 5-HT3a se expresa sólo en la superficie de axones y dendritas de células transfectadas en presencia del receptor P2X2 (Emerit et al., 2016), esto demuestra la función dominante del receptor P2X2 como una proteína específica para interactuar con el receptor 5-HT3a y entrega evidencia de una vía directa de interacción funcional receptor-receptor en el tráfico de proteínas y de la interacción del receptor 5-HT3a con otros receptores de la familia P2X,.
(18) 9. mediante mutagénesis de los receptores P2X3 y P2X4, el que presenta un aumento considerable de la expresión de este receptor en la superficie celular (Emerit et al., 2016). Investigaciones muestran la existencia de receptores heteroméricos P2X4/P2X7 (Antonio et al., 2011) y recientemente se ha observado interacción física y funcional entre estos receptores expresados en ovocitos de Xenopus mediante microscopía FRET (Schneider et al., 2017). Esta investigación sugiere que el receptor P2X4 y P2X7 interactúan no sólo como heterómeros, sino que también como homómeros. No obstante, en todos estos casos aún no se conoce la estequiometría de estos receptores ionotrópicos en los complejos y si la interacción está controlada por la unión del agonista. Los receptores 5-HT3a y P2X4 y su implicancia fisiológica Los antecedentes descritos sobre crosstalk hacen mención sobre los receptores 5-HT3a y P2X4 interactuando con otros receptores ionotrópicos, los cuales se encuentran ampliamente expresados en el SNC y sistema nervioso periférico (SNP) (Jackson & Yakel, 1995) (Suurväli, Boudinot, Kanellopoulos, & Rüütel Boudinot, 2017), pero es en el SNC donde distintas líneas de investigación han descrito que participan en una serie de patologías; lo que se desconoce aún es si la interacción entre ambos receptores podría modular alguna de estas patologías. Los receptores 5-HT3a participan en funciones que incluyen emesis, cognición y ansiedad. Se ubican en varias áreas del cerebro incluyendo la corteza, hipocampo y núcleo accumbens (Tecott, Maricq, & Julius, 1993). Estos receptores 5-HT3a también regulan la motilidad intestinal, secreción y peristalsis en el sistema nervioso entérico y están involucrados en la.
(19) 10. regulación de la información en el tracto gastrointestinal (Galligan, 2002). Los antagonistas del receptor de serotonina son usados en clínica para aliviar náuseas y vómitos causados por quimioterapia y radioterapia, y para el manejo de diarreas post infección y síndrome de colon irritable (Perwitasari et al., 2011). Los receptores P2X4 están involucrados en transmisión sináptica, contracción muscular, secreción exocrina, agregación plaquetaria, activación de macrófagos, diferenciación y proliferación celular, así como dolor neuropático, inflamación, función cardíaca, así como preferencia y consumo de alcohol (Suurväli et al., 2017) (Bortolato et al., 2013). Así también, una deficiencia de este receptor estaría asociada a discapacidad sociocomunicativa (Wyatt et al., 2013). El principal ligando del receptor P2X es ATP, esta molécula es reconocida como responsable de la transferencia de energía intracelular; sin embargo, el ATP puede actuar como una molécula de señalización extracelular, ya que se ha demostrado que ATP media la neurotransmisión en la mayoría de las células nerviosas del SNC y SNP (Suurväli et al., 2017). Por ejemplo, bajo condiciones fisiológicas, astrocitos y neuronas son responsables de la liberación de pequeñas cantidades de ATP, que puede actuar sobre los receptores purinérgicos P2X o P2Y, ambos expresados en neuronas y células gliales, regulando la señalización purinérgica. Sin embargo, cuando se libera ATP a partir de células dañadas, puede iniciar inflamación, además de amplificar y mantener la inmunidad celular a través de los receptores P2 (Stokes, Layhadi, Bibic, Dhuna, & Fountain, 2017). Entonces el aumento de ATP producto de daño celular podría afectar y modular la interacción de receptores P2X, como P2X4, con otros receptores co-expresados en el SNC..
(20) 11. Para poder investigar sobre las posibles interacciones físicas entre los receptores 5-HT3a y P2X4, la implicancia de esta interacción en patologías en el SNC, es necesario conocer la estructura de estos importantes canales iónicos activados por ligando y de esta manera determinar las bases de un posible crosstalk estructural. Estructura del receptor 5-HT3a Cada subunidad del receptor 5-HT3 tiene un gran dominio extracelular que forma el sitio de unión al ligando, un dominio transmembrana que consiste en cuatro α-hélices (M1-M4) que permiten a los iones cruzar la membrana y un dominio intracelular formado por un loop intracelular M3-M4, el cual es responsable de la modulación del receptor, clasificación y tráfico de moléculas e influye en la apertura y conductancia iónica (Boess, Beroukhim, & Martin, 1995). La estructura pentamérica del receptor 5-HT3 se observó por primera vez mediante microscopía electrónica (Boess et al., 1995), mientras que la estructura cristalizada del receptor se logró mediante rayos X difractados a una resolución de 3,5 Å. El modelo estructural comprende los residuos 9-275, 280-334 y 399-459 del receptor. Los iones extracelulares entran a un vestíbulo de 60 Å de alto y 20 Å de ancho, mientras que cuando los iones entran, atraviesan un poro transmembrana de ~ 40 Å (Hassaine et al., 2014). Dos tipos de subunidades del receptor 5-HT3; 5-HT3a y 5-HT3b tienen relevancia fisiológica (Thompson & Lummis, 2006). La subunidad a forma receptores homoméricos funcionales, mientras que la subunidad b no lo hace. El receptor pentamérico 5-HT3a tiene un peso molecular de ~ 250 kDa y está formado por cinco subunidades a homólogas o combinaciones heterólogas de la subunidad a con otras subunidades (b-e) arregladas alrededor de un seudo eje simétrico (Niesler, Frank, Kapeller,.
(21) 12. & Rappold, 2003). Estos canales se ubican en el complejo vagal dorsal del tronco cerebral y en el tracto gastrointestinal, forman un circuito que controla motilidad intestinal, secreción, percepción visceral y el reflejo de emesis (Engel, Smidt, & Hooft, 2013). Por otro lado, el receptor 5-HT3a participa en una serie trastornos psiquiátricos como ansiedad, depresión, trastorno bipolar y síndrome de colon irritable (Costall B. & Naylor RJ, 2004). La función principal del receptor homomérico 5-HT3a es participar en la rápida activación y desensibilización de corrientes de entrada, las cuales permiten el paso de iones sodio (Dubin et al., 1999). Los receptores también son permeables a calcio y otros pequeños cationes. (Maricq, Peterson, Brake, Myers, & Julius, 1991). El ciclo de apertura del canal involucra la transición entre tres estados conformacionales: estado inactivo, es una conformación no conductiva con baja afinidad al agonista; estado abierto, una conformación conductiva con alta afinidad al agonista y el estado desensibilizado, con una conformación no conductiva con la más alta afinidad al agonista entre los tres estados (Hilf & Dutzler, 2008). La unión de serotonina en el dominio extracelular induce un cambio conformacional que permite la apertura del poro en el dominio transmembrana. Además de estos dos dominios, el canal posee un gran dominio intracelular formado por la región entre las hélices M3 y M4 (Sauguet et al., 2014). El dominio intracelular está implicado en el tráfico y empaquetamiento del receptor en membrana plasmática, apertura del canal, modificación post traduccional y regulación de las funciones intracelulares del canal (Hilf & Dutzler, 2008). Este dominio intracelular es relativamente grande, modulando la conductancia de canal único, rectificación y desensibilización cinética (Peters et al., 2004)..
(22) 13. Estructura del receptor P2X4 Los diferentes subtipos de receptores P2X comparten la topología en forma de trímeros (Gouaux & Hattori, 2012). Estructuralmente, estos receptores están caracterizados por dos dominios transmembrana, un gran loop extracelular de unión a ligando y los extremos Cterminal y N-terminal ubicados en el citoplasma. El N-terminal contiene un sitio consenso para la fosforilación de la proteína quinasa C, mientras que el C-terminal es variable (Suurväli et al., 2017). La estructura cristalizada del receptor P2X4 ha sido resuelta en pez cebra (Kawate, Michel, Birdsong, & Gouaux, 2009) (Li, Kawate, Silberberg, & Swartz, 2010) y confirmada en rata (Igawa, Abe, Tsuda, Inoue, & Ueda, 2015). El receptor P2X4 en pez cebra presenta forma de cáliz y tiene una estructura trimérica con enlaces covalentes entre las subunidades que tienen una función importante en la apertura del canal, así como en el ensamblaje del receptor (Kawate et al., 2009). La estructura homomérica de pez cebra zfP2X4 tiene un dominio extracelular de alrededor de 70 Å desde la membrana y una pequeña región transmembrana que se extiende a través de la membrana de alrededor de 28 Å (Kawate et al., 2009). La parte extracelular de cada región del receptor puede ser descrita como una región rica en enlaces disulfuros, mientras que la región transmembrana del receptor está formada por seis hélices, dos de cada subunidad. Las hélices dentro de una subunidad están antiparalelas entre si formando un ángulo de 45° con la membrana (Kawate et al., 2009). Para la activación del receptor, tres moléculas de ATP se deben unir al dominio extracelular, induciendo la formación del canal catiónico en la región transmembrana (Gouaux & Hattori, 2012). El ATP es necesario para el cambio conformacional que permite la apertura del canal..
(23) 14. Los cationes al entrar a la célula inducen despolarización y activan numerosos procesos intracelulares (Wong et al., 2000). El comportamiento de los receptores P2X puede ser modificado por interacciones alostéricas. Estas interacciones ocurren en sitios adicionales al sitio activo y están vinculados a cambios conformacionales de la proteína (Christopoulos, May, Avlani, & Sexton, 2004). Entonces, la respuesta alostérica surge no sólo de la unión de pequeñas o grandes moléculas extracelulares, sino que también de cambios en temperatura, fuerza iónica y de modificaciones covalentes en la región transmembrana extracelular o intracelularmente (Tsai, del Sol, & Nussinov, 2009). En ausencia de calcio extracelular es posible observar un importante aumento de tamaño del poro transmembrana en el receptor P2X4 (Khakh, Proctor, Dunwiddie, Labarca, & Lester, 1999). Para entender la estructura del receptor, así como los cambios conformacionales que lo afectan al ser estimulado, mediante microscopía de fuerza atómica (AFM) se observó el comportamiento del receptor P2X4 a nivel de canal único. En presencia de calcio, el receptor P2X4 se abre sólo a cationes pequeños, permeables al poro del canal; por el contrario, en ausencia de calcio extracelular el tamaño del poro aumenta permitiendo el paso de moléculas como ioduro de propidio y bromuro de etidio (Shinozaki et al., 2009). Estos resultados demuestran que el dominio extracelular del receptor P2X4 está involucrado en la dinámica de la permeabilidad del canal. Está bien establecido que el receptor P2X4 es activado por ATP que en su totalidad está disociado en la forma ATP4-. Este hallazgo está de acuerdo con la estructura cristalizada del receptor P2X4-ATP, que muestra que los grupos fosfatos cargados negativamente α, β y γ.
(24) 15. interactúan con aminoácidos conservados polares y básicos de las dos subunidades de P2X4 (Gouaux & Hattori, 2012). Estudios topográficos mediante AFM como herramienta de Biología Estructural Si bien, la electrofisiología y farmacología han sido ampliamente usadas para describir las propiedades de los receptores ionotrópicos y proteínas de membrana en general (Gill, Peters, & Lambert, 1995); se han desarrollado continuamente una variedad de métodos para abordar la arquitectura molecular de proteínas de membrana que interactúan, desde una alta resolución como la cristalografía de rayos X hasta enfoques biofísicos complementarios que incluyen crio-microscopía electrónica (Cryo-EM), espectrometría de masas (MS) y AFM. Sobre la base de la exploración de la superficie de la muestra, AFM ya se ha utilizado para caracterizar la arquitectura molecular de los receptores individuales P2X2, P2X3, P2X4, P2X6 y 5-HT3a, 5-HT3ab (Barrera & Robinson, 2011); (Barrera, Herbert, Henderson, Martin, & Edwardson, 2005); (Barrera, Morales, et al., 2005); (Barrera, Ormond, Henderson, Murrell-Lagnado, & Edwardson, 2005); (Barrera, Henderson, Murrell-Lagnado, & Edwardson, 2007); (Barrera & Robinson, 2011); (Antonio et al., 2011), entre otros. Dentro de los métodos existentes para estudiar la topografía y propiedades físicas de las superficie de las membranas, la técnica de AFM ha sido desarrollada como una importante herramienta para medir fuerza y dinámicas de interacción entre moléculas individuales, (Shahin & Barrera, 2008) y por ende podría ser aplicada para estudiar la interacción funcional entre dos receptores ionotrópicos. Esta técnica produce imágenes tridimensionales de superficies biológicas con resolución atómica, pudiendo observar procesos biológicos y fisiológicos en tiempo real (Shahin & Barrera, 2008). AFM usa un pequeño tip, del orden de unos pocos nanómetros para escanear la superficie de la muestra la que es presionada con.
(25) 16. una pequeña fuerza de carga. El movimiento es controlado por un piezo eléctrico en la dimensión horizontal (x,y) y vertical (z). Cuando el tip entra en contacto con la muestra genera una deflexión del cantilever. La magnitud de ésta es capturada por un láser enfocado y centrado en la parte trasera de la superficie del cantilever, justo encima del tip que permite estimar la dirección angular de los cambios en la deflexión del tip. AFM funciona principalmente en el modo contacto y modo intermitente. El modo contacto se usa para análisis de muestras biológicas relativamente rígidas, como células o tejidos porque el continuo contacto entre el tip y la muestra puede producir daño. En cambio, en el modo intermitente el cantilever oscila a una frecuencia constante y la interacción entre el tip y la muestra se ve considerablemente reducida, siendo menos destructiva (Shahin & Barrera, 2008). Dentro de las ventajas de AFM en el área de microscopía, se pueden mencionar que 1) entrega un perfil de la superficie tridimensional en condiciones fisiológicas por lo tanto la muestra no requiere tratamientos especiales ni fijación, 2) obtiene información topográfica de moléculas biológicas en la superficie, donde la mayoría de las señales bioquímicas reguladoras y otras señales son dirigidas, 3) a nivel de la membrana plasmática, AFM permite determinar la fuerza y dinámicas de interacción en moléculas individuales y 4) puede ser aplicada para detectar cambios conformacionales, de estados oligoméricos y estequiometría de proteínas de membrana (Shahin & Barrera, 2008). Recientemente, otra ventaja de esta técnica ha revolucionado la investigación de procesos biológicos, AFM de alta velocidad (HS-AFM) (Rajendran, Endo, & Sugiyama, 2014) permite observar de manera directa procesos dinámicos de moléculas biológicas a una velocidad en el orden de 100 ms, con resolución lateral en nanómetros (Ando, 2013) (Eghiaian, Rico, Colom, Casuso,.
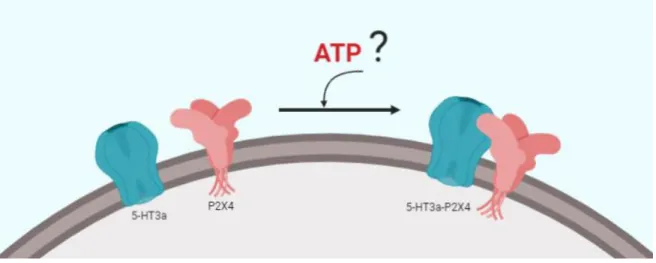
(26) 17. & Scheuring, 2014). Por ejemplo, HS-AFM ha sido utilizado para obtener imágenes de receptores P2X4 reconstituidos en bicapas, antes y después de adicionar ATP, para analizar los cambios en la estructura del poro del receptor (Shinozaki et al., 2009). Por lo tanto, AFM puede representar una buena herramienta para determinar la estequiometría y la movilidad del complejo de receptores 5-HT3a-P2X4 en ausencia y presencia de ATP. Los antecedentes presentados demuestran que existe co-localización entre receptores 5HT3a y P2X4; sin embargo, se desconoce si existe un crosstalk estructural estequiométrico en este complejo (FIGURA 1) y si su formación y función depende de la presencia del ligando ATP. Además, las imágenes de AFM podrían sugerir información sobre los cambios conformacionales que induciría ATP sobre el receptor P2X4, por lo tanto, en la formación de un complejo de receptores 5-HT3a-P2X4 en membrana.. FIGURA 1. Crosstalk estructural entre los receptores de membrana 5-HT3a y P2X4. En membrana plasmática los receptores 5-HT3a y P2X4 que se encuentran aislados pueden interactuar físicamente formando un complejo de receptores 5-HT3a-P2X4, aunque no se descarta que ATP al unirse al receptor P2X4 podría modular y favorecer la formación del complejo 5-HT3a-P2X4..
(27) 18. HIPÓTESIS. “Existe crosstalk estructural estequiométrico entre los receptores 5-HT3a y P2X4, donde el receptor 5-HT3a ejerce un efecto inhibitorio sobre la respuesta del receptor P2X4”. OBJETIVO GENERAL. Determinar el efecto de ATP en la estequiometría y función del complejo de receptores 5-HT3a-P2X4, a través de AFM y mediciones de calcio intracelular respectivamente.. OBJETIVOS ESPECÍFICOS. I. Evaluar la presencia del complejo de receptores 5-HT3a y P2X4 en la superficie de células tsA 201 mediante inmunofluorescencia y vía análisis de western blot del complejo purificado. II. Determinar el efecto de ATP en la abundancia del complejo de receptores 5-HT3a-P2X4, mediante imágenes topográficas de AFM.. III. Evaluar los cambios dinámicos del complejo de receptores 5-HT3a-P2X4 en la presencia de ATP mediante HS-AFM. IV. Determinar el efecto de ATP en la señalización de calcio intracelular mediada por el complejo de receptores 5-HT3a-P2X4..
(28) 19. MATERIALES Y MÉTODOS. Transformación de células competentes con pDNA del receptor 5-HT3a y P2X4 Se transformó bacterias E. coli DH5α competentes con DNA plasmidial (pDNA) purificado de 5-HT3a y P2X4. El cDNA que codifica el receptor 5-HT3a humano con un tag en Cterminal Myc/His-6 se clonó en el vector pcDNA3.1 (Invitrogen) usando HindIII/Xhol. El cDNA que codifica para el receptor de rata P2X4 con un tag en C-terminal hemaglutinina (HA) se clonó en el vector pEGFP sin el tag GFP (Invitrogen). Para la transformación se mezcló 10 µL de E. coli competente con 2 µL del plasmidio respectivo. Se incubó las células por 30 min en hielo, luego se sometió a un choque térmico por 45 seg a 42°C en un baño termorregulado y posteriormente se enfrió la muestra durante 2 min en hielo. Se incubó las células en 250 µL de medio LB (5 g triptona; 2,5 g extracto de levadura; 2,5 g NaCl) en una incubadora con agitación a 37ºC, 476 g durante 1 h. Luego, se transfirió el volumen en placas LB/ampicilina para 5-HT3a y placas LB/kanamicina para P2X4 y se incubó toda la noche (~12 h) en una incubadora a 37ºC para obtener colonias aisladas.. Purificación de pDNA del receptor 5-HT3a y P2X4 Se utilizó el kit QIAfilter™ Plasmid Maxi Kit (Cat. #122262, Hilder, DE) para la purificación. Se preparó un inóculo de bacterias picando una colonia y sometiéndola a agitación en una incubadora a 37°C, 476 g, en medio LB toda la noche (~12 h). El inóculo se transfirió a 250 mL de LB, y se incubó en agitación a 37°C, 476 g durante toda la noche (~12 h). Se cosechó las bacterias por centrifugación a 6.000 g por 15 min (centrifuga Sigma modelo 3-18K, rotor 19776-H). Se resuspendió el pellet bacteriano y se adicionó las.
(29) 20. soluciones correspondientes para obtener un lisado homogéneo. Se filtró el lisado y se incubó por 10 min a temperatura ambiente. Se filtró el lisado por un sistema de columnas, se lavó por flujo de gravedad y el eluído se precipitó adicionando isopropanol. La muestra se centrifugó a 15.000 g por 30 min a 4°C (centrifuga Sigma modelo 3-18K, rotor 12154-H) para obtener un pellet de pDNA, el cual se lavó con etanol 5%. El pellet se secó al aire, resuspendió en agua Milli-Q estéril y cuantificó (NanoPhotometer P-Class, IMPLEN, Munich, DE). Cultivo celular y transfección de células tsA 201 con pDNA del receptor 5-HT3a y P2X4 Se mantuvo las células tsA 201 (Sigma-Aldrish, St. Louis, MO. Cat. # 96121229-1VL) en 5 frascos para cultivo T175 con medio DMEM (Gibco, Grand Island, NY, USA), suplementado con 10% de suero fetal bovino (Gibco, Grand Island, NY, USA), 100 unidades/mL penicilina (Sigma-Aldrish, St. Louis, MO), 100 µg/mL estreptomicina (SigmaAldrish, St. Louis, MO) en estufa con 5% CO2 – 95% aire atmosférico y ajustada a 37°C hasta alcanzar un 60 - 70% de confluencia. Para transfectar las células con los pDNA de los receptores, se usó polietilenimina (PEI) (Sigma-Aldrish, St. Louis, MO) 1mg/mL. Se mezcló 10 mL de medio DMEM sin suero con 275 µL de PEI, se agregó 200 µL de los respectivos pDNA (25 µg cada uno) y se incubó durante 10 min a temperatura ambiente. Posteriormente, se agregó 2 mL de la mezcla en cada frasco para cultivo y se mantuvo por 24 h en estufa con 5% CO2 – 95% aire atmosférico y ajustada a 37°C. Al día siguiente de la transfección se realizó el proceso de purificación de proteínas..
(30) 21. Solubilización y purificación del receptor 5-HT3a conteniendo el tag Myc/His-6 Se descartó el medio de cultivo de los cinco frascos, y se lavó las células con 9 mL de solución salina HBS (composición (mM): HEPES 50; NaCl 100, pH 7,6) + EDTA 2 mM. Se removió las células por agitación usando 10 mL de solución salina HBS + EDTA 2 mM y se colocó en un tubo falcon de 50 mL. Se extrajo las células remanentes con 10 mL de la misma solución salina y se colocó en otro tubo falcon de 50 mL. Se obtuvo un pellet de células mediante centrifugación de ambos tubos falcon a 6.500 g por 5 min a 4°C (centrifuga Sigma modelo 3-18K, rotor 19776-H). Se descartó el sobrenadante y se resuspendió el pellet celular en solución de solubilización (solución de lisis 9 mL; Triton X-100 2 mL (SigmaAldrish, St. Louis, MO); 1 tableta cOmplete™, EDTA-free (Roche); PMSF 10 µL) (Solución de lisis (composición (mM): Tris 10; NaCl 100; EDTA 5, pH 7,6). La muestra se incubó durante 1 h en rotación en cámara fría. La muestra se centrifugó a 6.500 g por 5 min a 4°C (centrifuga Sigma modelo 3-18K, rotor 19776-H). El sobrenadante se colocó en un tubo para centrifugación Beckman y se sometió a ultracentrifugación (Ultracentrífuga Beckman Optima™ L80, rotor Ty80, USA) a 50.000 g por 1 h a 4°C. Se lavó las beads His (HisPure™ Ni-NTA superflow agarose. Cat. # 25215 (Thermo Scientific) dos veces con 8 mL de solución de lavado (HBS 9 mL; CHAPS 1%; 1 tableta cOmplete™, EDTA-free Roche). El sobrenadante de la centrifugación se agregó a las beads previamente lavadas y se incubó en cámara fría en rotación por 1 h. Se centrifugó a 6.500 g por 5 min a 4°C (centrifuga Sigma modelo 3-18K, rotor 19776-H), se descartó el sobrenadante y se lavó las beads tres veces con 8 mL de solución de lavado. Se resuspendió las beads en igual volumen de solución de lavado y se transfirió a una columna de purificación (Cat. # 450015, Invitrogen). Se decantó las beads, luego se dejó escurrir la.
(31) 22. solución de lavado. Se agregó 500 µL de solución de elución con imidazol; 200 mM imidazol para la elución 1 (E1) y 400 mM imidazol para la elución 2 (E2). Se colectó las fracciones de proteínas eluídas con ambas concentraciones de imidazol por separado en tubos eppendorf.. Solubilización y purificación del receptor P2X4 conteniendo el tag HA Para la purificación del receptor P2X4 se descartó el medio de cultivo de los cinco frascos y se lavó las células con 9 mL de solución salina HBS (composición (mM): HEPES 50; NaCl 100, pH 7,6) + EDTA 2 mM. Se removió las células por agitación usando 10 mL de solución salina HBS + EDTA 2 mM y se colocó en un tubo falcon de 50 mL. Se extrajo las células remanentes con 10 mL de la misma solución salina y se colocó en otro tubo falcon de 50 mL. Se obtuvo un pellet de células mediante centrifugación de ambos tubos falcon a 6.500 g por 5 min a 4°C (centrifuga Sigma modelo 3-18K, rotor 19776-H). Se descartó el sobrenadante y se resuspendió el pellet celular en solución de solubilización (solución de lisis: 9 mL; Triton X-100 2 mL (Sigma-Aldrish, St. Louis, MO); 1 tableta cOmplete™, EDTA-free (Roche); PMSF 10 µL) (Solución de lisis (composición (mM)): Tris 10; NaCl 100; EDTA 5, pH 7,6). La muestra se incubó durante 1 h en rotación en cámara fría. La muestra se centrifugó a 6.500 g por 5 min a 4°C (centrifuga Sigma modelo 3-18K, rotor 19776-H) El sobrenadante se colocó en un tubo para centrifugación Beckman y se sometió a ultracentrifugación (Ultracentrífuga Beckman Optima™ L80, rotor Ty80) a 50.000 g por 1 h a 4°C. Se lavó las beads HA (HA epitope tag agarose conjugate. Cat. # 26181. Thermo Scientific) con solución de lavado (solución de lisis 9 mL; Triton 1 mL) colocando en un tubo eppendorf.
(32) 23. 1 mL de solución de lavado y 50 µL de beads HA, luego se centrifugó a 22.000 g por 1 min a 4°C (centrifuga Sigma modelo 3-18K, rotor 12154-H), este procedimiento se realizó tres veces. Luego de la centrifugación, el sobrenadante se colocó en un tubo falcon limpio y se colectó 100 µL en un tubo eppendorf para usarlo como control de membrana en western blot. Se colocó las beads HA en el tubo falcon junto con el sobrenadante de la ultracentrifugación y se incubó en cámara fría en rotación durante 3 h. Se centrifugó a 6.500 g por 5 min (centrifuga Sigma modelo 3-18K, rotor 19776-H). Se descartó el sobrenadante y se transfirió las beads HA con 1 mL de CHAPS 1% en un tubo eppendorf y se centrifugó a 22.000 g por 1 min a 4°C (centrifuga Sigma modelo 3-18K, rotor 12154-H). Se descartó el sobrenadante se agregó nuevamente 1 mL de CHAPS 1% y se incubó en cámara fría en rotación durante 5 min. Se centrifugó el tubo eppendorf a 22.000 g por 1 min a 4°C. Se descartó el sobrenadante y el pellet se transfirió a un nuevo tubo eppendorf, se agregó 1 mL de CHAPS 1%. Se incubó nuevamente en cámara fría en rotación durante 5 min. Se centrifugó el tubo eppendorf a 22.000 g por 1 min a 4°C. Se descartó el sobrenadante y se agregó 200 µL de CHAPS 1% y 6 µL del tag HA (tag HA peptide. Cat. #26184. Thermo Scientific). Se incubó en cámara fría en rotación durante 1 hora. Se centrifugó el tubo eppendorf a 22.000 g por 1 min a 4°C. El sobrenadante obtenido corresponde a la elución que contiene al receptor P2X4 unido al tag HA.. Solubilización y purificación del complejo de receptores 5-HT3a-P2X4 conteniendo el tag HA Se realizó el mismo protocolo para purificar el receptor P2X4 y el complejo 5-HT3a-P2X4, a través de pull down por HA, puesto que el receptor P2X4 contiene el tag HA. Se usó.
(33) 24. células tsA 201 co-transfectadas con 25 µg de cada plasmidio. La purificación del complejo de receptores 5-HT3a-P2X4 se realizó 6 veces.. Análisis de inmunofluorescencia del receptor 5-HT3a y P2X4 Las células transfectadas con los receptores 5-HT3a y P2X4 crecieron en covers de vidrio estériles y se fijó con formaldehído al 1%. Se lavó los covers con solución salina PBS (composición (mM): NaCl 136,9; KCl 2,68; NaH2PO4 10,44; KH2PO4 1,76), pH 7,4 y se bloqueó con PBS + 0,1% suero fetal bovino (Gibco, Grand Island, NY, USA). Se incubó las células toda la noche (~12 h) con anticuerpo monoclonal anti-Myc para el receptor 5-HT3a y anti-HA para el receptor P2X4, ambos anticuerpos en la dilución 1:250. Se lavó las células tres veces por 10 minutos con solución salina PBS e incubó por una hora con anticuerpo secundario, Alexa Fluor®555 para anti-Myc y Alexa Fluor®488 para anti-HA, ambos anticuerpos en la dilución 1:500. Como control negativo, se incubó las células sólo con anticuerpo secundario. El anticuerpo anti-Myc se obtuvo de Thermo Scientific (Cat. # PA1981, Rockford, IL, USA); el anticuerpo anti-HA se obtuvo de Thermo Scientific (Cat. # 26183, Rockford, IL, USA) y los anticuerpos secundarios Alexa Fluor®488 (Cat. # A11029) y Alexa Fluor®555 (Cat. # A-21424) se compraron en Molecular Probes, Eugene, OR, USA. La señal fluorescente se observó con un microscopio invertido IX81 (Olympus, Tokio, Japón) y una cámara digital CCD Orca R2 (Hamamatsu, Japón).. Análisis de western blot del receptor 5-HT3a y P2X4 La muestra de proteínas purificadas producto de la transfección y co-transfección de 5-HT3a y P2X4 se mezcló con buffer de carga 2X (Glicerol 4 mL; SDS10% 8 mL; Tris HCl 1 M pH.
(34) 25. 6,8 1mL; agua destilada 5,8 mL; DTT 1M 1,2 mL; β-mercaptoetanol 2 mL y azul bromofenol 2 mg), se hirvió 5 min para desnaturalizar las proteínas que posteriormente se separó por electroforesis en gel de poliacrilamida con dodecil sulfato sódico (SDS-PAGE 10%) y se transfirió a una membrana de nitrocelulosa (Cat. # 9004-70, BioRad, CA, USA). Se incubó los anticuerpos primarios anti-Myc; anti-His6 y anti-HA (1:500) en una solución de leche descremada al 4% toda la noche (~12 h) en cámara fría y el anticuerpo secundario anti-HRP (1:1.000) (Cat. # 621040, Molecular Probes, Eugene, OR, USA) se incubó en solución de leche al 4% durante 1 h a temperatura ambiente. Para detectar las bandas de las proteínas se usó el sustrato quimioluminiscente SuperSignal® West Femto, Thermo Scientific (Cat. # 34095). La masa molecular se estimó con marcadores estándar (Cat. # 1610375, BioRad, CA, USA). Para revelar la membrana se usó el equipo MyECL™ Imager, Thermo Scientific (Cat. # 62267). Como control positivo, se usó la fracción de membrana total enriquecida que corresponde al sobrenadante que se obtuvo producto de la solubilización y posterior ultracentrifugación del proceso de purificación; así como las beads específicas para los tags de los distintos receptores.. Formación de liposomas e inserción de los complejos de receptores 5-HT3a-P2X4 Se obtuvo 1,2-dioleoil-sn-glicero-3-fosfocolina (DOPC) y 1,2-dioleoil-sn-glicero-3fosfoserina (DOPS), a partir de Avanti Polar Lipids, como solución de cloroformo. Se mezcló 30 µL de cada uno de estos lípidos de acuerdo sus radios molares en un tubo de vidrio. Se incubó la mezcla durante 30 min a temperatura ambiente y se secó con nitrógeno gaseoso. Se lavó los lípidos secos con 370 µL de buffer HBS 1X a una concentración total de 2 mg/mL y se agitó el tubo en un vórtex hasta que la suspensión lipídica se observó.
(35) 26. transparente. Mediante diálisis, se incubó la suspensión lipídica en buffer HBS 1X con los complejos purificados de receptores 5-HT3a y P2X4 en cámara fría durante dos días, usando membranas de diálisis SnakeSkin® Dialysis Tubing, Thermo Scientific, (Cat. # 68100) previamente hidratadas en agua destilada. Se montó las muestras de liposomas-proteínas sobre micas, y se llevaron al HS-AFM para la obtención de imágenes (FIGURA 2). Este ensayo se realizó 2 veces.. FIGURA 2. Esquema representativo de una muestra de receptores 5-HT3a y P2X4 insertos en liposoma y analizados mediante HS-AFM.. Imágenes de los complejos de receptores 5-HT3a-P2X4 por AFM Una vez purificados los complejos 5-HT3a-P2X4, 50 µL de la muestra se colocó sobre micas de muscovita (Electron Microscopy Sciences, Cat. # 71855-01, Hatfield, PA, USA) para ser adsorbidas. Después de una incubación por 10 minutos, la muestra se lavó con agua MilliQ, se secó con nitrógeno gaseoso para terminar el montaje. Este procedimiento también se realizó cuando la muestra de los complejos 5-HT3a-P2X4 se trató 30 min con ATP 1, 10 y 100 µM. y con antagonista de receptores P2X, ácido disulfónico piridoxalfosfato-6azofenol-2',4' (PPADS) 10 µM. Se obtuvo imágenes de las muestras en seco, usando el modo de ‘contacto intermitente’ en un AFM MFP-3D-SA Asylum (FIGURA 3). El cantilever se usó con una frecuencia de ~ 500 kHz y una constante elástica 40 N/m. La fuerza aplicada.
(36) 27. para obtener las imágenes se mantuvo lo más baja posible, target amplitude ~ 0,5 V y amplitude set point ~0,4 V). Se obtuvo una imagen completa en 15 minutos. El volumen molecular de las partículas de receptores se determinó desde las dimensiones obtenidas por AFM. La altura y radio de la partícula se calculó a partir de la ecuación: Vm= (πh/6)(3r2 + h2) Ecuación 1. donde h es la altura de la partícula y r es el radio a la mitad de la altura. La ecuación considera a la molécula de proteína como una esfera. Se calculó la concentración de un receptor individual o del complejo de receptores a partir de la siguiente ecuación:. Ecuación 2. donde M representa la molaridad de la muestra de partículas purificadas obtenidas de imágenes de AFM. Para los experimentos de HS-AFM en líquido, usando un instrumento Bruker AXS FastScan™, se usó cantilevers AC40 (Olympus Corporation) con una frecuencia entre ~ 600 y 1.000 kHz y una constante elástica de 0,09 N/m. Se obtuvo una imagen por minuto..
(37) 28. FIGURA 3. Esquema representativo de una muestra de receptores 5-HT3a y P2X4 en una micela de detergente (CHAPS) analizados mediante AFM.. Medición de la concentración intracelular de calcio Se detectó los cambios en la concentración de calcio intracelular usando el indicador fluorescente de calcio Fluo 4-AM (Thermo Fisher Scientific), como se describió recientemente por Lillo et al. 2018 (Lillo et al., 2018). Se incubó las células tsA 201 con 5 µM de Fluo 4-AM por 45 min a temperatura ambiente para cargar las células con este marcador, que se disolvió en DMSO al 0,17%, preparado en un buffer salino Tyrode MOPS (composición mM: 118 NaCl; 5.4 KCl; 2,5 CaCl2; 1,2 KH2PO4; 1,2 MgSO4; 11,1 glucosa y 5 MOPS, pH 7,4). Se evaluó los cambios en la concentración de calcio intracelular en preparaciones de células con buffer MOPS después de incubarlas 5 min con solución buffer sin calcio, que contiene 2 mM EGTA. La señal fluorescente se observó con un microscopio BX50WI (Olympus, Tokio, Japón) y una cámara acoplada (Retiga Fast 1394; QImaging, Surrey, BC, Canadá). Se obtuvo imágenes de las células cargadas cada 2 s. Se expresó los cambios en la concentración de calcio intracelular como variaciones de la intensidad de fluorescencia F/F0, donde F es la fluorescencia observada durante el periodo de estimulación y F0 es el valor de la fluorescencia basal. En este experimento se midió los cambios de calcio intracelular 45 min después del tratamiento con Fluo 4-AM 5 µM..
(38) 29. RESULTADOS. Co-localización de los receptores 5-HT3a y P2X4 expresados en células tsA 201 mediante inmunofluorescencia En un experimento inicial, la expresión de proteínas y localización intracelular se analizó mediante inmunofluorescencia. Las células tsA 201 se transfectaron con el pDNA del receptor 5-HT3a y con el pDNA del receptor P2X4. En el panel A de la FIGURA 4 se observó en rojo la marca contra el tag Myc-His-6 para el receptor 5-HT3a usando el anticuerpo anti-Myc. En B, se observó en verde la marca contra el tag HA para el receptor P2X4 usando el anticuerpo anti-HA. En la esquina inferior derecha se muestran los respectivos controles negativos que corresponden a las células incubadas sólo con anticuerpo secundario. En el panel C, se observó la co-localización de la marca para el receptor 5-HT3a y P2X4. Esto sugiere que el protocolo de co-transfección en esta línea celular es eficiente, que ambas proteínas son detectadas y es posible realizar el protocolo de purificación de proteínas de membrana. Por otro lado, el tipo de marca en forma de anillo característica de proteínas de membrana indicaría co-localización en la misma célula y en membrana plasmática e intracelular..
(39) 30. FIGURA 4. Co-localización de los receptores 5-HT3a y P2X4 en células tsA 201. En el panel A, se observó marca en rojo para el receptor 5-HT3a con el anticuerpo primario antiMyc (1:250) y anticuerpo secundario Alexa Fluor®555 (1:500). En el panel B, se observó marca en verde para el receptor P2X4 con el anticuerpo anti-HA (1:250) y Alexa Fluor®488 (1:500). En cada uno de estos paneles se muestra el control negativo que corresponde a las células incubadas sólo con el anticuerpo secundario. El panel C, muestra la respectiva colocalización de la marca para el receptor 5-HT3a y el receptor P2X4. Barra de escala representa 50 µm. (n=4).
(40) 31. Detección de los receptores 5-HT3a y P2X4 mediante análisis de western blot La membrana de las células tsA 201 transfectadas con el pDNA del receptor 5-HT3a, se solubilizó y posteriormente se aisló los receptores mediante su unión a columnas Ni2+agarosa vía los tags de Histidina. En la FIGURA 5A, se observó una banda leve a 55 kDa en la elución 1 y elución 2 obtenidas a partir de concentraciones crecientes de imidazol y en el control positivo, correspondiente a la fracción de membrana total, usando el anticuerpo anti-Myc contra el tag Myc/His-6 del receptor. En la FIGURA 5B, se observó bandas aproximadamente a 70 kDa para el monómero del receptor P2X4 en la elución de la purificación, en la fracción de membrana total y en las beads usadas en el proceso de purificación, usando el anticuerpo anti-HA contra el tag HA. Estos resultados sugieren que el receptor 5-HT3a y el receptor P2X4 están presentes en las muestras purificadas. Se evaluó la formación del complejo 5-HT3a-P2X4 de las muestras purificadas a partir de células co-transfectadas. En la FIGURA 5C, se observó una banda en la fracción de membrana total y una leve banda en la elución a 55 kDa, con el anticuerpo anti-Myc contra el tag Myc/His-6 del receptor 5-HT3a; mientras que en la FIGURA 5D, se usó el anticuerpo anti-HA contra el tag HA del receptor P2X4, donde se observó una clara banda tanto en la fracción de membrana, así como en la elución y en las beads a 70 kDa. Este resultado sugiere que el receptor P2X4 forma parte del complejo y que por el ensayo de pull-down como técnica de purificación de proteínas, éste debe estar unido al receptor 5-HT3a..
(41) 32. FIGURA 5. Análisis de western blot de los receptores 5-HT3a y P2X4 de muestras purificadas. Se analizó las muestras purificadas usando anticuerpos monoclonales antiMyc (1:500) contra el tag Myc/His-6 del receptor 5-HT3a (panel A) donde se observa una banda a 55 KDa y el anticuerpo anti-HA (1:500) contra el tag HA del receptor P2X4 (panel B) donde se observa una banda a 70 kDa. Para evaluar la formación de un complejo de proteínas entre 5-HT3a y P2X4 mediante pull down, se usó el anticuerpo monoclonal antiMyc (1:500) y se observó una banda en la fracción de membrana total y una banda muy leve en la elución a 55 kDa correspondiente al receptor 5-HT3a (panel C), mientras que el receptor P2X4, se evaluó con el anticuerpo monoclonal anti-HA (1:500) y se observó bandas en la fracción de membrana total y en la elución a 70 kDa (panel D). Se usó el anticuerpo secundario anti-HRP conjugado (1:1.000). Los controles positivos corresponden a la fracción de membrana total y las beads que se usaron en el proceso de purificación de proteínas de membrana. Las cabezas de flecha indican los marcadores de peso molecular en kDa. (n=5).
(42) 33. Análisis topográfico del complejo 5-HT3a-P2X4 mediante AFM. Para tener información estructural y analizar la estequiometría de la interacción, se registró los complejos formados por los receptores 5-HT3a-P2X4 mediante imágenes de AFM en aire. Estas imágenes de baja magnificación mostraron partículas de distinto tamaño, incluyendo subunidades individuales, así como subunidades ensambladas formando complejo de receptores. En la FIGURA 6A, la imagen mostró una población de partículas que difieren en tamaño, lo que representaría tanto receptores aislados como formando complejos. La flecha roja indica una partícula con un volumen molecular de 493 nm3, mientras que la flecha azul indica una partícula de mayor tamaño con un volumen molecular de 1.194 nm3. La FIGURA 6B mostró una representación tridimensional del panel A. La FIGURA 6C, mostró una población de partículas formando el complejo 5-HT3a-P2X4 en presencia de ATP 100 µM, agonista del receptor P2X4. En esta imagen la población de partículas difiere en tamaño, la flecha roja indica una partícula con volumen molecular de 486 nm3, mientras que la flecha azul indica una partícula con volumen molecular de 1.124 nm3. La FIGURA 6D mostró una representación tridimensional del panel C. Dentro de la población de partículas, las imágenes mostraron de manera representativa el tamaño y forma de las partículas que se consideran en los análisis de histogramas de volumen molecular..
(43) 34. FIGURA 6. Imagen mediante AFM del complejo 5-HT3a-P2X4. En el panel A, la imagen de AFM mostró partículas que corresponderían a receptores P2X4 y complejos 5-HT3aP2X4. En el panel C, la imagen de AFM mostró una población de partículas que corresponderían a receptores P2X4 y complejos 5-HT3a-P2X4 en presencia de ATP 100 µM. Los paneles B y D muestran una representación tridimensional de la zona indicada en A y B respectivamente.. Distribución de frecuencias de los volúmenes moleculares del complejo 5-HT3a-P2X4. Las dimensiones de las partículas y la Ecuación 1 se usaron para calcular los volúmenes moleculares. La FIGURA 7A mostró el volumen molecular para una población abundante del receptor 5-HT3a con en un peak a 787 ± 27 nm3, mientras que para el receptor P2X4, FIGURA 7B, el volumen molecular se observó en un peak a 491 ± 5 nm3. Estos resultados son similares a los volúmenes publicados para ambos receptores vía AFM, 757 nm3 para el receptor 5-HT3a (Barrera, Herbert, et al., 2005) y 417 nm3 para el receptor P2X4 (Antonio et al., 2011)..
(44) 35. FIGURA 7. Histogramas de los volúmenes moleculares de los receptores 5-HT3a y P2X4. La curva indica la función Gaussiana utilizada para ajustar los datos. La muestra obtenida de la transfección del receptor 5-HT3a se observa en el panel A, donde los valores de los promedios corresponden a 787 ± 27 nm3. La muestra obtenida de la transfección del receptor P2X4 se observa en el panel B, donde los valores de los promedios corresponden a 491 ± 5 nm3. (promedio ± DE) (n= 200 partículas) (P < 0,0001). La FIGURA 8A mostró los volúmenes moleculares de las muestras purificadas obtenidas de la co-transfección de ambos receptores en dos peaks; el primero a 489 ± 25 nm3, lo cual puede corresponder a una población abundante de P2X4 libre y el segundo peak a 1.100 ± 109 nm3, que sugiere una población del complejo 5-HT3a-P2X4 con una estequiometria de 1:1. En la FIGURA 7, si sumamos los dos peaks correspondientes a cada uno de los receptores, tendríamos un volumen molecular de 1.278 nm3, el que es cercano los 1.100 nm3 obtenidos a partir de la co-transfección. Cuando la misma muestra de la co-transfección se trató con el agonista del receptor P2X4, ATP, se observó en la FIGURA 8B, que la población del complejo aumentó de 42% a 51% (área bajo el ajuste de doble curva gaussiana, corte de 750 nm3) después de la estimulación.
(45) 36. con ATP 100 µM, sin cambios en los valores máximos, 505 ± 15 nm3 y 1.236 ± 64 nm3 respectivamente.. FIGURA 8. Histogramas de los volúmenes moleculares del complejo 5-HT3a-P2X4. La curva indica la función Gaussiana utilizada para ajustar los datos. La muestra producto de la co-transfeción se observa en el panel A, donde los valores de los promedios corresponden a 489 ± 25 nm3, 1.100 ± 109 nm3. La muestra producto de la co-transfección se trató con ATP 100 µM como se observa en el panel B, donde los valores de los promedios corresponden a 505 ± 15 nm3, 1.236 ± 64 nm3 (promedio ± DE) (n= 250 partículas) (P < 0,0001). Para determinar la concentración adecuada de ATP en la evaluación del complejo, se realizó una curva dosis-respuesta mediante AFM determinando los volúmenes moleculares de las muestras purificadas. Se trató con 1, 10 y 100 µM de ATP. En la FIGURA 9A y FIGURA 9C, se observó una curva gaussiana con dos peaks, con una tendencia similar a la observada sin presencia de ATP. En cambio, en la FIGURA 9B, se observó dos peaks donde predomina uno a 875 ± 109 nm3. Considerando los datos obtenidos, los experimentos se realizaron con 100 µM ATP para asegurar ver el efecto esperado en este modelo de muestras purificadas y adsorbidas en mica..
(46) 37. FIGURA 9. Histogramas de los volúmenes moleculares del complejo 5-HT3a-P2X4 a distintas concentraciones de ATP. Se realizó una curva dosis-respuesta para ATP. Las curvas indican la función Gaussiana utilizada para ajustar los datos. En el panel A, la concentración de ATP es de 1 µM. Los valores de los peaks corresponden a 567 ± 36 nm3 y 1.205 ± 27 nm3. En el panel B, la concentración de ATP es de 10 µM. Los valores de los peaks corresponden a 875 ± 109 nm3 y 1.298 ± 48 nm3. En el panel C, la concentración de ATP es de 100 µM. Los valores de los peaks corresponden a 448 ± 39 nm3, 1.100 ± 106 nm3. (promedio ± DE) (n=50 partículas) (P < 0,0001).
(47) 38. Cambios dinámicos del complejo de receptores 5-HT3a-P2X4 en presencia de ATP mediante HS-AFM La mezcla de liposomas-proteínas se analizó mediante HS-AFM, esto permite obtener imágenes en tiempo real de las interacciones y cambios dinámicos entre proteínas. En la FIGURA 10A-B, se observó la interacción entre dos receptores. Considerando la altura del dominio extracelular de las partículas, 70 Å para el receptor P2X4 (Kawate et al., 2009) y 60 Å para el receptor 5-HT3a (Hassaine et al., 2014), podemos estimar que ambos receptores corresponderían al complejo de receptores 5-HT3a-P2X4, puesto que la imagen muestra diferencia en 1 nm entre cada partícula, lo que es proporcional a la diferencia de 10 Å de los datos publicados; ahora si bien podemos observar que las alturas obtenidas no son similares a las publicadas, esto puede deberse a una mayor interacción que ejercería el tip sobre la muestra produciendo una reducción en la altura de las partículas. Para evaluar si ATP genera cambios dinámicos en la conformación estructural de las proteínas insertas en la muestra de liposomas-proteínas, se obtuvo imágenes, luego de 5 min de tratamiento de la muestra con ATP 100 µM. La imagen de la FIGURA 10C-D no mostró cambios conformacionales en la altura de las partículas ni en la disposición espacial en presencia de ATP..
(48) 39. FIGURA 10. Imagen mediante HS-AFM del complejo de receptores 5-HT3a-P2X4. Las imágenes A y B de HS-AFM mostraron dos partículas entre 3 y 4,5 nm de altura, que podrían corresponder al complejo de receptores 5-HT3a-P2X4. La imagen C mostró dos partículas de 4 nm y 5 nm antes de ser tratadas con ATP, que podrían corresponder al complejo de receptores 5-HT3a-P2X4. En la imagen D se observó que no hubo cambios conformacionales evidentes al tratar la muestra (imagen C) con 100 µM de ATP. Las imágenes se registraron cada un minuto. Barra de escala representa 8 nm. (n=2).
Figure



+7






Documento similar
La campaña ha consistido en la revisión del etiquetado e instrucciones de uso de todos los ter- mómetros digitales comunicados, así como de la documentación técnica adicional de
Debido al riesgo de producir malformaciones congénitas graves, en la Unión Europea se han establecido una serie de requisitos para su prescripción y dispensación con un Plan
Como medida de precaución, puesto que talidomida se encuentra en el semen, todos los pacientes varones deben usar preservativos durante el tratamiento, durante la interrupción
El nuevo Decreto reforzaba el poder militar al asumir el Comandante General del Reino Tserclaes de Tilly todos los poderes –militar, político, económico y gubernativo–; ampliaba
En estos últimos años, he tenido el privilegio, durante varias prolongadas visitas al extranjero, de hacer investigaciones sobre el teatro, y muchas veces he tenido la ocasión
que hasta que llegue el tiempo en que su regia planta ; | pise el hispano suelo... que hasta que el
En junio de 1980, el Departamento de Literatura Española de la Universi- dad de Sevilla, tras consultar con diversos estudiosos del poeta, decidió propo- ner al Claustro de la
E Clamades andaua sienpre sobre el caua- 11o de madera, y en poco tienpo fue tan lexos, que el no sabia en donde estaña; pero el tomo muy gran esfuergo en si, y pensó yendo assi
