Actividad antifúngica de metabolitos bacterianos frente a Mycosphaerella fijiensis Morelet
97
0
0
Texto completo
(2) TESIS DE DIPLOMA. Actividad antifúngica de metabolitos bacterianos frente a Mycosphaerella fijiensis Morelet. Autora: Eilyn Mena Méndez Tutora: Dr.C. Yelenys Alvarado Capó Consultante: MSc. Mileidy Cruz Martín. Santa Clara, 2012.
(3) Cuando quieras emprender algo, va haber un montón de gente que dirá que no lo hagas. Cuando vean que no te pueden detener, te dirán como lo tienes que hacer. Y cuando vean que finalmente lo has logrado, dirán que siempre han creído en ti. John C. Maxwell.
(4) . A mi tutora Yelenys Alvarado, por enseñarme a ver las cosas desde otro punto de vista. . A Mileidy Cruz, por su amistad y ayuda constante. . A los compañeros del laboratorio: Cynthia, Michel, Mayra, Berkis y Yoandy. . Al resto de los trabajadores del IBP que me brindaron su ayuda y apoyo. . A mi novio Dany, por su paciencia y apoyo incondicional. . A mi hermana (Evelyn), tía (Sabina), abuela (María) y padres (Idaimi y Eduardo), por su colaboración. . A los padres de mi novio (Cristina y Abel), por sus consejos. . A todos los que han hecho posible la realización de esta investigación,. Muchas gracias..
(5) Dedico este trabajo:. A mi familia y a mi novio, en especial a mi abuela María (Mima).
(6) Resumen RESUMEN Las bacterias asociadas a cultivos han sido empleadas para incrementar la nutrición de la planta y como fuentes de agentes de biocontrol, pero pocas son las investigaciones relacionadas con la interacción Musa spp.-microorganismo-microorganismo. Por ello, este trabajo tuvo como objetivo caracterizar la actividad antifúngica de metabolitos producidos por bacterias frente a Mycosphaerella fijiensis. Con este fin se obtuvieron filtrados de cultivo a partir de bacterias que inhibieron in vitro el crecimiento del patógeno y se evaluó su efecto sobre la respuesta de plantas de Musa sp. inoculadas con este hongo. Así como también, se caracterizó la actividad antifúngica de estos con respecto a la estabilidad ante variaciones de la temperatura, el pH, susceptibilidad a la desnaturalización proteica por detergentes y proteinasa K. Además, se semipurificó la fracción proteica mediante la precipitación con sulfato de amonio (70%) y diálisis con membrana de 12kDa de exclusión. Los filtrados de cultivo de las cepas, mostraron actividad antifúngica in vitro a partir de las 48 horas de incubación y tuvieron efecto sobre la respuesta de plantas inoculadas con M. fijiensis en casa de cultivo, en dependencia de la cepa bacteriana empleada y del momento de su aplicación. La actividad antifúngica se vio afectada por la temperatura, el pH, así como, en presencia de detergentes CTAB, Tritón x-100 y Tween-80 y la enzima proteinasa K en dependencia de la cepa. Estos resultados sugirieron la posible naturaleza proteica de los metabolitos con actividad antifúngica. Se comprobó que la actividad antifúngica de los filtrados de cultivo frente a Mycosphaerella fijiensis estuvo relacionada con metabolitos de naturaleza proteica presentes en su composición. Palabras claves: actividad antagonista, metabolitos antifúngicos, metabolitos secundarios, filtrados de cultivo, proteínas, bacterias epífitas..
(7) Abstract ABSTRACT Bacteria associated to crop have been employed to increase the nutrition of plants and as sources of biocontrol agent, but a few investigations are related the interaction Musa spp.microorganism- microorganism. For that reason, the aim of this work is to characterize the antifungal activity of metabolites produced by bacteria against Mycosphaerella fijiensis. With this objective were obtained cell-free culture filtrate from bacteria who inhibited the pathogen growth in vitro and was evaluated their effect on the Musa spp. plants response inoculated with this fungus. As well as, were characterized the antifungal activity of these with respect to thermal and pH stability, susceptibility to protein denaturing detergents and proteinase K. In addition, was half-purified protein fraction by means of ammonium sulphate precipitation to achieve 70% saturation and dialysis with membrane of 12kDa shutting out. Cell-free cultures filtrate from bacteria, showed antifungal activity in vitro from 48 hours of incubation. and. had. effect. on. the. Musa. spp.. plants. response. inoculated with this fungus in greenhouse, dependence of the strain bacterium employed and the moment of its application. The antifungal activity look affected by temperature, pH, as well as, in the presence of detergents CTAB, Triton x-100 and Tween-80 and enzyme proteinase K in dependency of the strain. These results suggested the possible protein nature of the metabolites with antifungal activity. In this study, was verified that the antifungal activity of cell-free cultures filtrate against Mycosphaerella fijiensis was related with metabolites of protein nature in its composition. Keywords: antagonistic activity, antifungal metabolites, secondary metabolites, cell-free cultures filtrate, proteins, endophytic bacteria..
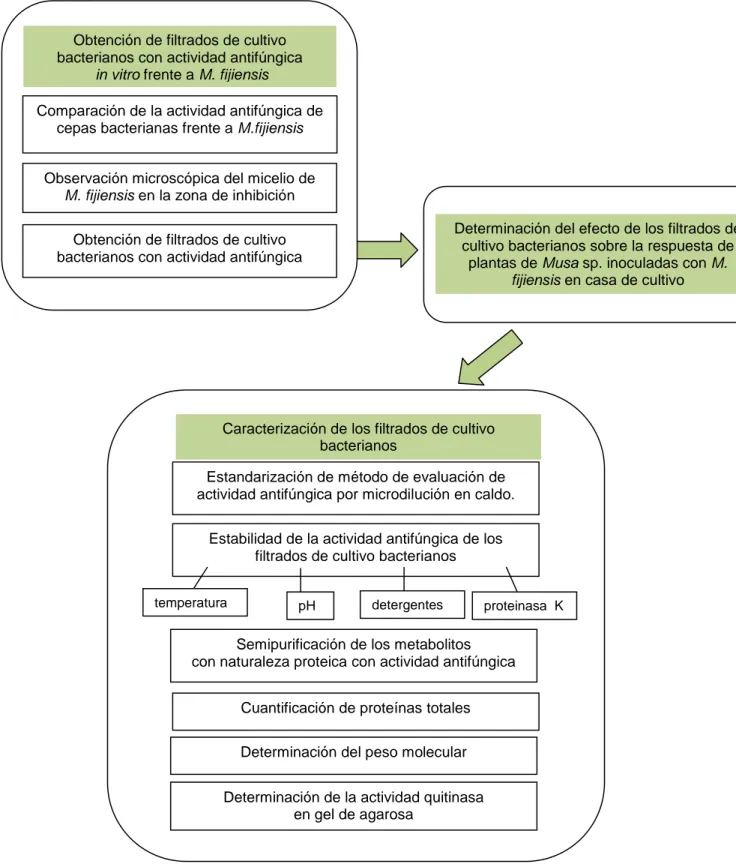
(8) Índice ÍNDICE. Pág.. 1. INTRODUCCIÓN……………………………………………………………………….. 1. 2. REVISIÓN BIBLIOGRÁFICA………………………………………………................ 4. 2.1 Plátanos y bananos………………………………………………………………... 4. 2.2 Enfermedades del cultivo………………………………………………………….. 4. 2.2.1 Mycosphaerella fijiensis……………………………………………………. 5. 2.3 Control……………………………………………………………………………... 7. 2.3.1 Control biológico…………………………………………………………….. 7. 2.3.2 Antagonismo microbiano………………………………………………….. 9. 2.3.2.1 Bacterias con actividad antifúngica…………………………….. 11. 2.4 Bacterias de la filosfera de Musa spp………………………………………….. 13. 2.4.1 Bacterias de la filosfera de Musa spp. como agentes de control biológico……………………………………………………………………. 14. 2.4.2 Aislamiento de bacterias de la filosfera………………………………….. 14. 2.5 Metabolitos y filtrados de cultivo bacterianos…………………………………... 15. 2.5.1 Caracterización de metabolitos con actividad antifúngica………………. 16. 2.5.2 Compuestos proteicos con actividad antifúngica………………………... 17. 3. MATERIALES Y MÉTODOS………………………………………………………….. 19. 3.1 Obtención de filtrados de cultivo bacterianos con antifúngica in vitro frente a M. fijiensis ………………………………………………………………... 20. 3.2 Determinación del efecto de filtrados de cultivo bacterianos sobre la respuesta de plantas de Musa sp. inoculadas con M. fijiensis en casa de cultivo……………………………………………………………. 22. 3.3 Caracterización de los filtrados de cultivo bacterianos con actividad antifúngica in vitro frente a M. fijiensis ……………………... 25. 3.3.1 Estandarización del método para evaluar el crecimiento de M. fijiensis…………………………………….……………………………. 26. 3.3.2 Determinación de la estabilidad de la actividad antifúngica de los filtrados de cultivo bacterianos…………………………………………….. 27. 3.3.2.1 Efecto de la temperatura sobre la actividad antifúngica de los filtrados de cultivo bacterianos……………………………………. 27. 3.3.2.2 Efecto del pH sobre la actividad antifúngica de los filtrados de cultivo bacterianos………………………………………………... 3.3.2.3 Efecto de detergentes sobre la actividad antifúngica de los. 28.
(9) Índice filtrados de cultivo bacterianos………………………...…………. 28. 3.3.2.4 Efecto de proteasas sobre la actividad antifúngica de los filtrados de cultivo bacterianos…………………………………………….... 29. 3.3.3 Caracterización de metabolitos de naturaleza proteica con actividad antifúngica in vitro frente a M. fijiensis…………………………………….. 29. 3.3.3.1 Semipurificación de proteínas con sulfato de amonio…………... 29. 3.3.3.2 Cuantificación de proteínas totales………………….…………….. 30. 3.3.3.3 Determinación del peso molecular……………………………….... 30. 3.3.3.4 Determinación de la actividad quitinasa en geles de agarosa…. 30. 4. RESULTADOS Y DISCUSIÓN……………………………………………………….. 33. 4.1 Obtención de filtrados de cultivo bacterianos con antifúngica in vitro frente a M. fijiensis ………………………………………………………………………... 33. 4.2 Determinación del efecto de filtrados de cultivo bacterianos sobre la respuesta de plantas de Musa sp. inoculadas con M. fijiensis en casa de cultivo……………………………………………………………………………….. 41. 4.3 Caracterización de los filtrados de cultivo bacterianos con actividad antifúngica in vitro frente a M. fijiensis …………………………………………... 51. 4.3.1 Estandarización del método para evaluar el crecimiento de M. fijiensis………………………………………………………… ……….. 51. 4.3.2 Determinación de la estabilidad de la actividad antifúngica de los filtrados de cultivo bacterianos…………………………………………….. 53. 4.3.2.1 Efecto de la temperatura sobre la actividad antifúngica de los filtrados de cultivo bacterianos……………………………………. 53. 4.3.2.2 Efecto del pH sobre la actividad antifúngica de los filtrados de cultivo bacterianos……………………………………………….... 56. 4.3.2.3 Efecto de detergentes sobre la actividad antifúngica de los filtrados de cultivo bacterianos………………………...…………. 59. 4.3.2.4 Efecto de proteasas sobre la actividad antifúngica de los filtrados de cultivo bacterianos…………………………………………….... 62. 4.3.3 Caracterización de metabolitos de naturaleza proteica con actividad antifúngica in vitro frente a M. fijiensis…………………………………….. 64. 4.3.3.1 Semipurificación de proteínas con sulfato de amonio…………... 64. 4.3.3.2 Cuantificación de proteínas totales………………….…………….. 65. 4.3.3.3 Determinación del peso molecular……………………………….... 65.
(10) Índice 4.3.3.4 Determinación de la actividad quitinasa en geles de agarosa…. 66. 5. CONCLUSIONES. 73. 6. RECOMENDACIONES. 74. 7. REFERENCIAS. 75.
(11) Introducción 1. INTRODUCCIÓN Los plátanos y bananos (Musa spp.) constituyen el cuarto cultivo económicamente importante en la alimentación después del arroz (Oryza sativa L.), el trigo (Triticum aestivum L.) y el maíz (Zea mays L.). La producción mundial anual de plátanos y bananos se encuentra en el orden de 102 toneladas métricas y para Cuba es de 249 200, lo que representa el 0,24 % (FAOSTAT, 2011). Las plantaciones de este cultivo son afectadas por Mycosphaerella fijiensis Morelet (anamorfo: Pseudocercospora fijiensis Deighton) que causa la Sigatoka negra. Esta enfermedad es considerada la más perjudicial, ya que provoca una necrosis severa en la hoja y es difícil de controlar (Churchill, 2010). El control de la Sigatoka negra está dado principalmente por el manejo cultural y la aplicación de fungicidas. El control basado sólo en las aplicaciones de fungicidas es posible, pero a un costo muy elevado para los pequeños y medianos productores y con el riesgo de generar resistencia en el hongo (Patiño et al., 2007). Por otra parte, el empleo de agroquímicos no es sustentable, debido a que contaminan el ambiente y el patógeno se ha hecho resistente a fungicidas como benomil, tiabendazol, azoxistrobina, trifloxistrobina, propiconazol (Fu et al., 2010); además de estrobilurinas y los inhibidores de desmetilación (Churchill, 2010). Por ello, se requiere la búsqueda de alternativas que permitan a los pequeños productores controlar la enfermedad y continuar dentro de la actividad agrícola sin afectar la sostenibilidad del medio ambiente. El interés en la conservación del ambiente y la creciente demanda de consumir productos naturales libres de compuestos químicos, ha impulsado investigaciones relacionadas con el control biológico de la Sigatoka negra. Con vistas a lo anterior se han propuesto alternativas como: el uso de microorganismos antagonistas, la adición de sustratos que permitan incrementar las poblaciones bacterianas y la inducción de resistencia a patógenos (Riveros et al., 2002).. 1.
(12) Introducción El empleo de bacterias antagonistas se centra principalmente en la búsqueda de candidatos productores de enzimas hidrolíticas (glucanasas y quitinasas), que degraden las hifas del patógeno. En correspondencia con ello se han realizado investigaciones en géneros bacterianos productores de quitinasas tales como: Bacillus, Pseudomonas y Serratia; con el objetivo de purificarlas para el control de hongos, insectos y artrópodos (Rao-Podile y Neeraja, 2010). Sin embargo, el único producto biológico en el mercado ha sido Serenade®, elaborado a partir de Bacillus subtilis para el control de hongos (Soffia, 2005). Aunque se han logrado avances en la búsqueda para el control biológico, a partir de bacterias nativas de la filosfera, contra Mycosphaerella fijiensis; aún es limitado el conocimiento de las características de los metabolitos producidos por estas y su efecto sobre el patógeno. Por ello, se requiere la caracterización de dichos metabolitos y analizar su efecto antifúngico tanto in vitro como ex vitro. Todo ello, contribuirá a elaborar un bioproducto menos agresivo al ambiente y al desarrollo de nuevas estrategias para el control de M. fijiensis. Teniendo en cuenta las razones expuestas anteriormente se estableció como hipótesis de trabajo: A partir de cepas con antagonismo in vitro frente a Mycosphaerella fijiensis y con el empleo de técnicas bioquímicas y analíticas se podrá caracterizar la actividad antifúngica de metabolitos presentes en filtrados de cultivo. Con vistas a ello, este trabajo se propone el siguiente objetivo general: Caracterizar la actividad antifúngica de metabolitos producidos por bacterias frente a Mycosphaerella fijiensis. Para dar cumplimiento a este se plantearon los siguientes objetivos de trabajo: 1. Obtener filtrados de cultivo bacterianos con actividad antifúngica in vitro frente a M. fijiensis.. 2.
(13) Introducción 2. Determinar el efecto de los filtrados de cultivo bacterianos sobre la respuesta de plantas de ‘Grande naine’ (Musa AAA) inoculadas con M. fijiensis en casa de cultivo. 3. Caracterizar filtrados de cultivo bacterianos con actividad antifúngica in vitro frente a M. fijiensis.. La tesis se estructuró en los siguientes acápites: Introducción, Revisión bibliográfica, Materiales. y. Métodos,. Resultados,. Discusión,. Conclusiones,. Recomendaciones. y. Referencias Bibliográficas. Dentro del texto se intercalaron tablas y figuras.. 3.
(14) Revisión Bibliográfica 2. REVISIÓN BIBLIOGRÁFICA 2.1 Plátanos y bananos Los plátanos y bananos agrupan un gran número de plantas herbáceas del género Musa, tanto híbridos obtenidos horticulturalmente a partir de las especies silvestres Musa acuminata y Musa balbisiana como cultivares genéticamente puros de estas especies. El género Musa pertenece a la familia Musaceae, orden Zingiberales subclase Zingiberidae, Clase Liliopsida, División Magnoliophyta, Reino Plantae (Simpson, 2006). La variedad de cultivares de estas especies está dada por la distribución genómica de Musa acuminata y Musa balbisiana, por lo que existen plantas diploides, triploides y tetraploides (Roux et al., 2006).. La diferencia existente entre plátanos y bananos no se basa en. características botánicas, sino en la forma de consumir la fruta. De forma general, las bananas son más dulces y se ingieren de forma cruda y los plátanos se consumen cocidos, generalmente verdes (López, 1989). Constituyen un alimento de alto consumo a nivel mundial porque contienen una gran cantidad de potasio, además de vitaminas A, E, K, C, B y aminas biogénicas como la dopamina y la serotonina. Este cultivo es utilizado también para la producción de alcohol y la extracción de fibras (Simpson, 2006).. 2.2 Enfermedades del cultivo La diversidad genética existente en los plátanos y bananos hace que los patógenos no ataquen uniformemente a todos los cultivares. Este cultivo es afectado por bacterias, nemátodos, insectos, virus y hongos. La bacteria Ralstonia solanacearum Smith, causa el hereque o moko bacteriano del banano. Entre los nemátodos que atacan los rizomas y causan su podredumbre se encuentran: el barrenador Radopholus similis Cobb; espirales como Helicotylenchus multicinctus Cobb y el de la raíz Meloidogyne javanica Orley. Existen insectos como el picudo negro (Cosmopolites sordidus German) que penetra la base suculenta del pseudotallo y se alimenta de la savia.. 4.
(15) Revisión Bibliográfica Además, el trips del banano (Chaetanaphothrips orchidii Haliday) afecta la cáscara de la banana, por lo cual la pulpa se expone y descompone. También el ácaro del banano (Tetranychus lambi Koch) afecta el fruto, lo que provoca quebraduras en la cáscara y el marchitamiento prematuro. Por otra parte, el Virus del mosaico del pepino y Banana bunchy top virus (BBTV), retrasan el crecimiento de las hojas e inhiben la formación de frutos. Entre los hongos que afectan el cultivo se encuentra Fusarium oxysporum f. sp cubense Schltdl, agente causal del mal de Panamá, que provoca el secado de las hojas y que el pseudotallo se agoste por la muerte progresiva del sistema vascular. Además, Gloeosporium musarum Cke. & Massee provoca antracnosis, forma lesiones negruzcas en la piel del pseudotallo y ocasiona la desecación de los frutos (Agrios, 2005). Por otra parte, se encuentran Mycosphaerella fijiensis Morelet, M. musicola Mulder y M. eumusae Crous, agentes causales del complejo de enfermedades de la Sigatoka en el plátano. De ellos,. M. fijiensis es. considerado el miembro más destructivo, causante de la Sigatoka negra. La enfermedad ha sido diagnosticada en muchos países bananeros, hasta esparcirse por todo el mundo (Churchill, 2010).. 2.2.1 Mycosphaerella fijiensis Mycosphaerella fijiensis, perteneciente a la división Ascomycota, es la forma sexual (telemorfa) del hongo, descrito en 1969 por Morelet. A la forma asexual (anamorfa) se le llama Pseudocercospora fijiensis descrita por Deighton. La Sigatoka negra destruye el área foliar, ya que disminuye la capacidad de las hojas para fotosintetizar y como provoca la maduración prematura del racimo, se reduce la calidad de la fruta. La diseminación de la enfermedad se hace por medio de esporas (ascosporas para la forma sexual y conidios para la forma asexual). Los conidios se forman en menor cuantía con respecto a las ascosporas. Las ascosporas requieren de una película de agua para que ocurra la diseminación, estas constituyen el primer medio de dispersión a largas distancias y durante. 5.
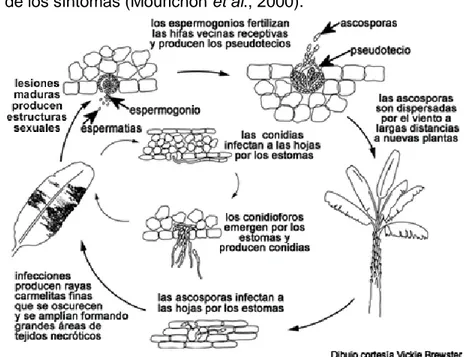
(16) Revisión Bibliográfica épocas lluviosas.. Los peritecios maduran cuando los tejidos muertos de la hoja están. saturados con agua por aproximadamente 48 horas (Marín et al., 2003). Los primeros síntomas de la Sigatoka negra son manchas cloróticas muy pequeñas que aparecen en el envés de la tercera o cuarta hoja abierta, entre los 14 y 20 días después de la infección. Las manchas crecen convirtiéndose en rayas de color marrón delimitadas por las nervaduras. La coloración va haciéndose más oscura y visible en el haz. Cuando el grado de severidad de la enfermedad es alto, grandes áreas de la hoja pueden ennegrecer y lucir mojadas. El ciclo de M. fijiensis consiste de cuatro estados distintos que incluyen la germinación de las esporas, penetración del hospedero, el desarrollo de síntomas y la producción de esporas (Meredith y Lawrence, 1969). En el patosistema Musa-M. fijiensis se observan dos tipos de interacciones: interacciones compatibles, donde se ubican los fenotipos susceptibles y parcialmente resistentes; y las interacciones incompatibles, donde están los fenotipos altamente resistentes. Las interacciones compatibles se caracterizan por un desarrollo completo del ciclo infectivo del patógeno (Figura 1), y las interacciones incompatibles se caracterizan por un bloqueo en el desarrollo de los síntomas (Mourichon et al., 2000).. Figura 1. Ciclo de la enfermedad en el patosistema Musa-M.fijiensis (Bennett y Arneson, 2003). 6.
(17) Revisión Bibliográfica 2.3 Control Mycosphaerella fijiensis tiene un alto impacto económico, social y ambiental, por lo que su control ha atraído la atención de muchos investigadores. El manejo de la Sigatoka negra está dado principalmente por medidas culturales, el uso de cultivares resistentes y el empleo de productos agroquímicos y compuestos de origen natural. El control químico se ha llevado a cabo con el uso de fungicidas protectores y sistémicos en suspensión acuosa, en emulsiones de aceite y agua, o solo con aceite mineral, con activadores de mecanismos de resistencia del hospedante. Entre los fungicidas más utilizados se encuentran el mancozeb, mofolinas, estrobilurinas, triazoles (Pérez, 2006). Según Fu et al. (2010) el control químico de M. fijiensis se basa en la aplicación de estrobilurinas, triazoles, espiroketalaminas e imidazoles, ya que es resistente a fungicidas como benomyl, thiabendazol, azoxystrobin, trifloxystrobin y propiconazol, siendo los triazoles y el procloraz los más efectivos. Churchill (2010) informó la resistencia a estrobilurinas y los inhibidores de desmetilación por parte de M. fijiensis. El control empleando fungicidas es a un alto costo para los pequeños y medianos productores y con el riesgo de generar resistencia en el hongo (Patiño et al., 2007). La constante aplicación de productos químicos no solo ha hecho más resistente al hongo, sino que ha alterado las concentraciones de nutrientes en los cultivos. Esto conjuntamente con el alto costo y las alteraciones del equilibrio ambiental han provocado que el control químico no sea sustentable. En este sentido, se requiere la búsqueda de alternativas que permitan a los pequeños productores controlar la enfermedad y continuar dentro de la actividad agrícola sin afectar la sostenibilidad del medio ambiente (Salazar et al., 2006).. 2.3.1 Control biológico En la agricultura, a nivel mundial, se han hecho cambios con el fin de reducir la contaminación ambiental. Esto ha sido propiciado por la preferencia de los consumidores de. 7.
(18) Revisión Bibliográfica ingerir productos orgánicos de alta calidad. En este sentido, se ha puesto en práctica la implementación de metodologías enfocadas al control biológico de las enfermedades y plagas más comunes que afectan las cosechas de importancia económica. El control biológico es un método del manejo integrado de plagas, enfermedades y malezas que consiste en utilizar organismos vivos con objeto de controlar las poblaciones de otro organismo. Además, presenta una serie de ventajas como: el poco efecto nocivo, la rara resistencia de las plagas, poseen un tiempo de eficacia relativamente largo y la relación costo/beneficio es favorable (Pérez, 2006) En adición, existen estrategias como: el uso de microorganismos antagonistas, la adición de sustratos que permitan incrementar las poblaciones bacterianas, la aplicación de microorganismos antagonistas que promuevan el crecimiento o induzcan resistencia a patógenos (Riveros et al., 2002). El control biológico de la Sigatoka negra es complejo y desafiante debido a la naturaleza policíclica de la enfermedad. Este surgió como una alternativa, debido a la resistencia de M. fijiensis a los fungicidas sistémicos y la creciente demanda por mantener un ambiente seguro. La selección de microorganismos para el control de la Sigatoka negra está enfocado principalmente en la búsqueda de bacterias capaces de secretar enzimas hidrolíticas para degradar la pared celular del hongo, teniendo en cuenta además, el potencial para colonizar la superficie foliar y formar estructuras resistentes que le permitan sobrevivir bajo condiciones ambientales adversas (Marín et al., 2003). Con el fin de aumentar las poblaciones nativas de bacterias con actividad lítica sobre M. fijiensis se han realizado estudios a partir de la modificación de las condiciones físicas y nutricionales de la filosfera de plátanos y bananos. En base a ello, Arango (2002) aplicó sustratos como melaza+quitina+leche y quitina+glucano+nitrato de calcio sobre la filosfera de banano. En este caso, el uso alternado de sustratos con los fungicidas tradicionales permitieron reducir en un 40% la aplicación de químicos. De forma similar, Patiño et al.. 8.
(19) Revisión Bibliográfica (2007) incrementaron las poblaciones de bacterias quitinolíticas y glucanolíticas con potencial biorregulador sobre M. fijiensis mediante la aplicación de sustratos foliares con base en quitina coloidal, harina de cebada, urea y una solución mineral base. Además, como alternativa de control biológico para la Sigatoka negra se han empleado extractos vegetales con actividad antifúngica. Los extractos metanólicos y de diclorometano de diferentes familias de plantas como Asteraceae, Euphorbiaceae, Rubiaceae, Solanaceae empleados por Niño et al. (2007) evidenciaron que algunos agentes protectores o inductores de resistencia por su actividad antifúngica son: cumarinas, compuestos fenólicos, flavonoides, saponinas y quinonas. Se determinó, además, que algunos de estos metabolitos secundarios hacen parte del repertorio de las sustancias que sirven de defensa en las plantas, como los alcaloides, terpenoides y fenilpropanoides. En esta misma línea de investigación, estos autores para el 2009 determinaron que los extractos de plantas de las familias Annonaceae, Apiaceae, Apocynaceae, Asclepiadaceae, Asteraceae, Clusiaceae, Costaceae, Euphorbiaceae, Lamiaceae, Malvaceae,. Melastomataceae,. Moraceae,. Passifloraceae y Urticaceae presentaban también actividad antifúngica. En ese estudio los extractos de diclorometano más activos fueron los de Ureara ballotaefolia (UTP-150, Urticaceae) y el de Costus sp. (UTP-187, Costaceae) y el más potente fue el extracto metanólico de Topobea cf discolor (UTP-160, Melastomataceae) que inhibió el crecimiento de M. fijiensis tanto en la fase sexual como en la sexual (Niño et al., 2009).. 2.3.2 Antagonismo microbiano El antagonismo microbiano no es más que el enfrentamiento entre dos. o más. microorganismos. El control de patógenos mediante el empleo de microorganismos constituye una línea de investigación a nivel mundial. Existe una interacción continua entre los patógenos potenciales y sus antagonistas, de forma tal que estos últimos contribuyen a que en la mayoría de los casos no se desarrolle la enfermedad. En condiciones naturales los. 9.
(20) Revisión Bibliográfica microorganismos están en un equilibrio dinámico en la superficie de las plantas (FernándezLarrea, 2001). Por ejemplo, Cain et al. (2000) demostraron que cepas pertenecientes al género Burkholderia inhibían el crecimiento de otras bacterias, hongos, levaduras y protozoos. En este caso la inhibición no estaba dada por el contacto celular o la presencia de células vivas, sino por la actividad antimicrobiana que se produce por excreción de compuestos extracelulares. Otro ejemplo lo constituyen los hongos Trichoderma y Gliocladium. Ambos ejercen su acción mediante varios mecanismos, entre los cuales tiene un rol importante el parasitismo. Son antagonistas de patógenos de suelos como: Botrytis cinerea Whetzel, Rhizoctonia solani Kühn, Sclerotium rolfsii Sacc y Sclerotium cepivorum Berk (Fernández-Larrea, 2001), entre otros. Varios antagonistas como Burkholderia cepacia (De Costa y Erabadupitiya, 2005), Trichoderma harzianum (Alvindia y Natsuaki, 2008), Pichia anomala, Candida oleophila (Williamson et al., 2008), Bacillus amyloliquefaciens, Pseudomona fluorescens, P. syringae, P. aeruginosa, P. aureofaciens, P. putida, P. pyrrocinia (Alvindia y Natsuaki, 2009), Pantoea agglomerans y Flavobacterium sp. (Niroshini-Gunasinghe y Karunaratne, 2009) han sido seleccionados para el control de enfermedades post cosecha en bananos. Autores como Osorio et al. (2004), realizaron pruebas de antagonismo sobre ascosporas de M. fijiensis, con bacterias quitinolíticas aisladas del filoplano de banano en un ensayo in situ sobre discos de hojas también de esta planta, y encontraron que algunos de los aislados inhibieron la germinación de las ascosporas o deformaron sus tubos germinativos, hasta en un 40 y 85% respectivamente. Tal efecto, en parte, fue explicado por la actividad de las enzimas quitinolíticas sobre la pared del patógeno. Por otra parte, Arzate-Vega et al. (2006) evaluaron el efecto antagónico de cepas de Trichoderma spp. sobre M. fijiensis in vitro y en invernadero, donde la inhibición del crecimiento del patógeno se debió al contacto entre los hongos. Como principal resultado. 10.
(21) Revisión Bibliográfica obtuvieron una disminución de los porcentajes de infección en un 70 y 88% para el experimento del invernadero. Los antagonistas no tienen un único modo de acción y la multiplicidad de éstos es una característica importante para su selección como agentes de control biológico. Si el antagonista posee varios modos de acción reduce los riesgos de desarrollo de resistencia en el patógeno (Fernández-Larrea, 2001). El antagonismo de los microorganismos está dado por diferentes mecanismos de acción. Entre ellos se encuentra la competencia por el nicho ecológico o sustrato, la producción de antibióticos y la producción de metabolitos que afectan directamente el patógeno o inducen resistencia sistémica en la planta (Hu et al., 2008; Lian et al., 2008). En este sentido, los mecanismos de prevención y eliminación del efecto de los patógenos están dados por la producción de sideróforos, cianuro de hidrógeno, reguladores del crecimiento (giberelina, auxina, citoquinina, etileno, ácido abscísico), la síntesis de antibióticos o de enzimas que degradan la pared celular, la solubilización de fosfatos y otros nutrientes (Hayat et al., 2010).. 2.3.2.1 Bacterias con actividad antifúngica Muchos géneros bacterianos han sido objeto de investigaciones por. poseer actividad. antifúngica frente a patógenos de plantas. Los más estudiados han sido las bacterias pertenecientes a los géneros Bacillus, Serratia y Pseudomonas (Marín et al., 2003). Estos géneros se destacan por sintetizan un gran número de compuestos antimicrobianos, que afectan el crecimiento de hongos patógenos, por lo que han utilizados tradicionalmente como agentes de biocontrol tanto las propias bacterias como los metabolitos purificados. Por ejemplo, ya existe en el mercado el biofungicida Serenade®, efectivo contra la Sigatoka negra en programas integrados con fungicidas convencionales. Este funciona frente a M. fijiensis por la acción biológica de la cepa Bacillus subtilis Cohn QST 173 en conjunto con los lipopéptidos que esta bacteria produce. Estos pertenecen a tres clases específicas: iturinas,. 11.
(22) Revisión Bibliográfica plipastatinas y surfactinas (Soffia, 2005). Este producto fue desarrollado por AgraQuest Inc.,. con los números de patentes para Estados Unidos 6.060.051; 6.103.228;. 6.291.426 y 6.417.163 y con el número de Registro de la EPA 69592-7 (Ceballos, 2009). La obtención de compuestos a partir del género Bacillus constituye una temática de investigación en el área del control biológico. Esto se debe al amplio espectro de microorganismos que puede controlar y a la diversidad de compuestos que producen con actividad antifúngica. En este sentido, autores como Munimbazi y Bullerman (1998) refieren cómo Bacillus pumilus Cohn inhibe el crecimiento de Aspergillus spp., Penicillium spp., Fusarium spp. y la producción por estos de aflatoxinas, ácido ciclopiazónico, ocratoxina A y patulina. Además de otras bacterias de este género frente a Aspergillus flavus Link (Moyne et al., 2001), Magnaporthe grisease Heber, Sclerotinia slerotirum Bary, Rhizoctonia solani, Alternaria oleracea, A. brassicae y Botrytis cinerea (Liu et al., 2007) y A. solani (Hu et al., 2008). Igualmente, Paenibacillus lentimorbus Ash NRRL B-30488 fue aislada de la leche de vaca e inoculada posteriormente en plantas de maíz, pepino (Cucumis sativus L.), sorgo (Sorghum bicolor L.), trigo y otros cultivos. El resultado fue un incremento en la masa seca, la promoción del crecimiento de las plantas inoculadas en comparación con aquellas que no fueron inoculadas con bacterias; además de mostrar competencia con los microorganismos nativos. La interacción entre Fusarium oxysporum f. sp ciceri y B-30488 mostró que la bacteria degradaba las hifas del hongo debido a la producción de enzimas hidrolíticas: quitinasas y β-1,3-glucanasas (DasGupta et al., 2006). En correspondencia con ello, RaoPodile y Neeraja (2010) informaron la producción de quitinasas en géneros bacterianos tales como: Aeromonas, Arthrobacter, Beneckea, Bacillus, Chromobacterium, Clostridium, Klebsiella, Pseudomonas, Serratia, Streptomyces y Vibrio. Los autores plantearon, además, que en el género Bacillus la actividad quitinasa es eficiente y se han realizado estudios en varias especies tales como: B. circulans, B. subtilis, B. alvei, B. lentus, B. cereus, B.. 12.
(23) Revisión Bibliográfica licheniformis y B. thuringiensis. Por otra parte, se han utilizado productos volátiles producidos por B. megaterium Cohn para controlar el nemátodo Meloidogyne incognita (Huang et al., 2009). Además, se han empleado lipopéptidos antibióticos de B. subtilis para el control de enfermedades causadas por Fusarium graminearum en maíz y trigo (Chan et al., 2009).. 2.4 Bacterias de la filosfera de Musa spp. El término filosfera fue propuesto por Last en 1955 y Ruinen un año después, para describir la superficie de las hojas de las plantas como un ambiente que es física, química y biológicamente diferente del resto de la hoja o del ambiente exterior que lo rodea (Riederer y Muller, 2006). Las bacterias asociadas a la hoja pueden estar localizadas en el tejido laminar o particularmente en las cámaras subestomáticas, en espacios intercelulares y en sitios protegidos en la superficie de la hoja (Jacques y Morris, 1995). Los microorganismos endófitos son aquellos que tienen todo o parte de su ciclo de vida invadiendo el tejido de la planta. Las bacterias epifíticas son las representantes mayoritarias de las filosfera y su densidad se ha sido estimada entre 106 y 107 unidades formadoras de colonia (UFC)/cm2 en una hoja típica (Gnanamanickam e Immanuel, 2007). Salazar (2005) realizó un análisis químico de la cantidad de minerales, carbohidratos y proteínas existentes en. la. filosfera de diferentes cultivares de Musa spp. en Urabá,. Colombia. Para todos los cultivares evaluados, esta presentó pobreza en la mayoría de los minerales, con excepción del sodio, potasio y fósforo. Según Salazar et al. (2006) el contenido de carbohidratos y proteínas en la filosfera es inferior al 0,05%; por lo que constituye un ambiente oligotrófico, donde las concentraciones de nutrientes son bajas. A pesar de que constituye un microclima extremo con variabilidad en los parámetros climáticos como la intensidad de la luz y la temperatura (Gnanamanickam e Immanuel, 2007) existen bacterias capacitadas para vivir ahí, debido a sus estrategias de supervivencia.. 13.
(24) Revisión Bibliográfica Según Suda et al. (2009) las comunidades de microorganismos presentes en la filosfera, fluctúan en dependencia del cultivo y del ambiente. A pesar de los estudios realizados, poco es el conocimiento sobre la interacción entre la comunidad de hongos y bacterias en la filosfera, en contraste con los conocimientos que se tienen acerca de la rizosfera.. 2.4.1 Bacterias de la filosfera de Musa spp. como agentes de control biológico Un método dentro del manejo integrado de plagas lo constituye la aplicación de agentes de control biológico (Pérez, 2006). Se han realizado algunas investigaciones basadas en la búsqueda de antagonistas nativos de la filosfera de musáceas. Por ejemplo, Osorio et al. (2004), realizaron una selección de bacterias quitinolíticas nativas del Urabá antioqueño, Colombia, con potencial antagonista contra M. fijiensis. De los aislados encontraron algunas con capacidad para producir enzimas quitinolíticas y glucanolíticas, que afectaron la germinación de las ascosporas del hongo en un 42% y deformaron sus tubos germinativos en un 87%. Por otra parte, Salazar. (2005). efectuó una determinación parcial de las. bacterias epífitas aisladas de la filosfera de Musa spp. En este proceso, se seleccionaron 80 cepas quitinolíticas y 15 glucanolíticas, con potencial biorregulador contra M. fijiensis. El estudio mostró que se encontraron bacilos Gram negativos de forma predominante y bacilos y cocos Gram positivos. Por su parte, Alvindia y Natsuaki (2009) lograron reducir significativamente la incidencia de la podredumbre de la corona en bananos aplicando, poscosecha,. Bacillus amyloliquefaciens aislado de la superficie del fruto. Estos autores. demostraron así, que la microbiota nativa puede influir en el crecimiento de los patógenos y reducir las enfermedades foliares de los cultivos.. 2.4.2 Aislamiento de bacterias de la filosfera Existen diversos métodos para aislar bacterias de la filosfera. Su selección adecuada está en estrecha correspondencia con el fin de la investigación, ya que presentan ventajas y. 14.
(25) Revisión Bibliográfica desventajas. Según Jacques y Morris (1995) los métodos se clasifican en directos o indirectos. Entre los directos se encuentran: la observación al microscopio de la superficie de la hoja y el análisis sobre esta. Los indirectos consisten en la impresión de la hoja en medio de cultivo agar nutriente y la posterior observación microscópica. Existen otros métodos como la maceración o pulverización, la emulsión estomática, el lavado en un líquido y la sonicación, con el fin de aislar los microorganismos adheridos a la superficie y cuantificar el tamaño de las poblaciones microbianas asociadas a las hojas. Para el método del lavado de las hojas en un líquido, es importante tener en cuenta la composición de este, la intensidad y duración del proceso. Generalmente se utiliza solución amortiguadora de fosfato de sodio estéril 0,1 M a pH=7 y la posterior agitación de las muestras (Ceballos, 2009). Por otra parte, la maceración de tejido de la hoja, permite una mayor recuperación de microorganismos ya que son liberados los endófitos también. Este método tiene como desventaja la posible liberación del contenido celular de la planta, pudiendo lisar o inhibir el crecimiento de algunos microorganismos (Jacques y Morris, 1995).. 2.5 Metabolitos y filtrados de cultivo bacterianos Los microorganismos utilizados para el control biológico han sido aplicados directamente sobre la filosfera o a partir de ellos se han obtenido filtrados de cultivo o metabolitos para la formulación de un bioproducto. La obtención de los metabolitos bacterianos está en correspondencia con el tipo de molécula en cuestión y los diferentes métodos para la extracción y purificación existentes. Mediante el uso de filtrados de cultivo bacterianos se ha logrado la inhibición del crecimiento micelial de diferentes hongos fitopatógenos. Por ejemplo, Munimbazi y Bullerman (1998) comprobaron la inhibición del crecimiento de: Aspergillus, Penicillium y Fusarium, a partir del uso de filtrados de cultivo de B. pumilus. De forma similar, han sido empleados a partir del género Bacillus frente al patógeno Curvularia lunata (Basha y Ulaganathan, 2002), en un estudio de la cepa B.subtilis QM3 frente a. 15.
(26) Revisión Bibliográfica Alternaria solani (Hu et al., 2008) y en B. subtilis cepa B 106 contra la Sigatoka amarilla (Fu et al., 2010). Por otra parte, se han aislado y caracterizado metabolitos de naturaleza proteica con actividad antifúngica lo que constituye un avance para la posterior elaboración de productos de biocontrol. Un ejemplo es la producción de metabolitos difundidos por parte de B. amyloliquefaciens RC2 (Hatakeda y Shirata, 2001) así como en la obtención de bacillomycina D, una iturina contra Aspergillus flavus (Moyne et al., 2001) o de bacisubina, a partir de la cepa B-916 de Bacillus subtilis, con actividad inhibitoria del crecimiento micelial de Magnaporthe grisease, Sclerotinia sclerotioum, Rhizoctonia solani, Alternaria oleracea Milbr, A. brassicae Berk y Botrytis cinerea (Liu et al., 2007).. 2.5.1 Caracterización de metabolitos con actividad antifúngica La caracterización de metabolitos con actividad antifúngica se realiza con el fin de determinar su naturaleza química y su estabilidad ante factores abióticos, para la optimización en la producción y el escalado en la elaboración de productos de biocontrol, así como, para la dilucidación de los mecanismos por los que ejercen la actividad antifúngica. Para ello, se analiza la pérdida o no de actividad antifúngica ante diferentes valores de pH y temperatura, la degradación de estos por enzimas o detergentes, además de su solubilidad en solventes orgánicos o inorgánicos. Por ejemplo, Munimbazi y Bullerman (1998) determinaron la estabilidad de metabolitos antifúngicos producidos por B. pumillus a 121°C, además de analizar la pérdida o no de actividad antifúngica ante diferentes valores de pH (2, 4, 6, 8 y 10). También, evaluaron la resistencia a la hidrólisis por enzimas, la sensibilidad ante detergentes (aniónicos, catiónicos y no iónicos) y la solubilidad en solventes orgánicos. Los autores determinaron que los metabolitos producidos por B. pumillus eran estables ante variaciones de la temperatura, el pH, resistentes a la hidrólisis enzimática y a la acción de algunos de los detergentes empleados.. 16.
(27) Revisión Bibliográfica De igual forma Lee et al. (2008) caracterizaron metabolitos antifúngicos producidos por Paenibacillus lentimorbus respecto a la estabilidad térmica a 60°C y 121°C, a valores de pH (2 y 13), la desnaturalización por detergentes y enzimas. Los resultados mostraron que solo hubo pérdida de la actividad cuando los metabolitos fueron sometidos a 121°C, el detergente CTAB y la lisozima. Por su parte, Hu et al. (2008) determinaron el efecto de la temperatura (25, 60, 100 y 121°C) sobre la producción de compuestos por una cepa de B. subtilis, donde solo se afectó la actividad a temperaturas altas. Por otra parte, Fu et al. (2010) analizaron el efecto del tiempo, la temperatura, el pH y el crecimiento en diferentes medios de cultivo sobre la actividad inhibitoria de la cepa B 106 de B. subtilis frente a Pseudocercospora musae. Los autores, seleccionaron un tiempo de incubación, con una temperatura, pH y medio de cultivo óptimo para potenciar la actividad antifúngica de esta cepa bacteriana. Por otra parte, para la caracterización de metabolitos, se puede determinar su estructura por resonancia magnética nuclear (NMR) y espectroscopia de masa (MS). Existen también pruebas para analizar la actividad hidrolítica (celulasa, pectinasa, fosfatasa, proteolítica o lipasa), la producción de ácido indol acético (AIA) y cianuro de hidrógeno (HCN), además de la detección de genes que codifican para antibióticos, por PCR (del inglés: Polymerase Chain Reaction) (Sunish Kumar et al., 2005). Además, los autores determinaron que el metabolito producido por Pseudomona aeruginosa PUPa3 era fenazin-1-carboxamida según los resultados de la NMR (del inglés: Nuclear Magnetic Resonance) y MS (del inglés: Mass spectroscopy), además de la producción de AIA, sideróforos, fosfatasas y proteasas.. 2.5.2 Compuestos proteicos con actividad antifúngica Entre los metabolitos bacterianos con actividad antifúngica se destacan los de naturaleza proteica, entre los que se encuentran enzimas líticas y péptidos antibióticos. Por ejemplo, Lee et al. (2008) caracterizaron los metabolitos extracelulares de naturaleza proteica. 17.
(28) Revisión Bibliográfica producidos al final de la fase exponencial, previamente semipurificados, a partir de Paenibacillus lentimorbus. Estudios de péptidos antimicrobianos frente a M. fijiensis evidencian que estos son capaces de inhibir procesos celulares específicos, causar formación anormal del tubo germinativo e inhibir el crecimiento micelial del patógeno (Vásquez et al., 2009). Las bacterias del género Bacillus han sido empleadas para el control biológico, debido a la capacidad para producir metabolitos secundarios con un amplio espectro de actividad antibiótica. B. subtilis sintetiza pequeños péptidos no ribosomales entre los que se encuentran: iturina, surfactina, fengicina, plipastatina (Moyne et al., 2001) además de bacisubina (Liu et al., 2007). Entre los compuestos liberados al medio de cultivo por las bacterias con actividad antifúngica, se encuentran enzimas que degradan la pared de ascomicetos y basidiomicetos. Por ello, para el control de Mycosphaerella fijiensis entre los métodos más empleados se encuentra el potenciamiento de la colonización de bacterias glucanolíticas o quitinolíticas, nativas de la filosfera de banano, a través de sustratos foliares (Peláez et al., 2006).. 18.
(29) Materiales y Métodos 3. MATERIALES Y MÉTODOS El trabajo se realizó en el Laboratorio de Microbiología Aplicada del Instituto de Biotecnología de las Plantas (IBP) de la Universidad Central ¨Marta Abreu¨ de Las Villas (UCLV), en el período comprendido entre los años 2010-2012. Aislado de Mycosphaerella fijiensis El aislado de M. fijiensis empleado en todos los ensayos fue el CCIBP-Pf-83 perteneciente a la Colección de Cultivos Microbianos del Laboratorio de Microbiología Aplicada del IBP. Para su cultivo el aislado fue inoculado en Enlermeyers de 100 mL de volumen con 50 mL de Caldo Papa Dextrosa (PDB) e incubado en agitación a 120 rpm y 28oC, durante 15 días. Cepas bacterianas Para el estudio se emplearon 19 cepas bacterianas pertenecientes a la Colección de Cultivos Microbianos del IBP. Estas fueron aisladas de la filosfera de Musa spp. y se seleccionaron previamente por poseer actividad antifúngica in vitro frente a M. fijiensis (Poveda et al., 2010). Además, se incluyó en todos los ensayos la cepa de Bacillus subtilis (CCIBP-M27), aislada como contaminante del cultivo in vitro, con actividad antifúngica in vitro frente a este patógeno (Cruz-Martín et al., 2010). Material vegetal Se utilizaron plantas de Musa spp., pertenecientes al cultivar ‘Grande naine’ (Musa AAA), propagadas in vitro vía organogénesis según el protocolo descrito por Orellana (1994). Procesamiento estadístico de los datos El procesamiento estadístico de los datos de las variables evaluadas se realizó con el paquete estadístico Statistic Package for Social Science (SPSS) versión 19,0 para Windows. Las pruebas utilizadas para cada ensayo, previa comparación de normalidad y heterogeneidad de varianza, se detallan más adelante.. 19.
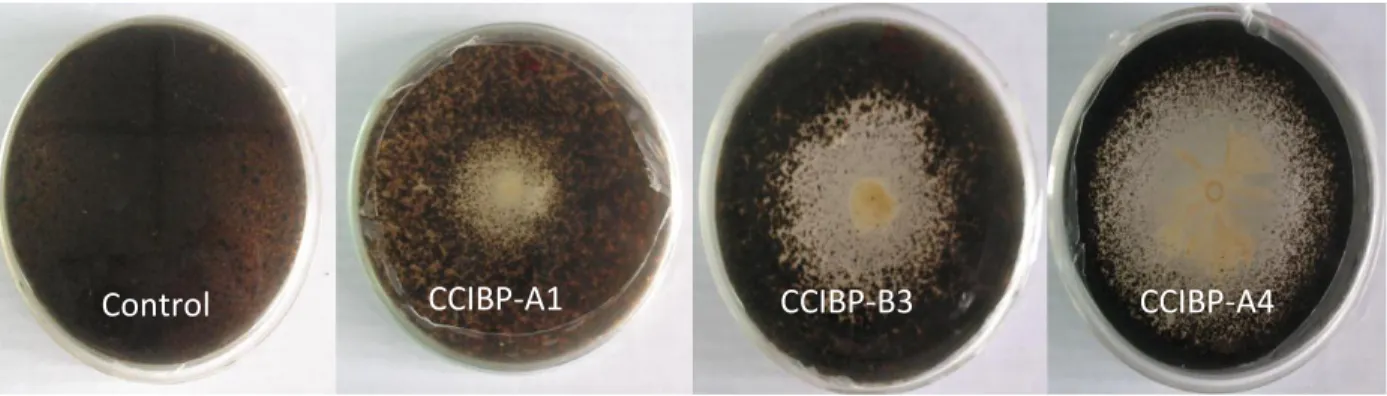
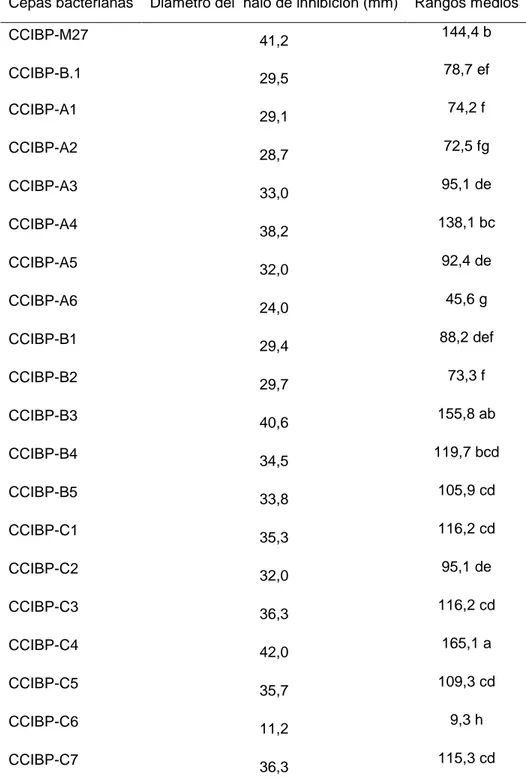
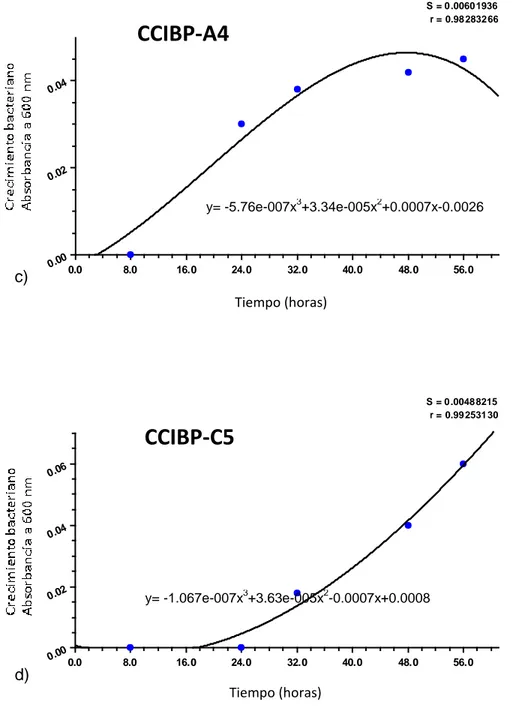
(30) Materiales y Métodos 3.1 Obtención de filtrados de cultivo bacterianos con actividad antifúngica in vitro frente a M. fijiensis Para obtener filtrados de cultivo bacterianos con actividad antifúngica in vitro frente a M. fijiensis se procedió como se describe a continuación: a. Selección de cepas con mayor actividad antifúngica in vitro frente a M. fijiensis b. Curva de crecimiento en medio de cultivo líquido c. Obtención de filtrados de cultivo y comprobación de la actividad antifúngica frente a M. fijiensis. Selección de cepas Para comparar la actividad antifúngica de las cepas frente a M. fijiensis se empleó el método de cultivo dual. Se prepararon placas de Petri (90,0 mm) para el cultivo dual como se describe a continuación. En un Erlermeyer con 200 mL de Papa Dextrosa Agar (PDA) (Duchefa) fundido a 40oC se añadieron 20 mL de una suspensión micelial de M. fijiensis (para una concentración final de 5,0x105 fragmentos de micelio/mL). Se homogenizó la mezcla y se vertió en las placas de Petri. Cuando las placas estuvieron secas, se añadieron en el centro de la placa 7,0 µL de cada suspensión bacteriana (un cepa por placa de Petri) ajustada a DO600=0,1 (Eppendorf Biophotometer), a partir de cultivos bacterianos de 24 horas de crecidos en Agar Nutriente (AN). Como control se emplearon 7,0 µL de agua desionizada estéril en lugar de la suspensión bacteriana. Las placas se incubaron a 28ºC durante 72 horas. La evaluación se realizó mediante la medición del halo de inhibición (mm) del crecimiento del patógeno a las 72 horas de incubación. Se emplearon tres réplicas por cada cepa bacteriana y el ensayo se repitió tres veces. Los valores obtenidos fueron analizados estadísticamente mediante las pruebas de Kruskal-Wallis y Mann Whitney, previa comprobación de los supuestos de normalidad y heterogeneidad de varianza.. 20.
(31) Materiales y Métodos Se clasificaron como positivas las cepas bacterianas que produjeron halo de inhibición del crecimiento del patógeno. Además, se realizó observación microscópica de las hifas del patógeno presentes en la zona de inhibición, para describir sus características. A partir de los resultados de este acápite se seleccionaron cuatro cepas bacterianas para continuar los experimentos. Curva de crecimiento en medio de cultivo líquido Para obtener un filtrado de cultivo con actividad antifúngica, se elaboró la curva de crecimiento de las cepas en medio de cultivo Caldo Nutriente (CN) (BioCen). Se empleó como inóculo 1,0 mL de suspensión bacteriana ajustada a DO600nm=0,1 (Eppendorf Biophotometer) (~ 109 ufc/mL) en Erlenmeyers con 100 mL de medio de cultivo. Las muestras se incubaron a 30ºC, a 120 rpm durante 72 horas. Cada ocho horas y hasta las 72 horas se midió la absorbancia a DO600nm con un espectrofotómetro (Eppendorf Biophotometer). Como blanco se empleó el medio de cultivo sin inocular. Se realizaron tres réplicas para cada una de las cepas y el experimento se realizó dos veces. Las curvas de crecimiento (Absorbancia a DO600nm en función del tiempo), se realizaron con el software Curvaexpert. Obtención de filtrados de cultivo y comprobación de la actividad antifúngica frente a M. fijiensis Teniendo en cuenta la curva de crecimiento de las cepas a las 24 y 48 horas de incubación los cultivos bacterianos se centrifugaron por 15 minutos a 4ºC y 10 000 rpm (Eppendorf Centrifuge 5810 R). Para eliminar las células bacterianas, el sobrenadante fue filtrado (0,22 µm) y se conservó a 4ºC. El sobrenadante libre de células bacterianas se denominó filtrado de cultivo bacteriano. La actividad antifúngica in vitro de cada filtrado se comprobó mediante el método de dilución en Agar (dilución 1:1). Para ello, se utilizó una placa de 96 pocillos en base a resultados previos no mostrados. En cada pocillo se colocaron 125 µL de una suspensión micelial de M.. 21.
(32) Materiales y Métodos fijiensis en medio de cultivo Papa Dextrosa Agar (PDA) (Duchefa) (para una concentración final aproximada de 5,0x105 fragmentos de micelio/mL) y 125 µL de filtrado de cultivo estéril de cada una de las cepas. En el control se aplicó, en lugar de los filtrados bacterianos, 125 µL de medio de cultivo CN. Las placas se incubaron a 28ºC y oscuridad durante 48 horas y se colocaron ocho réplicas por tratamiento. Se informó que los filtrados tenían actividad antifúngica si se observaba inhibición del crecimiento micelial con respecto al control, según lo descrito por Poveda (2009). Se seleccionó el tiempo de incubación (24 o 48 horas) de las suspensiones bacterianas de las cepas seleccionadas, donde todos los filtrados de cultivo presentaron actividad antifúngica.. 3.2 Determinación del efecto de los filtrados de cultivo bacterianos sobre la respuesta de plantas de Musa spp. inoculadas con M. fijiensis en casa de cultivo Con el objetivo de determinar el efecto de los filtrados de cultivo bacterianos, con actividad antifúngica in vitro frente a M. fijiensis, en plantas de ‘Grande naine’ (Musa AAA) inoculadas con este patógeno en casa de cultivo se desarrolló este experimento. Los filtrados de cultivo bacterianos para la inoculación se obtuvieron como se describió en el acápite anterior. Se emplearon dos tratamientos para cada una de las cepas: 1) inoculación de los filtrados de cultivo bacterianos de cada cepa tres días antes (A) de la inoculación con M. fijiensis y 2) la aplicación de los filtrados de cultivo tres días posteriores (D) a la inoculación de M. fijiensis. Para un total de nueve tratamientos, incluyendo el control. Para este estudio se tuvo en cuenta el procedimiento descrito por Leiva-Mora et al. (2010) que se describe brevemente a continuación. Las plantas se colocaron en bolsas de polietileno con sustrato compuesto por 50% de humus de lombriz, 30% de compost y 20% de zeolita a una razón de 5:3:2 (v/v). Se mantuvieron en fase de aclimatización durante 45 días. Posteriormente, se transfirieron a macetas plásticas. 22.
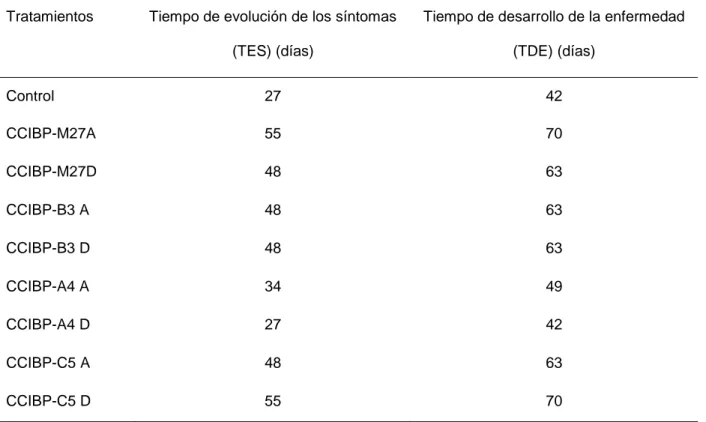
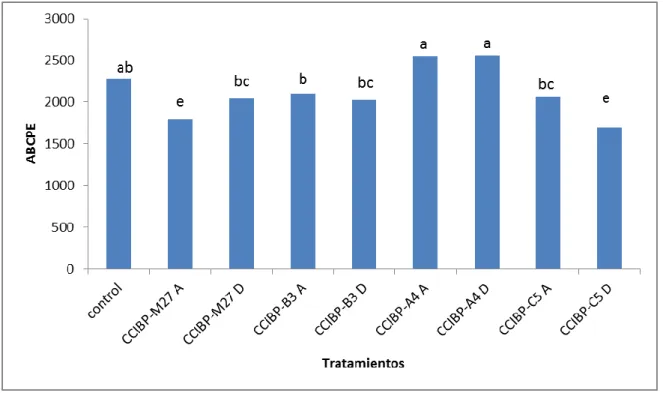
(33) Materiales y Métodos de 20 cm de diámetro con 500 mL de capacidad con igual sustrato por 45 días hasta alcanzar como mínimo 20 cm de altura y tres hojas activas. En la casa de cultivo el riego se efectuó por aspersión tres veces al día y la iluminación fue solar, con una media en la intensidad luminosa de 3 841 µmol.m2/s (medido con Extech Light Meter 401025, USA). La suspensión micelial de M. fijiensis se ajustó a 5,0x105 fragmentos de micelio/mL. La inoculación se realizó por la parte abaxial de las hojas y con la ayuda de un pincel tanto para la inoculación con M. fijiensis como para los filtrados de cultivo. Se inocularon las tres primeras hojas de cinco plantas para cada uno de los tratamientos, (n=15). Se emplearon como controles cinco plantas inoculadas con M. fijiensis y medio de cultivo CN. Las plantas se ubicaron completamente al azar en la casa de cultivo. La evaluación del desarrollo de la enfermedad se realizó cada siete días hasta los 70 días posteriores a la inoculación (dpi). Para ello se utilizó la escala propuesta por Alvarado-Capó et al. (2003) (Figura 2). Según lo propuesto por Leiva-Mora et al. (2010) en cada tratamiento se determinó las variables epifitiológicas: período de incubación. (PI) (días), tiempo de. evolución de los síntomas (TES) (días), tiempo de desarrollo de la enfermedad (TDE) (días) y área bajo la curva del progreso de la enfermedad (ABCPE). Esta última variable se calculó de acuerdo con la fórmula propuesta por Shaner y Finney (1977):. Donde: Yi = expresa la severidad (en función del índice de infección acorde con la escala), Xi =tiempo (días) a la i-ésima observación y n = número total de observaciones.. El índice de infección (II) se calculó según la fórmula:. 23.
(34) Materiales y Métodos. donde n= número de hojas en cada nivel de la escala b= valor de la escala N= 6 (grados de la escala) T= número total de hojas evaluadas por plantas, y se utilizará la escala evaluativa que aparece en la figura 2. Período de incubación (PI): Tiempo entre la inoculación y la aparición de las primeras lesiones puntiformes por el envés de la hoja (días). Tiempo de evolución de los síntomas (TES): número de días entre la aparición de los primeros síntomas (lesiones puntiformes) y la aparición de manchas necróticas con centros secos. Tiempo de desarrollo de la enfermedad (TDE): período entre la inoculación y la aparición de lesiones maduras (manchas necróticas con centros secos). Además, se evaluaron los componentes de la resistencia: número de lesiones necróticas y área de la lesión. Se le determinó el área a 50 lesiones por tratamiento mediante la siguiente fórmula para una elipse, a partir de mediciones obtenidas con una regla graduada:. donde a=largo de la lesión y b=ancho de la lesión. 24.
(35) Materiales y Métodos. Figura 2. Descripción de los estados de desarrollo de los síntomas en plantas de Musa spp. propagadas in vitro inoculadas con suspensión micelial de M. fijiensis de acuerdo con Alvarado-Capó et al. (2003). Los valores obtenidos fueron comparados con el control y analizados estadísticamente mediante las pruebas de Kruskal-Wallis y Mann Whitney, previa comprobación de los supuestos de normalidad y heterogeneidad de varianza.. 3.3 Caracterización de los filtrados de cultivo bacterianos con actividad antifúngica in vitro frente a M. fijiensis Con el objetivo de caracterizar los filtrados de cultivo obtenidos previamente se desarrollaron los siguientes experimentos.. 25.
(36) Materiales y Métodos 3.3.1 Estandarización del método para evaluar el crecimiento de M. fijiensis Con el objetivo de estandarizar un método que permitiera evaluar el crecimiento de M. fijiensis en presencia de volúmenes reducidos de filtrado de cultivo bacterianos se comparó el crecimiento de este patógeno en medio de cultivo líquido en frascos de 250 mL de capacidad con 100 mL de medio de cultivo (Erlenmeyers) y placas de 96 pocillos. Se realizaron las dos curvas de crecimiento de M. fijiensis, una a partir de las mediciones de masa seca (g) del crecimiento micelial en los Erlenmeyer y la otra a partir de la medición de la absorbancia a DO595nm en placas de 96 pocillos. Se compararon los valores de masa seca del micelio y de absorbancia. Para ello, en placas de 96 pocillos estériles, se inocularon por pocillo 250 μL de una suspensión micelial de M. fijiensis en medio de cultivo cultivo Caldo Papa Dextrosa, pH 5,6; (para una concentración final aproximada de 5,0x105 fragmentos de micelio/mL). Estas se incubaron a 28ºC y oscuridad durante diez días. Simultáneamente se inocularon Erlenmeyer (250 mL de volumen) con 100 mL medio de cultivo PDB, con una suspensión micelial de M. fijiensis a igual concentración. Los frascos se colocaron en zaranda (Gerhardt), a 28ºC y 120 rpm durante diez días. Para obtener los valores de masa seca se eliminó el medio de cultivo por filtración y el micelio se secó en estufa a 60ºC, hasta obtener masa constante. Los valores de Absorbancia a DO595nm fueron obtenidos mediante un espectrofotómetro (Thermo Labsystems Opsys MR). Las mediciones se hicieron a los tres, cinco, siete y diez días de incubación. Finalmente, se determinó la relación entre los valores de absorbancia con respecto a la masa seca mediante un análisis de regresión lineal según el método propuesto por Broekaert et al. (1990).. 26.
(37) Materiales y Métodos 3.3.2 Determinación de la estabilidad de la actividad antifúngica de los filtrados de cultivo bacterianos Este ensayo se realizó con el objetivo de determinar la estabilidad de la actividad antifúngica de los filtrados de cultivos bacterianos, de acuerdo con el efecto de la temperatura, el pH, y la presencia de detergentes y proteasas. Se emplearon filtrados de cultivos estériles por filtración, obtenidos de igual forma al acápite 3.1, de las cepas de estudio en el medio de cultivo y tiempo de incubación seleccionado. En todos los casos, para determinar la actividad. antifúngica de los filtrados de cultivo. expuestos a los diferentes tratamientos, se empleó el método de microdilución en placas de 96 pocillos. Se utilizó una dilución [1:10]; 25 µL de filtrado de cultivo sometido a variaciones de temperatura, pH y la presencia de detergentes y proteasa respectivamente y 225 µL una suspensión micelial de M. fijiensis en medio de cultivo PDB (a una concentración de 5,0x105 fragmentos de micelio/mL). Se tomó como control CN estéril sometido a similares condiciones. Además, incluyó un control de crecimiento, el cual consistía en 225 µL de medio de cultivo PDB con suspensión micelial de M. fijiensis y 25 µL de CN. Las placas se incubaron durante el tiempo seleccionado, a 28ºC y oscuridad. La evaluación se realizó mediante la lectura de absorbancia a DO595nm, en espectrofotómetro (Thermo Labsystems Opsys MR). Los valores obtenidos para cada caso, fueron analizados estadísticamente mediante la prueba de Kruskal-Wallis-Mann Whitney, previa comprobación de los supuestos de normalidad y heterogeneidad de varianza.. 3.3.2.1 Efecto de la temperatura sobre la actividad antifúngica de los filtrados de cultivo bacterianos Para determinar el efecto de la temperatura sobre la actividad antifúngica, se tomaron 250 µL de filtrado de cultivo estéril, proveniente de cada una de las cepas bacterianas, y se. 27.
(38) Materiales y Métodos colocaron a diferentes temperaturas (-80, 4, 28, 60 y 121ºC) durante 20 minutos. Pasado este tiempo se inocularon y evaluaron los filtrados según lo descrito en el acápite 3.3.2. Se utilizaron ocho réplicas por tratamiento.. 3.3.2.2 Efecto del pH sobre la actividad antifúngica de los filtrados de cultivo bacterianos Para determinar el efecto del pH sobre la actividad antifúngica de los filtrados de cultivo bacterianos se siguió el protocolo propuesto por Munimbazi y Bullerman (1998). Se disolvieron los filtrados bacterianos en agua destilada estéril (en proporción 1:1), ajustada a diferentes pH (2, 4, 6, 7 y 8) y se mantuvieron en estas condiciones durante 20 minutos. Posteriormente, se mezclaron con la suspensión micelial de M. fijiensis. Se incubaron y evaluaron según lo descrito en el acápite 3.3.2. Se realizaron cuatro réplicas por tratamiento.. 3.3.2.3 Efecto de detergentes sobre la actividad antifúngica de los filtrados de cultivo bacterianos Para determinar el efecto de los detergentes sobre la actividad antifúngica de los filtrados de cultivo bacterianos se tomaron 100 µL de filtrado estéril y se les añadieron 4 µL de detergente, para una concentración final de 1%. Los detergentes utilizados fueron: el aniónico SDS (del inglés: Sodium Dodecyl Sulphate),. el catiónico CTAB (del inglés:. Hexadecyltrimethyl Ammonium Bromide), además de los no iónicos Triton x 100 y Tween-80. Las mezclas se incubaron a 28ºC y oscuridad, durante cuatro horas. Pasado este tiempo, se mezclaron con la suspensión micelial de M. fijiensis. Se incubaron y evaluaron según lo descrito en el acápite 3.3.2. Se realizaron tres réplicas por tratamiento.. 28.
(39) Materiales y Métodos 3.3.2.4 Efecto de proteasas sobre la actividad antifúngica de los filtrados de cultivo bacterianos Para determinar el efecto de proteasas sobre la actividad antifúngica, se tomaron 950 µL de filtrado de cultivo estéril, proveniente de cada una de las cepas bacterianas, y se les añadió 50 µL de proteinasa K de Engyodontium álbum (Sigma) (100 µg/mL). Las muestras se colocaron a 37ºC, durante dos horas. Finalmente, se mezclaron con la suspensión micelial de M. fijiensis. Se incubaron y evaluaron según lo descrito en el acápite 3.3.2. Además, se incluyeron los filtrados de cultivo sin presencia de la proteasa. Se realizaron ocho réplicas por tratamiento.. 3.3.3 Caracterización de metabolitos de naturaleza proteica con actividad antifúngica in Vitro frente a M. fijiensis Con el objetivo de obtener metabolitos de la fase proteica de los filtrados, con actividad antifúngica caracterizados, se realizaron los siguientes experimentos.. 3.3.3.1 Semipurificación de proteínas con sulfato de amonio Para obtener los metabolitos de naturaleza proteica semipuricados, se obtuvo el filtrado de cultivo esterilizado, para cada una de las cepas bacterianas en estudio, de igual forma al acápite 3.2.1. Este fue sometido a una precipitación con sulfato de amonio (70%) durante 24 horas en agitación a 4ºC, posteriormente fueron centrifugados a 4ºC por 15 min a 10 000 rpm (Eppendorf Centrifuge 5810 R). El sobrenandante se desechó y los precipitados fueron resuspendidos en 2 mL de Solución Amortiguadora (SA) fosfato de sodio (0,01 M, pH=7) (Sigma-Aldrich) y dializados extensivamente mediante membranas de diálisis de 12 kDa contra la misma SA. Se chequeó la actividad antifúngica en todas las etapas de purificación de los metabolitos (filtrado de cultivo y las muestras dializadas) mediante el método de descrito en el acápite 3.3.2.. 29.
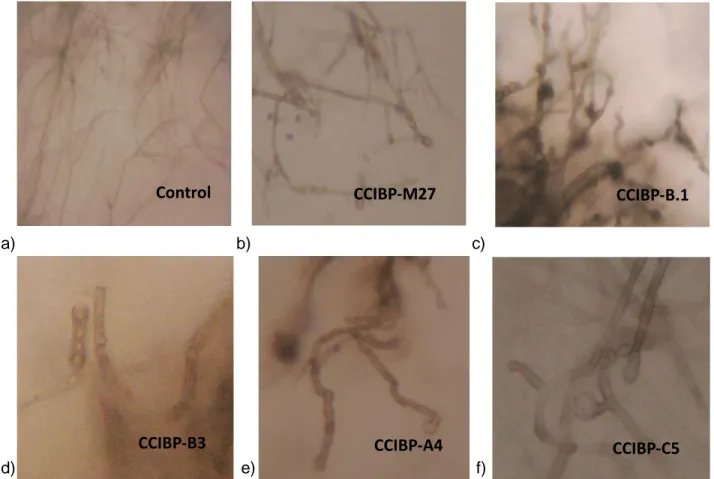
(40) Materiales y Métodos Además, se realizó observación microscópica de las hifas del patógeno incubadas conjuntamente con los metabolitos semipurificados, para describir sus características.. 3.3.3.2 Cuantificación de proteínas totales Las proteínas totales semipurificadas fueron cuantificadas mediante el método de Bradford (1976), respecto a una curva patrón a partir de concentraciones conocidas de BSA (del inglés: Bovine Serum Albumin), utilizando el reactivo Coomassie Plus-Bradfod Assay (Thermo Scientific). Los resultados se expresaron en µg/ml de proteína. Se emplearon tres réplicas por cada fracción de metabolitos semipurificados y se repitió dos veces. Los valores obtenidos fueron analizados estadísticamente mediante las pruebas de Kruskal-Wallis y Mann Whitney, previa comprobación de los supuestos de normalidad y heterogeneidad de varianza.. 3.3.3.3 Determinación del peso molecular Se realizó una electroforesis de proteínas bajo condiciones desnaturalizantes en gel de poliacrilamida (12%) (SDS-PAGE) de acuerdo con el procedimiento propuesto por Laemmli y Favre (1973). Después de la electroforesis, el gel fue teñido con el reactivo Coomassie brilliant blue (Spectrum). Para determinar el peso molecular de los metabolitos de naturaleza proteica extraídos de los filtrados bacterianos se compararon las bandas de la corrida electroforética con el marcador de peso molecular de proteínas proporcionado por FERMENTA (11-170 kDa). Los resultados se muestran en las bandas obtenidas en la corrida electroforética.. 3.3.3.4 Determinación de la actividad quitinasa en geles de agarosa Con el objetivo de caracterizar las proteínas presentes en las fracciones proteicas de los diferentes filtrados bacterianos se determinó la actividad quitinasa. Se utilizó el método. 30.
Figure

+7

Documento similar
que hasta que llegue el tiempo en que su regia planta ; | pise el hispano suelo... que hasta que el
dente: algunas decían que doña Leonor, "con muy grand rescelo e miedo que avía del rey don Pedro que nueva- mente regnaba, e de la reyna doña María, su madre del dicho rey,
Entre nosotros anda un escritor de cosas de filología, paisano de Costa, que no deja de tener ingenio y garbo; pero cuyas obras tienen de todo menos de ciencia, y aun
E Clamades andaua sienpre sobre el caua- 11o de madera, y en poco tienpo fue tan lexos, que el no sabia en donde estaña; pero el tomo muy gran esfuergo en si, y pensó yendo assi
Luis Miguel Utrera Navarrete ha presentado la relación de Bienes y Actividades siguientes para la legislatura de 2015-2019, según constan inscritos en el
Fuente de emisión secundaria que afecta a la estación: Combustión en sector residencial y comercial Distancia a la primera vía de tráfico: 3 metros (15 m de ancho)..
La campaña ha consistido en la revisión del etiquetado e instrucciones de uso de todos los ter- mómetros digitales comunicados, así como de la documentación técnica adicional de
This section provides guidance with examples on encoding medicinal product packaging information, together with the relationship between Pack Size, Package Item (container)
