M’atorgo a mi mateix el privilegi de descomptar el que no tinc del que tinc.
Així el que em quedi, ni que sigui poc, serà el nervi d’allò que he perseguit durant molts anys amb més força que traça.
Algú potser dirà que estigmatitzo, però jo invocaré codis antics i el valor oblidat de la paraula que mai no he transgredit volgudament,
i amb gest d’aparent desimboltura saltaré, sense moure’m, més ençà dels límits que m’imposin. Aleshores, si em criden respondré, però no crec que cap veu m’emocioni ni m’alteri.
Potser serà un regal immerescut, però goso esperar que ningú es queixi.
Més convincent que convençut, faré, de franc si cal, tots els papers de l’auca,
sense deixar amablement de somriure, per fer-me perdonar pecats i deutes.
Un cop arribat aquest moment és complicat escriure i fer memòria de tota la gent que m’ha ajudat al llarg d’aquests 4 anys. Per tant si m’oblido d’algú espero que no s’enfadi.
Primer de tot voldria agrair al meu tutor i director de tesi, al Dr. Pere Garriga per la oportunitat que m’ha donat per dur a terme aquest treball. Els seus consells per treballar en rodopsina han estat realment molt útils encara que a vegades seguís fent la meva. També gràcies per la seva paciència en els errors comesos senzillament per estupideses i per l’oportunitat que m’ha donat per ampliar els meus coneixements i les meves expectatives.
També voldria agrair a la Laia la seva perspectiva de veure les coses i per ser l’alegria del lab, i la paciència d’aguantar el meu caràcter el primer any així com la Laura, en Dasiel i als nou vinguts Darwyn i Wilber pel seu suport moral en la realització d’aquesta tesi. Som pocs però ben avinguts...
En aquesta llista no hi pot faltar tampoc el Dr. Luís del Valle, Lucho, pels consells “biològics” en el lab o via telèfon i el seu punt de vista de determinats aspectes que una principiant no veu. També em va ajudar a entrar en el mon de la rodopsina des de el punt de vista pràctic, manipulació de les cèl·lules, purificació dels DNA, etc.... Sense ell el lab no seria el que és. I que consti que no és “peloteo”.
També dono les gràcies al Dr. Joan Manyosa i la Dra. Anna Andrés de la UAB per permetre’ns anar al seu laboratori i ajudar a fer part d’alguns experiments, especialment la purificació d’alguns dels mutants que es presenten en aquesta tesi. També dono les gràcies als membres de la unitat de Biofísica del departament de Bioquímica i Biologia Molecular de la UAB per la seva hospitalitat i ajuda tècnica en qualsevol dels moments en que els necessitàvem (que no són pocs!!!).
En el nostre departament d’Enginyeria Química de la UPC, he trobat l’ajuda de la Dra. Núria Saperas en la construcció dels mutants, i també de l’Arnau Cordomí per les simulacions i modelització d’alguns dels mutants d’aquesta tesi i que esperem que acabin dins un
paper
.
científiques. I també a la Sandra i la Nadia pel seu suport tots aquests anys.
D’altra banda vull agraïr als meus pares, sense els quals aquesta tesi no hagués pogut sortir i per la seva ajuda incondicional sense saber gairebé que feia. A partir d’ara em veuran més sovint i
“no necessitaran una foto per
recordar la meva cara”
. També als meus germans Gemma, Lluís i Dani, tres persones i tres mons diferents. Gràcies mama pel poema de Martí i Pol.Finalment voldria agrair el suport incondicional, logístic, moral i informàtic de la persona que ha estat tots aquest anys recolzant-me. De fet, molts dels dibuixos d’aquesta tesi no s’haguessin realitzat sense la seva paciència. Gràcies Joan.
Per últim haig d’agraïr el suport econòmic a través d’un projecte de recerca finançat per la Direcció General de la ONCE.
Annex 1: Medis Bacterians
1.1.-Medi 2YT líquid (per 1 litre)
Aquest medi juntament amb el terrific broth es caracteritzen pel seu elevat contingut en triptona i extracte de llevat proporcionen un elevat creixement dels cultius bacterians.
1- Fer la barreja dels següents components: 16 g Triptona (Pronadisa)
10 g Extracte de Llevat (Pronadisa) 5 g NaCl (Panreac)
2-Es resuspèn tot en 800 ml d’aigua destil·lada.
3-S’ajusta a pH 7.0-7.4 amb NaOH 5N i s’enrasa a 1000 ml amb aigua destil·lada.
4-S’autoclava (30 min a 120ºC) i es guarda a temperatura ambient.
1.2.-Medi 2YT sòlid + 100 ug/ml ampicil·lina (per 500 ml)
1-Es barregen els següents components en 300 ml d’aigua destil·lada en un erlenmeier:
8 g Triptona (Pronadisa)
5 g Extracte de Llevat (Pronadisa) 5 g Agar Bacteriològic (Pronadisa) 2.5 g NaCl (Panreac)
2-S’ajusta a pH 7.0-7.4 i s’enrasa a 500 ml amb aigua destil·lada. Es tapa l’erlenmeier amb paper de plata.
3-S’autoclava (30 min a 120ºC) i es deixa refredar però sense solidificar (és a dir, fins que es pugui aguantar l’erlenmeier en el palmell de la mà). Quan cal refredar ràpidament el medi, es posa sota l’aixeta d’aigua freda agitant l’erlenmeier per a que el refredament sigui homogeni.
4-S’afegeixen 500 µl d’ampicil·lina (USB) 100mg/ml, es barreja suaument i es reparteix en plaques (15-20 ml/placa).
Annex 2: Medis eucariotes
2.1.-El sèrum fetal boví
El SFB (Gibco) ha estat obtingut a partir de la sang extreta d’una punció cardíaca de fetus boví. Posteriorment, es separa el sèrum respecte el coàgul i es congela. Tot el procediment es realitza a temperatures refrigerades. El sèrum utilitzat en els experiments descrits està lliure de virus i micoplasma. Per la presència en el sèrum dels factors complement, prèviament s’ha d’inactivar mitjançant un tractament tèrmic a 56ºC durant 30 min. Després s’aliquota en tubs de 50 ml en 50 ml i en guarden a -20ºC fins a la seva utilització.
2.2.-La tripsina-EDTA
La tripsina és un dels tres enzims proteolítics digestius i actua trencant les proteïnes de la dieta en pèptids i aminoàcids i en l’organisme és produït pel pàncrees. Presenta histidines i serines en el centre catalític i digereix pels enllaços peptídics d’aminoàcids que contenen arginina i lisina. La tripsina també s’utilitza per a la determinació de la seqüència de determinades proteïnes. En els nostres experiments és utilitzada per a desenganxar les cèl·lules de la capa adherent de la placa de cultiu. La casa comercial (BioMedia) ens el proporciona amb EDTA, un agent quelant per evitar la inactivació de l’enzim per efecte de determinats ions. Es troba concentrat 10 X en ampolles de 100 ml, es reparteixen de 10 ml en 10 ml i cada vegada i es guarda a -20ºC fins a la seva utilització. Per la seva utilització, es dilueix amb PBS.
2.3.-La penicil·lina + estreptomicina + glutamina
2.4.-El DEAE-Dextra
El DEAE-Dextrà és un compost format per DEAE un compost policatiònic derivat del dextrà i el dextrà pròpiament dit en una relació 1:3 respectivament. Quan es troba en forma sòlida és molt estable i la seva efectivitat pot durar més de cinc anys. En solució a temperatura ambient també és estable en un rang de pH de 4 a 10, per fora d’aquest rang de pH i a temperatures elevades el dextrà es podria despolimeritzar.
El compost pot tenir moltes funcions, una d’elles és la transfecció transitòria de gen clonats en cèl·lules de mamífer des de fa 40 anys (Vaheri i Pagano, 1965) . A partir d’aquell moment s’han realitzat diverses millores en el procés de transfecció i s’han arribat a definir les condicions òptimes ambientals de les cèl·lules i la composició del medi (Gonzalez i Joly, 1995).
La preparació del DEAE-Dextrà (2.5mg/ml), es realitza per a 100 ml:
1- Es pesen 0.250 g de DEAE-Dextrà (Sigma)
2-S’enrasa a 100 ml amb 90 ml DEMEM + 1 ml Strp+Gln+Pen + 10 ml FCS 3-S’agita amb vigorositat per tal de desfer la pasta que forma el DEAE en aquesta solució i posteriorment es filtra en una xeringa connectada a un filtre de 0.22 µm de porus (Millipore).
4-Es guarda a 4ºC fins a la seva utilització.
2.5.-La cloroquina
llum. En la transfecció, la chlo s’ha relacionat amb la millora de l’efectivitat de la transfecció, tot i que no se sap quina és la seva funció concreta.
La preparació de la chlo (1 mM) en 100 ml:
1-Es pesen 0.052 g de chlo en un vas de precipitat protegit de la llum amb paper de plata.
2-S’afegeixen 60 ml d’aigua milliQ i s’ajusta el pH=7.2 amb NaHCO3 en pols. 3-S’acaba d’enrasar amb aigua milliQ fins als 100 ml.
Annex 3: Encebadors utilitzats per a la construcció dels
mutants de rodopsina
3.1.-Oligonucleòtids sintètics utilitzats en la construcció dels mutants en la posició 94.
T94I1 5’-gAT CTC TTC Atg gTC TTC ggT ggC TTC ACC ACC ATC CTC TAC ACC TCT CTC-3’
T94I2 5’-C ATg gAg AgA ggT gTA gAg gAT ggT gAA gAA gCC gAA gAC CAT gAA gAC CAT gAA gA-3’
T94D1 5’-gAT CTC TTC Atg gTC TTC ggT ggC TTC ACC ACC GAT CTC TAC ACC TCT CTC-3’
T94D2 5’-C ATg gAg AgA ggT gTA gAg ATC ggT gAA gAA gCC gAA gAC CAT gAA gAC CAT gAA gA-3’
T94S1 5’-gAT CTC TTC Atg gTC TTC ggT ggC TTC ACC ACC TCT CTC TAC ACC TCT CTC-3’
T94S2 5’-C ATg gAg AgA ggT gTA gAg AgA ggT gAA gAA gCC gAA gAC CAT gAA gAC CAT gAA gA-3’
T94K1 5’-gAT CTC TTC Atg gTC TTC ggT ggC TTC ACC ACC AAg CTC TAC ACC TCT CTC-3’
T94K2 5’-C ATg gAg AgA ggT gTA gAg CTT ggT gAA gAA gCC gAA gAC CAT gAA gAC CAT gAA gA-3’
3.2.-Encebadors utilitzats en la construcció dels mutants E247A i E247Q.
E247Q1 5’-gCCCAgCAgCAggAgAgCgCCACCACTCAgAAggCCCAgAAggA ggTCA-3’
E247Q2 5’-CgCgTgACCTCCTTCTgggCCTTCTgAgTggTggCgCTCTCCTgC TgCTgggCTgCA-3’
E247A1 5’-gCCCAgCAgCAggAgAgCgCCACCACTCAgAAggCCgCgAAggA ggTCA-3’
3.3.-Encebadors utilitzats en la construcció del mutant L46R
L46R1 5’-CgCCTACATgTTCCggCTgATCATgCTTgg-3’ L46R2 5’-CCAAgCATgATCAgCCggAACATgTAggCg-3’
3.4.-Encebadors utilitzats en la construcció dels mutants T251A, T251E i T251K simples, dobles i triples mutants.
Annex 4. Gels d’agarosa analítics i preparatius.
Un cop formats els fragments de DNA digerit o obtingut el DNA d’una purificació a partir de cèl·lules bacterianes es poden visualitzar en un gel d’agarosa. L’agarosa en els gels es forma per què les cadenes del polímer s’uneixen entre si formant una xarxa rígida tridimensional de superfibres helicoidals que li confereixen rigidesa al gel. La mida dels porus del gel és relativament gran i permet el pas del DNA, lipoproteïnes i partícules víriques. És possible formar gels útils per electroforesi utilitzant quantitats d’agarosa compreses entre 0.4% fins 5% d’agarosa. Concentracions superiors no són recomanables ja que els gels resulten poc elàstics i difícilment manejables i llavors es substitueixen pels PAGE.
El gel que utilitzem és de baixa electroendosmosi (EEO). Aquest fenomen consisteix en que l’agarosa presenta determinats grups aniònics estabilitzats per càrregues positives, aquestes càrregues es troben solvatades per molècules d’aigua. En presència d’un corrent elèctric, les càrregues positives corren cap al càtode, enduent-se les molècules d’aigua amb elles, això provoca que es distorsioni tant el moviment de les mostres com l’estructura del propi gel. En el nostre cas, l’agarosa està modificada amb la presència de substitucions que fan abaixar el grau d’EEO.
El bromur d'etidi és un colorant vermell fluorescent carcinogen de la família de les sals de fenantridini. També se l’anomena bromur de 3,8-diamino-5-etil-6-fenilfenantridini o EtBr, la seva fórmula molecular és C21H20N3Br i el seu pes molecular és de 394.3. És un agent intercal·lant de les bases nitrogenades del DNA, el que permet detectar-lo en gel d’agarosa quan s’irradia amb llum UV.
Material i equipament utilitzat
1-Agarosa D-1 Baja EEO (Pronadisa), EtBr (Sigma; 10 mg/ml), el marcador de pesos moleculars λ DNA-HindIII digest (Pharmacia) i Kb DNA Ladder (Stratagene).
tampó de càrrega (6x) (en 5 ml conté 3.5 ml TAE 1X, 1.5 ml de glicerol (Sigma), 0.25% de blau de bromofenol (Sigma) i s’aliquota de 1ml en 1 ml i es guarda a -20ºC).
3-Els aparells d’electroforesi utilitzats són Sub-Cell GT mini i Sub-Cell GT wide mini (BioRad), la càmara fotogràfica Kodak DC290 Zoom Digital CameraTM i pel tractament dels gels s’utilitza el programa Quantity One (BioRad).
Procediment experimental
1-Es pesen 0.5 g d’agarosa i es resuspenen en 50 ml de TAE1X.
2-Es dissol l’agarosa durant 2 minuts al microones. Es deixar refredar (sense solidificar) i s’afegeix el EtBr (a una concentració final de 0.5 µg/ml).
3-Abans que es solidifiqui, es passa l’agarosa en la plantilla del aparell d’electroforesi que te la pinta corresponent en funció de les mostres que es vulguin carregar i es deixa solidificar.
4-A cada mostra s’afegeix la quantitat necessària de tampó de càrrega (6x) i es carreguen en el gel.
5-El gel es resol a 75V i 500 mA.
Abdulaev N. G. (2003) Building a stage for interhelical play in rhodopsin.
Trends Biochem. Sci. 28; 399-402.
Alberts B., Bray P., Lewis J., Raff M., Roberts R. i Watson J. D. Molecular Biology of the cell (1994). 3th edition. Garland Publishing, Inc.
Alewijnse A. E., Timmerman H., Jacobs E. H., Smit M. J., Roovers E., Cotecchia S. i Leurs R. (2000) The effect of mutations in the DRY motif on the constitutive activity and structural instability of the histamine H(2) receptor. Mol. Pharmacol.57; 890-898.
al-Jandal N., Farrar G. J., Kiang A. S., Humphries M. M., Bannon N., Findlay J. B., Humphries P. i Kenna P. F. (1999) A novel mutation within the rhodopsin gene (Thr-94-Ile) causing autosomal dominant congenital stationary night blindness. Hum. Mutat. 13; 75-81.
Andres A., Garriga P. i Manyosa J. (2003) Altered functionality in rhodopsin point mutants associated with retinitis pigmentosa. Biochem. Biophys. Res. Commun. 303; 294-301.
Anzenbacherova E., Bec N., Anzenbacher P., Hudecek J., Soucek P., Jung C., Munro A. W. i Lange R. (2000) Flexibility and stability of the structure of cytochromes P450 3A4 and BM-3. Eur. J. Biochem.267; 2916-2920.
Arnis S., Fahmy K., Hofmann K. P. i Sakmar, T. P. (1994) A conserved carboxylic acid group mediates light-dependent proton uptake and signaling by rhodopsin. J. Biol. Chem.269; 23879-23881.
Arora K. K., Cheng Z. i Catt K. J. (1997) Mutations of the conserved DRS motif in the second intracellular loop of the gonadotropin-releasing hormone receptor affect expression, activation, and internalization. Mol. Endocrinol.11; 203-212.
Ashwood-Smith M. J. (1967) Current concept concerning radioprotective and cryoprotective properties of dimethyl sulfoxide in cel·lular systems. Ann. N. Y. Acad. Sci. USA141;41-62.
Atwood C. S., Moir R. D. Huang X., Scarpa R. C., Bacarra N. M., Romano D. M., Hartshorn M. A., Tanzi R. E. i Bush A. I. (1998) Dramatic aggregation of Alzheimer Aβ by Cu(II) is induced by conditions representing physiological
acidosis. J. Biol. Chem.273; 12817-18226.
Ballesteros J. A. i Weinstein H. (1995) Integrated Methods for the Construction of Three-Dimensional Models and Computational Probing of Structure-Function Relations in G Protein-Coupled Receptors. Methods Neuroscience25;
366-428.
Ballesteros J. A., Jensen A. D., Liapakis G., Rasmussen S. G., Shi L., Gether U. i Javitch J. A. (2001) Activation of the beta 2-adrenergic receptor involves disruption of an ionic lock between the cytoplasmic ends of transmembrane segments 3 and 6. J. Biol. Chem.276; 29171-29177.
Berson, E. L. (2000) Nutrition and retinal degenerations. Int. Ophthalmol. Clin.
40; 93-111.
Boeckaert J. i Pen J. P. (1999) Molecular thinckering of G protein-coupled receptors: an evolutionary success. The EMBO Journal 18; 1723-1729.
Bosch L., Ramon E., Del Valle L. J. i Garriga P. (2003) Structural and functional role of helices I and II in rhodopsin. A novel interplay evidenced by mutations at Gly-51 and Gly-89 in the transmembrane domain. J. Biol. Chem. 278;
20203-20209.
Breit A., Lagace M. i Bouvier M. (2004) Hetero-oligomerization between beta2- and beta3-adrenergic receptors generates a beta-adrenergic signaling unit with distinct functional properties. J. Biol. Chem.279; 28756-28765.
Bubis J. (1998) Effect of detergents and lipids on transducin photoactivation by rhodopsin. Biol. Res.31; 59-71.
Buss V., Sugihara M., Entel P. i Hafner J. (2003) Thr94 and Wat2b effect protonation of the retinal chromophore in rhodopsin. Angew. Chem. Int. Ed Engl. 42; 3245-3247.
Cha K., Reeves P. J. i Khorana H. G. (2000) Structure and function in rhodopsin: Destabilization of rhodopsin by the binding of an antibody at the N-terminal segment provides support for involvement of the latter in an intradiscal tertiary structure. Proc. Natl. Acad. Sci. USA97; 3016–3021.
Chen S., Lin F., Xu M. i Graham R. M. (2002) Phe(303) in TMVI of the α1b adrenergic receptor is a key residue coupling TM helical movements to G-protein activation. Biochemistry 41;588-596.
Coddou C., Morales B. i Huidobro-Toro J. P. (2003) Neuromodulator role of zinc and copper during prolonged ATP applications to P2X4 purinoceptors.
Eur. J. Pharmacol.472; 49-56.
Cohen S. N., Chang A. C. Y., i Hsu L. (1972) Nonchromosomal antibiotic resistance in bacteria: genetic transformation of Escherichia coli by R-factor DNA. Proc. Natl. Acad. Sci. USA69; 2110-2114 .
Cohen G. B., Yang T., Robinson P. R i Oprian D. D. (1993) Constitutive activation of opsin: influence of charge at position 134 and size at position 296.
Courageot M. P., Lepine S., Hours M., Giraud F. i Sulpice J. C. (2004) Involvement of sodium in early phosphatidylserine exposure and phospholipid scrambling induced by P2X7 purinoceptor activation in thymocytes. J. Biol. Chem.279; 21815-21823.
Creemers A. F., Kiihne S., Bovee-Geurts P. H., DeGrip W.J., Lugtenburg J. i de Groot H. J. (2002) (1)H and (13)C MAS NMR evidence for pronounced ligand-protein interactions involving the ionone ring of the retinylidene chromophore in rhodopsin. Proc. Natl. Acad. Sci. USA99; 9101-9106.
Cuatrecasas P., Wilcheck M. i Anfinsen C. D. (1968) Selective enzyme purification by affinity chromatography. Proc. Natl. Acad. Sci. USA 61;
636-643.
Davidson F. F., Loewen P. C. i Khorana H. G. (1994) Structure and function in rhodopsin: replacement by alanine of cysteine residues 110 and 187, components of a conserved disulfide bond in rhodopsin, affects the light-activated metarhodopsin II state. Proc. Natl. Acad. Sci. USA91; 4029-4033.
DeGrip, W. J. i Bovee-Geurts P. H. M. (1979) Synthesis and properties of alkylglucosides with mild detergent action: Improved synthesis and purification of Octyl, -Nonyl, and Decyl-Glucose. Synthesis of Undeglycose and 2-1-Dodecylmaltoside. Chemistry and Physics of Lipids 23; 321-335.
DeGrip W. J., Vanoostrum J. i Bovee-Geurts P. H. (1998) Selective detergent-extraction from mixed detergent/lipid/protein micelles, using cyclodextrin inclusion compounds: a novel generic approach for the preparation of proteoliposomes. Biochem. J.330; 667-674.
Del Valle L. J., Escribano C., Perez J. J. i Garriga P. (2002) Calcium-induced
decrease of the thermal stability and chaperone activity of alpha-crystallin.
Biochim. Biophys. Acta.1601; 100-109.
Doi T., Molday R. S. i Khorana H. G. (1990) Role of the intradiscal domain in rhodopsin assembly and function. Proc. Natl. Acad. Sci. USA87; 4991-4995.
Dryja T. P., Berson E. L., Rao V. R. i Oprian D. D. (1993) Hetereogeneous missense in the rhodopsin gene as a cause of Congenital Stationary Night Blindness. Nature Genetics 4; 280-282.
Duñach M., Sabés M. i Padrós E. (1983) Fourth derivative spectrophotometry analysis in tryptophan environment in proteins. Biochemistry 134;
17448-17457.
Eason M. G. i Ligget S. B. (1996) Chimeric Mutagenesis af putative G-protein Coupling Domains of the α2-adrenèrgic receptors. J. Biol. Chem. 271;
Fahmy K. i Sakmar T. P. (1993) Regulation of the rhodopsin-transducin interaction by a highly conserved carboxylic acid group. Biochemistry32;
7229-7236.
Fahmy K., Siebert F. i Sakmar T. P. (1995) Photoactivated state of rhodopsin and how it can form. Biophys. Chem.56; 171-181.
Fahmy K., Sakmar T. P i Siebert F. (2000) Transducin-dependent protonation of glutamic acid 134 in rhodopsin. Biochemistry39; 10607-10612.
Farrar G. J., Kennan P. F. i Humphries P. (2002) On the genetics of Retinitis Pigmentosa and on mutation-independent approaches to therapeutic intervention. The EMBO Journal 21; 857-864.
Farrens D. L. i Khorana H. G. (1995) Structure and function in rhodopsin. Measurement of the rate of Metarhodopsin II decay by fluorescence spectroscopy. J. Biol. Chem.270; 5073-5076.
Farrens D. L., Altenbach C., Yang K., Hubbell W. i Khorana H. G. (1996) Requeriment of Rigid-Body Motion of Transmembrane helices for light activation of rhodopsin. Science276; 768-770.
Feng W. i Song Z. H. (2003) Effects of D3.49A, R3.50A, and A6.34E mutations on ligand binding and activation of the cannabinoid-2 (CB2) receptor. Biochem. Pharmacol.65; 1077-1085.
Ferretti L., Karnik S. S., Khorana H. G., Nassal M. i Oprian D. D. (1986) Total synthesis of a gene for bovine rhodopsin. Proc. Natl. Acad. Sci. USA 83;
599-603.
Filipek S., Krzysko K. A., Fotiadis D., Liang Y., Saperstein D. A., Engel A. i Palczewski K. (2004) A concept for G protein activation by G protein-coupled receptor dimers: the transducin/rhodopsin interface. Photochem. Photobiol. Sci. 3, 628-638.
Fotiadis D., Liang Y., Filipek S., Saperstein D. A., Engel A. i Palczewski K. (2003) Rhodopsin dimers in native disc membranes. Nature 421; 127-128.
Francesconi A. i Duvoisin R. M. (2004) Divalent cations modulate the activity of metabotropic glutamate receptors. Neurosci. Res.75; 472-479.
Franke R. R., Sakmar T. P., Oprian D. D. i Khorana H. G. (1988) A single amino acid substitution in Rhodopsin (Lysine 248Æ Leucine) prevents activation of transducin. J. Biol. Chem. 268; 2119-2122.
Franke R. R., König B., Sakmar T. P. Khorana H. G. i Hofmann K. P. (1990) Rhodopsin mutants that bind but fail to active transducin. Science 250;
123-125.
Fraser C. M., Chung F. Z., Wang C. D. i Venter J. C. (1988) Site-Directed Mutagenesis of Human β-adrenergic receptors. Substitution of Aspartic acid 130 by Asparagina produces a receptor with affinity agonist binding that in unclouped from adenilate cyclasa. Prog. Natl. Acad. Sci. USA 85; 5478-5482.
Futterman S. i Rollins M. H. (1973) The catalytic isomerization of all-trans-retinal to 9-cis-all-trans-retinal and 13-cis-all-trans-retinal. J. Biol. Chem. 248; 7773-7779.
Garcia-Segura J. M., Gavilanes J. G., Martínez del Pozo A., Montero F., Oñaderra M. i Vivanco F. (1996) Emisión de fluorescencia. En Técnicas instrumentales de análisis bioquímica. Proyecto Editorial Ciencias Químicas.
pp 120-166.
Garriga P., Liu X. i Khorana H. G. (1996) Structure and function in rhodopsin: correct folding and misfolding in point mutants at and in proximity to the site of the retinitis pigmentosa mutation Leu-125-->Arg in the transmembrane helix C.
Proc. Natl. Acad. Sci. USA93; 4560-4564.
Garwin G. G. i Saari J. C. (2000) High-performance liquid chromatography analysis of visual cycle retinoids. Methods Enzymol.316; 313-324.
Gerlach L. O., Jakobsen J. S., Jensen K. P., Rosenkilde M. R., Skerlj R. T., Ryde U., Bridger G.J. i Schwartz T.W. (2003) Metal ion enhanced binding of AMD3100 to Asp262 in the CXCR4 receptor. Biochemistry42; 710-717.
Gether U., Lin S., Ghanouni P., Ballesteros J. A., Weinstein H. i Kobilka B. K. (1997) Agonists induce conformational changes in transmembrane domains III and VI of the beta2 adrenoceptor. EMBO J.16; 6737-6747.
Gonzalez A. L. i Joly E. (1995) A simple procedure to increase efficiency of DEAE-dextran transfection of COS cells. Trends Genet.11; 216-217.
Grahn B. H., Paterson P. G., Gottschall-Pass K..T i Zhang Z. (2001) Zinc and the eye. J. Am. Coll. Nutr.20; 106-118.
Greasley P. J, Fanelli F., Rossier O., Abuin L. i Cotecchia S. (2002) Mutagenesis and modelling of the alpha(1b)-adrenergic receptor highlight the role of the helix 3/helix 6 interface in receptor activation. Mol. Pharmacol.61; 1025-1032.
Gross A. K., Rao V. R. i Oprian D. D. (2003) Characterization of rhodopsin congenital night blindness mutant T94I. Biochemistry 42; 2009-2015.
Guzman, Y. (1981) Transformed simian cells support the replication of Early SV40 mutants. Cell23; 175-182.
Han M., Smith S. O. i Sakmar T. P. (1998) Constitutive activation of opsin by mutation of methionine 257 on transmembrane helix 6. Biochemistry37;
8253-8261.
Han M. i Sakmar T. P. (2000) Assays for activation of recombinant expressed opsins by all-trans-retinals. Methods Enzymol.315; 251-267.
Hardgrave P. A. (2001) Rhodopsin Structure, Function and Topography. The FriedenWald Lecture. Invest. Ophthalmol. Vis. Sci.42; 3-9.
Heck M., Schadel S. A., Maretzki D., Bartl F. J., Ritter E., Palczewski K. i Hofmann H. P. (2003) Signalling states of rhodopsin. Formation of the storage form, metarhodopsin III, from active metarhodopsin II. J. Biol. Chem. 278;
3163-3169.
Heck M., Schadel S. A., Maretzki D. i Hofmann K. P. (2003b) Secondary binding sites of retinoids in opsin: characterization and role in regeneration.
Vision Res.43; 3003-3010.
Heckenlively J. R., Boughman J. A. i Friedman L. H. (1988) Pedigree analisis. In
Heckenlively JR. Retinitis Pigmentosa. Philadelphia PA, Lippincott, pp 6-24. Herbert T. E., Moffett S., Morello J. P., Loise T.P., Bichet D.G., Barret C., i Bouvier M. (1996) A Peptide from a β2-Adrenèrgic Receptor Transmembrane
Domain Inhibits Both Receptor Dimeritzation and Activation. J. Biol. Chem.
271; 16384-16392.
Hessel E., Muller P., Herrmann A. i Hofmann K. P. (2001) Light-induced reorganization of phospholipids in rod disc membranes. J. Biol. Chem. 276;
2538-2543.
Hogger P., Shockley M. S., Lameh J. i Sadee W. (1995) Activating and inactivating mutations in N- and C-terminal i3 loop junctions of muscarinic acetylcholine Hm1 receptors. J. Biol. Chem.270; 7405-7410.
Holst B. i Schwartz T. W. (2003) Molecular mechanism of agonism and inverse agonism in the melanocortin receptors: Zn(2+) as a structural and functional probe. Ann. N. Y. Acad. Sci.994; 1-11.
Hong K., Knudsen P. J. i Hubbell W. L. (1982) Purification of rhodopsin on hydroxyapatite columns, detergent exchange, and recombination with phospholipids. Methods Enzymol.81; 144-150.
Huang P., Visiers I., Weinstein H. i Liu-Chen L. Y. (2002) The local environment at the cytoplasmic end of TM6 of the mu opioid receptor differs from those of rhodopsin and monoamine receptors: introduction of an ionic lock between the cytoplasmic ends of helices 3 and 6 by a L6.30(275)E mutation inactivates the mu opioid receptor and reduces the constitutive activity of its T6.34(279)K mutant. Biochemistry41, 11972-11980.
Humphries M. M., Rancourt D., Farrar G. J., Kenna P., Hazel M., Bush R. A., Sieving P. A., Sheils D. M., McNally N., Creighton P., Erven A., Boros A., Gulya K., Capecchi M. R. i Humphries P. (1997) Retinopathy induced in mice by targeted disruption of the rhodopsin gene. Nat. Genet.15, 216-219.
Hwa J., Garriga P., Liu X. i Khorana H. G. (1997) Structure and function in rhodopsin. Packing of the helices in the transmembrane domain and folding to a tertiary structure in the intradiscal domain are coupled. Proc. Natl. Acad. Sci. USA 94; 10571-10576.
Jäger S., Palczewski K. i Hofmann K. P. (1996) Opsin/all-trans-retinal complex activates transducin by different mechanisms than photolyzed rhodopsin.
Biochemistry35; 2901-2908.
Jansen J. J., Mulder W. R., De Caluwe G. L., Vlak J. M. i DeGrip W. J. (1991) In vitro expression of bovine opsin using recombinant baculovirus: the role of glutamic acid (134) in opsin biosynthesis and glycosylation. Biochim. Biophys. Acta1089; 68-76.
Janz J. M. i Farrens D. L. (2004) Role of retinal hydrogen bond network in rhodopsin schiff base stability and hydrolysis. J. Biol. Chem. Epub October 8; M408766200.
Jastrzebska B., Maeda T., Zhu L., Fotiadis D., Filipek S., Ángel A., Stenkamp R. E. i Palczewski K. (2004) Functional characterization of rhodopsin monomers ans dimmers in detergent. J. Biol. Chem. Epub 15 October; M408691200.
Javitch J. A., Fu D., Liapakis G. i Chen J. (1997) Constitutive activation of the beta2 adrenergic receptor alters the orientation of its sixth membrane-spanning segment. J. Biol. Chem. 272; 18546-18549.
Jensen A. D., Guarnieri F., Rasmussen S. G., Asmar F., Ballesteros J. A. i Gether U. (2001) Agonist-induced conformational changes at the cytoplasmic side of transmembrane segment 6 in the beta 2 adrenergic receptor mapped by site-selective fluorescent labeling. J. Biol. Chem.276; 9279-9290.
Jobling M. F., Huang X., Stewart L. R., Barnham K. J., Curtain C., Volitakis I., Perugini M., White A. R., Cherny R. A., Masters C. L., Barrow C. J., Collins S. J., Bush A. I. i Cappai R. (2001) Copper and zinc binding modulates the aggregation and neurotoxic properties of the prion peptide PrP106-126.
Kalloniatis M. i Fletcher E. L. (2004) Retinitis pigmentosa: understanding the clinical presentation, mechanisms and treatment options. Clin. Exp. Optom.87;
65-80.
Kaushal S. i Khorana H. G. (1994) Structure and function in rhodopsin. 7. Point mutations associated with autosomal dominant retinitis pigmentosa. Biochemistry 33; 6121-6128.
Kaushal S., Ridge K. D. i Khorana H. G. (1994) Structure and Function in Rhodopsin. The Role of Asparagine-linked glycosilation. Proc. Natl. Acad. Sci. USA 91; 4024-4028.
Kelleher D. J. i Johnson G. L. (1988) Transducin inhibition of light-dependent rhodopsin phosphorylation: evidence for beta gamma subunit interaction with rhodopsin. Mol. Pharmacol.34; 452-460.
Kim J. M., Altenbach C., Thurmond R. L., Khorana H. G. i Hubbell W. L. (1997) Structure and function in rhodopsin: rhodopsin mutants with a neutral amino acid at E134 have a partially activated conformation in the dark state. Proc. Natl. Acad. Sci. USA94; 14273-14278.
Kjelsberg M. A., Cotecchia S., Ostrowski J., Caron M. G. i Lefkowitz R. J. (1992) Constitutive activation of the alpha 1B-adrenergic receptor by all amino acid substitutions at a single site. Evidence for a region which constrains receptoractivation. J. Biol. Chem.267; 1430-1433.
Konig B., Arendt A., McDowell J. H., Kahlert M., Hardgrave P. A i Hoffmann K. P. (1989) Three Cytoplasmic loops of Rhodopsin interact with transducin. Proc. Natl. Acad, Sci. USA 86; 6878-6882.
Kosugi S., Mori T. i Shenker A. (1998) An anionic residue at position 564 is important for maintaining the inactive conformation of the human lutropin/choriogonadotropin receptor. Mol. Pharmacol.53; 894-901.
Kroeger K. M., Pfleger K. D. i Eidne K. A. (2003) G-protein coupled receptor oligomerization in neuroendocrine pathways. Front. Neuroendocrinol.24; 254-278.
Laemmli U. (1970) Cleavage of Structural Proteins during the Assemby of the Head of Bacteriophage T4. Nature227; 680-685.
Lagerstrom M. C., Jovins J., Fredriksson R., Fridmanis D., Haitina T., Ling M. K., Berglund M. M. i Schioth H. B. (2003) High affinity agonistic metal ion binding sites within the melanocortin 4 receptor illustrate conformational change of transmembrane region 3. J. Biol. Chem. 278; 51521-51526.
Leure-duPree A. E. (1981) Electron-Opaque inclusions in the rat retinal pigment epithelium after treatment with chelators of zinc. Invest. Ophthalmol. Vis. Sci.
21; 1-9.
Leure-duPree A. E. i McClain C. J. (1982) The effect of severe Zinc deficiency on the morphology of the rat retinal pigment ephitelium. Invest. Ophthalmol. Vis. Sci. 23; 425-434.
Li J., Huang P., Chen C., de Riel J. K., Weinstein H. i Liu-Chen L. Y. (2001) Constitutive activation of the mu opioid receptor by mutation of D3.49(164), but not D3.32(147): D3.49(164) is critical for stabilization of the inactive form of the receptor and for its expression. Biochemistry. 40; 12039-12050.
Liang Y., Fotiadis D., Filipek S., Saperstein D. A., Palczewski K. i Engel A. (2003) Organization of the G protein-coupled receptors rhodopsin and opsin in native membranes. J. Biol. Chem.278; 21655-21662.
Liang Y., Fotiadis D., Maeda T., Maeda A., Modzelewska A., Filipek S., Saperstein D.A., Engel A. i Palczewski K. (2004) Rhodopsin signaling and organization in heterozygote rhodopsin knockout mice. J. Biol. Chem. 279;
48189-48196.
Lin S. W. i Sakmar T. P. (1996) Specific tryptophan UV-absorbance changes are probes of the transition of rhodopsin to its active state. Biochemistry 35;
11149-11159.
Lu B., Kwan T., Kurimoto Y., Shatos M., Lund R. D. i Young M. J. (2002) Transplantation of EGF-responsive neurospheres from GFP transgenic mice into the eyes of rd mice. Brain Res.943, 292-300.
Luecke H., Schobert B., Richter H. T., Cartailler J. P. i Lanyi J. K. (1999) Structure of bacteriorhodopsin at 1.55 A resolution. J. Mol. Biol. 291; 899-911.
Mandel M. i Higa A. (1970) Calcium dependent bacteriophage DNA infection. J. Mol. Biol.53, 154-159.
Marin E. P., Krishna G., Zyaga T. A., Isele J., Siebert F. i Sakmar T.P. (2000) The amino terminus of the fourth cytoplasmic loop of rhodopsin modulates rhodopsin-transducin interaction. J. Biol. Chem.275; 1930-1936.
Marrón J. (1996) Aplicació de tècniques espectroscòpiques per a l’anàlisi funcional del fotorreceptor rodopsina i la caracterització estructural dels seus intermediaris. Tesina dirigida per Joan Manyosa i David Garcia-Quintana. Universitat Autònoma de Barcelona.
Molday R. S. i MacKenzie D. (1983) Monoclonal antibodies to rhodopsin: characterization, cross-reactivity, and application as structural probes.
Biochemistry 12; 4517-4523.
Molday R. S. (1998) Photoreceptor Membrane proteins, phototransduction, and retinal degenerative diseases. Invest. Opthlmol. & Vis. Sci.39; 2493-2513.
Nair K. S., Balasubramanian N. i Slepak V. Z. (2002) Signal-dependent translocation of transducin. RGS9-1-Gbeta complex, and arrestin to detergent-resistant membrane rafts photoreceptors. Curr. Biol.12; 421-425.
Naash M. I., Wu T. H., Chakraborty D., Fliesler S. J., Ding X. Q., Nour M., Peachey N. S., Lem J., Qtaishat N., Al-Ubaidi M. R. i Ripps H. (2004) Retinal abnormalities associated with the G90D mutation in opsin. J. Comp. Neurol.
478; 149-163.
Nathans J. i Hogness D. S. (1984) Isolation and nucleotide sequence of the gene encoding human rhodopsin. Proc. Nalt. Acad. Sci. USA 81; 4851-4855.
Nathans J. (1990) Determinants of visual pigment absorbance: identification of the retinylidene Schiff's base counterion in bovine rhodopsin. Biochemistry29; 9746-9752.
Nicolas C., Ghedira I., Stiemer R., Teufel B., Hecquet C., Faure J. P. i Mirshahi M. (2000) Identification of visual arrestin (S-antigen) in retinal pigmented epithelial cells. Curr. Eye Res.21; 677-683.
Noorwez S. M., Malhotra R., McDowell J. H., Smith K. A., Krebs M. P. i Kaushal S. (2004) Retinoids assist the cellular folding of the autosomal dominant retinitis pigmentosa opsin mutant P23H. J. Biol. Chem. 279; 16278-16284.
Oishi M. i Cosloy S. D. (1972) The genetic and biochemical basis of the transformability of Escherichia coli K12. Biochem. Biophys. Res. Commun.49;
1568-1572.
Okada T., Fujiyoshi Y., Silow M., Navarro J., Landau E. M i Shichida Y. (2002) Functional role of internal water molecules in rhodopsin revealed by X-ray crystallography. Proc. Natl. Acad. Sci. 99; 5982-5987.
Oprian D. D., Molday R. S., Kaufman R.J. i Khorana H.G. (1987) Expression of a synthetic bovine rhodopsin gene in monkey kidney cells. Proc. Natl. Acad. Sci. USA84, 8874-8878.
Padrós E., Duñach M., Morros A., Sabés M. i Mañosa J. (1982). The state of tyrosine and phenylalanine residues in proteins analyzed by fourth-derivative spectrophotometry. Histone H1 and ribonuclease A. Eur. J. Biochem.127; 117–
Palczewski K., Kumasaka T., Hori T., Behnke C. A., Motoshima H., Fox B. A., Le Trong I., Teller D. C., Okada T., Stenkamp R. E., Yamamoto M. i Miyano M.
(2000) Crystal structure of rhodopsin: A G protein-coupled receptor.
Science. 289; 739-745.
Papermaster D. S. i Dreyer W. J. (1974) Rhodopsin Content in the Outer Segment Memvranes of Bovine and Frog Retinal Rods. Biochemistry 13;
2438-2444.
Parma J., Duprez L., Van Sande J., Cochaux P., Gervy C., Mockel J., Dumont J. i Vassart G. (1993) Somatic mutations in the thyrotropin receptor gene cause hyperfunctioning thyroid adenomas. Nature365; 649-651.
Patel A. B., Crocker E., Eilers M., Hirshfeld A., Sheves M. i Smith S. O. (2004) Coupling of retinal isomerization to the activation of rhodopsin. Proc. Natl. Acad. Sci. USA.101; 10048-10053.
Pauwels P. J., Gouble A. i Wurch T. (1999) Activation of constitutive 5-hydroxytryptamine(1B) receptor by a series of mutations in the BBXXB motif: positioning of the third intracellular loop distal junction and its G(o)alpha protein interactions. Biochem. J.343; 435-442.
Polozova A. i Litman B. J. (2000) Cholesterol dependent recruitment of di22:6-PC by a G protein-coupled receptor into lateral domains. Biophys. J.79;
2632-2643.
Pucadyil T. J. i Chattopadhyay A. (2004) Cholesterol modulates ligand binding and G-protein coupling to serotonin(1A) receptors from bovine hippocampus.
Biochim Biophys Acta1663;188-200.
Ramon E., Marron J., del Valle J. L., Bosch L., Andres A., Manyosa J. i Garriga P. (2003) Effect of dodecil maltoside detergent on rhodopsin stability and function. Vision Research43; 3055-3061.
Rao R. V., Cohen G. B. i Oprian D. D. (1994) Rhodopsin mutation G90D and a molecular mechanism for congenital night blindness. Nature367; 639-642.
Rasmussen S. G., Jensen A. D., Liapakis G., Ghanouni P., Javitch J. A. i Gether U. (1999) Mutation of a highly conserved aspartic acid in the beta2 adrenergic receptor: constitutive activation, structural instability, and conformational rearrangement of transmembrane segment 6. Mol. Pharmacol. 56;175-184.
Ren Q., Kurose H., Lefkowitz R. J. i Cotecchia S. (1993) Constitutively active mutants of the alpha 2-adrenergic receptor. J. Biol. Chem. 268; 16483-16487.
Rodriguez J. A., Herrera C. A., Birch D. G. i Daiger S. P. (1993) A leucine to arginine amino acid substitution at codon 46 of rhodopsin is responsible for a severe form of autosomal dominant retinitis pigmentosa. Hum. Mutat. 2;
205-213.
Rosenberg I. M. (1996) Protein Analysis and Purification, Benchtop techniques; Massachusetts General Hospital.
Ruiz A., Borrego S., Marcos I. i Antiñolo G. (2001) Anatomia y Fisiología de la retina. En La Retinosis Pigmentaria en España. Coord. Diana Valverde. pp 11-38.
Sagdullaev B. T., Aramant R. B., Seiler M. J., Woch G. i McCall M. A. (2003) Retinal transplantion-induced recovery of retintectal visual funstion in a rodent model of retinitis pigmentosa. Invest. Ophthalmol. Vis. Sci. 44, 1686-1695.
Sakamoto T. i Khorana H. G. (1995) Structure and function in rhodopsin. The fate of opsin formed upon the decay of light-activated metarhodopsin II in vitro. Proc. Natl. Acad. Sci. USA92; 249-253.
Sakmar T. P., Franke R. R i Khorana H. G. (1989) Glutamic acid-113 serves as the retylidene Schiff Base Counterion in Bovine Rhodopsin. Proc. Natl. Acad. Sci. USA86; 8309-8313.
Sakmar T. P. (1998) Rhodopsin: A prototypical G Protein-Coupled Receptor.
Prog. Nucleic Acid Res. Mol. Biol. 59;1-34.
Salesse C., Boucher F. i Leblanc R. M. (1984) An evaluation of purity criteria for bovine rod outer segment membranes. Anal. Biochem.142; 258-266.
Samama P., Cotecchia S., Costa T. i Lefkowitz R. J. (1993) A mutation-induced activated state of the beta 2-adrenergic receptor. Extending the ternary complex model. J. Biol. Chem.268; 4625-4636.
Schadel S. A., Heck M., Maretzki D., Filipek S., Teller D. C., Palczewski K. i Hofmann K. P. (2003) Ligand channeling within a G-protein-coupled receptor. The entry and exit of retinals in native opsin. J. Biol. Chem.278; 24896-24903.
Scheer A., Fanelli F., Costa T., De Benedetti P. G. i Cotecchia S. (1996) Constitutively active mutants of the alpha 1B-adrenergic receptor: role of highly conserved polar amino acids in receptor activation.EMBO J.15; 3566-2578.
Schetz J. A. i Sibley D. R. (1997) Zinc allosterically modulates antagonist binding to cloned D1 and D2 dopamine receptors. J. Neurochem. 68;
1990-1997.
Schetz J. A. i Sibley D. R. (1999) Metal-ions as atomic scale probes of protein structure in Structure-function analysis of G-protein poupled receptors. Ed. J.
Shapiro D. A., Kristiansen K., Weiner D. M., Kroeze W. K. i Roth B. L. (2002) Evidence for a model of agonist-induced activation of 5-hydroxytryptamine 2A serotonin receptors that involves the disruption of a strong ionic interaction between helices 3 and 6. J. Biol. Chem. 277; 11441-11449.
Sheikh S. P., Zyaga T. A., Lichtarge O., Sakmar T. P. i Bourne H. R. (1996) Rhodopsin activation blocked by metal-ion-binding sites linking transmembrane helices C and F. Nature.383; 347-350.
Shi L., Liapakis G., Xu R., Guarnieri F. i Ballesteros J. A. (2002) Javitch JA. Beta2 adrenergic receptor activation. Modulation of the proline kink in transmembrane 6 by a rotamer toggle switch. J. Biol. Chem.277; 40989-40996.
Shuster T. A., Nagy A. K., Conly D. C. i Farber D. B. (1992) Direct zinc Binding to purified rhodopsin and discs membranes. Biochem. J. 282; 123-128.
Shuster T. A., Martin F. i Nagy A. K. (1996) Zinc causes an apparent increase in rhodopsin phosphorilation. Curr. Eye. Res. 15; 1019-1024.
Sieving P. A., Richards J. E., Naarendorf F., Bingham E. L., Scott K. i Alpern M. (1995) Dark-light: Model for nightblindness from de human rhodopsin Gly90— Asp mutations. Proc. Natl. Acad. Sci. USA 92; 880-884.
Smith W. C., Dinculescu A., Peterson J. J. i McDowell J. H. (2004) The surface of visual arrestin that binds to rhodopsin. Mol. Vis.10; 392-398.
Spooner P. J., Sharples J. M., Verhoeven M. A., Lugtenburg J., Glaubitz C. i Watts A. (2002) Relative orientation between the beta-ionone ring and the polyene chain for the chromophore of rhodopsin in native membranes.
Biochemistry.41; 7549-7555.
Stanasila L., Perez J. B., Vogel H. i Cotecchia S. (2003) Oligomerization of the alpha 1a-and alpha 1b-adrenergic receptor subtypes. Potential implications in receptor internalization. J. Biol. Chem. 278; 40239-40251.
Stojanovic A., Stitham J. i Hwa J. (2004) Critical role of transmembrane segment zinc binding in the structure and function of rhodopsin. J. Biol. Chem.
279; 35932-35941.
Strosberg A. D. (1991) Structure/Function Relationship of Protein Belonging to the family of Receptors Coupled to GTP-Binding Protein. Eur. J. Biochem. 196;
1-10.
Sugihara M., Buss V., Entel P., Elstner M. i Frauenheim T. (2002) 11-cis-retinal protonated Schiff base: influence of the protein environment on the geometry of the rhodopsin chromophore. Biochemistry. 41; 15259-15266.
Swaminath G., Lee T. W. i Kobilka B. (2003) Identification of an allosteric binding site for Zn2+ on the beta2 adrenergic receptor. J. Biol. Chem.278;
352-356.
Tam S. W., Wilber K. E. i Wagner F. W. (1976) Light Sensitive Zinc Content of protein fractions from Bovine Rod Outer Segments. Biochem. Biophys. Res. Commun. 72; 302-309.
Teller D. C., Okada T., Behnke C. A. i Palczewski K. (2001) Advances in Determination of a High Resolution Three-Dimensional Structure of Rhodopsin, a Model of g-protein-Coupled Receptors (GPCR’s). Biochemistry
40; 7761-7772.
Terakita A., Koyanagi M., Tsukamoto H., Yamashita T., Miyata, T. i Shichida Y. (2004) Counterion displacement in the molecular evolution of the rhodopsin family. Nat. Struct. Mol. Biol.11; 284-289.
Trumpp-Kallmeyer S., Chini B., Mouillac B., Barberis C., Hoflack J. i Hibert M. (1995) Towards understanding the role of the first extracellular loop for the binding of peptide hormones to G-protein coupled receptors. Pharm. Acta Helv.
70; 255-262.
Vaheri A. i Pagano J. S. (1965) Infectious poliovirus RNA: a sensitive method of assay. Virology27; 434-436.
Vogel R., Siebert F., Zhang X. Y., Fan G. i Sheves M. (2004). Formation of Meta III during the Decay of Activated Rhodopsin Proceeds via Meta I and Not via Meta II. Biochemistry43; 9457-9466.
Vorobjev V. S., Sharonova I. N., Sergeeva O. A. i Haas H. L. (2003) Modulation of ATP-induced currents by zinc in acutely isolated hypothalamic neurons of the rat. Br. J. Pharmacol.139; 919-926.
Wald G. i Brown P. K. (1953) The molar extinction of rhodopsin. J. Gen. Physiol.
37; 189-200.
Wessling-Resnick M. i Johnson G. L. (1987) Allosteric behaviour in transducin activation mediated by rhodopsin. Initial rate analysis of guanine nucleotide exchange. J. Biol. Chem.262; 3697-3705.
Wray W., Boulikas T., Wray V. P. i Hancock R. (1981) Silver staining of proteins in polyacrylamide gels. Anal. Biochem.118; 197-203.
Wu S. H., Qiao X., Noebels J. L. i Yang X. L. (1993) Localization and Modulatory actions of zinc in vertebrate retina. Vision Res. 33; 2611-2616.
Yan E. C., Kazmi M. A., De S., Chang B. S., Seibert C., Marin E. P., Mathies R. A. i Sakmar T. P. (2002) Function of extracellular loop 2 in rhodopsin: glutamic acid 181 modulates stability and absorption wavelength of metarhodopsin II.
Biochemistry41; 3620-3627.
Zeng F. Y. i Wess J. (1999) Identification and Molecular Characteritzation of m3 Muscarinic receptor Dimers. J. Biol. Chem. 274; 19487-19497.
IV.1.-
Efecte de la temperatura, el zenc i l’entorn lipídic en
la conformació de la rodopsina i el seu cromòfor.
Per a la caracterització i estudi de la rodopsina nativa és necessària primer l’obtenció de la matèria primera, rodopsina a partir de retines bovines utilitzant tècniques d’extracció i d’immunopurificació.
IV.1.1.-Obtenció de la rodopsina a partir de retines bovines
Per obtenir la rodopsina a partir de retines bovines primer cal fer una extracció de ROS.
IV.1.1.1.-Extracció de ROS a partir de retines bovines
Seguint el procediment descrit en III.2.1 s’obté l’espectre de la figura 4.1.
Fig. 4.1: Espectre de ROS en tampó de fosfat potassi 70 mM pH 6.9, EDTA 0.1mM i 1% DM a 20ºC.
A partir de l’absorbància a 500nm, sabent que ε500nm de rodopsina de ROS és
40600 cm-1M-1 (Wald and Brown, 1953) i que la massa molecular és de 40000
[image:32.595.131.447.399.633.2]obtinguda de les 100 retines utilitzades és de 32 mg de proteïna. Prèviament s’ha descrit que per aquest número de retines s’haurien d’obtenir al voltant de 35 mg (Papermaster i Dreyer, 1974). Aquest resultat demostraria que no hi ha hagut pèrdues de proteïna durant el llarg procés d’extracció de ROS.
La relació (o ratio) A280nm/A500nm, que és un dels criteris que ens indica
quina és la puresa de rodopsina en la mostra de ROS (Salesse et al., 1984), és de
3.6. Per intentar disminuir la relació a 2.4-2.6 com s’ha descrit anteriorment (Papermaster i Dreyer, 1974) i eliminar part de proteïna que encara es troba en la mostra i que no ens interessa ja que no és rodopsina, es va realitzar un rentat de la mostra amb urea.
Un cop realitzat el rentat la proteïna presenta una lleugera disminució de la relació A280nm/A500nm obtenint-la de 3.4 (espectre no mostrat).
Tot i que aquesta mostra de ROS ja es pot utilitzar, es pot augmentar la puresa de rodopsina utilitzant cromatografia d’immunoafinitat. Prèviament però s’ha d’acoblar una matriu de sefarosa amb l’anticòs Rho-1D4.
IV.1.1.2.-Acoblament de l’anticòs Rho-1D4 a CNBr-sefarosa i determinació de la seva capacitat d’unió
Abans de l’acoblament és necessari realitzar un espectre de l’anticòs Rho-1D4 per calcular la quantitat de sefarosa necessària, descrit en IV.2.1.4. A partir de l’absorbància a 280 nm (Fig. 4.2), amb un valor de ε1%=1.383 mg/ml-1·cm-1 es
calcula que la quantitat d’anticòs inicial és de 126 mg.
Es realitza l’acoblament de l’anticòs a sefarosa i posteriorment es calcula la capacitat de la reïna per unir rodopsina. Per això, s’incuba ROS solubilitzada amb la reïna i s’elueix la proteïna amb dos tampons. Les 6 primeres elucions s’han obtingut amb tampó de baixa força iònica (Na2HPO4 2mM pH=6) mentre
que en l’última mostra, el rentat E, s’ha eluït la proteïna amb tampó d’alta força iònica (Na2HPO4 2mM pH=6 i 150 mM NaCl) a on es desprèn tot el que encara
Fig. 4.2: Espectre de l’anticòs Rho-1D4 en NaHCO3 0.25M i NaCl 0.9M (pH=9).
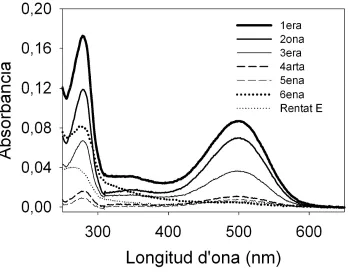
Fig. 4.3: Espectres de la rodopsina de ROS purificada en Na2HPO4 2mM pH=6 i 0.05% DM o Na2HPO4 2mM pH=6, 150 mM NaCl i 0.05% DM (Rentat E) a partir de la reïna acoblada amb l’anticòs Rho-1D4.
Longitud d'ona (nm)
260
280
300
320
340
Absorbància
[image:34.595.122.468.87.359.2] [image:34.595.120.469.409.683.2]A partir de l’absorbància a 500 nm dels espectres (Fig. 4.3) es calcula la quantitat de proteïna obtenint el resultats de la taula 4.1.
Taula 4.1: Quantitat de rodopsina purificada amb la reïna acoblada a l’anticòs Rho-1D4.
De la proteïna utilitzada inicialment, s’ha recuperat un 60% després de la purificació i majoritàriament entre la 1era i la 2ona elució (78% de la proteïna total purificada).
Calculant la diferència de la proteïna inicial, la total eluïda i sabent la quantitat de reïna que s’ha utilitzat en la purificació s’obté que la capacitat d’unió de la reïna és d’1.26 µg de proteïna/µl de reïna, similar a valors obtinguts anteriorment (Cha et al., 2000 i Hwa et al., 1997).
Aquesta reïna es podrà utilitzar per a la purificació de rodopsina a partir de retines bovines o bé rodopsina recombinant expressada en cèl·lules COS-1.
Elució Volum
(ml) Absorbància Quantitat de Proteïna (µg)
1era 2ona 3era 4era 5ena 6ena Rentat tampó E
1.0 0.5 0.5 0.5 0.5 0.5 0.5 0,087 0,070 0.036 0.011 0.008 0.005 0.005 86 34.6 18 5.3 3.7 2.7 2.6
Proteïna total obtinguda 152.9
IV.1.1.3.-Purificació de la rodopsina a partir de ROS
Un cop obtingut ROS a partir de les retines, es purifica rodopsina mitjançant el procediment explicat en III.2.1.5, es realitza un espectre de cadascuna de les elucions i el resultat d’aquests espectres és a la figura 4.4.
Fig. 4.4: Espectres d’absorció de les elucions de la rodopsina purificada en Na2HPO4 2mM pH=6 i DM 0.05% a 20ºC.
[image:36.595.112.469.212.475.2]Elucions A280nm/ A500nm
1 1.6 2 1.8 3 2.5 4 2.4 5 3.1
Taula 4.2: Relació A280nm/A500nm obtinguda en les diferents elucions de la rodopsina purificada.
L’avantatge d’utilitzar rodopsina i ROS de retina és treballar amb la proteïna nativa pròpiament dita, a més que se n’obté amb més quantitat. Això facilita el seu estudi si es compara amb les proteïnes recombinants, que requereixen més manipulació, més temps i on la quantitat de proteïna obtinguda és més petita que a partir de retines.
Saber quins factors poden influir en l’estabilitat de la rodopsina és important per tenir un coneixement més profund de la proteïna en les condicions de treball i aplicar-ho en l’estudi de proteïnes mutants associades a malalties de la retina.
En aquest treball es mostrarà quina influència té la temperatura en la isomerització de l’11-cis-retinal i quin és l’efecte del zenc i del detergent DM,
utilitzat en la purificació de la proteïna, en l’estabilitat de rodopsina.
IV.1.2.- Efecte del detergent DM en l’estabilitat de la rodopsina
En aquest apartat s’estudia com la concentració de DM afecta l’estabilitat de la rodopsina, concretament la dels fotointermediaris MetaII i MetaIII seguint el protocol descrit en III.2.5.3.3.
IV.1.2.1.-Estabilitat de la MetaII a diferents concentracions de DM
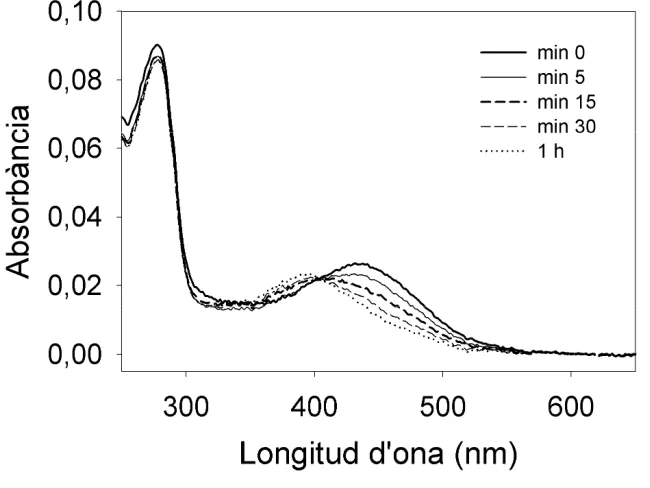
L’estabilitat de la MetaII es mesura a partir del valor d’absorbància a 440nm d’una mostra de rodopsina acidificada després de ser il·luminada. Entre la il·luminació i l’acidificació es deixen passar diferents temps, des de 0 minuts fins a 1 o 2 h. A mesura que passa el temps, s’observa que la mostra de rodopsina acidificada presenta un màxim desplaçat a longituds d’ona més baixes de 440 nm (Fig. 4.5), degut a que la MetaII es va separant en opsina i tot-trans-retinal.
Fig. 4.5: Espectres obtinguts després d’acidificar a diferents temps rodopsina il·luminada en tampó fosfat potassi 70 mM pH 6.9, EDTA 0.1mM i 0.05% DM a 20ºC.
[image:38.595.128.453.363.607.2]Fig. 4.6: Decay de la MetaII en tampó fosfat potassi 70 mM pH 6.9 i EDTA 0.1mM a les
diferents concentracions de DM, determinat a partir del mètode d’acidificació (Sakamoto i Khorana, 1995).
En aquest gràfic s’observa com baixes concentracions de DM promouen la separació de l’opsina del tot-trans-retinal mentre que l’estabilitat de MetaII és
similar en 0.05% i 0.025% de DM, que es confirma amb el càlcul del temps de vida mitjana (t½) (taula 4.3).
% DM t½ MetaII (min)
0.012 9.1 0.025 18.2 0.05 18.1
Taula 4.3. Temps de vida mitja de la Meta II a diferents concentracions de DM.
[image:39.595.96.456.86.352.2]IV.1.2.2.-Formació de la MetaIII en presència de diferents concentracions de DM
Pel seguiment de la formació de MetaIII es segueix el valor d’absorbància a 465 nm a les diferents concentracions de detergent (Fig. 4.7) i es calcula el % de formació del fotointermediari respecte el valor d’absorbància a 465 nm de la mostra inicial en foscor. El percentatge de formació de la MetaIII en les diferents concentracions de detergent es mostra a la taula 4.4.
%DM % Formació MetaIII
0.012 36 0.0175 30
0.025 24 0.05 15
Taula 4.4. Percentatge de formació de MetaIII a diferents concentracions de DM.
Fig. 4.7: Espectres de la formació de la MetaIII en tampó fosfat potassi 70 mM (pH 6.9) i EDTA 0.1mM a les diferents concentracions de DM. A) Formació de MetaIII en 0.05%DM 1) foscor, 2) espectre il·luminat minut 0, 3) minut 3, 4) minut 6 i 5) 36 minuts. B) Formació de MetaIII en 0.012%DM 1) en foscor, 2) il·luminat minut 0, 3) minut 6, 4) minut 12 i 5) minut 24.
[image:41.595.138.452.92.595.2]IV.1.2.3.-Com afecta la concentració de detergent a l’estabilitat i l’activitat de la rodopsina?
A mesura que disminuïm la concentració de detergent, observem una disminució de l’estabilitat de la MetaII, mentre que la formació de MetaIII es veu disminuïda per l’increment de concentració del DM.
La concentració de detergent condiciona l’equilibri entre MetaI i MetaII i s’ha observat que el DM desplaça l’equilibri cap a la formació de MetaII (Vogel et al.
2004), tal i com mostren els nostres resultats. En canvi en un altre detergent, la digitonina, s’ha vist que dóna rigidesa a les micel·les de rodopsina-detergent-membranes, i que fa desplaçar l’equilibri cap a la formació de MetaI (Vogel et al.
2004). En els nostres resultats, aquesta rigidesa dels complexos es podria comparar amb la de la membrana de ROS sense solubilitzar o amb poca presència de detergent, provocant que es formés MetaIII, via MetaI (Vogel et al,
2004).
Els resultats obtinguts es poden complementar amb dues característiques més com són la funcionalitat i l’estabilitat de rodopsina mesurada per calorimetria diferencial de rastreig (DSC, difference scanning calorimetry) (Marrón, 1996).
Els resultats referents a la capacitat de rodopsina per activar la proteïna G si es realitza a diferents concentracions de DM, des de 0% fins a 0.05% de DM, mostren que augmenta l’activitat del receptor des de 0% DM fins a 0.01% on s’observa una màxima activació i a partir d’aquí, al augmentar la concentració l’activitat disminueix.
Quan es comparen aquests resultats amb els de l’estabilitat de MetaII i la formació de MetaIII ens trobem que un paper important el realitzaria la presència de transducina a l’entorn, que actua desplaçant l’equilibri de MetaIII a MetaII provocant que aquesta rodopsina inactiva però unida al tot-trans
-retinal, passi a la forma activa (Zimmermann et al. 2004). Així doncs, mentre
des del punt de vista espectroscòpic, sigui veritablement MetaII la que activi a la transducina en els experiments de funcionalitat.
El perquè a altes concentracions de detergent la Meta II és més estable però activa menys a la transducina, s’explicaria per què les micel·les formades pel DM poden alterar l’equilibri de proteïna-membrana-detergent de cada micel·la donant un entorn a la rodopsina no favorable per a la unió i/o activació de la proteïna G (Ramon et al.,2003 i Mitchell et al, 2001; Bubis, 1998), i també per
que aquestes condicions poden afectar l’estabilitat de la transducina.
En resum, altes concentracions de detergent afavoreixen una millor estabilitat de la MetaII però no afavoreixen les condicions per a que aquesta MetaII formada pugui activar a la proteïna G. És per això que els experiments d’activació es realitzen en les concentracions més baixes de DM, però per sobre de la concentració micel·lar crítica (0.01%DM).
[image:43.595.112.449.481.745.2]Tots aquests factors aporten més evidències de que l’entorn lipídic condiciona l’estabilitat dels fotointermediaris i la funcionalitat de la rodopsina.
IV.1.3.- Isomerització específica de l’11-cis-retinal unit a la rodopsina
sota condicions de desnaturalització
La majoria de treballs publicats de rodopsina es centren en la part proteica, és a dir, en realitzar mutacions en la proteïna per estudiar l’efecte en l’estructura i/o funció de l’opsina, però cal considerar la importància de l’altra part que constitueix la rodopsina, l’11-cis-retinal. En aquest apartat s’estudia que li
succeeix a l’11-cis-retinal un cop és tractat a 55ºC, en solució i quan està unit a la rodopsina, per això, s’han realitzat els experiments descrits en III.2.5.3.2 i s’obtenen els resultats de les figures 4.9 i 4.10.
Fig. 4.9: Espectres de diferència entre abans i després d’il·luminar de a) 11-cis-retinal en etanol b) tot-trans-retinal en etanol c) rodopsina en Na2HPO4 2mM pH=6.0 tractada a 55ºC durant 30 min.
S’observa que el retinal extret de rodopsina tractada a 55ºC (C) presenta un comportament molt similar al tot-trans-retinal (B). Per estudiar si aquesta
[image:44.595.108.465.355.620.2]-retinal, s’ha repetit l’experiment utilitzant 11-cis-retinal tractat a 55ºC i els
resultats es mostren en la figura 4.10.
Fig. 4.10: Espectres de diferència d’abans i després d’il·luminar de a) 11-cis-retinal en etanol b)
tot-trans-retinal en les mateixes condicions que l’11-cis-retinal i c) 11-cis-retinal en etanol i tractat a 55ºC durant 30 min.
L’espectre de diferència de l’11-cis-retinal tractat a 55ºC (C) és similar al del cromòfor sense tractar (A).
Aquests resultats demostren que l’11-cis-retinal i el tot-trans-retinal presenten un diferent patró d’espectre de diferència i d’aquesta manera els podem distingir en aquests experiments. En les figures 4.9 i 4.10 observem que l’espectre de diferència del retinal extret de la rodopsina tractada a 55ºC no presenta un mateix patró que el de l’11-cis-retinal en solució sinó similar al del
tot-trans-retinal. Mentre que quan es mostra l’espectre de diferència de l’11-cis
-retinal en solució tractat a 55ºC, aquest és molt similar al obtingut en el cas d’aquest mateix pigment sense tractar.
[image:45.595.97.471.120.407.2]Fig. 4.11: Cromatograma d’HPLC d’11-cis-retinal lliure i de rodopsina tractat a 55ºC. En les abscisses es mostren el temps de retenció dels diferents retinals en 72% acetonitril aquós (volum/volum) i acetat amoni 0.015 M pH=5.5 (A: mostres d’11-cis-retinal en foscor (a), il·luminat (b) i tot-trans-retinal en foscor; B: barreja d’11-cis-retinal i tot-trans-retinal en foscor
(a), 11-cis-retinal tractat 1h a 55ºC (b) i tot-trans-retinal tractat 1h a 55ºC; C: barreja d’11-cis -retinal i tot-trans-retinal en foscor (a), retinal extret de rodopsina sense tractar (b) i retinal
extret de rodopsina tractada 30 min a 55ºC (c)).
Els resultats obtinguts a partir de l’HPLC confirmen el que ja s’havia observat en l’anàlisi espectroscòpica. En la figura 4.11A es mostra una barreja d’11-cis
-retinal en foscor (a) i il·luminat (b) i tot-trans-retinal en foscor (c). En les mostres en foscor s’observen 2 temps de retenció diferents; l’11-cis-retinal a 14.5
min i 15.8 min en el cas del tot-trans-retinal. La mostra il·luminada ens
[image:46.595.228.379.73.474.2]11-cis-retinal il·luminat, és a dir tot-trans-retinal, a 15.8 min, causat per que la mostra
d’11-cis-retinal es troba parcialment il·luminada.
La figura 4.11B mostra una barreja d’11-cis-retinal i tot-trans-retinal sense
tractar (a), 11-cis-retinal tractada a 55ºC (b) i tot-trans-retinal tractat a 55ºC (c).
En el cromatograma, observem que les mostres sense tractar i les mostres tractades presenten el mateix comportament que els respectius sense tractar. Per tant, l’11-cis-retinal i el tot-trans-retinal mantenen la seva configuració a
55ºC.
La figura 4.11C mostra el cromatograma d’una barreja d’11-cis-retinal i
tot-trans-retinal sense tractar (a), una mostra del retinal extret a partir de
rodopsina sense tractar (descrit en l’apartat III.2.8.) (b) i mostra del retinal extret que prové de rodopsina tractada i que el seu espectre de diferència correspon al C de la figura 4.9. S’observa que el temps de retenció del retinal extret de rodopsina no tractada correspon al temps de l’11-cis-retinal, 14.5 min,
mentre que el retinal extret de la mostra de rodopsina tractada a 55ºC correspon al temps de retenció del tot-trans-retinal, 15.8 min.
Els cromatogrames confirmen el que s’havia observat en les figures 4.9 i 4.10, és a dir, que l’11-cis-retinal tractat durant 1 h a 55ºC en solució és més estable
que quan es troba unit a la opsina, on s’isomeritza a tot-trans-retinal.
Aquests resultats s’expliquen per que l’energia llumínica necessària pel canvi d’isomerització entre l’11-cis i el tot-trans és molt similar a la energia tèrmica
que s’ha proporcionar al sistema per a permetre el mateix efecte (Ala-Laurita et
al. 2004) i sembla ser que l’energia tèrmica equivalent a 55ºC permet la
isomerització.
Segons aquests resultats, quan la rodopsina és tractada a 55ºC i a mesura que es va desplegant, va provocant el canvi d’11-cis-retinal a tot-trans-retinal. Això
IV.1.4.- Efecte del zenc en l’estabilitat de la rodopsina
En aquest apartat s’estudia l’efecte del zenc, un ió present en elevades concentracions a la retina, en l’estabilitat de la conformació inactiva i activa de rodopsina purificada i de ROS, la capacitat de la rodopsina il·luminada per tornar a unir-se a retinal i observar si provoca un canvi intern en la proteïna mitjançant el càlcul de la quarta derivada de l’espectre d’absorció que ens determinarà si el zenc produeix un canvi en l’entorn dels aminoàcids aromàtics (vegeu l’apartat III.2.5.3.1).
IV.1.4.1.-Estabilitat tèrmica de la forma inactiva de la rodopsina en presència de zenc i d’altres cations.
A partir de l’absorbància a 500 nm (que disminueix al llarg del temps, vegeu figura 4.7) es representen les corbes de desnaturalització en absència o presència de 15 i 50 µM ZnCl2 (Fig. 4.12).
[image:48.595.128.473.417.699.2]A mesura que s’augmenta la concentració de zenc es produeix un efecte desestabilitzador en la conformació inactiva de rodopsina. Aquest resultat també s’ha observat en una publicació recent en que es treballa amb rodopsines mutants d’RP i on el resultat de la rodopsina wild type (WT) a 10µM i 50µM
confirma l’augment de la velocitat de decay en experiments realitzats en
presència de zenc (Stojanovic et al. 2004).
Per tal de comprovar si aquest efecte s’observa només en presència de zenc, es realitza el mateix assaig en presència d’altres cations que formen part del grup dels elements de transició dins de la taula periòdica (tals com el cadmi, el cobalt i el coure), el calci i la colina. Aquest darrer és per descartar l’efecte de la càrrega. Aquests cations ja s’havien utilitzat anteriorment en rodopsina per estudiar la seva competència respecte el zenc (Schuster et al., 1992). Els
resultats obtinguts es representen en la figura 4.13.
Fig. 4.13: Corba de desnaturalització tèrmica a 55ºC de rodopsina purificada en Hepes 20 mM (pH 7.4), NaCl 145 M, MgCl2 2 mM i 0.02%DM en presència dels diferents cations a 50 µM.
[image:49.595.98.457.363.663.2]observat que competeix amb el zenc per la unió de rodopsina (Schuster et al.,
1992). El calci, cadmi o cobalt provoquen un efecte similar al de la proteïna en absència de cations i en el cas de la colina, sembla que hi ha un augment de l’estabilitat al llarg del temps, a causa de la seva condició de component de les membranes que estabilitzarien a la rodopsina dins les micel·les.
Paral·lelament es comprova si aquest resultat obtingut en rodopsina purificada, on les condicions no són naturals, s’observa amb rodopsina en les membranes de ROS (Fig. 4.14).
Fig. 4.14: Estabilitat tèrmica de rodopsina de membranes de ROS en Hepes 20 mM (pH 7.4), NaCl 145 M, MgCl2 2 mM en absència i presència de zenc a 55ºC.
Aquests resultats mostren que una mostra de ROS en membranes presenta el mateix comportament que una de rodopsina purificada i en detergent ja que el zenc augmenta la velocitat de desnaturalització tèrmica de la rodopsina.
IV.1.4.2.-Determinació dels paràmetres termodinàmics de la rodopsina en presència de zenc
[image:50.595.110.461.242.521.2]µM de zenc mitjançant el gràfic d’Arrhenius per estudiar quins paràmetres energètics canvien i relacionar-los amb canvis en la conformació de la rodopsina entre ambdues condicions (Fig. 4.15).
Fig. 4.15: Gràfic d’Arrhenius de la desnaturalització tèrmica de rodopsina purificada en Hepes
20 mM (pH 7.4), NaCl 145 M, MgCl2 2 mM i 0.02%DM en presència de 50 µM (●) i absència de zenc (0).
Considerant que els valors de constant de velocitat (k) alts corresponen a menys estabilitat per part de la proteïna, aquests resultats confirmen que la proteïna control és més estable que en presència de zenc a 50 µM. A partir del valor de k i seguint les fórmules definides en la taula 3.3 es calculen els paràmetres termodinàmics d’Ea, ∆H, ∆G i ∆S, (taula 4.5).
Paràmetre Absència de Zenc Presència de Zenc (50µM)
d’Ea 99.09 ± 0.06 156.18 ± 0.20
∆H 98.44 ± 0.05 155.53 ± 0.19
∆G 5.74 ± 0.03 4.77 ± 0.04
∆S(10-3) 282.62 ± 0.06 460.00 ± 0.09
[image:51.595.115.414.148.374.2]