TESIS
ANÁLISIS FUNCIONAL DEL SECUESTRO DE CARBONO EN EL GRADIENTE SUCESIONAL DE UN BOSQUE SECO TROPICAL DEL VALLE DEL RÍO MAGDALENA
MAESTRIA EN MANEJO, USO Y CONSERVACIÓN DEL BOSQUE UNIVERSIDAD DISTRITAL FRANCISCO JOSÉ DE CALDAS
AUTOR
WILBER ALFONSO TRIANA DIOSA INGENIERO FORESTAL
DIRECTOR EXTERNO JHON JAIRO SANCHEZ
MAGISTER EN MANEJO Y CONSERVACIÓN DE BOSQUES TROPICALES Y BIODIVERSIDAD
TESIS SOMETIDA A CONSIDERACIÓN DEL CONSEJO CURRICULAR DE LA MAESTRIA EN MANEJO, USO Y CONSERVACIÓN DEL BOSQUE COMO REQUISITO PARA OPTAR POR EL GRADO DE
MAGISTER EN MANEJO, USO Y CONSERVACIÓN DEL BOSQUE
2017
Esta tesis ha sido aprobada por el consejo curricular de la MAESTRIA EN MANEJO USO Y CONSERVACION DEL BOSQUE como requisito para optar por el grado de MAGISTER EN MANEJO, USO Y CONSERVACIÓN DEL BOSQUE
FIRMANTES:
__________________________________ Edgar Andrés Avella
Jurado
____________________________________ Germán Andrés González
Jurado
__________________________________ JHON JAIRO SANCHEZ CORREA
Director de Tesis
___________________________________ WILBER ALFONSO TRIANA DIOSA
TABLA DE CONTENIDO
ANÁLISIS FUNCIONAL DEL SECUESTRO DE CARBONO EN EL GRADIENTE SUCESIONAL DE UN
BOSQUE SECO TROPICAL DEL VALLE DEL RÍO MAGDALENA ... 6
RESUMEN ... 6
INTRODUCCIÓN ... 7
MATERIALES Y MÉTODOS ... 10
Área de estudio ... 10
Determinación de Estados Sucesionales ... 11
Selección de Especies ... 12
Selección de Individuos ... 12
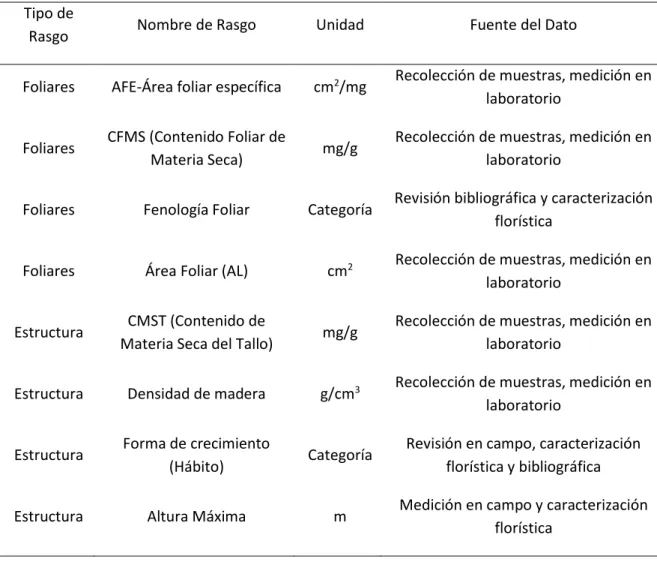
Determinación de Rasgos Funcionales ... 12
Manejo de Datos ... 18
RESULTADOS ... 19
Especies seleccionadas para cada estado sucesional ... 19
Valores de Rasgos Funcionales... 20
Determinación de Tipos Funcionales de Plantas (TFPs) ... 20
Composición de TFPs en el gradiente sucesional ... 25
Índices de Diversidad Funcional ... 29
DISCUSIÓN ... 31
Tipos Funcionales de Plantas asociadas a secuestro de carbono en Bs-T ... 31
Variación de TFPs en el gradiente sucesional ... 32
Variación de los rasgos de madera en el gradiente sucesional ... 34
Variación de índices multirasgo en el gradiente sucesional ... 34
Diversidad Funcional como indicadora de secuestro biológico de carbono ... 36
CONCLUSIONES ... 37
RECOMENDACIONES ... 38
REFERENCIAS BIBLIOGRÁFICAS ... 39
LISTA DE TABLAS
Tabla 1. Rasgos funcionales relacionados con el secuestro biológico de carbono ... 13 Tabla 2. Especies seleccionadas para el estudio con base en su dominancia en cada estado sucesional. AB%: Área Basal Relativa. IVI%: Índice de Valor de Importancia Relativo. ... 19 Tabla 3. MANOVA para comparación de TFPs a partir de Rasgos Funcionales. Medias con una letra común no son significativamente diferentes (P< 0.05). AF: Área Foliar. AFE: Área Foliar Específica. CFMS: Contenido de Foliar de Materia Seca. CMST: Contenido de Materia Seca en el Tallo. DM: Densidad de la Madera. Hmax: Altura Máxima. ... 22 Tabla 4. Análisis de tablas de contingencia a partir de Chi Cuadrado entre TFPs, fenología foliar y forma de crecimiento. Asociaciones significativas, p<0.05(*) ... 24 Tabla 5. Rasgos dominantes y nombre de los TFPs. AF: Área Foliar. AFE: Área Foliar Específica. CFMS: Contenido de Foliar de Materia Seca. CMST: Contenido de Materia Seca en el Tallo. DM: Densidad de la madera. Hmax: Altura Máxima. ... 25
LISTA DE ANEXOS
LISTA DE FIGURAS
ANÁLISIS FUNCIONAL DEL SECUESTRO DE CARBONO EN EL GRADIENTE SUCESIONAL DE UN BOSQUE SECO TROPICAL DEL VALLE DEL RÍO MAGDALENA
RESUMEN
INTRODUCCIÓN
La distribución del bosque seco tropical en Colombia se ha visto fuertemente reducida por procesos de deforestación y colonización desde hace más de 500 años (Etter et al. 2008). En la actualidad se ha visto impactado por nuevos tensionantes como el turismo, el desarrollo urbano y la minería, ocasionando que se encuentre totalmente fragmentado y rodeado de matrices de transformación (Portillo y Sánchez, 2010). Estas afectaciones lo han llevado a ser considerado uno de los ecosistemas más amenazados del neotrópico (Miles et al.2006, Pennington, 2012). Es así, que la restauración ecológica del bosque seco tropical se convierte en una necesidad imperante, búsqueda que se debe fundamentar en la recuperación de los atributos esenciales que definen el ecosistema y le dan su identidad (composición, estructura y función), los cuales influyen de manera determinante en los servicios ecosistémicos que este provee (Murcia y Guariguata, 2014).
Pese a tener tal nivel de importancia, tanto la planificación como el monitoreo de los procesos de restauración ecológica han estado enfocados en evaluar la composición y estructura de los ecosistemas restaurados. En Colombia, según Murcia y Guariguata (2014), dentro de 81 proyectos que establecían una línea base para la planificación de acciones de restauración, sólo tres incluyeron estudios ecosistémicos relacionados con características funcionales. De igual forma, el éxito de los mismos se ha medido a partir del área sembrada o plantada, la tasa de supervivencia del material plantado, el cambio de una cobertura por otra o la densidad y porcentaje de cobertura de las plantas en un determinado período de tiempo; lo cual, según Aguilar y Ramírez (2015) subestima el proceso de restauración, al no medir su impacto en otros compartimientos del ecosistema, en los procesos ecosistémicos y en los servicios que de estos se derivan.
Pennington (2012) y Portillo y Sánchez (2010) argumentan que se presentan profundas falencias en cuanto al conocimiento de los procesos ecológicos y los servicios ecosistémicos que genera el bosque seco tropical, por lo que a fin de lograr procesos de restauración ecológica efectivos, en los que se recuperen dichos procesos y servicios, es importante conocer su dinámica en procesos de sucesión posteriores al abandono y las consecuencias que esto tiene para su conservación (Torres et al. 2012). La mayor parte de los estudios que se han desarrollado en este aspecto, se han enfocado en bosques húmedos tropicales, los cuales presentan diferencias importantes en la forma en la que se desarrolla la sucesión, sus valores de rasgos y sus relaciones entre estos rasgos (Wright y Westoby, 2002; Poorter y Kitajima, 2007), particularmente se ha observado que al presentar estaciones secas, el Bs-T presenta periodos de tiempo en los que la irradiancia sobre el sotobosque aumenta de manera importante, sugiriendo que otros factores diferentes a la disponibilidad de luz podrían describir mejor las variaciones de rasgos en este ecosistema que en su homólogo húmedo (Lebrija-Trejos, 2009). Considerando lo anterior, de determinar patrones en el comportamiento de la sucesión pueden partir acciones de manejo adecuadas y la medición de su impacto una vez se busca inducir estos cambios. Profundizar en estas áreas del conocimiento, particularmente en los cambios funcionales que se presentan durante la sucesión, permitirá ofrecer lineamientos para la restauración de estos ecosistemas no solo desde la composición sino desde la función (Portillo y Sánchez, 2010).
recuperación, conocer la manera en la que varía su capacidad para prestar este servicio durante esta sucesión es especialmente importante, ya que como plantean Aguilar y Ramírez (2015), es necesario utilizar indicadores de captura de CO2 que determinen el impacto de las acciones de restauración a nivel de comunidades y ecosistemas.
Los rasgos funcionales son el medio por el cual se determina la diversidad funcional de un ecosistema. Por rasgo se entiende cualquier característica morfológica, fisiológica o fenológica medible a nivel individual para todo organismo, independiente del ambiente o cualquier otro nivel de organización (Violle et al., 2007), estos se han propuesto como el vínculo más directo entre la biodiversidad y los procesos de los ecosistemas (Díaz et al. 2006).
En el caso específico del secuestro carbono, Cornelissen et al. (2003) y Casanoves et al. (2011) plantean que los rasgos de plantas más relacionados son el área foliar (AF), área foliar específica (AFE), contenido foliar de materia seca (CFMS), densidad de la madera (Dm), contenido de materia seca del tallo (CMST), altura máxima (Hmax), forma de crecimiento y fenología foliar, relaciones que podrían ser entendidas de la siguiente manera:
En primera instancia, se entiende que los valores de área foliar configuran de manera importante la capacidad de las plantas para interceptar la radiación fotosintéticamente activa (RFA), siendo esta la fuente primaria de energía utilizada para la elaboración de compuestos y tejidos (Warnock et al., 2006), elementos en los cuales se ve almacenado el carbono finalmente (Taiz y Zeiger, 2006). Sin embargo, no toda la radiación fotosintéticamente activa es utilizada para la incorporación en los tejidos vegetales y por tanto el crecimiento, también en esta ecuación influye de manera determinante la tasa de asimilación neta (NAR) y en función de esta, la tasa de crecimiento relativo (RGR), la cual se encuentra estrechamente relacionada de manera positiva con el área foliar específica (Villar, 2004) y de manera negativa con el contenido foliar de materia seca (De La Riva et al.,2014).
La forma de crecimiento (hábito) y la altura máxima identifican el estrato que ocupa una especie en un gradiente lumínico, debido a que determinan el acceso a la luz (Poorter, Bongers y Bongers, 2006) y su tasa potencial de crecimiento en función de este, factores que influyen en su capacidad de captura de CO2 (Pérez-Harguindeguy et al. 2013). De igual forma, se ha conocido que las especies caducifolias suelen presentar un rápido crecimiento (Cornelissen, 2003) y tasas fotosintéticas altas (Easmus, 1999), al ser comparadas con las especies siempreverdes o perenes, por lo que las primeras presentan tasas de fotosíntesis más elevadas en periodos de tiempo reducidos, lo que conlleva a una mayor fijación en términos netos. Por otra parte, la densidad de la madera suele ser utilizada para estimar el contenido de diversos elementos biogeoquímicos en la madera (Návar, 2009) entre ellos, el contenido de carbono (Chavé, 2002), así como la estabilidad y la tasa de crecimiento volumétrica (Chave, 2009).
Pese a lo anterior, el entendimiento de las relaciones que se presentan entre los rasgos funcionales y la captura o almacenamiento no es suficiente al abarcarse de manera singular, considerando que las especies vegetales no presentan uno sólo de estos rasgos, sino una combinación de los mismos. Dichas combinaciones, conocidas como síndromes funcionales (Martín et al., 2007), buscan ser simplificadas para un mejor entendimiento en dos estrategias de inversión de recursos: especies adquisitivas, que presentan una captura, pero también una liberación de recursos prematura, y conservativas, que presentan una captura de recursos más lenta, pero de igual forma su liberación requiere de mayor tiempo (Diaz et al., 2004, Wright et al., 2004).
En Colombia una gran cantidad de estudios se han enfocado en estimar el almacenamiento de carbono en la biomasa aérea (Duque et al., 2011a, Duque et al., 2011b, Duque et al., 2012, Vásquez, 2009), la caída de hojarasca (Mosquera et al. 2007), madera muerta (Duque et al., 2011c) y biomasa subterránea en bosques sucesionales y maduros (Sierra et al., 2001). De la misma manera, se han realizado estudios a fin de determinar los flujos de carbono en la Amazonía colombiana (Rangel, 2009) y buscando validar modelos de acumulación de biomasa en diversos ecosistemas (Duque et al., 2010). Por otro lado, se han realizado algunos estudios buscando relacionar valores de algunos rasgos relacionados con la captura de CO2 a partir de la madera (Rodríguez, 2012), y la madera y hojas (Turriago, 2008). Estudios más recientes (López, 2015) ha relacionado la diversidad funcional con la captura de CO2. Por otro lado, se han presentado aproximaciones importantes que detallan a nivel regional y local diversos servicios ecosistémicos que ofrecen los Bs-T (Maass et al., 2004), así como las relaciones de costo-beneficio que representan los procesos de restauración ecológica que se adelantan en estos ecosistemas (Birch et al., 2010). Inclusive, se han abordado servicios de aprovisionamiento en bosques secos tropicales secundarios en diferentes estados sucesionales en el caribe colombiano (Castellanos, 2013). Sin embargo, no se conoce si los cambios presentes en la diversidad funcional a medida que transcurre la sucesión pudiesen estar relacionados con el secuestro biológico de carbono y si de presentarse, estas variaciones podrían servir para inferir variaciones en el mismo.
El ensamblaje comunitario presente en los procesos sucesionales que acontecen después del abandono en ecosistemas en recuperación es producto de la respuesta de las especies a diferentes filtros ambientales (McGill, 2016), que bien pueden ser bióticos (competencia, herbivoría, enfermedades, etc.) o abióticos (disponibilidad de nutrientes, agua, luz, temperatura, etc.) (Cornwell y Askerly, 2009). Al presentarse dichos cambios ambientales, se espera que paralelamente a los cambios en composición y estructura (Mayfield et al., 2010; Putz et al., 2012) se evidencien valores de rasgos que respondan a las nuevas condiciones (Díaz et al., 1997; Petchey et al., 2007), y que de igual forma influyan de manera diferencial sobre los procesos ecosistémicos subyacentes (Grime et al., 1997).
En algunos estudios se ha encontrado que la diversidad funcional, expresada en rasgos como la fenología foliar, la densidad de la madera y las ramas, el área foliar específica, el contenido foliar de materia seca y el contenido de nutrientes varía en diferentes estados sucesionales (Carreño et al., 2012; Castellanos, 2013; López, 2015, Sanaphre-Villanueva et al., 2017), algunos inclusive han hallado diferencias importantes entre gradientes sucesionales de ecosistemas tropicales húmedos y secos (Lohbeck et al., 2013). Pese a lo anterior, son muy pocos los estudios en bosques secos interandinos (Pennington, 2012), y más aún, que evalúen la influencia que pudiesen tener estos cambios en la generación de servicios ecosistémicos como el de secuestro de carbono.
MATERIALES Y MÉTODOS
Área de estudio
El estudio se desarrolló en un bosque seco tropical (Bs-T) de la cuenca alta del río Magdalena, en el área de compensación del Proyecto Hidroeléctrico el Quimbo, entre las coordenadas Oeste 75°41’44.84’’-75°41’26.23’’ y Norte 2°18’38.50’’-2°18’.19.21’’. En esta zona se viene adelantando el proyecto piloto de restauración ecológica del Bosque Seco Tropical del Quimbo (Figura 1).
Figura 1. Ubicación del área de estudio. Coordenadas Oeste 75°41’44.84’’-75°41’26.23’’ y Norte 2°18’38.50’’-2°18’.19.21’’
El área de estudio se encuentra entre 775 a 978 msnm, con una temperatura promedio de 24.3°C, presenta una precipitación media anual de 1063 mm, distribuyéndose en un régimen monomodal con 180 días de lluvia al año que comprende un período húmedo de octubre a marzo y uno de sequía marcada de abril a septiembre (IDEAM, 2010).
Determinación de Estados Sucesionales
Los estados sucesionales se determinaron a partir del estudio de la caracterización florística y estructural de la zona 2 del proyecto hidroeléctrico El Quimbo, realizada por Fundación Natura (ver García et al., 2015) y revisión en campo, considerando los valores de área basal, estratificación vertical, alturas máximas y diversidad de plantas, para tener mayor claridad de la composición y estructura general de cada estado sucesional evaluado, en los Anexo 1, Anexo 2, Anexo 3 se muestra el índice de valor de importancia (IVI) de las especies presentes en cada uno.
1. Pastos arbolados con dominio de Guazuma ulmifolia
En los levantamientos realizados en este bosque se registraron 160 individuos distribuidos en 22 especies, 18 géneros y 13 familias. Las familias que caracterizan florísticamente la zona son Malvaceae con Guazuma ulmifolia, Salicaceae con Casearia corymbosa, C. praecox y C. sylvestris, y Moraceae con Maclura tinctoria. Presentan una altura promedio de 6.1 m y 6.0 m2/hectárea de área basal, con presencia de especies importantes como Triplaris cummingiana y Spondias mombin. Presenta dominancia del estrato Arbolitos (5 a 12 m) con un 23.4%, seguido del estrato arbustivo (1.5 a 5 m) con 6.04% y del estrato Arbóreo Inferior (12 a 25 m) con 4.53%, siendo este su estrato menos abundante.
2. Arbustales cerrados con presencia de individuos arbóreos
En los levantamientos realizados en este bosque se registraron 242 individuos distribuidos en 45 especies, 41 géneros y 26 familias. Las familias que caracterizan florísticamente la zona son Fabaceae con individuos de la especie Machaerium capote principalmente, con presencia de otras especies importantes como Calliandra tolimensis, Albizia guachapele, Chloroleucon mangense, Erythrina poeppigiana e Inga bourgonii. También como familia importante se encontró la familia Rubiaceae, con individuos presentes de las especies Randia armata, Guettarda malacophylla y Simira cordifolia, y Salicaceae con Casearia corymbosa, Casearia mollis y Casearia praecox.
Estos arbustales presentan una altura promedio de 7.2 m y un área basal de 7.1 m2/hectárea, con presencia de especies importantes como Astronium graveolens, Hirtella americana y Spondias mombin. Presenta dominancia del estrato Arbustivo (1.5 a 5 m) con un 17.4%, seguido del estrato arbolitos (5 a 12 m) con 16.0% y del estrato Arbóreo Inferior (12 a 25 m) con 0.86%, siendo este su estrato menos abundante.
3. Bosques
individuos en 7 especies. Las especies más abundantes son Trichilia hirta con 211 individuos, Casearia praecox con 103 y Eugenia egensis con 72 individuos.Estructuralmente, este bosque se conforma por tres estratos, dentro de los que sobresale el estrato de Arbolitos (Ar) con dominancia de la especie Caseria praecox.
Selección de Especies
Teniendo en cuenta que las especies dominantes son las que aportan en una mayor proporción a los procesos ecosistémicos (Díaz et al. 2006), se ha propuesto que las especies que abarcan el 80% de la cobertura de un ecosistema son las que más contribuyen en la generación de servicios ecosistémicos (Cornelissen et al. 2003: Paqueman y Quested, 2007; Pérez-Harguidenguy et al. 2013). Tomando en cuenta este principio, a partir de la caracterización florística realizada por García et al. (2015) se determinó para la cada uno de los tres estados sucesionales evaluados, las especies que representan el 80% del área basal (dominantes), y que por tanto aportan en mayor grado al proceso de secuestro biológico de carbono.
Selección de Individuos
Los individuos seleccionados fueron en la medida de lo posible, árboles robustos, maduros y bien desarrollados, localizados en ambientes bien iluminados, de acuerdo a lo recomendado por Cornelissen et al. (2003), los árboles que presentaron afectaciones severas por herbivoría y patógenos se excluyeron cuando sus cantidades poblacionales lo permitieron. Se tomaron cinco individuos por especie en cada estado sucesional para los rasgos foliares y de madera, de acuerdo a lo recomendado por Cornelissen et al. (2003) y Pérez-Harguindeguy et al. (2013), para la determinación de la altura máxima y para la forma de crecimiento se tomaron 12 individuos. Determinación de Rasgos Funcionales
Tabla 1. Rasgos funcionales relacionados con el secuestro biológico de carbono Tipo de
Rasgo Nombre de Rasgo Unidad Fuente del Dato
Foliares AFE-Área foliar específica cm2/mg Recolección de muestras, medición en laboratorio
Foliares CFMS (Contenido Foliar de
Materia Seca) mg/g
Recolección de muestras, medición en laboratorio
Foliares Fenología Foliar Categoría Revisión bibliográfica y caracterización florística
Foliares Área Foliar (AL) cm2 Recolección de muestras, medición en laboratorio
Estructura CMST (Contenido de
Materia Seca del Tallo) mg/g
Recolección de muestras, medición en laboratorio
Estructura Densidad de madera g/cm3 Recolección de muestras, medición en laboratorio
Estructura Forma de crecimiento
(Hábito) Categoría
Revisión en campo, caracterización florística y bibliográfica
Estructura Altura Máxima m Medición en campo y caracterización florística
Todos los rasgos a excepción de la fenología foliar y la forma de crecimiento, fueron medidos en los meses de junio y julio, meses en los que se presentan precipitaciones medias mensuales entre 45 y 83 mm (IDEAM, 2017), haciendo parte de la estación seca.
Rasgos Foliares
A B C
D E F
Figura 2. A. Toma de muestras foliares. B. Hidratación. C. Etiquetado y almacenamiento en campo. D. Almacenamiento para laboratorio. E. Fotografías para cálculo de área foliar. F. Determinación de área foliar.
Área foliar (AF)
Es un rasgo particularmente útil para analizar la economía del carbono (Salgado-Negret et al., 2016), se asocia con diversos procesos biológicos y ambientales tales como el crecimiento, la fotosíntesis, la transpiración, la asignación de luz y la interceptación de luz (Kucharik et al. 1998), se ha demostrado ampliamente su importancia para la estimación del crecimiento vegetal y el potencial del rendimiento biológico, así como el uso eficiente de la radiación solar y los recursos hídricos (Sonnentag et al. 2008).
La medición del área foliar se realizó por medio del Software ImageJ (Schneider et al., 2012), para la determinación del área proyectada de cada hoja, previa calibración del instrumento con áreas conocidas por medio de la ubicación de una regla en las fotografías utilizadas (Figura 2F). Las unidades de medida fueron centímetros cuadrados (cm2). Cuando las especies presentaron hojas simples se midió la totalidad de la lámina, cuando se contó con hojas compuestas se midieron las láminas de los foliolos, excluyendo raquis y peciolos o peciolulos según cada caso.
Área Foliar Específica (AFE)
Posteriormente se utilizaron los datos de peso seco y área foliar de cada hoja para obtener el área foliar específica de cada hoja, se estimó el peso específico de acuerdo con lo propuesto por Veneklaas (1985). Empleando la siguiente fórmula:
AFE (g/cm2)= Psh/Aph (1)
Donde AFE: área foliar específica
Psph: Promedio de peso seco por hoja Aph: Promedio de área por hoja Contenido Foliar de Materia Seca (CFMS)
Es una medida que está relacionada de manera inversa con la tasa de crecimiento relativo, mostrando así una relación importante con respecto a la productividad de la planta (Pérez-Harguindeguy et al. 2013).
Para el cálculo de este rasgo se consideró el dato de peso en húmedo de la hoja (g), y posterior a la fase del secado de las muestras foliares a 70°C y por 48 horas, se pesaron las mismas para obtener la masa seca al horno (mg), con lo cual se calculó dicha relación, entendiendo que se calcula como la masa seca al horno, de una hoja dividida por su masa fresca saturada de agua (Pérez-Harguindeguy et al. 2013).
Fenología foliar
La fenología foliar influye de manera importante en la tasa fotosintética de la planta, las especies caducifolias suelen presentar un rápido crecimiento (Cornelissen, 2003) y tasas fotosintéticas altas (Easmus, 1999), es de tipo cualitativa y excluyente (perene, semicaducifolia o caducifolia), se determinó a partir de la revisión de densidad foliar de los individuos encontrados en campo tanto en época húmeda como en época seca.
Rasgos vegetativos
Forma de crecimiento (Hábito)
La forma de crecimiento identifica el estrato que ocupa una especie en el eje vertical, con lo cual determina su acceso a la luz y su tasa potencial de crecimiento (Pérez-Harguindeguy et al. 2013), se relaciona generalmente con la altura o el biotipo, a partir de este rasgos se pueden identificar los estratos que se encuentran en los diferentes tipos de vegetación, lo que sumado a la altura y a la cobertura de copa indica la dominancia energética de un determinado individuo de acuerdo a una posición determinada (Rangel y Velásquez, 1997)
Se determinó a partir de la consulta de bibliografía (Geilfus, 1989), (CATIE, 1994), (Figueroa y Galeano, 2007), (Scholz et al., 2008), (Gómez y Toro, 2008), (Rendón et al., 2013) y según la altura promedio que alcanzan los individuos dentro del área de estudio, siguiendo las categorías propuestas por Rangel y Lozano (1986): Arbóreo (Arbóreo superior (As) > 25 m, Arbóreo inferior (Ai) 25 – 12 m), Subarbóreo (Ar) 12 - 5 m, Arbustivo (Arb) 5 – 1,5 m, Herbáceo (H) 1.5 – 0.25 m y Rasante (R) < 0.25 m.
Densidad de la Madera (Dm)
Es entendida como la masa seca al horno dividida por el volumen verde de la misma, suele utilizarse para estimar el contenido de diversos elementos biogeoquímicos en la madera (Návar, 2009) entre ellos, el contenido de carbono (Chavé, 2002). Para la determinación de la densidad de la madera de una especie, se pueden utilizar un núcleo de madera extraído con un barreno o las ramas del árbol o planta (Salgado et al., 2016), a esta conclusión llegaron Swenson y Enquist (2008) al comparar los datos obtenidos en los 2 métodos descritos tanto a nivel de arbustos como de árboles. En campo se tomaron muestras de ramas de al menos 1 cm de diámetro de 5 individuos sanos por cada especie, previamente identificados. Se removió la corteza externa de cada una de las ramas para evitar la proliferación de hongos (Figura 3A) y se dimensionó a 5 cm de longitud aproximadamente (Figura 3B). Cada muestra se etiquetó y depositó en bolsas resellables para su traslado a laboratorio (Figura 3C).
C D
Figura 3. Toma de muestras vegetativas en campo. A. Descortezado. B. Dimensionado. C. Etiquetado. D. Almacenamiento para laboratorio.
Posteriormente se almacenaron las muestras debidamente etiquetadas, en agua con hipoclorito de sodio para evitar la proliferación de hongos (Figura 3D), de acuerdo a lo recomendado por Salgado et al. (2016).
En la fase de laboratorio se determinó la densidad básica, puesto que es la más utilizada en cálculos de biomasa. Se llevaron las ramas a contenido de humedad en punto de saturación de fibras y se utilizó la siguiente fórmula:
Db=Po/Vv (2)
Donde Db: Densidad básica Po: Peso anhidro Vv: Volumen verde
Con la muestra de madera en un contenido de humedad en el punto de saturación de fibras se realizó la medición del volumen verde, el cual se determinó con un método de medición indirecta de volumen “método de inmersión en agua”, este método consiste en tarar una probeta de plástico y llenarla de agua con la suficiente cantidad para que la muestra de madera sumergida no derrame el líquido, introduciendo la muestra en la probeta con la ayuda de una aguja, cuidando que la muestra de madera no entre en contacto con el recipiente, ya que esto alteraría la medición del volumen. La medición que muestra la probeta representa el volumen de la rama, esto se explica fácilmente debido a que el agua tiene una densidad de 1.0 g/cm3 (Chave, 2006). El peso anhidro (o peso seco) se midió a partir del pesaje de cada una de las probetas de madera una vez fueron extraídas del horno, una vez las muestras mantuvieron un peso constante, de acuerdo a lo recomendado por Pérez-Harguindeguy et al. (2013).
Contenido de Materia Seca del Tallo (CMST)
Para la medición de esta variable se utilizaron las mismas ramas utilizadas para la medición de la densidad de la madera, de tal manera que la colecta en campo fue la misma utilizada para la variable mencionada. En el laboratorio, la medición del peso en el punto de saturación de las fibras se llevó a cabo por medio del pesaje de la rama saturada de agua, se llevó a la estufa durante 48 a 72 horas y a una temperatura de 70-72 °C de acuerdo a lo recomendado por Pérez-Harguindeguy et al. (2013), posteriormente el peso en seco se determinó a partir de mediciones constantes del peso luego de las 48 horas.
Altura Máxima (Hmax)
La altura máxima expresa la posición de una determinada especie a través de un gradiente lumínico, debido a que de su altura depende en gran medida la capacidad de captación de la radiación directa, con lo cual se limita su capacidad para llevar a cabo los procesos fotosintéticos que determinan su capacidad de captura de CO2 (Pérez-Harguindeguy et al. 2013). Este rasgo es uno de los que mayor influencia tiene en la determinación del almacenamiento potencial de carbono por parte de un determinado individuo.
El valor del rasgo de altura máxima para cada una de las especies, se determinó con base en la información registrada en los levantamientos de la caracterización florística y estructural realizada por la García et al., (2015), y los levantamientos del diagnóstico silvicultural realizado por García et al., (2016), tomando en cuenta para cada especie el promedio del valor de sus 12 individuos más altos, de acuerdo a (Pérez-Harguindeguy et al. 2013).
Manejo de Datos
Considerando que se buscó determinar si se presentaban variaciones funcionales en el gradiente sucesional, tanto a nivel comunitario como a nivel de TFPs, se procedió a calcular a nivel comunitario índices monorasgos y multirasgo; y a nivel de TFPs la abundancia y área basal relativas de cada uno de los mismo.
Se estandarizaron los datos y a través de análisis multivariado, se realizó un análisis de conglomerados jerárquicos para determinar TFPs en la totalidad del gradiente sucesional. El método de encadenamiento utilizado fue el de Ward, y se utilizó la medida de distancia euclidea. Por medio de estadística inferencial, se evaluó si dichos grupos presentaban diferencias significativas entre sí por medio de un Análisis de Varianza Multivariado (MANOVA) utilizando la prueba de Hotelling.
La caracterización de los TFPs se realizó por medio de análisis de Tablas de contingencia para el caso de las variables categóricas, y Análisis de Componentes Principales (ACP) para evidenciar gráficamente relaciones entre los TFPs y los rasgos cuantitativos. Posteriormente se determinó si había diferencias significativas en los valores de los rasgos cuantitativos entre los TFPs a partir de comparaciones de medias con un análisis de varianzas producto de modelos lineales mixtos. Esta determinación se realizó por medio del software estadístico InfoStat (Di Rienzo et al., 2011). Se determinó si se presentaban diferencias significativas en las abundancias y dominancias de TFPs en cada estado sucesional por medio de comparaciones de medias con un análisis de varianza a partir de modelos lineales mixtos.
Legendre, 2010), y los propuestos por Villéger et al. (2008): Riqueza funcional (FRic), equidad funcional (FEve, por sus siglas en inglés) y divergencia funcional (FDiv). Esta determinación se realizó por medio del software estadístico F-Diversity (Casanoves et al., 2010). Por medio de comparaciones de medias a partir de un análisis de varianza a partir de modelos lineales mixtos, se determinó si había diferencias significativas en los valores de dichos índices entre los estados sucesionales evaluados.
RESULTADOS
Especies seleccionadas para cada estado sucesional
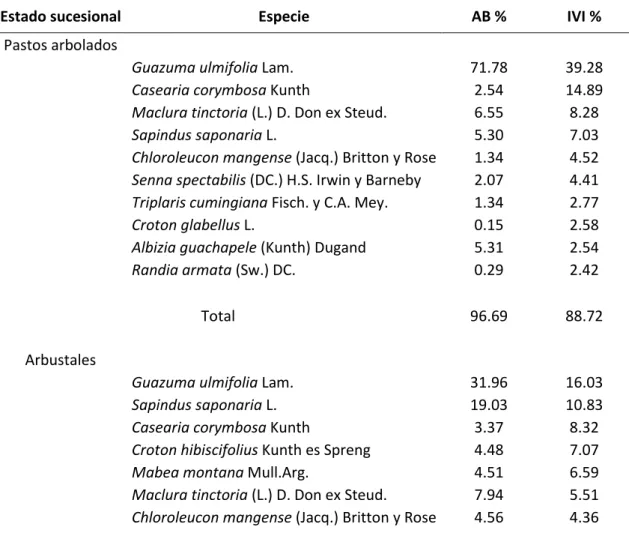
En cada uno de los tres estados sucesionales se obtuvieron 26 especies dominantes, las cuales representan más del 80% del área basal e índice valor de importancia de cada estado sucesional (Tabla 2). De estas, ocho especies pertenecen a los pastos arbolados, 21 especies a los arbustales y 13 al bosque propiamente dicho. Importante es señalar que, de las 26 especies evaluadas, tres especies se presentaron en los tres estados sucesionales evaluados, siete se encontraron presentes en dos estados sucesionales, y 16 se encontraron en un único estado sucesional, por lo que al sumar los biotipos encontrados en los tres estados sucesionales evaluados se obtuvo un total de 42.
Tabla 2. Especies seleccionadas para el estudio con base en su dominancia en cada estado sucesional. AB%: Área Basal Relativa. IVI%: Índice de Valor de Importancia Relativo.
Estado sucesional Especie AB % IVI %
Pastos arbolados
Guazuma ulmifolia Lam. 71.78 39.28
Casearia corymbosa Kunth 2.54 14.89
Maclura tinctoria (L.) D. Don ex Steud. 6.55 8.28
Sapindus saponaria L. 5.30 7.03
Chloroleucon mangense (Jacq.) Britton y Rose 1.34 4.52 Senna spectabilis (DC.) H.S. Irwin y Barneby 2.07 4.41 Triplaris cumingiana Fisch. y C.A. Mey. 1.34 2.77
Croton glabellus L. 0.15 2.58
Albizia guachapele (Kunth) Dugand 5.31 2.54
Randia armata (Sw.) DC. 0.29 2.42
Total 96.69 88.72
Arbustales
Guazuma ulmifolia Lam. 31.96 16.03
Sapindus saponaria L. 19.03 10.83
Casearia corymbosa Kunth 3.37 8.32
Croton hibiscifolius Kunth es Spreng 4.48 7.07
Mabea montana Mull.Arg. 4.51 6.59
Estado sucesional Especie AB % IVI % Triplaris cumingiana Fisch. y C.A. Mey 1.16 3.25
Machaerium capote Dugand 3.74 3.22
Guapira pubescens (Kunth) Lundell 0.68 3.09 Miconia spicellata Bonpl. Ex Naudin 3.73 3.05
Astronium graveolens Jacq. 1.92 2.77
Hirtella americana L. 0.69 2.75
Byrsonima crassifolia (L.) Kunth 1.65 2.54
Albizia guachapele (Kunth) Dugand 4.34 2.16
Croton glabellus L. 0.26 1.52
Cordia alliodora (Ruiz y Pav.) Oken 0.16 1.48
Randia armata (Sw.) DC. 1.00 1.23
Total 95.18 85.76
Bosques
Trichilia hirta L. 37.55 20.34
Casearia praecox Griseb. 19.78 18.07
Machaerium capote Dugand 7.67 9.15
Pseudobombax septenatum (Jacq.) Dugand 0.67 5.76 Triplaris cumingiana Fisch. y C.A. Mey 3.23 4.90
Guettarda malacophylla Standl. 3.63 3.77
Eugenia egensis DC. 7.13 6.40
Trichanthera gigantea (Humb. y Bonpl.) Nees 1.35 3.28 Simira cordifolia (Hook.f.) Steyerm 3.50 2.70
Astronium graveolens Jacq. 1.88 2.51
Randia armata (Sw.) DC. 1.21 1.96
Maclura tinctoria (L.) D. Don ex Steud. 0.81 1.82
Amyris pinnata Kunth 0.94 1.36
Total 89.37 82.03
Valores de Rasgos Funcionales
Dentro de los valores de rasgos encontrados, se evidenciaron rasgos con una alta variabilidad interespecífica, en especial para el caso de los rasgos foliares, presentándose en rangos entre 12.36 a 311.17 cm2 de Área Foliar, 184.5 a 450.4 mg/g de Contenido Foliar de Materia Seca, 86.33 a 208.72 cm2/g de AFE. Los resultados para los que se muestran los valores medios en el caso de variables cuantitativas, o categóricos en el caso de variables cualitativas en cada uno de los estados sucesionales, se pueden observar en el Anexo 4.
Determinación de Tipos Funcionales de Plantas (TFPs)
especies (Pseudobombax septenatum, Trichanthera gigantea y Maclura tinctoria). El TFP 3 se conforma por siete especies (Amyris pinnata, Cordia alliodora, Hirtella americana, Mabea montana, Eugenia egensis, Byrsonima crassifolia y Miconia spicellata). Por último, el TFP 4 se compone de nueve especies (Guapira pubescens, Randia armata, Croton glabellus, Guettarda malacophylla, Chloroleucon mangense, Senna spectabilis, Croton hibiscifolius, Guazuma ulmifolia y Casearia corymbosa).
Figura 4. Dendrograma de Tipos Funcionales de Plantas (TFPs)
Figura 5. Análisis de Componentes Principales (ACP) para rasgos funcionales cuantitativos y TFPs. AF: Área Foliar. AFE: Área Foliar Específica. CFMS: Contenido Foliar de Materia Seca. CMST: Contenido de Materia Seca en el Tallo. DM: Densidad de la madera. Hmax: Altura Máxima.
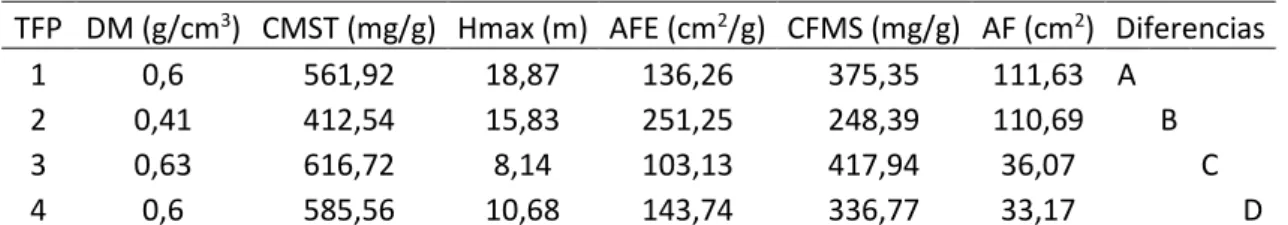
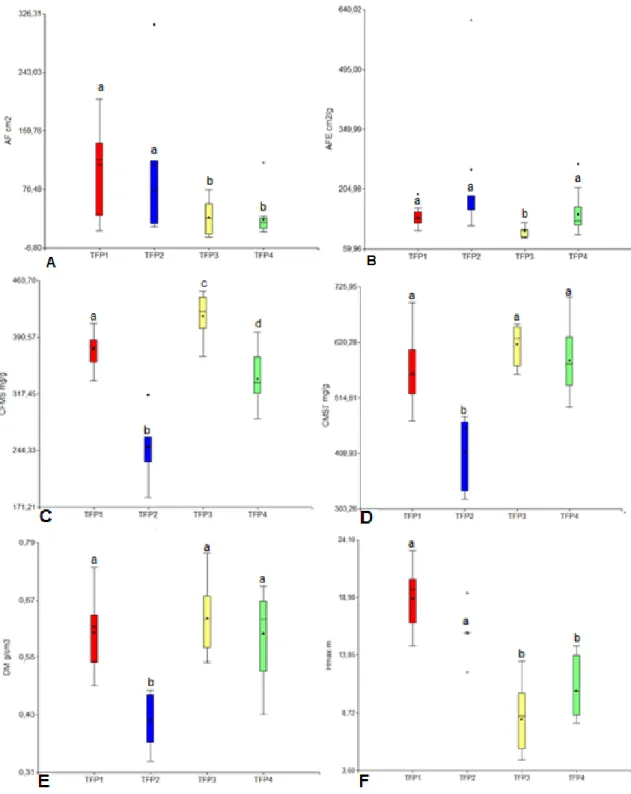
Tabla 3. MANOVA para comparación de TFPs a partir de Rasgos Funcionales. Medias con una letra común no son significativamente diferentes (P< 0.05). AF: Área Foliar. AFE: Área Foliar Específica. CFMS: Contenido de Foliar de Materia Seca. CMST: Contenido de Materia Seca en el Tallo. DM: Densidad de la Madera. Hmax: Altura Máxima.
TFP DM (g/cm3) CMST (mg/g) Hmax (m) AFE (cm2/g) CFMS (mg/g) AF (cm2) Diferencias
1 0,6 561,92 18,87 136,26 375,35 111,63 A
2 0,41 412,54 15,83 251,25 248,39 110,69 B
3 0,63 616,72 8,14 103,13 417,94 36,07 C
4 0,6 585,56 10,68 143,74 336,77 33,17 D
dispersión al interior de los grupos encontrados son la densidad de la madera y el contenido de materia seca en el tallo, mientras que el área foliar específica muestra una dispersión menos marcada al interior de los grupos evaluados.
Figura 6. Box-plots para Rasgos Cuantitativos por Tipos Funcionales de Plantas (TFPs). A: Área Foliar. B: Área Foliar Específica. C: Contenido de Foliar de Materia Seca. D: Contenido de Materia Seca en el Tallo. E: Densidad de la madera. F: Altura Máxima. Letras diferentes muestran diferencias significativas (p<0.05).
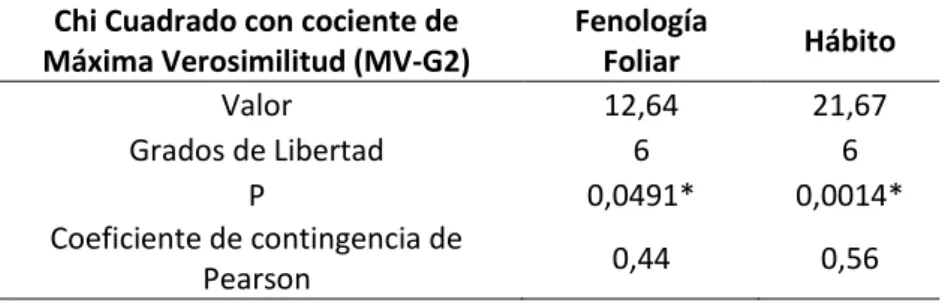
fenología semicaducifolia (P=0.0491), el TFP 3 al estrato arbustivo y la fenología perene, el TFP 4 al estrato arbustivo y la fenología caducifolia.
Tabla 4. Análisis de tablas de contingencia a partir de Chi Cuadrado entre TFPs, fenología foliar y forma de crecimiento. Asociaciones significativas, p<0.05(*)
Chi Cuadrado con cociente de Máxima Verosimilitud (MV-G2)
Fenología
Foliar Hábito
Valor 12,64 21,67
Grados de Libertad 6 6
P 0,0491* 0,0014*
Coeficiente de contingencia de
Pearson 0,44 0,56
Figura 7. Análisis de Correspondencias (AC) para TFPs, forma de crecimiento y fenología foliar
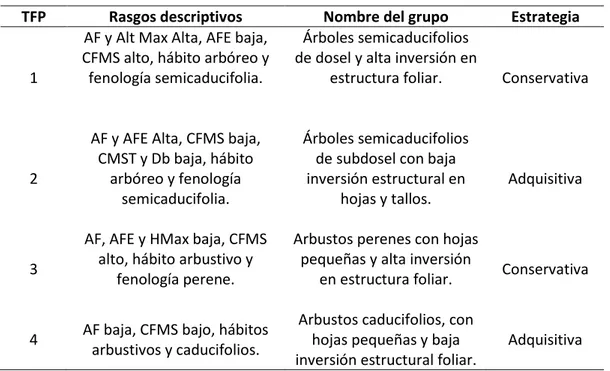
valores de contenido foliar de materia seca, alta área foliar específica, con tendencia a la caducifolía y densidad de la madera de media a baja (TFPs 2 y 4), y adquisitivas con altos valores de contenido foliar de materia seca, baja área foliar específica y con tendencia a mantener las hojas durante el año (TFP 1 y 3).
Tabla 5. Rasgos dominantes y nombre de los TFPs. AF: Área Foliar. AFE: Área Foliar Específica. CFMS: Contenido de Foliar de Materia Seca. CMST: Contenido de Materia Seca en el Tallo. DM: Densidad de la madera. Hmax: Altura Máxima.
TFP Rasgos descriptivos Nombre del grupo Estrategia
1
AF y Alt Max Alta, AFE baja, CFMS alto, hábito arbóreo y fenología semicaducifolia.
Árboles semicaducifolios de dosel y alta inversión en
estructura foliar. Conservativa
2
AF y AFE Alta, CFMS baja, CMST y Db baja, hábito
arbóreo y fenología semicaducifolia.
Árboles semicaducifolios de subdosel con baja inversión estructural en
hojas y tallos.
Adquisitiva
3
AF, AFE y HMax baja, CFMS alto, hábito arbustivo y
fenología perene.
Arbustos perenes con hojas pequeñas y alta inversión
en estructura foliar. Conservativa
4 AF baja, CFMS bajo, hábitos arbustivos y caducifolios.
Arbustos caducifolios, con hojas pequeñas y baja inversión estructural foliar.
Adquisitiva
Composición de TFPs en el gradiente sucesional
Figura 8. Abundancia relativa de Tipos Funcionales de Plantas (TFPs) en cada estado sucesional. Letras diferentes al interior de cada estado sucesional significan diferencias significativas (p<0.05).
Como se observa en la Figura 9, la abundancia de los TFPs cambia de manera significativa en el gradiente sucesional para todos los grupos a excepción de los árboles semicaducifolios de dosel (TFP 1, P=0.0691). Los cambios más relevantes se presentan en la abundancia de los TFPs asociados a estrategias adquisitivas, el TFP 2 (árboles semicaducifolios de subdosel con baja inversión estructural en hojas y tallos) y 4 (arbustos caducifolios, con hojas pequeñas y baja inversión estructural foliar), la cual es significativamente mayor en los pastos arbolados (P=0.0006 y P=0.0002, respectivamente), y se observa una disminución significativa de la misma en el gradiente sucesional.
Figura 9. Abundancia relativa de cada Tipo Funcional de Plantas (TFPs) en el gradiente sucesional. Letras diferentes al interior de cada estado sucesional significan diferencias significativas (p<0.05).
Al comparar el área basal relativa (Figura 10) de cada TFP en cada uno de los tres estados sucesionales, se encontró que en los pastos arbolados, los arbustos caducifolios, con hojas pequeñas y baja inversión estructural foliar (TFP 4) son significativamente los más dominantes (P=0.0001), mientras que los arbustos perenes con hojas pequeñas y alta inversión en estructura foliar (TFP 3) son significativamente los menos dominantes en este estado sucesional por no contar con individuos en el mismo.
Figura 10. Área Basal relativa de Tipos Funcionales de Plantas (TFPs) en cada estado sucesional. Letras diferentes al interior de cada estado sucesional significan diferencias significativas (p<0.05).
Figura 11. Área Basal relativa de cada Tipo Funcional de Plantas (TFPs) en el gradiente sucesional. Letras diferentes al interior de cada estado sucesional significan diferencias significativas (p<0.05).
De acuerdo a lo observado en la Figura 9 y en la Figura 11, el área basal se muestra menos sensible a los cambios sucesionales que la abundancia. Puesto que el TFP 4, conformado por arbustos caducifolios, con hojas pequeñas y baja inversión estructural foliar, presenta cambios significativos en su abundancia en el gradiente sucesional, lo cual no se muestra de una manera significativamente evidente en el área basal.
Índices de Diversidad Funcional
Como se evidencia en la Figura 12, al comparar los índices de diversidad funcional entre estados sucesionales se encontró que la media ponderada de la comunidad (CWM, por sus siglas en inglés) de todos los rasgos cuantitativos muestra un aumento (en ocasiones significativo) a medida que avanza la sucesión. Particularmente importantes se muestran los rasgos relacionados con la madera, tales como el contenido de materia seca en el tallo (CMST) y la densidad de la madera (DM), cuya media ponderada de la comunidad se incrementa de manera significativa a medida que se avanza la sucesión (P=0.0001, en los dos casos).
La media ponderada de la comunidad de los rasgos AF y Hmax muestran valores estadísticamente similares en los estados de pastos arbolados y arbustales; sin embargo, los mismos presentan valores significativamente más bajos que los presentados en los bosques (P=0.0001, en los dos casos). Por otro lado, el área foliar específica (AFE) y el contenido foliar de materia seca (CFMS) muestran valores similares en los arbustales y bosques, pero son significativamente mayores (P=0.0001, en los dos casos) a los encontrados en los pastos arbolados (Figura 12).
homogeneidad funcional (FEve) de los pastos arbolados y arbustales con los bosques, índice en el cual muestran valores significativamente más bajos (Figura 13, P=0.015).
Figura 13. Comparación de índices de diversidad funcional multirasgo entre estados sucesionales. FRic: Riqueza funcional. FEve: Homogeneidad funcional. FDiv: Diversidad funcional multirasgo. FDis: Dispersión funcional. Letras diferentes indican diferencias significativas (p <0,05).
DISCUSIÓN
Tipos Funcionales de Plantas asociadas a secuestro de carbono en Bs-T
Estas diferencias en la composición de los rasgos entre los grupos funcionales denotan de igual forma una singularidad en su aporte al servicio ecosistémico de secuestro de carbono (Lavorel et ál. 1997; Díaz y Cabido 2001; Fornara y Tilman 2009). En el caso del secuestro biológico de carbono, las especies con rasgos que favorecen la adquisición de recursos (adquisitivas) presentan una tasa de fotosíntesis más alta, reteniendo y liberando el carbono y otros elementos también de una manera más prematura (Grime, 1997; Reich et al., 1999). De manera contraria, los TFPs asociados a estrategias conservativas presentan tasas fotosintéticas y de captura de carbono más reducidas, pero una mayor retención y almacenamiento del carbono en sus estructuras, en especial en el fuste.
Realizar comparaciones de TFPs entre estudios no es una tarea sencilla (Hooper et al., 2005), debido a que se realizan presentando diferencias importantes en cuanto a escalas de trabajo, enfoques (Díaz et al., 2002; Casanoves, 2011). En contraste, resultados comparables se evidencian en estudios que realizan inferencias de acuerdo a índices de diversidad funcional (en especial monorasgo) (Conti y Diaz, 2013; Castellanos, 2013; Finegan et al., 2015; Lohbeck et al., 2015; López, 2015). Pese a lo anterior, Jiang y Dong (2000) identificaron TFPs en un gradiente climático de acuerdo a su capacidad para capturar carbono, encontrando como principal parámetro diferenciador entre grupos la forma de vida, rasgo que sirvió en el presente estudio para separar los TFPs 1 y 2 (árboles) de los TFPs 3 y 4 (arbustos).
De igual forma, Montes et al.,(2017) identificaron en bosques secos del caribe colombiano dos tipos funcionales de plantas, uno ligado a estrategias conservativas como baja AFE y alto CFMS, el cual se mostraría como un homólogo a los TFP 1 y 3 del presente estudio; mientras que el segundo presenta valores de rasgos asociados a estrategias adquisitivas (alta AFE y bajo CFMS), el cual por su composición de rasgos se mostraría como un homólogo a los TFP 2 y 4 del presente estudio, compartiendo de esta manera entre los dos estudios, dos de los rasgos que generaron mayores diferencias entre los grupos.
Si bien se identifican diferencias importantes entre los TFPs encontrados, y se evidencia como se ubican a lo largo de un eje de especialización en la adquisición e inversión de recursos, por lo que los valores de los rasgos de los TFPs no son completamente opuestos entre sí. Esto quiere decir, que se pueden encontrar rasgos tanto adquisitivos como conservativos al interior de cada grupo, evidenciándose así lo que se conoce como un desacoplamiento de los rasgos (Baraloto et al., 2010, Fortunel et al., 2012). Esto se puede observar de manera particular en el área foliar, en la cual al expresar valores altos se asoció tanto a estrategias conservativas como adquisitivas (TFPs 1 y 2), y la forma de crecimiento, en la cual los arbustales se asociaron tanto a estrategias conservativas como adquisitivas (TFPs 3 y 4), por lo que indicaría que se podrían encontrar especies con comportamientos intermedios entre adquisitivas y conservadoras. Lo encontrado confirma la necesidad de realizar estudios enfocados en los síndromes de rasgos, y no en su comportamiento individual entre cada uno de los grupos evaluados, para así lograr entender qué condiciones generan dicho desacoplamiento, o a determinar cuál podría ser el orden detrás de estos.
Variación de TFPs en el gradiente sucesional
De acuerdo a los resultados evidenciados, se observa que los arbustos caducifolios con hojas pequeñas y baja inversión en estructura foliar (TFP 4) son los más abundantes en los pastos arbolados, lo cual obedecería a una mayor capacidad de adaptación a condiciones que exigen alta eficiencia hídrica, expresada en rasgos que contribuyen a tolerar o evadir la sequía (Aranda et al., 2014), en este caso mediante la caducifolía (Salgado-Negret y Paz, 2016), o la reducción del área foliar, lo cual contribuye de manera significativa a disminuir la respiración y por tanto la pérdida de agua (Kucharik et al. 1998; Sonnentag et al., 2008), por lo que se podría decir que el principal filtro ambiental, que configura la composición de rasgos en la comunidad es abiótico, particularmente la disponibilidad de agua (Ceccon y Hernández, 2009). Pese a lo anterior, debe considerarse la influencia del hombre y en especial su perturbación en ecosistemas naturales como un motor que condiciona la trayectoria sucesional que tomará el ecosistema en cuestión (Foley et al., 2005), puesto que, en este caso particular, por ser un ecosistema sometido a ganadería intensiva (Avella et al., 2014) la especie más abundante y dominante en los pastos arbolados (Guazuma ulmifolia) no fue sujeta de aprovechamientos con el mismo grado de intensidad de otras especies comúnmente presentes en estados sucesionales iniciales en el área de estudio, debido a que fue utilizada como especie forrajera y para sombrío de ganado producto de la sombra que genera su copa.
Por otro lado, los resultados encontrados muestran que a excepción de los arbustos perenes con hojas pequeñas y alta inversión en estructura foliar (TFP 3) en los pastos arbolados, todos los TFPs se encuentran presentes (aunque varíen sus proporciones) en el gradiente sucesional. Al encontrar que en el gradiente sucesional disminuyó la abundancia de los TFP 2 (árboles semicaducifolios de subdosel con baja inversión estructural en hojas y tallos) y 4 (arbustos caducifolios, con hojas pequeñas y baja inversión estructural foliar), grupos que presentan como característica común una baja inversión estructural (Salgado et al., 2016) y altas tasas fotosintéticas (Reich, 1999, Wright y Westoby, 2002), debido a su madera blanda (TFP 2) y caducifolía (TFP 4), presentando por ende rápido crecimiento, se evidencia una disminución en la proporción de grupos funcionales relacionados con estrategias adquisitivas. Caso opuesto al crecimiento en la dominancia de los Árboles semicaducifolios de dosel y alta inversión en estructura foliar (TFP 1), este último, un rasgo común en especies con bajas tasas fotosintéticas (Salgado et al., 2016), cuya área basal aumenta en el gradiente sucesional.
Por otra parte, la presencia de especies de rápido crecimiento y valores altos de tamaño de copa y altura en los primeros estados sucesionales, particularmente Guazuma ulmifolia, se explicarían en la asignación temprana a la adquisición de recursos, para pasar posteriormente a la formación de materiales estructurales (Guariguata y Ostertag, 2001). Sin embargo, Lebrija-Trejos y Bongers (2008) no encontraron grupos de especies que pudiesen ser considerados como pioneras de larga vida o de rápido crecimiento al inicio de la sucesión, sino cuando ya se había presentado una etapa con presencia de arbustos y hierbas durante cerca de 10 años, diferencia que se entiende al presentar periodos de lluvia mucho más cortos y con condiciones más abruptas que las evidenciadas en el presente estudio. Pese a lo anterior, se puede observar una presencia importante de especies comunes en los pastos arbolados y los arbustales, lo cual llevaría a pensar que podrían ser especies que se presenten comúnmente en la etapa de arbustales, pero que en procesos de degradación por tala o quemas expresen sinergismos ante perturbaciones recurrentes (pisoteo y forrajeo producto de la presencia de ganado) y la capacidad de rebrote común en los bosques secos tropicales (Vesk y Westoby, 2004).
Variación de los rasgos de madera en el gradiente sucesional
Los dos rasgos cuyos valores promedio mostraron un aumento significativo al comparar los tres estados sucesionales evaluados fueron los relacionados con la madera (densidad y contenido de materia seca del tallo). Especialmente relevante es la densidad de la madera, considerando que fue un rasgo que presentó una variabilidad reducida al interior de cada uno de los estados sucesionales evaluados, y que presentó cambios importantes estadísticamente.
Resultados que concuerdan por lo encontrado por Castellanos (2013) en bosques secos tropicales del Caribe colombiano, en donde se evidenció un aumento en la abundancia de especies con maderas densas a medida que avanzaba la sucesión, por lo cual la densidad de la madera podría servir como un predictor de la capacidad de la vegetación de secuestrar carbono. Sin embargo, es necesario tomar en cuenta difiere de lo encontrado por López (2015), en bosques secundarios altoandinos en Colombia, donde no se evidenció una correlación significativa entre la acumulación de biomasa (la cual refleja cambios sucesionales) y la densidad de la madera, lo cual podría deberse a la diferencia en términos ambientales entre los dos estudios.
Variación de índices multirasgo en el gradiente sucesional
comparar bosques secundarios altoandinos tempranos y tardíos, pero no para los índices FDiv, FEve y FRic.
La estabilidad de los valores de los índices multirasgo se podría explicar de dos formas, las cuales no son mutuamente excluyentes entre sí, sino que, por el contrario, podrían llegar a ser complementarias. La primera teoría podría ser la del establecimiento en estados sucesionales avanzados de especies con valores de rasgos similares o ya presentes en la comunidad. Esta teoría podría evidenciarse en que, si bien los valores de rasgos varían a través de la comunidad, la presencia de la totalidad de los grupos es constante en todo el gradiente sucesional aun cuando varíen sus proporciones de abundancia o área basal (a excepción del TFP 1 en los pastos arbolados), con lo cual se entendería que el hipervolúmen de valores de rasgos que expresa con el índice de riqueza funcional se mantenga constante. De igual forma, la formación de cuatro grupos funcionales (TFPs) claramente definidos, con especies que presentan valores de rasgos similares entre sí, y que tendrían la misma respuesta a los cambios ambientales (Díaz y Cabido 2001, Díaz et al. 2011) implica de facto una alta redundancia funcional (Petchey y Gaston, 2009; Salmerón et al., 2016), la cual garantiza a partir de la presencia de un TFP determinado, que esos valores de rasgos se mantengan presentes en la sucesión y por tanto no se vean afectados los índices. Un ejemplo claro de lo anterior, es que pese a que la abundancia relativa del guácimo (Guazuma ulmifolia), perteneciente al TFP 4, es alta en los estados sucesionales iniciales como pastos arbolados y arbustales (Ver Anexos 1 y 2), se reduce progresivamente en el gradiente sucesional (Ver Anexo 3), por lo que se podría pensar que, en los bosques los valores de rasgos de Guazuma ulmifolia no tendrían una alta representatividad. Sin embargo, esta especie es reemplazada en los bosques por otra especie perteneciente al TFP 4 como Guettarda malacophylla, por lo que los valores de dichos rasgos se conservarían en la comunidad en su conjunto.
Por otro lado, una explicación alternativa a la baja variabilidad de los índices a través del gradiente sucesional es un recambio reducido de especies a través de la sucesión debido a su presencia en varios estados sucesionales, lo cual al considerar que de las 26 especies evaluadas, tres estaban presentes en todos los estados y ocho en al menos dos, y que precisamente el estado sucesional que mostró diferencias con los demás estados al comparar el índice de homogeneidad funcional (FEve) fue el bosque, al contar con la mayor cantidad de especies exclusivas de ese estado (ocho), comparado con los dos estados sucesionales restantes (pastos arbolados uno, y arbustales seis).
Un factor adicional que podría explicar la estabilidad en el gradiente sucesional de los índices multirasgo, podría ser la baja escala temporal de la sucesión evaluada, puesto que al haber sido ecosistemas intervenidos recientemente (5 años en el caso de los pastos arbolados, por ejemplo) (García et al., 2014), podrían llegar a ser menos evidentes los cambios presentes en el conjunto de la comunidad, factor que ha sido documentado en otros trabajos previamente (Lohbeck et al., 2013).
Lo anterior podría estar revelando que pese a presentar cambios en la composición y abundancia de los diversos elementos en el ecosistema, las especies que entran a participar de manera decisiva en diferentes estados de la sucesión, llegan a ocupar espacios vacíos en el nicho, dejados por otras que fueron eliminadas o las cuales disminuyeron su presencia en el gradiente sucesional.
la variabilidad de los atributos funcionales considerando los valores de las especies más abundantes (Casanoves et al., 2011), ni la diferenciación del nicho y por tanto la competencia (Mason et al., 2003; Mason et al., 2005) pese a presentar diferencias importantes en la composición y dominancia de las especies en cada estado sucesional.
Sería de esperarse que, debido a una mayor riqueza de especies la amplitud del nicho de la comunidad sea mayor en los bosques que en los estados sucesionales más tempranos, por lo cual, producto de una mayor estratificación vertical y a la variabilidad de los rasgos que generaría esta riqueza, se presente un mayor uso de la energía lumínica y el agua, generando así una mayor fotosíntesis y diferenciales de almacenamiento de carbono. Adicional a esto, Lebrija-Trejos y Bongers (2008) encontraron que, en bosques muy secos tropicales en México, variables como la densidad de individuos por hectárea, el tamaño de copas y la altura de dosel presentaron una recuperación más rápida a través de la sucesión que el área basal o la riqueza de especies. Sin embargo, se encontró que en el gradiente sucesional no se evidencia mayor variabilidad a nivel multirasgo ni en términos de su divergencia ni de su dispersión, y que tampoco hay una mayor amplitud del nicho comunitario, lo que implicaría que por presentarse un trade off (compensación) en estos atributos, no sea adecuada su utilización como indicador en lo que atañe a procesos ecosistémicos como el secuestro de carbono, pese a mostrar sensibilidad en otros aspectos importantes como la respuesta a factores ambientales por parte de la comunidad (Villeger et al., 2008).
Diversidad Funcional como indicadora de secuestro biológico de carbono
La utilización de la diversidad funcional como indicadora de ganancia o pérdida de servicios ecosistémicos o los procesos que los generan, ha sido relativamente reciente, y en consecuencia no ha sido estudiada de una manera tan profunda como otros métodos o mecanismos para inferir o calcular estos cambios. Sin embargo, de acuerdo a lo encontrado en este estudio, la abundancia/dominancia de tipos funcionales de plantas ligados a estrategias conservativas y adquisitivas es útil para aproximarse a la capacidad del ecosistema y sus diferentes estadios sucesionales para captar, liberar y conservar el carbono. Su utilidad no está determinada únicamente por no precisar de cálculos de cantidad de biomasa por hectárea para inferir diferencias entre estados sucesionales (lo cual contribuye de manera directa a la conservación), sino también porque se puede extender a otras especies, debido a que a partir de métodos estadísticos se pueden incorporar especies adicionales a la base con la que se cuenta.
El monitoreo a los procesos de restauración ecológica comúnmente se ha enfocado en evaluar la cantidad de hectáreas plantadas, individuos plantados y en ocasiones en la cobertura de los sitios intervenidos (Aguilar y Ramírez, 2015). Sin embargo, estos son parámetros en los que se evalúa realmente la gestión o intervención realizada, y no la reacción del ecosistema a este estímulo, por lo que se restringe dicha evaluación a la recuperación en términos de composición y estructura, pero no en la función que de estas puede derivar.
Considerando lo anterior, las especies asociadas a estrategias adquisitivas (TFPs 2 y 4) tales como Pseudobombax septenatum, Trichanthera gigantea, Maclura tinctoria, Guapira pubescens, Randia armata, Croton glabellus, Guettarda malacophylla, Chloroleucon mangense, Senna spectabilis, Croton hibiscifolius, Guazuma ulmifolia y Casearia corymbosa), podrían estar capturando mayores cantidades de carbono pero liberándolo también de una manera prematura. Mientras que especies asociadas a estrategias conservativas (TFPs 1 y 3) tales como Albizia guachapele, Sapindus saponaria, Triplaris cumingiana, Astronium graveolens, Machaerium capote, Casearia praecox, Simira cordifolia y Trichilia hirta, Amyris pinnata, Cordia alliodora, Hirtella americana, Mabea montana, Eugenia egensis, Byrsonima crassifolia y Miconia spicellata podrían estar aportando en un mayor grado al almacenamiento de carbono y a la acumulación de biomasa.
CONCLUSIONES
Se identificaron cuatro tipos funcionales de plantas (TFPs) con roles diferenciables producto de su composición de rasgos. Se agruparon en dos estrategias de inversión de recursos: dos grupos funcionales se asociaron a estrategias adquisitivas, los cuales presentan una tendencia a captar y liberar carbono de manera más prematura: Árboles semicaducifolios de subdosel con baja inversión estructural en hojas y tallos (TFP 2) y arbustos caducifolios, con hojas pequeñas y baja inversión estructural foliar (TFP 4).Por otro lado, los dos grupos restantes se asociaron a estrategias conservativas, aportando más en el almacenamiento que en la captura de carbono, siendo estos los árboles semicaducifolios de dosel y alta inversión en estructura foliar (TFP 1) y arbustos perenes con hojas pequeñas y alta inversión en estructura foliar (TFP 3).
Los grupos con estrategias adquisitivas (TFPs 2 y 4) mostraron una reducción en sus proporciones de abundancia en el gradiente sucesional, mientras que el TFP 1 (asociado a estrategias conservativas) se mantuvo estable, y el TFP 3 aumentó su abundancia en los arbustales pero la redujo en los bosques. El área basal relativa de los grupos de tendencia adquisitiva TFPs 1 y 3 mostró un aumento significativa en diferentes etapas de la sucesión, mientras se mantuvo constante para el TFP 2 y se redujo para el TFP 4. Esto sugiere una disminución en la proporción de especies adquisitivas que presentarán mayores tasas de capturo de carbono, mientras aumenta la proporción de especies conservativas con mayor potencial para almacenarlo; patrón que se ha encontrado en otros gradientes sucesionales mediante métodos destructivos y ecuaciones alométricas. Lo anterior sugiere que las proporciones de TFPs en gradientes sucesionales podrían ser consideradas como variables importantes para inferir el éxito o no en la ganancia de procesos ecosistémicos como el secuestro biológico de carbono, en procesos de restauración ecológica en el bosque seco tropical de la cuenca alta del río Magdalena.
Se encontró un incremento significativo en la media ponderada de la comunidad (CWM) de los rasgos cuantitativos evaluados en el gradiente sucesional, encontrando que son especialmente sensibles a estos cambios los rasgos en la madera: contenido de materia seca en el tallo y densidad de la madera, lo cual vislumbra la posibilidad de utilizar la densidad de madera como un indicador de éxito en procesos de restauración ecológica, considerando su baja variabilidad intraespecífica y cantidad de mediciones en bases de datos.
funcional (FRic), dispersión funcional (FDis) y divergencia funcional (FDiv), lo cual denotaría una especie de trade off de características funcionales a nivel comunitario pese a los evidentes cambios en composición y estructura.
RECOMENDACIONES
REFERENCIAS BIBLIOGRÁFICAS
• Aguilar, M. y Ramírez, W. (Eds). 2015. Monitoreo a procesos de restauración ecológica, aplicado a ecosistemas terrestres. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt (IIAvH). Bogotá DC. Colombia. 250 pp.
• Aranda, I., Ramírez-Valiente, J. y Rodríguez-Calcerrada, J. 2014. Características funcionales que influyen en la respuesta a la sequía de las especies del género Quercus: variación inter- e intra-específica. Ecosistemas 23 (2): 27-36
• Avella, A., García, N., Torres, S. 2014. Aproximaciones al ecosistema de referencia para los procesos de restauración ecológica del proyecto hidroeléctrico El Quimbo. Plan Piloto de Restauración Ecológica El Quimbo. Bogotá DC, Colombia.
• Baraloto, C., Timothe, C., Poorter, L., Beauchene, J., Bonas, D., Domenach,, A. 2010. Decoupled leaf and stem economics in rain forest trees. Ecology letters 13 (11): 1338-1347.
• Birch, J., Newton, A., Álvarez, C., Cantarello, E., Echeverría, C., Kitzberger, T., Schiappacasse, I. y Tejedor, N. 2010. Cost-effectiveness of dryland forest restoration evaluated by spatial analysis of ecosystem services. Proceedings of the National Academy of Sciences, 107(50): 21925 –21930.
• Carreño, G., Peña-Claros, M., Bongers, F., Alarcón, A., Licona, J. and Poorter, L. 2012. Effects of disturbance intensity on species and functional diversity in a tropical forest. Journal of Ecology 100: 1453–1463
• Casanoves, F., Di Rienzo, J., Pla, L. 2008. User manual FDiversity: statistical software for the analysis of functional diversity. First edition, Argentina. ‹www.fdiversity.nucleodiversus.org/›
• Casanoves, F., Pla, L., Di Rienzo, J., Díaz, S. 2010. FDiversity: un paquete de software para el análisis integrado de la diversidad funcional.
• Casanoves, F., Pla, L., Di Rienzo, J., Chan-Dzul, A., Bermeo, D., Delgado, D., y Finegan, B. 2011. Valoración y análisis de la diversidad funcional y su relación con los servicios ecosistémicos. In Valoración y análisis de la diversidad funcional y su relación con los servicios ecosistémicos. 9–31 pp.
• Castellanos, C. 2013. Functional analysis of secundary tropical dry forest in a región of the Colombian Caribbean. Thesis Doctor of Philosophy. Bounemouth University. 174 p
• CATIE. 1994. Laurel, Cordia alliodora (Ruiz y Pavón) Oken, especie de árbol de uso múltiple en América Central. Colección de guías silviculturales No. 16. CATIE, Turrialba, Costa Rica. 41 p.
• Ceccon, E. y Hernández, P. 2009. Seed raind dynamics following disturbance exclusion in a secondary tropical dry forest in Morelos, Mexico. Revista de Biología Tropical 2: 257-269
• Chavé, J. 2002. Medición de densidad de madera en árboles tropicales. Manual de campo. PAN-AMAZONIA. 7 pp.
• Chave, J., Muller, H., Baker, T., Easdale, T., Ter Steege, H. y Webb, C. 2006. Regional and phylogenetic variation of wood density across 2456 neotropical tree species. Ecological Applications 16: 2356–2367.
• Chave, J., Coomes, D., Jansen, S., Lewis, S., Swenson, N. y Zanne, A. 2009. Towards a worldwide wood economics spectrum. Ecology Letters 12: 351–366