PONTIFICIA UNIVERSIDAD JAVERIANA FACULTAD DE CIENCIAS
PROGRAMA DE POSGRADO MAESTRÍA EN CIENCIAS BIOLÓGICAS
DISEÑO DE UN MÉTODO DE DETECCIÓN MÚLTIPLE PARA LA IDENTIFICACIÓN DE ORGANISMOS GENÉTICAMENTE MODIFICADOS EN
ALIMENTOS PROCESADOS
ANDRÉS FELIPE VELA ROJAS
ÉNFASIS EN BIOTECNOLOGÍA AGRICOLA E INDUSTRIAL
PONTIFICIA UNIVERSIDAD JAVERIANA FACULTAD DE CIENCIAS
PROGRAMA DE POSGRADO MAESTRÍA EN CIENCIAS BIOLÓGICAS
DISEÑO DE UN MÉTODO DE DETECCIÓN MÚLTIPLE PARA LA IDENTIFICACIÓN DE ORGANISMOS GENÉTICAMENTE MODIFICADOS EN
ALIMENTOS PROCESADOS
ANDRÉS FELIPE VELA ROJAS
Director:
JAVIER DAVID CASTELLANOS PULIDO
MSc. Biotecnología
Trabajo de Grado presentado como requisito parcial para optar al título de:
MAGISTER EN CIENCIAS BIOLÓGICAS Énfasis en Biotecnología Agrícola e Industrial
NOTA DE ADVERTENCIA
“La Pontificia Universidad Javeriana no se hace responsable por los conceptos, criterios, opiniones y conclusiones expresados por sus alumnos, solo velará por que el trabajo no tenga ataques personales y únicamente se vea en él, el anhelo de buscar la verdad y justicia”.
DISEÑO DE UN MÉTODO DE DETECCIÓN MÚLTIPLE PARA LA IDENTIFICACIÓN DE ORGANISMOS GENÉTICAMENTE MODIFICADOS EN
ALIMENTOS PROCESADOS
Andrés Felipe Vela Rojas
___________________________ __________________________ Javier David Castellanos M.Sc. Natalhie Campos Reales Ph.D
Director del trabajo de Grado Jurado
________________________ ___________________________
Wilson Terán Pérez Ph.D. Martín Emilio Rodriguez M.Sc.
Jurado Jurado
_________________________ _____________________________ Dra. Ingrid Schuler PhD. Dr. Manuel Franco PhD.
Decana Académica Director de Posgrados
TABLA DE CONTENIDO
RESUMEN ... 14
ABSTRACT ... 16
1. INTRODUCCIÓN ... 17
2. ANTECEDENTES BIBLIOGRÁFICOS ... 19
2.1. Revolución Verde ... 19
2.2. Biotecnología moderna ... 19
2.3. Obtención de un organismo genéticamente modificado ... 20
2.3.1. Identificación y aislamiento del gen de interés ... 20
2.3.2. Inserción del segmento de ADN en la célula receptora ... 23
2.3.2.1. Transformación mediada por Agrobacterium. ... 25
2.3.2.2. Bombardeo de partículas. ... 25
2.3.2.3. Bigotes de Carburo de Silicio. ... 26
2.3.2.4. Transformación de protoplastos. ... 26
2.3.2.5. Electroporación de tejidos. ... 26
2.3.2.6. Inyección de ADN... 26
2.3.2.7. Vectores virales. ... 27
2.3.2.8. Nuevas técnicas de modificación genética ... 27
2.3.3. Regeneración y selección de los tejidos vegetales que han recibido el gen de interés ... 28
2.4. Caracterización del nuevo organismo vivo. ... 29
2.5. Clasificación de los OGM ... 29
2.5.1. Basada en el origen de los elementos insertados ... 30
2.5.2. Basada en la característica insertada ... 30
2.5.2.1. Organismos con características agronómicas mejoradas. ... 31
2.5.2.2. Organismos con mejoras en las características nutricionales. ... 34
2.5.3. Basados en el conocimiento de la información insertada ... 35
2.5.4. Basada en la clasificación legal. ... 35
2.6. Bases de datos con información de OGM ... 36
2.7. Requisitos legales para la comercialización de OGM en Colombia. ... 36
2.7.1. Legislación aplicable a alimentos ... 36
2.7.2. Legislación aplicable a Alimentos Genéticamente Modificados ... 37
2.8.1. Enfoque para la detección de OGM ... 40
2.8.2. Técnicas analíticas para la detección de OGM ... 42
2.8.2.1. Detección basada en expresión proteica ... 42
2.8.2.2. Detección basada en Ácidos Nucleicos... 43
2.8.2.2.1. PCR ... 44
2.8.2.2.2. PCR cuantitativa en tiempo real (qRT-PCR) ... 46
2.8.2.2.3. PCR simplex, multiplex y multi-target ... 48
2.8.2.2.4. Interpretación de resultados de una PCR tiempo real ... 50
2.8.2.2.5. Características de la PCR ... 52
2.9. Nuevas tendencias para la detección de OGM ... 54
2.10. Genotipificación... 55
2.11. Validación del método de Detección de OGM basado en PCR tiempo real ... 58
2.11.1. Modelo de validación y verificación de métodos de la EURL-GMFF ... 59
2.11.2. Verificación de métodos ... 60
3. OBJETIVOS... 62
3.1. Objetivo general ... 62
3.2. Objetivos específicos ... 62
4. MATERIALES Y MÉTODOS ... 63
4.1. Diseño del método Multi-target de maíz ... 63
4.2. Primers y sondas ... 63
4.3. Materiales de Referencia y certificados para la validación ... 64
4.4. Acondicionamiento de la muestra y Extracción de ADN ... 64
4.5. Condiciones de PCR-HRM ... 64
4.6. Verificación preliminar de primers y sondas ... 65
4.7. Estandarización de PCR-HRM ... 65
4.8. Validación de la Multi-target de maíz ... 66
4.9. Comparación de la Multi-target de maíz del INVIMA con otras Multi-target existentes ... 66
5. RESULTADOS Y DISCUSIÓN ... 67
5.1. Diseño del método multi-target de maíz ... 67
5.1.1. Definición del gen de referencia para la Multi-Target de maíz ... 67 5.1.2. Definición de los elementos específicos para el Screening de la Multi-Target de
5.1.3. Definición de los eventos para la detección Evento específica de la Multi-Target de maíz 72
5.2. Verificación preliminar de primers y sondas ... 76
5.2.1. Nuevas secuencias para p35S ... 81
5.2.2. Nuevo diseño del Screening para la Multi-target de maíz. ... 82
5.3. Estandarización de PCR-HRM ... 85
5.3.1. Parámetros de corrida de Melt Curve Genotyping ... 85
5.3.2. Variables a evaluar para la estandarización de la PCR-HRM ... 85
5.3.3. Parámetros de análisis de Melt Curve Genotyping para el Screening de la Multi-target. 88 5.3.4. Criterios de evaluación para la estandarización de la PCR-HRM ... 88
5.3.5. Definición de la concentración de ADN óptima para PCR-HRM ... 92
5.4. Validación de la multi-target de maíz. ... 102
5.5. Comparación de la multi-target de maíz del INVIMA con otras Multi-target existentes ... 117
6. CONCLUSIONES ... 120
7. RECOMENDACIONES ... 121
BIBLIOGRAFÍA ... 123
LISTA DE TABLAS
Tabla 1. Promotores comúnmente utilizados en la transformación genética. ... 226
Tabla 2. Promotores comúnmente utilizados en la transformación genética. ... 23
Tabla 3. Promotores, gen de interés y terminadores de los eventos aprobados para
consumo humano para Colombia. ... 24
Tabla 4. Eventos de transformación genética aprobados para consumo humano en
Colombia ... 36
Tabla 5. Bases de datos de OGM y sus enfoques principales. Información basada en ENGL 2011a. ... 40
Tabla 6. Actividades y criterios de aceptación de las características a evaluar para la verificación de método de Detección de OGM validado por la JRC bajo la metodología de pre-validación e inter-laboratorio. ... 64
Tabla 7. Concentraciones de trabajo de los primers y las sondas utilizadas en la
Multi-target de maíz. ... 69
Tabla 8. Gen de referencia reportado en el informe de validación de los eventos de transformación genética. Información basada en el EURL-GMFF y ENGL, 2010... 73
Tabla 9. Características de evaluación de adh1, hmgA y zSSllb como genes de referencia.
Basada en Scholdberg T., et. al., 2008). ... 74
Tabla 10. Promotores y terminadores de los eventos de transformación genética en maíz.
... 75
Tabla 11. Disponibilidad de Material de Referencia y Certificado de los eventos de transformación genética aprobados en el país. ... 77
Tabla 12. Total de eventos que componen los Stacks de las solicitudes al CTN Salud al 24 de Abril de 2013 ... 78
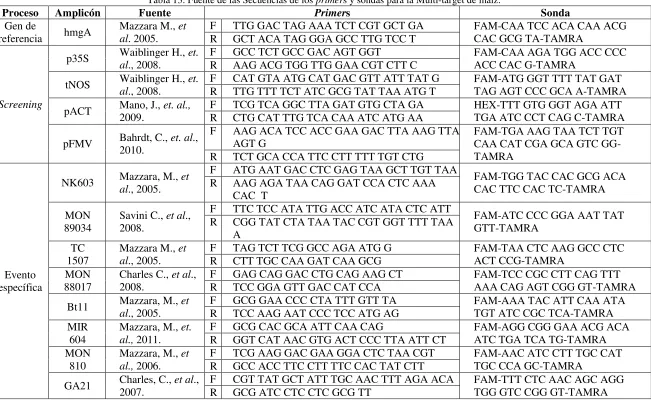
Tabla 13. Fuente de las Secuencias de los primers y sondas para la Multi-target de maíz. ... 81
Tabla 14. Valores de coeficiente de determinación R2 para evaluar el comportamiento de los métodos de detección de OGM por PCR tiempo real por sonda para la secuencia de p35S (Basada en Holden M. y colaboradores en 2010). ... 85
Tabla 15. Nueva secuencia para la detección de p35S (Zeitler R., et. al., 2002) ... 86
Tabla 16. Resultados de la prueba Hetero-Dimer por el analizador de secuencias
OligoAnalyzer 3.1 ... 87
Tabla 17. Opciones de picos para la metodología de ponderación de dispersión de picos.
... 97
Tabla 18. Evaluación de 40, 80, 120, 160 y 200 ng de ADN para PCR-HRM de pFMV.
Tabla 19. Evaluación de 40, 80, 120, 160 y 200 ng de ADN para PCR-HRM de p35S.100
Tabla 20. Evaluación de 40, 80, 120, 160 y 200 ng de ADN para PCR-HRM de tNOS.
... 101
Tabla 21. Evaluación de 40, 80, 120, 160 y 200 ng de ADN para PCR-HRM de pACT. ... 102
Tabla 22. Evaluación de 80, 100 y 120 ng de ADN para PCR-HRM de pFMV. ... 103
Tabla 23. Evaluación de 80, 100 y 120 ng de ADN para PCR-HRM p35S. ... 103
Tabla 24. Evaluación de 80, 100 y 120 ng de ADN para PCR-HRM de tNOS. ... 104
Tabla 25. Evaluación de 80, 100 y 120 ng de ADN para PCR-HRM de pACT. ... 104
Tabla 26. Especificidad del gen de referencia hmgA por PCR-HRM para la Multi-Target de maíz. ... 106
Tabla 27. Especificidad de los primers de los Dúplex pFMV-p35S y tNOS-pACT para los eventos dentro del alcance del Screening de la Multi-target de maíz. ... 109
Tabla 28. Especificidad de los primers Evento específicos para los eventos dentro del alcance del Screening de la Multi-target de maíz. ... 111
Tabla 29. Sensibilidad del gen de referencia hmgA por PCR-HRM para la Multi-Target de maíz. ... 114
Tabla 30. Sensibilidad de pFMV del Dúplex pFMV-p35S por PCR-HRM para la Multi-Target de maíz. ... 116
Tabla 31. Sensibilidad de p35S del Dúplex pFMV-p35S por PCR-HRM para la Multi-Target de maíz. ... 117
Tabla 32. Sensibilidad de tNOS del Dúplex tNOS-pACT por PCR-HRM para la Multi-Target de maíz. ... 118
Tabla 33. Sensibilidad de pACT del Dúplex tNOS-pACT por PCR-HRM para la Multi-Target de maíz. ... 118
LISTA DE FIGURAS
Figura 1. Ruta más común de creación de un OGM. Tomado de: Scheifler, Mertxe.
2009. ... 25
Figura 2. Niveles de detección de acuerdo a los constructos para identificar si es genéticamente modificado o no (Screening), que OGM es (Detección Evento Específica) o cuanta concentración de OGM tiene una muestra (Detección Cuantitativa) (NTWG, 2010). ... 45
Figura 3. Diagrama de temperaturas por tiempo de una PCR. Basado en Querci M., et. al., 2007. ... 49
Figura 4. Amplificación exponencial del ADN basado en PCR. Tomado de Querci M., et. al., 2007. ... 49
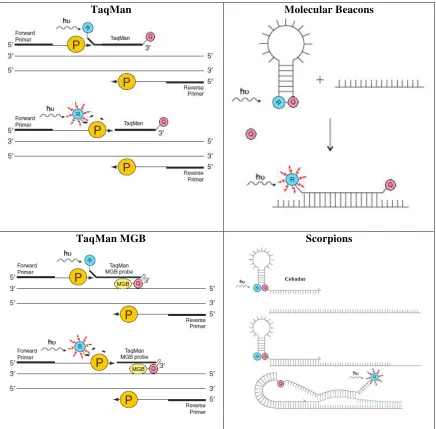
Figura 5. Tipos de sondas para PCR. Basado en Applied Biosystems, 2010; Querci M., et. al., 2007. R: Fluorocromo Reportero. Q: Fluorocromo Quencher. P: ADN Polimerasa. MGB: Enlazante del Surco Menor. ... 51
Figura 6. Formatos para la PCR. A. Formato simplex; B. Formato en dúplex; C. Formato en Multiplex; C. Formato Multi-Target. ... 53
Figura 7. Fluorescencia durante la amplificación. ... 55
Figura 8. Fluorescencia en función de los ciclos de amplificación y la regresión lineal de una curva de calibración... 56
Figura 9. Diagrama Fluoresencia porTemperatura y Derivada Negativa de la fluorescenia por temperatura para la identificación gráfica de la Tm. Tomada del ROCHE 2010a. .. 56
Figura 10. Procedimiento de PCR-HRM. A: Denaturación. B: Amplificación (Anillamiento y extensión). C. Curva de fusión. ... 57
Figura 11. Integración de la Multi-target de maíz al esquema de detección del laboratorio ... 69
Figura 12. Diseño de la Multi-target de maíz. Inhi: control de inhibición. Ext: control de extracción. ... 74
Figura 13. Verificación preliminar de p35S. ... 78
Figura 14. Verificación preliminar de pFMV. ... 80
Figura 15. Verificación preliminar de tNOS. ... 80
Figura 16. Verificación preliminar de pACT. ... 81
Figura 17. Nuevo diseño de la Multi-target de maíz con los dos Dúplex del Screening. 84 Inhi: control de inhibición. Ext: control de extracción. X: pozos libres. ... 84
Figura 19. Gráfica de segunda derivada para el programa de amplificación (a),
comportamiento de la fluorescencia al aumento de temperatura (b) y la derivada negativa de la fluorescencia (c) del dúplex tNOS-pACT. ... 88
Figura 20. Diagrama causa-efecto de acuerdo a las posibles contribuciones a la Medición de la Incertidumbre para los ensayos cuantitativos. Lo que se encuentra dentro del círculo son las contribuciones por reproducibilidad. Tomado del Procedimiento de estimación de la incertidumbre de los ensayos de cuantificación de OGM PA02-PM-OGM-P002
Versión 00. ... 90
Figura 21. Diagrama causa efecto de las posibles contribuciones a la incertidumbre en la medición volumétrica para el montaje de la detección de OGM. ... 90
Figura 22. Partes del pico esperado para HRM en la detección de OGM ... 92
Figura 23. Gráfica de segunda derivada para el programa de amplificación (a),
comportamiento de la fluorescencia al aumento de temperatura (b) y la derivada negativa de la fluorescencia (c) de la especificidad del gen de referencia hmgA. ... 103
Figura 24. Gel de la electroforesis de la amplificación de hmgA con: 1) Marcador de peso, 2)Non Template Contro (NTC) o control de reactivos, 3) Control -, 4) Arroz, 5) Soya, 6) Algodón, 7) MON89034xNK603, 8) MON88017, 9)NK603, 10) MON810, 11) GA21, 12) Bt11, 13) MIR604 y 14) TC1507 ... 104
Figura 25. Gráfica de segunda derivada para el programa de amplificación (a),
comportamiento de la fluorescencia al aumento de temperatura (b) y la derivada negativa de la fluorescencia (c) de la especificidad del dúplex pFMV-p35S. A. pFMV. B. p35S
... 105
Figura 26. Gráfica de segunda derivada para el programa de amplificación (a),
comportamiento de la fluorescencia al aumento de temperatura (b) y la derivada negativa de la fluorescencia (c) de la especificidad del dúplex tNOS-pACT. A. tNOS. B. pACT
... 106
Figura 27. Gráfica de segunda derivada para el programa de amplificación (a),
comportamiento de la fluorescencia al aumento de temperatura (b) y la derivada negativa de la fluorescencia (c) de la especificidad del método Evento específico de detección de MIR604 (A) y MON810 (B). ... 107
Figura 29. Gráfica de segunda derivada para el programa de amplificación (a),
comportamiento de la fluorescencia al aumento de temperatura (b) y la derivada negativa de la fluorescencia (c) de la especificidad del método Evento específico de detección de NK603 (A) y TC1507 (B). ... 109
Figura 30. Gráfica de segunda derivada para el programa de amplificación (a),
comportamiento de la fluorescencia al aumento de temperatura (b) y la derivada negativa de la fluorescencia (c) de la especificidad del método Evento específico de detección de GA21 (A) y Bt11 (B). ... 110
Figura 31. Gráfica de segunda derivada para el programa de amplificación (a),
Figura 32. Gráfica de segunda derivada para el programa de amplificación (a),
LISTA DE ANEXOS
Anexo 1. Datos de la especificidad de los hmgA por PCR-HRM para Multi-targer de
maíz. ... 145
Anexo 2. Datos de la especificidad de los Duplex pFMV-p35S y tNOS-pACT por PCR-HRM para Multi-targer de maíz. ... 146
Anexo 3. Datos de especificidad de los primers Evento específicos para los eventos
dentro del alcance de la Multi-target de maíz ... 147
Anexo 4. Datos de la sensibilidad de hmgA por PCR-HRM para el Gen de referencia de la Multi-Target de maíz. ... 147
Anexo 5. Datos de la sensibilidad de pFMV del Duplex pFMV-p35S por PCR-HRM para la Multi-Target de maíz. ... 147
Anexo 6. Datos de la sensibilidad de p35S del Duplex pFMV-p35S por PCR-HRM para la Multi-Target de maíz. ... 148
Anexo 7. Datos de la sensibilidad de tNOS del Duplex tNOS-pACT por PCR-HRM para la Multi-Target de maíz. ... 149
Anexo 8. Datos de la sensibilidad de pACT del Duplex tNOS-pACT por PCR-HRM para la Multi-Target de maíz. ... 149
RESUMEN
Uno de los resultados más importantes de la Biotecnología moderna para la agricultura, es el desarrollo de los Organismos Genéticamente Modificados (OGM). El ingreso de los OGM al mercado de los alimentos para consumo humano, ha generado un cambio acelerado en la forma de ver, comprender y controlar los alimentos en función de la seguridad alimentaria. Es tal el desarrollo en este campo del conocimiento, que se comercializan a nivel mundial eventos simples que son la expresión de una característica nueva a través de modificación genética y eventos apilados o Stacks que son la expresión
de dos o más características nuevas en comparación con su parental silvestre no modificado Hace aproximadamente dos años, era necesario detectar la presencia de eventos simples en alimentos y granos. Sin embargo, hoy en día, es necesario detectar múltiples eventos de transformación genética que pueda contener un alimento procesado o un grano en la fuente de almacenamiento. De acuerdo a lo anterior, se diseñó un esquema de detección eficiente el cual consta de cuatro pasos sucesivos: 1) Detección cualitativa del gen de referencia, 2)
Screening, 3) Detección evento específica y 4) Cuantificación del transgen. Para armonizar
el esquema de detección con la identificación cualitativa de múltiples eventos en alimentos procesados, articulado con la oportunidad de análisis de muestras del laboratorio, se hizo necesario diseñar una metodología robusta que permita la optimización de recursos, tales como tiempo, personal, consumibles, reactivos y uso de equipos, para la detección de múltiples eventos utilizando la técnica de Reacción en Cadena de la Polimerasa en tiempo real.
Esta es la primera aproximación a la detección de múltiples eventos de transformación
genética en maíz bajo un esquema de identificación altamente efectivo y eficiente. El alcance del método está dirigido a la detección cualitativa del gen de referencia de maíz
hmgA, a nivel de Screening con el promotor 35S del Virus del Mosaico de la Coliflor (p35S
CaMV), el promotor 35S del Virus del mosaico del higo (pFMV), el terminador NOS de la nopalina sintasa de Agrobacterium tumefacines (tNOS) y el promotor de la Actina del
Arroz (pACT) y a nivel Evento específico de los eventos NK603, MON810, MON88017, MON89034, Bt-11, GA21, MIR604 y TC1507 los cuales son eventos aprobados para consumo humano para Colombia.
El objeto del método es detectar ya sea de manera escalonada (primero gen de referencia,
luego del Screening y finalmente identificación Evento Especifica) ó única, en la cual se
puede identificar el gen de referencia, el Screening y el evento especifico que contienen la
muestra en la misma corrida utilizando PCR en tiempo real.
La técnica se diseñó bajo la metodología de la Multi-target de PCR, lo cual significa que se adicionan primers y sondas diferentes a cada pozo o grupos de pozos para poder identificar
distintos amplicones anillando eficientemente a las mismas condiciones de tiempo y temperatura. Por otro lado, para aumentar la eficiencia del método y demostrar que el método es fácilmente actualizable, se simplificó el Screening implementando dos Dúplex
o del 50% en los amplicones del Screening, según sea el caso. Finalmente, se implementó
el análisis del perfil de fusión por High Resolution Melting para el control de falsos
positivos y falsos negativos. La técnica PCR-HRM demostró ser altamente confiable para determinar productos inespecíficos de amplificación y algunos falsos negativos a partir del análisis de segunda derivada, convirtiéndose en la primera aproximación para la detección
de OGM.
La técnica de PCR-HRM por PCR Multi-target integrada con PCR Multipex, demostró ser eficiente por la baja inversión para lograr la validación de 13 analitos de detección cualitativa de OGM, sensible con un Límite de Detección (LOD) absoluto de 74 copias para la detección del Gen de Referencia y un LOD relativo de 0.1% para el Screening y la
ABSTRACT
The technology derived from recombinant ADN produced a rapid change in the way that we control food and understand it. Two years ago, it was necessary to detect the presence of simple events in foods or grains. However now, it is necessary to detect multiple genetic transformation events than can compose a processed food or in the grain storage source. According to the above, an efficient detection scheme has been designed which consists of four sequential steps: 1) qualitative detection of the reference gene, 2) Screening, 3)
specific event detection and 4) Quantification of the transgene. To harmonize this detection scheme in processed foods, taking into account the laboratory analysis opportunity and sampling plans at ports and border crossings, it was necessary to design a robust methodology that enables the optimization of resources, defined as time, staff, supplies, reagents and equipment used for the detection of multiple events using the Quantitative Real Time Polymerase Chain Reaction (qRT-PCR) technique.
This is the first approach to detect multiple genetic transformation events in maize with a highly effective and efficient system of identification. The scope of the method is directed to the qualitative detection of the reference gene of maize hmgA, Screening level with
Cauliflower Mosaic Virus p35S promoter (p35SCaMV) figworth mosaic virus promoter (pFMV), the terminator NOS to nopaline synthase from Agrobacterium tumefacines
(tNOS) and actin promoter Rice (pACT) and of the specific events NK603, MON810, MON88017, MON89034, Bt11, GA21, MIR604and TC1507, which are approved for human consumption for Colombia.
The aim of the method is detecting though a stepwise approach (first reference gene,
Screening finally specific event detection) or a single approach, which identifies the
reference gene, the specific event Screening and in the sample in the same run using real
time PCR.
The technique was designed based on the Multi-targetPCR methodology, which means that different probes and primers are added to each well or group of wells to efficiently identify
different amplicons at the same conditions of time and temperature. In the other hand, to increase the efficiency of the method and to show the upgradeability of the method, the
Screening was simplified by the implementation of two Duplex Reactions (pFMV-p35S
and tNOS-pACT) causing a decrease in the use of the master mix near 16% and allowing an expansion of the events identification scope in 25% or 50% in the Screening amplicons,
as applicable. Finally, a profile analysis by High Resolution Melting fusion was implemented to control false positives and false negatives. HRM PCR technique proved to be highly reliable to determine nonspecific amplification products and some false negatives from second derivative analysis.
1. INTRODUCCIÓN
Históricamente, la agricultura ha sido la estrategia principal de subsistencia humana (Ladizinsky G., 1998). Desde el desarrollo de la agricultura, los seres humanos han buscado mejorar los cultivos que son de su interés. Estos cambios se han producido a través de la selección, ya sea consciente o no, de los rasgos específicos que sean ventajosos para su consumo o para su cultivo (Mantell S., et al., 2008). Inicialmente, la comparación
entre las diferentes variedades de un mismo cultivo, generó la selección de algunas especies con capacidades para ser cultivados en una amplia variedad de ambientes (Benitez A., 2005). Principalmente, la base genética para la mejora tradicional de las plantas se constituía en las mutaciones aparecidas espontáneamente, la recombinación genética a partir de cruzamientos intra e interespecíficos y la poliploidía (Benitez A., 2005). De esta manera fue como el cruzamiento tradicional de especies apalancó la evolución de los cultivos modernos en el mundo (Mantell S., et al., 2008). Ya en el siglo XX, el desarrollo
de nuevas herramientas da origen a la Biotecnología moderna siendo uno de sus principales exponentes los Organismos Genéticamente Modificados (OGM). Estos OGM son organismos vivos cuya composición genética ha sido alterada por medio de la tecnología genética (Miraglia M., et. al., 2004) generándole características diferentes a su contraparte
isogénica no modificada. Luego del ingreso de los OGM a las cadenas de fabricación de alimentos, se genera una preocupación mundial acerca de qué posición, el análisis de información y la detección de estas modificaciones genéticas. Colombia por su parte, delega al Laboratorio Central Interinstitucional de Detección y Monitoreo de OGM y específicamente al INVIMA para que realice la detección de OGM en productos para consumo humano. El laboratorio por parte del INVIMA, diseñó un esquema de detección basado en la cadena de valor y en el flujo analítico que tiene la muestra a partir del momento que se le entrega al analista hasta la interpretación de resultados del ensayo. Desde el punto de vista analítico, las muestras son acondicionadas para lograr normalizar lo más posible sus características físicas previas a la extracción. Este paso consiste principalmente, en retirar la mayoría de fuentes de inhibición o submuestras que pueda alterar el resultado del ensayo, garantizando la viabilidad del ADN. Luego de ese proceso, se procede a una molienda/homogenización para garantizar la homogeneidad y el tamaño de partícula idóneo para realizar una extracción de ADN óptima en criterios de concentración y calidad del analíto de interés. Una vez se cuente con el ADN óptimo para la detección, se procede a hacer la amplificación de acuerdo al objeto del análisis solicitado por el cliente.
El cliente puede buscar únicamente la identificación de si su muestra es genéticamente modificada o no, por lo cual lo más recomendado es realizar una prueba de Screening con
evento tienen su muestra y necesita saber la concentración específica a la cual se encuentra. Para estos casos, el laboratorio realiza la Detección Cuantitativa a partir de protocolos validados.
De la misma manera, el cliente puede solicitar la prueba de Screening acompañada de una
2. ANTECEDENTES BIBLIOGRÁFICOS
2.1. Revolución Verde
La necesidad de producir alimento para una mayor población, ha hecho que el hombre desarrolle herramientas que permitan aprovechar al máximo las tierras destinadas para el trabajo agrícola (Mertxe S., 2010). Ya en el siglo XX, la Agricultura se inició a soportar fuertemente en la ciencia (Instituto Novel de Noruega, 2000). En este sentido, Fritz Harber y Carl Bosch encontraron la manera de sintetizar químicamente el amoniaco para el desarrollo de fertilizantes nitrogenados, Paul Muller descubre las propiedades insecticidas del DDT y Norman Borlaug desarrolla el trigo de alta productividad resistente a la roya (Mertxe S., 2010). Por su parte, en México se inició una revolución silenciosa liderada por Norman Borlaug en 1943 denominada la Revolución Verde. Luego de desarrollar plantas resistentes a la roya y otras enfermedades, era necesario desarrollar plantas que mostraran buena respuesta a fertilizantes sintéticos debido a que las tierras destinadas para cultivo en el Tercer Mundo, habían sido explotadas sistemáticamente por siglos de agricultura tradicional y era necesario fertilizarlas para hacerlas productivas. Adicionalmente, el uso de fertilización completa de los cultivos, hacía que un aumento en la cantidad de granos colapsara el tallo de la planta. Por tal razón, Borlaug decidió buscar una variedad de trigo más pequeña que pudiera resistir más eficientemente la producción. El nuevo híbrido resultó ser un trigo altamente adaptable con tallo fuerte y corto. Para el inicio de la década de los 60s, en la inmensa cantidad de tierras cultivables en México crecía trigo enano lo cual aumentó la cosecha 6 veces a comparación de las presentadas en 1944. Rápidamente, la revolución verde se esparció por Asia en países que se encontraban al borde del colapso alimentario.
De la misma forma en que se desarrolló el trigo de mayor producción, en Filipinas se estableció el Instituto Internacional de Investigación de Arroz IRRI (por sus siglas en inglés) el cual desarrolló el cultivar IR. Este cultivar de arroz duplicó la producción de arroz en dos décadas por el uso de fertilizantes y pesticidas indicados por el instituto de investigación.
2.2. Biotecnología moderna
El efecto de las técnicas de la Biotecnología Moderna en la agricultura ha sido el ampliar la gama de material hereditario que puede ser utilizado por la agricultura convencional para la mejora genética de plantas, animales y microorganismos (Persley G., et al., 1993). Ese
material se puede insertar, utilizando las herramientas de la Biotecnología Moderna, con una mayor precisión y en menor tiempo a comparación del uso de la agricultura tradicional (Persley G., et al., 1993). Internacionalmente, el protocolo de Cartagena define la
que superan las barreras fisiológicas naturales de la reproducción o de la recombinación y que no son técnicas utilizadas en la reproducción y selección tradicional”.
Uno de los resultados más importantes de la Biotecnología moderna para la agricultura, es el desarrollo de los Organismos Genéticamente Modificados (OGM). Existen muchas definiciones de OGM, sin embargo se podría resumir, que los OGM son organismos vivos cuya composición genética ha sido alterada por medio de la tecnología genética (Miraglia M., et. al., 2004) generándole características diferentes a su contraparte isogénica no
modificada. Cada nueva modificación genética se conoce como un evento y la combinación de varios rasgos transgénicos en una planta se conoce como evento apilado o
Stack (Stein A. y Rodríguez-Cerezo E., 2009).
2.3. Obtención de un organismo genéticamente modificado
La obtención de un OGM se basa en el uso sistemático de tres desarrollos científicos del ADN recombinante:
1. Identificación y aislamiento del gen de interés.
2. La inserción del segmento de ADN en la célula receptora.
3. Regeneración y selección de los tejidos vegetales que han recibido el gen de interés (Mertxe S., 2010).
Este proceso puede diferir dependiendo de la técnica de inserción del segmento de ADN y la necesidad de regeneración de tejidos. Sin embargo, en la figura 1, se puede observar un proceso general de transformación genética para la obtención de un OGM. Es importante aclarar, que en el paso 7 de la figura 1, se tiene implícito un fitomejoramiento de las plantas modificadas genéticamente.
2.3.1. Identificación y aislamiento del gen de interés
Principalmente, se han desarrollado OGMs con tolerancia a herbicidas, resistencia a enfermedades e insectos, mejora de calidad y tolerancia a estrés abiótico (Griffiths K., et al., 2004). Para conferirle estas nuevas características a los productos, es necesario
identificar los genes de interés, los cuales, pueden ser identificados de cualquier tipo de organismo. Por ejemplo, se pueden identificar genes de interés en microorganismos como los genes que codifican las proteínas de la familia Cry de Bacillus turingiensis, las cuales
enzimas de restricción, las cuales son útiles para el corte y ensamble en el vector de transformación (Walker J. y Gingold E., 1997).
Figura 1. Ruta más común de creación de un OGM. Tomado de: Mertxe S., 2010.
La secuencia aislada y purificada, generalmente se ensambla o clona en un plásmido el cual cuenta con el promotor y terminador escogido de acuerdo al rasgo genético a mejorar buscado en el OGM (Mertxe S., 2010).
Para lograr obtener el rasgo deseado en la planta transgénica, no solo es necesario utilizar el método de transformación óptimo, sino que es preciso definir qué tipo de secuencia de interés es la óptima, y cómo su expresión será regulada dependiendo del promotor y terminador elegidos. Con eso, se busca determinar en qué tejido de la planta se desea la expresión del transgen asociado al nuevo rasgo (Streatfield S., et al., 2010).
Los promotores son secuencias de ADN utilizadas para controlar la expresión de los genes introducidos, en búsqueda de la sobre expresión en algunos casos, o para la síntesis de marcadores de selección de la transformación (Potenza C., et al., 2004; Streatfield S., et al.,
2010). Los terminadores son secuencias que codifican señales de finalización de la transcripción (Hemmer W., 1997; Holden M., et al., 2010).
Hasta ahora, los promotores más utilizados para las transformaciones genéticas son el Promotor 35S del Virus del Mosaico de la Coliflor (CaMV), el Promotor de la actina del arroz 1 (pRACT) y el Promotor 35S del Virus del Mosaico del Higo (pFMV). El terminador utilizado comúnmente como parte del inserto en modificaciones genéticas es el Terminador de la secuencia de la Nopalina Sintasa de Agrobacterium tumefaciens (tNOS)
ampliamente utilizados para la transformación genética de plantas, se encuentran en la tabla 1 y 2.
Tabla 1. Promotores comúnmente utilizados en la transformación genética.
Promotor Descripción Expresión Referencias
Adh1 deshidrogenasa de maíz. Promotor de la Alcohol Constitutiva
Terada R., et al., 2000; Prieszner J., et al., 2001; Piquemal J., et al.,
2002; Jones H. y Shewry P., 2009.
ZmUbi1 Promotor de la Ubiquitina 1
de maíz. Constitutiva
Christensen A., et al., 1992; Jones
H. y Shewry P., 2009; Park S., et al., 2010; Stewart C., et al., 2011.
RACT Promotor de la actina del arroz. Constitutiva
McElroy D., et al., 1990; Xu D., et al., 1996b; Jones H. y Shewry P.,
2009; Park S., et al., 2010.
CaMV 35S Promotor 35S del virus del
mosaico de la coliflor. Constitutiva
Odell J., et al., 1985; Jones H. y
Shewry P., 2009.
H2B Promotor de la histona 2
del maíz. Constitutiva
Ping Y., et. al., 1995; Rasco-Gaunt
S., et al., 2003; Jones H. y Shewry
P., 2009
Hmwg alto peso molecular del trigo. Promotor de la Glutenina de Endospermo específica Norre F., Shewry P., 2009. et al., 2002; Jones H. y
ZmGlb1 Promotor de la Globulina 1 de maíz.
Semilla específica
Mei C., et al., 2004;Streatfield S., et al., 2010.
RUBQ1 Promotor de la Ubiquitina 1 del arroz. Constitutiva Wang J. y Oard J., 2003; Park S., et
al., 2010.
OsCc1 Promotor de la citocromo c1 de
Oryza sativa. Constitutiva
Jang I., et al., 2002; Park S., et al.,
2010; Stewart C., et al., 2011
pFMV Promotor 35S del virus del
mosaico del higo. Constitutiva
Maiti I. et. al., 1997;
Govindarajulu M., et al., 2008.
El p35S de CaMV, fue descrito inicialmente por Odell y colaboradores en 1985, es un promotor constitutivo de alta expresión para dicotiledóneas con baja actividad en monocotiledoneas, usado en maíz, arroz, algodón y soya modificada genéticamente (Hemmer W., 1997; Jones H. y Shewry P., 2009; Park S., et al., 2010; Stewart C., et al.,
2011). De acuerdo a la base de datos del Centro para la Evaluación del Riesgo Ambiental (CERA por sus siglas en ingles, http://cera-gmc.org/index.php?action=gm_crop_database ), el 66% de los eventos contienen al menos una copia de p35S en su genoma.
El pRACT, descrito inicialmente por McElroy y colaboradores en 1990, es un promotor constituvo de alta expresión debido a que es un componente fundamental del citoesqueleto de las cédulas de las plantas en monocotiledoneas y usado en dicotiledóneas con buenos resultados (McElroy D., et al., 1990; Park S., et al., 2010). Adicionalmente, el promotor
puede tener expresión transitoria o permanente si se encuentra acompañado del intrón (Peremarti A., et al., 2010). De acuerdo a la base de datos de CERA, el 14% de los eventos
El pFMV caracterizado inicialmente por Sanger y colaboradores en 1990, es un promotor constitutivo de alta expresión utilizado para la transformación de monocotiledoneas y dicotiledóneas (Bhattacharyya S., et al., 2002; Peremarti A., et al., 2010). De acuerdo a la
base de datos de CERA, el 11.33% de los eventos contienen al menos una copia del pFMV.
Tabla 2. Terminadores comúnmente utilizados en la transformación genética.
Terminador Descripción Referencias
NOS
Terminador de la Nopalina sintasa de
Agrobacterium tumefaciens.
Depicker A., et al., 1982; Bevan M., et al., 1983
35S CaMV Terminador 35S del Virus del mosaico de la coliflor.
Franck A., et. al., 1980; Pietrzak
M., et. al., 1986.
PINII proteinasa de la papa Inhibidor II de la Keil M., et al., 1986; An G.,
et.al., 1989.
Glb1 Globulina I del maíz Terminador de la Lucas D., T., et al., 2007; Kawakatsu
et. al., 2012.
tahsp17
Región no transcrita de la proteína de choque térmico 17.3 del trigo
McElwain E. y Spiker S., 1989; Vaughn T., et. al., 2005.
ORF25
Señal de poliadenilación ORF25 de Agrobacterium
tumefaciens.
Barker R., et. al., 1983; Meilan R., et. al., 2000.
7S ´3
Señal de poliadenilación del gen de la subunidad
alfa de la beta-conglicinina de la Soya
Schuler M., et al., 1982; Betz, F.,
et. al., 2000.
T-E9
Región no transcrita del gen de la ribulosa-1,5-bifosfato carboxilasa sub
unidad E9 de arveja.
Nida D., et. al., 1996;El-Gewely,
2004.
El tNOS, es la secuencia más ampliamente utilizada para los constructos de los OGM (Hemmer W., 1997; Holden M. et al., 2010). De acuerdo a la base de datos de CERA, el
52.67% de los eventos contienen al menos una copia del tNOS.
Los eventos aprobados para consumo humano por el Ministerio de Protección Social para Colombia, tienen los promotores y terminadores que se encuentran en la tabla 3.
2.3.2. Inserción del segmento de ADN en la célula receptora
Tabla 3. Promotores, gen de interés y terminadores de los eventos aprobados para consumo humano para Colombia.
Nombre del Evento Constructo
Promotor Gen de interés Terminador
MON810 p35S CaMV cry1Ab
Ninguno. Región que se perdió durante la integración
NK603 p-RACT CP4epsps tNOS
p35S CaMV CP4epsps tNOS
TC1507 p35S CaMV pat t35S
ZMUbi1 cry1Fa2 ORF25
Bt11 p35S CaMV pat tNOS
p35S CaMV cry1Ab tNOS
MON 89034 p35S CaMV cry1A.105
Región no traducida de la proteína de choque térmico 17.3 del trigo
p35S FMV cry2Ab tNOS
LY 038
ZMGlb1, intrón de RACT y secuencia transito del cloroplasto DHDPS
cordapA Región no traducida de ZM1Gbl1
MON 863 p35S CaMV (4-A51) cry3Bb1
Región no traducida de la proteína de choque térmico 17.3 del trigo
p35S FMV nptll tNOS
MON 88017
pRACT CP4epsps tNOS
p35S CaMV con la región
duplicada de unión cry3Bb1
Región no traducida de la proteína de choque térmico 17.3 del trigo
DAS 59122
ZMUbi1 cry34Ab1 pinII
Promotor del gen de la peroxidasa de raíz de Triticum aestivum
cry35Ab1 pinII
p35S CaMV pat t35S
MON 87460 pRACT cspB
Transcrito 7 del gen T-tr7
de A. tumefaciens
p35S CaMV nptII tNOS
MIR604
Promotor derivado del gen de la metalotioneina de Maíz
mcry3A tNOS
ZMUbi1 pmi tNOS
T25
p35S CaMV pat t35S
Promotor bacteriano no
expresado en eucariotas bla Terminador truncado
GA21 pRACT epsps tNOS
C., 2000), y la transformación estable, en algunos casos puede llegar a requerir cultivos celulares o de tejidos y se basa en la integración del ADN foráneo en el material genético de la planta, por lo cual es transmitido en las generaciones sucesivas (Gheysen G., et al.,
1998). Un ejemplo de esa transferencia de ADN es la inserción del precursor de ADN del Ácido Ribonucleico de Interferencia (ARNi) para realizar silenciamiento de genes, utilizando métodos de transformación mediada por Agrobacterium, por Bombardeo de
particulas o por cualquier método que muestre buenos resultados. El resutlado de la transformación es la inducción de la metilación del ADN o la inactivación del ARN mensajero en el citoplasma a nivel traduccional o post-transcripcional respectivamente (COGEM, 2006; Lusser M., et. al., 2011).
2.3.2.1. Transformación mediada por Agrobacterium.
Las bacterias del género Agrobacterium sp. son unas bacterias de la familia Rhizobiaceae
las cuales tienen la habilidad de transferir un segmento de ADN a células de plantas, por medio de un plásmido, generando la introducción e integración de ADN foráneo a las células de la planta obteniendo una planta transgénica (Newell C., 2000; Stewart C., et al.,
2011). En el caso de Agrobacterium tumefaciens, el plásmido contiene información
necesaria para inducir un tumor (factor Ti) y para Agrobacterium rhizogenes genera la
inducción de raíz pilosas (factor Ri) (Stewart C., et al., 2011).
Principalmente, el uso de Agrobacterium tumefaciens a atrapado la atención de la
comunidad científica debido a que el inserto se pasa en una sola pieza y en menos copias que los otros métodos, convirtiéndose en un método de rutina (Gheysen G., et al., 1998).
Su facilidad para insertar ADN en Monocotiledóneas, económicamente importantes como maíz, arroz, trigo y cebada, le ha valido estar involucrado en la gran cantidad de transgénesis relacionadas con estos productos. Por su parte, Agrobaterium rhizogenes es
usado para la transformación de dicotiledóneas por la incorporación de T-ADN del plásmido en el ADN de la planta de una manera análoga al tumor que genera
Agrobacterium tumefaciens. Sin embargo, las plantas transformadas por A. rhizogenes y
su progenie muestran tumoraciones en las raíces lo cual se consideró como una transformación anormales (Newell C., 2000). Adicionalmente, una vez se logró eliminar los genes responsables del genotipo tumoral del plásmido Ti de Agrobacterium tumefaciens, se generalizó el uso de este microorganimos para las transformaciones
genéticas de plantas.
2.3.2.2. Bombardeo de partículas.
El método de bombardeo de partículas se desarrolló como una alternativa a las restricciones de huésped presentadas por A. tumefaciens y los dispendiosos procesos de regeneración de
2011). Este método claramente evita la interacción entre la bacteria y el tejido vegetal lo cual no implica el uso de secuencias específicas para la replicación (Newell C., 2000). Sin embargo, el uso del bombardeo de partículas está limitado por la capacidad de regeneración del tejido bombardeado y la eficiencia en la integración estable del ADN (Newell, 2000). Adicionalmente, debido a la dispersión interna del ADN recombinante en el ADN del hospedero, se presentan múltiple copias de las secuencias objetivo (Wojciech P. y Somers D., 1998).
2.3.2.3. Bigotes de Carburo de Silicio.
Los Bigotes de Carburo de Silicio son Microfibras de 10-80 µm de largo por 0.6 µm de diámetro, las cuales se ponen en solución con el ADN a insertar y los embriones a modificar para realizar la transformación (Petolino J., et al., 2000). Para esta
transformación, es necesario el uso de cultivos embriogénicos en suspensión; por lo cual, las limitaciones de genotipo y los perdida de fertilidad son dos problemas técnicos de los cultivos en suspensión lo cual limita de cierta manera el método (Petolino J., et al., 2000).
2.3.2.4. Transformación de protoplastos.
La transformación de protoplastos es lograr la transformación genética de una célula vegetal suprimiendo de la pared celular utilizando enzimas o por procedimientos mecánicos, para luego por electroporación, microinyección o concentraciones de sales insertar el ADN exógeno (Newell C., 2000, Davey M., et al., 2005 Fernández F., 2006).
Sin embargo, este método de transformación requiere una regeneración de plantas que puede ser compleja y dispendiosa, contrario a la transformación con A. tumefaciens y el
bombardeo de partículas, lo cual rezagó su uso (Newell C., 2000; Davey M., et al., 2005).
2.3.2.5. Electroporación de tejidos.
La electroporación es una técnica que usa pulsos eléctricos para hacer temporalmente permeables las paredes y membranas celulares (Newell C., 2000; Fernández F., 2006). Un impulso eléctrico de alto voltaje, permite a las células tomar el ADN plasmídico lo que genera una expresión estable o transitoria. Este método se ha usado con resultados satisfactorios para la transformación de tejidos regenerables en algunas líneas de maíz (D´Halluin K., et al., 1992; Newell C., 2000) y en tejidos de leguminosas (Chowrira G., et al., 1995; Newell C., 2000). Sin embargo, esta técnica de modificación genética tiene baja
reproducibilidad (Fernández F., 2006).
2.3.2.6. Inyección de ADN.
protoplastos y de embriones derivados de microesporas. Sin embargo, el uso rutinario de esta técnica requiere un esfuerzo técnico muy grande mostrando una eficiencia muy baja (Newell C., 2000).
2.3.2.7. Vectores virales.
Los vectores virales son una alternativa de integración estable de información genética. (Hiatt A., 1993). Los virus pueden tolerar información genética exógena independiente de la fuente de la secuencia de interés, sin perder la capacidad de replicarse (Hiatt A., 1993). La proteína producto puede ser inducida con mayor rapidez y eficiencia (Gleba Y., et al.,
2007).
2.3.2.8. Nuevas técnicas de modificación genética
Las nuevas técnicas de modificación genética responden básicamente a algunas inquietudes de la comunidad científica y de los consumidores finales de OGM, como el control de las mutaciones en función de la especificidad de la mutación en el genoma (no sea al azar), el manejo de genes más compatibles con su hospedero y la expresión de nuevas proteínas, que en algunos casos son sinteticas, pero en el contexto comercial tendría un gran valor.
La técnica de Nucleasa de Dedo de Zinc o ZFN (por sus siglas en inglés; Zinc Finger Nuclease) consiste en una recombinación homóloga de algún fragmento de ADN en un sitio deseado del genoma del hospedero (Lusser M., et. al., 2011; EFSA, 2012). Para lograr
esta recombinación, es necesario insertar en el hospedero por transfección, electroporación o vectores virales, un plásmido de expresión transitoria con la secuencia molde y la secuencia a insertar. La información que contienen el plásmido, expresa para un heterodímero: el dominio Dedo de Zinc, que contiene el ADN recombinante el cual se construye con los tramos flanqueantes del ADN del hospedero para anillarse específicamente, y en el dominio de la Nucleasa, que corta la doble cadena de ADN para realizar la inserción en una secuencia dirigida (Lusser M., et. al., 2011; EFSA, 2012).
Estos dos dominios realizan una modificación que va a ser heredada de forma estable. Principalmente, la ZFN es utilizada para realizar reparaciones, deleciones e inserciones en el material genético del hospedero (EFSA, 2012).
La mutagénesis mediada por oligonucleótidos (Laible G., et al., 2006) está basada en el uso
de cadenas de ADN o ARN de 20 a 100 nucleótidos que no contienen la(s) base(s) que se van a eliminar, los cuales se anillan específicamente al genoma (excepto en la(s) base(s) que se van a eliminar), provocando la formación de horquillas. Una vez formada la horquilla, la Polimerasa replica todo el genoma modificado a partir del oligonucleótido anillado (Laible G., et al., 2006; Lusser M., et. al., 2011).
Inicialmente es necesario realizar la modificación genética del porta injerto, utilizando las técnicas tradicionales de modificación genética como Biolística o Agrobacterium tumefaciens. Una vez se caracterice completamente el porta injerto, el cual debe expresar
de manera estable la característica que se insertó, se procede a realizar el injerto de la especie que se desee modificar (COGEM, 2006; Lusser M., et. al., 2011). Esta técnica se
ha usado para realizar el silenciamiento de genes por medio de RNA de interferencia y la generación de pepinos resistentes a virus del moteado del Mosaico del Pepino (Gal-On A.,
et. al., 2005; COGEM, 2006).
Finalmente, gracias a los últimos desarrollos en las técnicas de Biología Molecular, se han diseñado genomas sintéticos, estos métodos pueden potencialmente utilizarse a futuro en la producción de cultivos genéticemente modificados. Un genoma sintético es el diseño de componentes y sistemas biológicos que no existen en la naturaleza y la reingeniería de los actuales elementos por medio del diseño de sistemas biológicos artificiales (Symbiology, 2005). Recientemente se diseñó el genoma de Mycoplasma genitalium el cual se ensambló
con algunas piezas que se sintetizan comercialmente (Lusser M., et. al., 2011). En un
futuro cercano, se esperan desarrollos en productos y plataformas bioquímicas para la producción de biocombustibles, productos farmacéuticos y modelos de biorremediación eficientes (Lusser M., et. al., 2011).
2.3.3. Regeneración y selección de los tejidos vegetales que han recibido el gen de interés
Dependiendo del tipo de transformación, es necesario regenerar los tejidos modificados. Para este caso, se utilizan técnicas de cultivo in vitro de tejidos, las cuales hacen posible
que a partir de la célula o el tejido modificado, se puedan regenerar plantas completamente a partir de fitohormonas y otros factores añadidos al medio de cultivo (Muñoz J., 2004; Fernández F., 2006; Mertxe S., 2010) gracias a la totipotencia de las células vegetales (Arunraj D. y Babu G., 2002).
Para diferenciar entre las células o plantas modificadas de las no modificadas, se utilizan medios de cultivo celular con antibióticos o sustratos, para los cuales, las plantas modificadas tienen resistencia o tolerancia (Gheysen G., et al., 1998; Newell C., 2000;
Fernández F., 2006) o marcadores de selección con agentes fluorescentes (Sheen J., et al.,
2002). Esta resistencia o tolerancia es un acompañante cotidiano de los insertos de interés. Los marcadores de selección de antibióticos más ampliamente utilizados para la selección de células o planta modificadas son el gen de la neomicina fosfotransferasa II (nptII), la
higromicina fosfotransferasa (hpt) y el Fosfinotricina acetil transferasa (bar/pat) (Gheysen
G., et al., 1998; Newell C., 2000). Los genes reporteros que tienen actividad sobre
sustratos que no están de manera endógena en el hospedero. Los más utilizados es el gen
manA el cual codifica para la proteína fosfomanosa isomerasa, el gen sylA que codifica para
la Xilosa isomerasa y el gen uidA que codifica para la -glucoronidasa. Estas proteínas
Gheysen G., et al., 1998; Newell C., 2000; Fernández F., 2006). En algunos casos, cuando
el gen de interés insertado es la tolerancia a algún herbicida, la selección de las células o plantas modificadas se basa en una prueba de tolerancia al herbicida insertado.
2.4. Caracterización del nuevo organismo vivo.
Los nuevos organismos genéticamente modificados, son analizados detalladamente para determinar sus características agronómicas y de seguridad para los consumidores y el medio ambiente (Windels P. et al., 2001; Mertxe S., 2010). Pese a que el proceso de
inserción puede ser estandarizado y desarrollado por personal altamente capacitado, controlar todos los aspectos relacionados con la transformación es una tarea imposible. Por esta razón, de acuerdo a los lineamientos del Codex Alimentarius (2009), en los Principios para el análisis de riesgos de alimentos obtenidos por medios biotecnológicos modernos (CAC/Gl 33-2003), la empresa productora de OGM, “deberá proporcionar suficiente información sobre la modificación genética a fin de que sea posible identificar todo el material genético que puede haberse aportado a la planta huésped, y suministrar la información necesaria para el análisis de los datos que apoyan la caracterización del ADN insertado en la planta”. Particularmente esta información debe incluir la descripción del
proceso de transformación, información sobre el ADN que ha de introducirse, caracterización molecular y bioquímica completa de la modificación genética, información sobre la inserción del ADN en el genoma de la planta, información de todas las sustancias que se hayan expresado en la planta de ADN recombinante y alguna información adicional (estabilidad del ADN del huésped, presencia de modificaciones postraduccionales no objetivo, funcionalidad del objeto de la modificación genética, afectación por la inserción del material genético del huésped e identidad y modalidades de expresión de la nueva proteína). Finalmente se debe realizar la evaluación de la toxicidad, alergenicidad, alteración en la tasa de expresión de metabolitos secundarios en la planta, efecto de la elaboración de alimentos en la proteína insertada y las modificaciones nutricionales derivadas de la inserción genética (OMS-FAO, 2009).
2.5. Clasificación de los OGM
Existen cuatro clasificaciones de los OGM:
1) Basada en el origen de los elementos insertados. 2) Basada en la característica insertada.
2.5.1. Basada en el origen de los elementos insertados
Esta clasificación se divide en cuatro generaciones. La primera generación denominada
Eventos de transformación individual, comprende los eventos de transformación genética en los cuales la secuencia de interés era aislada con enzimas de restricción e insertada en un plásmido bacteriano de alta propagación construido generalmente con los promotores p35S CaMV o pNOS, los terminadores t35S CaMV o tNOS y en algunos casos intrones que contribuyen a la regulación transcripccional y post-transcripccional de la señal (Arunraj D. y Babu B., 2002; Holst-Jensen A. et. al., 2012). Adicionalmente, esos OGM contienen
genes marcadores de selección los cuales se usaron inicialmente para evidenciar la transformación exitosa de la planta modificada.
La segunda generación comprende los Eventos apilados o Stacks. Los Stacks se dividen en
tres grandes grupos: Stacks por transformación, Stacks moleculares y Stacks por cultivo.
Los Stacks por transformación es la inserción de un nuevo gen a un evento de
transformación genética ya existente. Los Stacks moleculares son la modificación de un
parental no modificado genéticamente con un vector que contiene dos o más genes de interés para la transformación. Finalmente, los Stacks por cultivo es el cruzamiento de dos
eventos de transformación genética para generar una nueva variedad con las dos caracteristicas de sus parentales (Rashmi S., 2012).
La tercera generación es denominada cerca de los intragénicos. Estos eventos de transformación genética cuentan con una secuencia diana generalmente tomada de la misma especie a modificar, pero con constructos externos (plásmidos convencionales). Un ejemplo de estos OGM es la papa AV43-6-G7, la cual codifica una secuencia antisentido de la enzima sintasa del gránulo del almidón GBSS (por sus siglas en ingles Granule Bound Starch Synthase) y fue transformada con el plásmido pKGBA50mf-IR1.1 que contienen
fragmentos de Agrobacteriun tumefaciens, Escherichia coli y fragmentos modificados del
fago M13 ( http://www.gmo-compass.org/eng/gmo/db/150.docu.html ).
Finalmente, los OGM de cuarta generación son conocidos como intragénicos y cisgénicos (Holst-Jensen A. et. al., 2012). Estos OGM son aquellos en los cuales el origen de los
insertos son asociados a especies que son compatibles y de la misma especie respectivamente (Lusser M., et. al., 2011; Holst-Jensen A. et. al., 2012).
2.5.2. Basada en la característica insertada
La clasificación de los OGM de acuerdo a la característica insertada se divide en dos grandes grupos. El primero son los cultivos con características agronómicas mejoradas o primera generación y los organismos con mejoras en las características nutricionales o
2.5.2.1. Organismos con características agronómicas mejoradas.
Este tipo de organismos son los más ampliamente desarrollados y son los que al mejorar las características agronómicas se reduce la pérdida por ataque de algunas especies de insectos y virus, o para incentivar el uso de herbicidas menos fuertes y más amigables con el planeta, o por retrasar la degradación del producto de interés comercial, o para tener la posibilidad de soportar estrés abiótico, lo cual representa una mejora en el rendimiento del cultivo (Jiang Y. y Fu J., 2000; Robinson C., 2001; Mertxe S., 2010; Kolodyazhnaya Y.,
et al., 2009). Esto se consigue por la inserción y expresión de genes que generen
resistencia a virus, hongos, bacterias, insectos o mediante la tolerancia a un herbicida, o por la inserción de un gen anti sentido que silencie la producción de etileno o pectina (Jiang Y. y Fu J., 2000; Robinson C., 2001).
Los OGM con resistencia a insectos, principalmente expresan delta endotóxinas o comúnmente llamadas proteínas Cry, que son derivadas de Bacillus thuringiensis (Romeis
J., et al., 2008; Sum S., 2008; Mertxe S., 2010), las cuales luego de ser activadas por
proteasas y el pH alcalino del intestino del insecto, se une a receptores específicos de membrana en las células epiteliales del intestino medio formando un poro, lo que genera la muerte por septicemia al insecto (Schnepf E., et al., 1998). Históricamente, el manejo
integrado de plagas busca mejorar el control biológico mediante la conservación de los enemigos naturales existentes, o estableciendo otros nuevos a través de la inoculación o inundación (Romeis J., et al., 2008); sin embargo, el uso de las proteínas cry se popularizó
debido a su efectivo modo de acción, a un historial de uso seguro y a la practicidad agronómica que ofrece su manejo dentro del OGM. Otro mecanismo insecticida que se emplea para OGM, es la expresión de lecitinas o inhibidores de la proteasa y amilasas lo cual favorece la defensa química propia de la planta (Robinson C., 2001; Romeis J., et al.,
2008). Los eventos de transformación genética con resistencia a insectos aprobados para consumo humano en Colombia, se encuentran en la Tabla 4.
Los OGM resistentes a virus y enfermedades, se desarrollaron por la dificultad para controlar el efecto letal de estas partículas (Sum S., 2008; Mertxe S., 2010). Esta transformación consiste en la inserción de genes que expresan proteínas como la de la cápside del virus en el genoma de la planta, replicasas y ARN interferentes (Bolivar F., 2007; Sum S., 2008). Históricamente, se desarrolló sandía resistente al virus del mosaico de la sandía (WMV), calabacín resistente al virus del mosaico del calabacín (ZYMV) por la introducción y expresión de genes de cubierta de los virus WMV y ZYMV. Sin embargo, el OGM más emblemático resistente a virus es la papaya resistente al virus de la mancha anular (PRSV) por la expresión de la proteína VMAP de la capa del virus, lo cual representó la recuperación de la producción de papaya en Hawái, la cual había sido arrasada en un 80% por el virus PRSV (Ferreira S. y Pits K., 2002; Sum S., 2008).
Tabla4. Eventos de transformación genética aprobados para consumo humano en Colombia
Nombre del Evento Característica insertada Matriz Día de aprobación
MON531 Resistencia a lepidópteros Algodón 24/06/2003 MON88913 Resistencia a lepidópteros Algodón 27/10/2003 MON810 Resistencia al barrenador europeo del maíz Maíz 27/10/2003 MON71800 Tolerancia al herbicida glifosato Trigo 01/04/2004 NK603 Tolerancia al herbicida glifosato Maíz 01/04/2004 GTS 40-3-2 Tolerancia al herbicida glifosato Soya 30/12/2005
H7-1 Tolerancia al herbicida glifosato Remolacha
Azucarera 30/12/2005
TC1507 Resistencia al barrenador europeo del maíz, tolerancia al
herbicida glufosinato de amonio Maíz 17/10/2006 MON531 x MON1445 Resistencia a lepidópteros y tolerancia al herbicida glifosato Algodón 16/06/2008 LLRICE601 Tolerancia al herbicida glifosinato de amonio Arroz 26/09/2008 LLRICE62 Tolerancia al herbicida glifosinato de amonio Arroz 26/12/2008
BT 11 Resistencia al barrenador europeo del maíz, tolerancia al
herbicida glufosinato de amonio Maíz 13/04/2009 MON 810 x NK 603 Resistencia a lepidópteros y tolerancia al herbicida glifosato Maíz 26/11/2009 MON 15985 Resistencia a lepidópteros Algodón 01/12/2009
TC1507 x NK603 Resistencia a lepidópteros y tolerancia a los herbicidas
glufosinato de amonio y glifosato Maíz 09/02/2010
MON 89788 Tolerancia al herbicida glifosato Soya 24/06/2010
MON 88913 Tolerancia al herbicida glifosato Algodón 24/06/2010 MON 15985 x MON 88913 Resistencia a lepidópteros y tolerancia al herbicida glifosato Algodón 24/06/2010 MON 89034 Resistencia a lepidópteros Maíz 24/06/2010
DP 356043 Tolerancia a los herbicidas glifosato y los derivados de la imidazolinona. Soya 24/06/2010
MON 89034 x NK603 Resistencia a lepidópteros y tolerancia al herbicida glifosato Maíz 24/06/2010
MON 89034 x TC 1507 x MON 88017 x DAS 59122
Resistencia a coleópteros, lepidópteros, el barrenador europeo del maíz y tolerancia a los herbicidas glifosato y
glufosinato de amonio
Maíz 24/06/2010
LY 038 Mayor contenido de lisina Maíz 19/07/2010
MON 863 Resistencia al gusano de la raíz del maíz Maíz 18/05/2011
MON 88017 Resistencia al gusano de la raíz del maíz y tolerancia al
herbicida glifosato Maíz 18/05/2011
DAS 59122 Resistencia al gusano de la raíz del maíz y tolerancia al
herbicida glulfosinato de amonio Maíz 18/05/2011
MON 89034 x MON 88017 Resistencia a coleópteros y lepidópteros, y tolerancia al
glifosato Maíz 18/05/2011
MON 87460 Uso eficiente de agua Maíz 18/05/2011
MON 810 x MON 88017 Resistencia a coleópteros y lepidópteros, y tolerancia al
glifosato Maíz 24/05/2011
T25 x NK603 Tolerancia a los herbicidas glufosinato de amonio y glifosato Maíz 26/01/2012 MON 87701 x MON 89788 Resistencia a lepidópteros y tolerancia al glifosato Soya 26/01/2012 CV127 Tolerancia a los herbicidas derivados de la imidazolinona Soya 26/01/2012 MIR604 Resistencia al gusano de la raíz del maíz Maíz 26/01/2012
BT11 x MIR162 x MIR604 x GA21
Resistencia a coleópteros, lepidópteros, al barrenador europeo, y tolerancia a los herbicidas glifosato y glufosinato
de amonio
Maíz 26/01/2012
Bt11 x MIR604 Resistencia a coleópteros y lepidópteros y tolerancia al
herbicida glufosinato de amonio Maíz 26/01/2012
T25 Resistencia a le lepidópteros y tolerancia al glufosinato de
amonio Maíz 26/01/2012
caso de Glifosato que es el principio activo del herbicida Roundup® de Monsanto Company, interfieren alguna vía metabólica importante para el desarrollo de la planta, generando que la planta no se desarrolle con normalidad y muera (Mertxe S., 2010). Para lograr la tolerancia a estos herbicidas, se han insertado genes que expresan proteínas como la CP4EPSPS de Agrobacterium cepa CP4, la cual es una
5-enolpiruvil-shiquimato-3-fosfato sintetasa resistente al efecto del glifosato, el cual, inhibe la enzima EPSPS nativa de las plantas generando que no se sintetice aminoácidos aromáticos como la fenilalanina, tirosina y triptófano (Arunraj D. y Babu G.; 2002; Sum S., 2008). Otro gen que se inserta para generar la tolerancia a un herbicida específico, es el gen que codifica para la proteína PAT de Streptomyces viridochromogenes el cual genera la resistencia al glufosinato de
amonio (Freyssinet G., 2003). El glufosinato de amonio es convertido por las plantas en L-fosfinocitrina, el cual es un inhibidor fuerte de la Glutamina Sintetasa, siendo esta precursora de la asimilación de amonio (Torres J., et al., 2009). La proteína PAT, que es
una fosfinotricin-acetil transferasa, acetila la molécula de L-fosfinotricina en N-acetil-L-glufosinato la cual no es tóxica para la planta (Sum S., 2008; Torres J., et al., 2009). Los
eventos de transformación genética con tolerancia a herbicidas aprobados para consumo humano en Colombia, se encuentran en la Tabla 1.
Los OGM con un retraso en su degradación, son OGM en los cuales se les ha silenciado el gen de la expresión del Etileno o Pectina. Para llegar a este desarrollo tecnológico, fue necesario entender profundamente el metabolismo y la expresión de los genes específicos involucrados en la maduración del fruto (Bleecker A. y Schaller G., 1996; Sisler E. y Serek M., 2007; Jiang Y. y Fu J., 2000). Una vez se comprendió el metabolismo y se determinaron los genes involucrados, se insertó un gen antisentido en la planta para silenciar la expresión del gen (Jiang Y. y Fu J., 2000; Sum S., 2008) lo cual generó un retrazo en la maduración del fruto aumentando directamente la vida útil del alimento.
Finalmente, el estrés abiótico es conocido como el efecto del frio, la disponibilidad de agua, la salinidad del suelo, las temperaturas críticas, la concentración tóxica a metales pesados, acidez y alcalinidad, déficit de nutrientes inorgánicos en los cultivos, los cuales afectan directamente la productividad (Xu D., et al., 1996a; Kolodyazhnaya Y., et al., 2009; Allen
R., 2010). Para generar plantas con tolerancia al estrés abiótico, fue necesario identificar y controlar los genes que son inducidos en estrés abiótico. Estos genes incluyen los relacionados con la degradación de osmolitos compatibles como aminoácidos (prolina y alanina), iones cuaternarios (betaína, glicina betaína), azúcares y alcoholes de azúcares (manitol, sorbitol, trealosa e inositol) y carbohidratos (Delauney J. y Verma D., 1993; Queitsch C., et al., 2000; Zhou H., et al., 2004; Allen R., 2010; Sundaram S. y
Rathinasabapathi B, 2010). Al controlar la expresión de estos genes, se disminuye el potencial hídrico de las células, se protegen las enzimas de la inactivación y se mantiene la integridad estructural de la proteína (Kolodyazhnaya Y., et al., 2009). La estrategia para