ANÁLISIS DE LA DINÁMICA HÍDRICA DE PLANTAS TIPO C
3Caso de estudio: Cultivo de Maracuyá (Passiflora edulis variedad
flavicarpa) en condiciones de clima templado.
JONATHAN ROMERO CUELLAR (I.A)
Trabajo de grado para optar al título de:
Magíster en Hidrosistemas
PONTIFICIA UNIVERSIDAD JAVERIANA
FACULTAD DE INGENIERÍA
DEPARTAMENTO DE INGENIERÍA CIVIL
MAESTRÍA EN HIDROSISTEMAS
JONATHAN ROMERO CUELLAR Página 2
ANÁLISIS DE LA DINÁMICA HÍDRICA DE PLANTAS TIPO C
3Caso de estudio: Cultivo de Maracuyá (Passiflora edulis variedad
flavicarpa) en condiciones de clima templado.
JONATHAN ROMERO CUELLAR (I. A)
Director:
NELSON OBREGÓN NEIRA
(I.C, M.Sc, Ph.D)
PONTIFICIA UNIVERSIDAD JAVERIANA
SEDE BOGOTÁ
FACULTAD DE INGENIERÍA
DEPARTAMENTO DE INGENIERÍA CIVIL
MAESTRÍA EN HIDROSISTEMAS
JONATHAN ROMERO CUELLAR Página 3
REGLAMENTO DE LA
PONTIFICIA UNIVERSIDAD
JAVERIANA.
Artículo 23. "La Universidad no se hace
responsable por los conceptos emitidos
por sus alumnos en sus trabajos de tesis.
Sólo velará porque no se publique nada
contrario al dogma y a la moral católica
y porque las tesis no contengan ataques
personales contra persona alguna, antes
bien se vea en ellas el anhelo de buscar
la verdad y la justicia".
JONATHAN ROMERO CUELLAR Página 4 El proyecto de grado titulado:
ANÁLISIS DE LA DINÁMICA HÍDRICA DE
PLANTAS TIPO C3. Caso de estudio: Cultivo de
Maracuyá (Passiflora edulis variedad flavicarpa)
en condiciones de clima templado.” presentado
por Jonathan Romero Cuellar, en cumplimiento parcial de los requisitos exigidos para optar el título de Magíster en Hidrosistemas, fue aprobado el día 05 del mes de Junio de 2012.
_________________________
PhD. MSc. Ing. Nelson Obregón Neira Director
_________________________
MSc. Ing. Francisco Boshell Villamarín Jurado
_________________________ MSc. Ing. Carlos Devia Castillo Jurado
JONATHAN ROMERO CUELLAR Página 6
Agradecimientos
Especial agradecimiento al grupo de investigación GHIDA (Hidroingeniería y Desarrollo Agropecuario), a la Universidad Surcolombiana, a Colciencias, al CEPASS, la Gobernación del Huila.
Igualmente a la PUJ, los profesores (Andrés Torres, Jaime Lara, Efraín Dominguez, Ramon, etc.) y amigos de la Maestría en Hidrosistemas, a los amigos del Instituto Geofísico (Yolanda, Iliana, Felipe, Jorge Luis, Francisco, Eder, Hugo, Santiago, Don Fernando, Ana María, Javier, Thomas, Mario, Jaime, Edgar, Alfonso, Pedro).
También a CENIGAA por su apoyo y amistad (Jorge, Oscar, Gustavo, Helmer, Amalia).
A Caro (yogo) por su apoyo, comprensión, amor y cariño.
Finalmente y no menos importante al gran maestro, al profe, a Nelson, por todo su apoyo y conocimiento transmitido, por enseñarme lecciones para la vida, sin el este sueño no hubiese sido posible. Un gran maestro dedica su vida a estructurar otras vidas, por lo tanto lo que enseña no tiene precio.
En la cadena de las especies el árbol es un logro en sí. Es una escultura viviente y perfecta.
Es un desafío a la gravedad. El árbol es el único elemento natural en movimiento perpetuo hacia el cielo. Crece sin prisa hacia la luz de la cual se alimenta su follaje.
JONATHAN ROMERO CUELLAR Página 7
Contenido
Listado de Figuras ... 9
Listado de Tablas ... 10
1. Introducción ... 11
2. Objetivos ... 12
2.1 Objetivo General ... 12
2.2 Objetivos Específicos ... 13
3. Marco Conceptual ... 13
3.1 Potencial hídrico en el sistema suelo-planta-atmósfera. ... 13
3.2 Transpiración de la planta ... 15
3.3 Eficiencia fotosintética en el uso del agua ... 16
3.4 Fotosíntesis ... 17
3.4.1 Ruta Fotosintética C3 ... 18
3.4.2 Ruta Fotosintética C4 ... 21
3.4.3 Ruta Fotosintética CAM ... 22
3.4.4 Fotosíntesis en el contexto del cambio climático ... 22
3.5 Interacciones de la capa límite en la hoja ... 23
3.6 Descripción del modelo implementado ... 25
3.6.1 Fotosíntesis C3 ... 26
3.6.2 Conductancia Estomática ... 30
3.6.3 Condiciones intercelular y superficial de la hoja ... 31
3.6.4 Conductancia de la capa límite en la hoja ... 32
3.6.5 Balance de energía en la hoja ... 32
3.6.6 Acople de procesos en la hoja ... 33
3.7 Cultivo de Maracuyá ... 34
3.7.1 Descripción botánica ... 34
3.7.2 Fotosíntesis y Transpiración del cultivo Maracuyá... 34
3.7.3 Requerimientos climáticos y edáficos ... 35
4. Metodología, diseño experimental y descripción del caso de estudio ... 35
4.1 Ubicación zona experimental ... 36
JONATHAN ROMERO CUELLAR Página 8
4.3 Protocolo de modelación implementado ... 37
4.4 Caso de estudio ... 39
4.4.1 Información meteorológica ... 39
5. Herramienta computacional ... 44
5.1 Diseño algorítmico del modelo implementado ... 45
5.2 Descripción del programa ... 47
5.3 Verificación del código de programación ... 47
6. Análisis de Resultados ... 48
6.1 Matriz de Adyacencia ... 48
6.2 Aplicación del Modelo para el cultivo de maracuyá en el departamento del Huila. ... 49
6.2.1 Calibración del modelo implementado ... 49
6.2.2 Validación del modelo implementado ... 52
6.2.3 Simulación del modelo implementado ... 53
6.3 Simulación de escenarios ... 54
7. Propuesta de protocolo de medición ... 62
7.1 Análisis de las variables del modelo implementado con su posibilidad de medición ... 62
7.2 Técnica Eddy covariance y equipos de medición ... 65
8. Conclusiones ... 67
9. Recomendaciones y trabajos futuros ... 68
Referencias bibliográficas ... 69
Definiciones ... 72
Anexos ... 73
Anexo 1. Código de la herramienta computacional implementada. ... 73
Anexo 2 Características del Lisímetro ... 83
Anexo 3 Detalles del caso de estudio ... 84
Anexo 4 Aporte de la herramienta diseña ... 86
JONATHAN ROMERO CUELLAR Página 9
Listado de Figuras
Figura 1 Valores generales del potencial hídrico en el sistema
suelo-planta-atmósfera. ... 14
Figura 2 Elementos que interactúan en la fotosíntesis (Bieto y Talón, 2008)... 18
Figura 3 Representación de la ruta fotosintética C3 (Smith y Smith, 2007). ... 20
Figura 4 Representación de la ruta fotosintética C4 (Smith y Smith, 2007). ... 21
Figura 5 Diagrama causal de las interacciones entre el ambiente y la planta. Adaptado de (Golberg, 2010). ... 24
Figura 6 Esquema de la anatomía de la hoja de una planta C3 indicando las resistencias al flujo de entrada de C02 y de salida de vapor de agua a través del estoma (Gracia, 2011). ... 30
Figura 7 Localización del proyecto. ... 36
Figura 8 Representación de la serie de Temperatura ambiente registrada por la estación automatizada de la zona experimental registros horarios a lo largo del día. ... 40
Figura 9 Representación de la serie de humedad relativa registrada por la estación automatizada de la zona experimental registros horarios a lo largo del día. ... 41
Figura 10 Representación de la serie de velocidad del viento registrada por la estación automatizada de la zona experimental registros horarios a lo largo del día. ... 42
Figura 11 Representación de la serie de radiación neta registrada por la estación automatizada de la zona experimental registros horarios a lo largo del día. ... 43
Figura 12 Apariencia de las señales registradas por la estación automatizada de la zona experimental cada hora. ... 44
Figura 13 Diagrama de flujo de la solución del modelo. ... 46
Figura 14 Matriz de Adyacencia Modelo Acople Biofísico y Bioquímico a nivel de hoja. ... 48
Figura 15 Puntos obtenidos en la exploración del espacio paramétrico del modelo... 50
Figura 16 Análisis de sensibilidad regional del modelo. ... 51
Figura 17 Simulación y medición de la transpiración diaria del cultivo de maracuyá, Huila, Colombia. ... 52
JONATHAN ROMERO CUELLAR Página 10
Figura 19 Simulación de An, Gs, ET y UEA a nivel diario para el cultivo de maracuyá en el Huila... 54
Figura 20 Respuesta de la conductancia estomática (Gs) respecto a la variación de la velocidad del viento para el cultivo de maracuyá (Ta=23°C y HR=60%). ... 55
Figura 21 Simulación del efecto de PPFD sobre la temperatura y la respuesta de la tasa de fotosíntesis (An) para el cultivo de maracuyá (HR=60%). ... 56
Figura 22 Respuesta de la Gs frente a cambios de Ta bajo tres escenarios de PPFD para el cultivo de maracuyá. ... 57
Figura 23 Simulación de la respuesta de la (An) frente a cambios en el CO2
atmosférico para el cultivo de maracuyá. ... 57
Figura 24 Simulación de la respuesta de (Gs) frente a cambios en el CO2
atmosférico para el cultivo de maracuyá. ... 58
Figura 25 Simulación de la tasa de fotosíntesis frente a tres escenarios de CO2 en la
atmosfera para el cultivo de maracuyá. ... 59
Figura 26 Simulación de la respuesta del uso eficiente del agua (EFA) respecto a la variación de PPFD y la velocidad del viento para el cultivo de maracuyá. ... 60
Figura 27 Simulación de la respuesta del uso eficiente del agua (EFA) respecto a la variación de la temperatura ambiente y la velocidad del viento para el cultivo de maracuyá. ... 61
Figura 28 Matriz de adyacencia del sistema si se mide la temperatura de la hoja. 63
Listado de Tablas
Tabla 1 Ubicación de la zona experimental... 37
Tabla 2 Combinación de los 10 mejores parámetros del modelo. ... 51
Tabla 3 Identificación de parámetros del modelo para el cultivo de maracuyá en el departamento del Huila. ... 53
JONATHAN ROMERO CUELLAR Página 11
1.
Introducción
El invento de la agricultura cambio radicalmente la historia. La agricultura fue la primera gran revolución, con ella se establecieron las ciudades. Como todas las especies la principal preocupación es el alimento diario por ende la supervivencia de la humanidad depende directamente de la agricultura (Bertrand, 2009).
No obstante, en la gran mayoría de zonas agrícolas y principalmente en regiones áridas y semiáridas el manejo eficiente del agua de riego es un problema crítico (Hanks y Hill, 1980). Las fuentes del recurso hídrico se agotan y esto genera un aumento en los costos de producción de los alimentos (Hanks y Hill, 1980).
Por esta razón, es necesario realizar un manejo eficiente del agua de riego. Una de las principales tareas a realizar es estimar adecuadamente los requerimientos hídricos de los cultivos (Villalobos et al., 2000). De hecho los principales componentes de los requerimientos hídricos son la evaporación y la transpiración para los cuales existen diferentes modelos.
Sin embargo los modelos no predicen lo que va suceder, pero si permiten entender mejor a partir de hoy lo que puede suceder mañana. Uno de los aspectos importantes de los modelos que se presentan en este trabajo de grado es la estimación de la traspiración de los cultivos, esta aplicación es una herramienta potente para una programación óptima del agua de riego (Babajimopoulos C. et al., 1995).
En este sentido, el departamento del Huila no es ajeno a las problemáticas planteadas anteriormente, principalmente porque es un departamento productor de alimentos. Una de las principales apuestas productivas es el cultivo de Maracuyá. En el año 2009 el cultivo de maracuyá reportó una de los mayores aportes en el producto interno bruto (1.36%) del departamento del Huila (Torrente, 2011).
JONATHAN ROMERO CUELLAR Página 12
cultivos (transpiración, fotosíntesis y conductancia estomática) y las condiciones hidrometeorológicas (De Oliveira et al., 2005). Así mismo, el manejo adecuado del agua de riego mejora la calidad de los cultivos y esto incrementa la competitividad de los productores (Acevedo-Opazo et al., 2010).
Por lo tanto, el documento presentado cuenta con 7 capítulos principales los cuales son: el capítulo 3 donde se condensa todo el marco conceptual y teórico del trabajo de grado, el capítulo 4 es la contextualización en el caso de estudio, el capítulo 5 es el desarrollo computacional implementado, el capítulo 6 son los resultados (las simulaciones), el capítulo 7 es la propuesta de protocolo de medición y finalmente conclusiones y recomendaciones.
2.
Objetivos
JONATHAN ROMERO CUELLAR Página 13
Analizar las interacciones ecofisiológicas (fotosíntesis, conductancia
estomática) de las plantas tipo C3 con variables ambientales (velocidad
del viento, temperatura del ambiente, humedad relativa) con miras al uso eficiente del agua en el contexto del caso de estudio.
2.2Objetivos Específicos
Desarrollar herramientas que faciliten el análisis y la simulación de
interacciones clima-suelo-biota en el marco del uso eficiente del agua de riego agrícola.
Proponer un protocolo de medición de variables
hidrometeorológicas y ecofisiológicas que faciliten la
implementación de modelos que representen la interacción clima-suelo-biota.
3.
Marco Conceptual
JONATHAN ROMERO CUELLAR Página 14
El movimiento del agua en el sistema suelo–planta–atmósfera obedece a
los planteamientos de la segunda ley de la termodinámica. Aplicando los principios de esta ley al contexto del potencial hídrico el desplazamiento del agua siempre será de una zona de mayor potencial a una de menor potencial, provocando una disminución de energía potencial (Larqué y Trejo, 1990).
Es decir en el interior de la planta el potencial hídrico es más elevado en las raíces, disminuyendo progresivamente en el tallo, presentando los valores más bajos en las hojas (Bieto y Talón, 2008). Teniendo en cuenta el ambiente el potencial más negativo se encuentra en la atmósfera. En
síntesis la base del continuo suelo–planta–atmósfera consiste en un
[image:14.612.81.529.290.539.2]gradiente negativo, presentando el potencial más negativo en las hojas y menos negativo en las raíces (Figuras 1).
Figura 1 Valores generales del potencial hídrico en el sistema suelo-planta-atmósfera. Tomado de (Bieto y Talón, 2008).
Adicionalmente, Bieto y Talón (2008) indican que el movimiento del agua se manifiesta por dos fenómenos conocidos como: flujo másico y difusión. El flujo másico se presenta debido a una diferencia de presión que promueve el movimiento conjunto de moléculas de agua y solutos en una dirección. Por el contrario, en el transporte difusivo es en todas las direcciones y rige el movimiento de agua entre las células vivas.
JONATHAN ROMERO CUELLAR Página 15
que el movimiento en la fase del vapor de agua es proporcional al gradiente de concentración del vapor de agua (García y Fuentes, 1994).
3.2 Transpiración de la planta
Estudiar el proceso de la transpiración puede tener varias justificaciones, pero desde un punto de vista muy aplicado se puede decir que la principal pérdida económica y el fracaso de los cultivos del mundo se deben a la presencia de déficits hídricos producidos por el exceso de la transpiración (Bieto y Talón, 2008).
En este sentido la transpiración es principalmente un proceso de evaporación desde la carpeta vegetal y más específicamente desde los estomas. También es un proceso controlado por factores físicos y fisiológicos. En síntesis la transpiración es el proceso dominante en las relaciones hídricas de una planta, y altas tasas de transpiración conlleva a
una intensa absorción de CO2 (De Oliveira et al., 2005).
Tradicionalmente se presenta la transpiración como un proceso de perdida de agua, pero es una visión un poco simple. Incluso la transpiración es un mecanismo eficiente de transporte de nutrientes a las diferentes partes de las plantas y también cumplen un papel vital en la regulación térmica pues la energía requerida para el cambio de fase del agua de su estado líquido a gaseoso genera una sensación refrigerante para las plantas (Bieto y Talón, 2008).
De acuerdo con Larqué y Trejo (1990) la tasa de transpiración depende del suministro de energía para evaporar el agua, el gradiente de presión de vapor y principalmente de la magnitud de la resistencia a lo largo del tejido vascular de la planta.
De hecho la diferencia de presión de vapor de agua entre la hoja y el aire es la fuerza conductora del proceso de transpiración. La presión de vapor del aire se relaciona con la humedad absoluta y la temperatura, por otro lado, la presión de vapor de la hoja se asocia con la temperatura y el potencial hídrico(Campbell y Norman, 1997; Bietoy Talón, 2008).
Por último, en el campo de disciplinas como la hidrología y las ciencias agrícolas se planteó unir el concepto de evaporación del suelo y la
transpiración de las plantas en un solo término llamado
JONATHAN ROMERO CUELLAR Página 16
3.3 Eficiencia fotosintética en el uso del agua
En la actualidad hay una tendencia mundial de convertir la agricultura en una actividad sustentable. Básicamente porque esta, es una necesidad primaria de los seres vivos, y también por la influencia de factores como: el incremento descontrolado de la población humana y los efectos del cambio climático.
Debido a que, con frecuencia, el agua es un recurso limitante de la producción vegetal, existe un interés considerable en determinar la eficiencia en el uso del agua en relación con la fotosíntesis y la productividad (Bieto y Talón, 2008).
La eficiencia en el uso del agua es un parámetro que se puede considerar a diferentes niveles, para este trabajo se relaciona con la hoja, suele tener diferentes nombres: eficiencia en el uso del agua de la fotosíntesis, ó eficiencia intrínseca del uso del agua, eficiencia fotosintética en el uso del agua.
Pero en general para este trabajo se identifica este parámetro como
“Eficiencia fotosintética en el uso del agua (UEA)” y conceptualmente se
conoce como la cantidad neta de CO2 absorbido por unidad de agua
transpirada por una hoja, este parámetro combina el efecto de la bioquímica de la hoja con la respuesta estomática (Nikolov et al., 1995).
En términos matemáticos ⁄ , en otras palabras el UEA es una
medida de la efectividad de los estomas en maximizar la fotosíntesis reduciendo, al mismo tiempo, la perdida de agua (Bieto y Talón, 2008).
Por otro lado, a nivel de hoja los factores ambientales y de la planta
influyen en el UEA. Además algunos estudios demuestran que el UEA
disminuye exponencialmente con el incremento del déficit de presión del vapor de agua (disminución humedad relativa) porque se eleva la transpiración sin aumento simultáneo de fotosíntesis. Por tanto, un aumento en la temperatura del aire, a través de su efecto en el déficit de presión de
vapor reducirá el UEA (Baldocchi et al., 1987; Nikolov et al., 1995; Bieto and
JONATHAN ROMERO CUELLAR Página 17
Un manejo eficiente del agua de riego requiere información del consumo de agua por parte de los cultivos (Hanks y Hill, 1980). Además Bieto y Talón (2008) indican que el uso eficiente del agua de riego requiere por definición una buena cuantificación de la evaporación y la transpiración.
Finalmente, el UEA es una característica funcional de la vegetación. A
escala de individuo esta limita el potencial de crecimiento de una especie en condiciones de escasez hídrica. A nivel de paisaje determina la relación entre la hidrología de la cuenca y la producción primaria del ecosistema (Baldocchi y Meyers, 1998).
3.4 Fotosíntesis
Todos los seres vivos están compuestos por moléculas construidas sobre una estructura de átomos de carbono. Por lo tanto, toda la vida sobre la tierra se basa en el carbono (Smith y Smith, 2007).
Luego, escribir sobre el carbono es necesariamente escribir sobre plantas y sin lugar a dudas es conveniente pensar el proceso que acopla las plantas con el carbono, en este caso la fotosíntesis. La fotosíntesis tiene muchas definiciones e hipótesis, la intención de estos párrafos es aproximarse de manera sencilla hacia algunas de ellas.
La fotosíntesis es el proceso por el cual se transforma el CO2 en moléculas
orgánicas y tejidos vivos. En otras palabras es la conversión de dióxido de carbono en monosacáridos. También se conoce como un proceso que aprovecha la energía de onda corta (PAR) para conducir una serie de
reacciones químicas que resulta en la fijación de CO2 en los carbohidratos
(monosacáridos) y en liberación de O2 como subproducto (Smith y Smith,
2007).
Además y de acuerdo con Bieto y Talón (2008) la fotosíntesis es un proceso biológico complejo, que implica dos fases, la fase de foto-absorción de energía y la de foto-asimilación de elementos esenciales de la materia orgánica. En la primera fase la energía luminosa es absorbida por biomoléculas fotosensibles y transformadas en una forma de energía bioquímicamente estable. En la segunda fase los elementos constitutivos
son tomados de fuentes minerales inorgánicas (agua H2O, dióxido de
carbono CO2, nitratos NO3, sulfatos SO4, etc.) e incorporados en
biomoléculas orgánicas metabolizables.
JONATHAN ROMERO CUELLAR Página 18 →
Agua + Dióxido de Oxígeno + Glucosa Carbono
[image:18.612.86.532.247.377.2]La ecuación anterior indica que la unión del agua (proveniente de las raíces de la planta) más dióxido de carbono (proveniente de la atmósfera) y en presencia de luz solar se produce oxígeno (hacia la atmósfera) y algún tipo de carbohidrato o azúcar (para la planta, incrementando su biomasa)(Baldocchi et al., 1987) (Figura 2).
Figura 2 Elementos que interactúan en la fotosíntesis (Bieto y Talón, 2008).
En otras palabras el proceso de fotosíntesis tiene una primera fase de absorción y conversión de energía luminosa en energía electroquímica redox, en esta fase la luz es el principal valor limitante. La segunda fase se conoce como de asimilación de elementos químicos esenciales para la síntesis bioquímica en donde la asimilación de los nutrientes inorgánicos,
incluidos el CO2 y el agua son los principales factores limitantes (Bieto y
Talón, 2008).
3.4.1 Ruta Fotosintética C3
La fotosíntesis es una serie de reacciones metabólicas que ocurren en presencia y ausencia de luz. A continuación se describen las dos tipos de reacciones.
JONATHAN ROMERO CUELLAR Página 19
células del cloroplasto. Luego la absorción de un fotón de luz eleva el nivel de energía de la molécula de clorofila. La molécula excitada no es estable y los electrones regresan rápidamente a su estado fundamental, liberando así la energía del fotón absorbido. Está energía se transfiere a otra molécula receptora para resultar en un proceso denominado transporte fotosintético de electrones. Mediante este proceso se produce la síntesis de trifosfato de adenosina (ATP) a partir del adenosín difosfato (ADP) y del nicotinamida adenina dinucleótido fosfato (NADPH) quien es un reductor fuerte (Smith y Smith, 2007).
Por otro lado, en las reacciones con ausencia de luz el CO2 se incorpora
bioquímicamente en los monosacáridos. Las reacciones oscuras no dependen directamente de la luz, pero sí de los productos de las reacciones lumínicas. El proceso en la fase oscura comienza cuando la molécula de Ribulosa 1,5 bifosfato (RuBP) de cinco carbonos se combina
con el CO2 para formar dos moléculas de un compuesto de tres carbonos
llamado fosfoglicolato (3-PGA):
→ Molécula de + Molécula de 2 moléculas de 1 Carbono 5 Carbonos 3 Carbonos
La ecuación anterior es una reacción denominada carboxilación y es catalizada por la enzima rubisco (ribulosa bifosfato carboxilasa-oxigenasa).
Las plantas convierten el 3-PGA en azúcar gliceraldehído 3-fosfato (G3P), para sintetizar G3P se necesita bastante ATP y NADPH. Del G3P una parte se utiliza para producir glucosa (mantenimiento de la planta) y el resto para sintetizar el nuevo RuBP con el fin de continuar el ciclo. Este ciclo se
conoce como el “ciclo de Calvin-Benson o ciclo C3” (Smith y Smith, 2007)
JONATHAN ROMERO CUELLAR Página 20 Figura 3 Representación de la ruta fotosintética C3 (Smith y Smith, 2007).
Según Bieto y Talón (2008) principalmente el ciclo de Calvin es manejado por los mecanismos reguladores de la rubisco y de la disponibilidad de
sustratos (CO2 y RuBP). De acuerdo con los compuestos obtenidos en el
ciclo de Calvin se sintetizan la sacarosa y el almidón. El proceso de fotosíntesis compite con el proceso de fotorespiración porque se producen
en simultánea en la hoja. La tasa de asimilación de CO2 (fotosíntesis neta)
es un balance entre el CO2 inicialmente fijado (fotosíntesis bruta) y las
pérdidas fotorespiratorias. Esta competencia se presenta por la competición entre carboxilación y oxigenación de rubisco, que depende de las propiedades cinéticas de la enzima, la temperatura ambiente y las
concentraciones de O2 y CO2.
En este sentido Smith y Smith (2007) agregan que la reacción competitiva
para el proceso de carboxilación reduce la eficiencia de las plantas C3.
Este dilema es un problema para las plantas porque algunos carbohidratos producidos en la fotosíntesis se utilizan en el proceso de respiración celular.
Finalmente bajo algunas condiciones ambientales por ejemplo: al incrementarse la temperatura, se favorece la actividad oxigenasa y se
disminuye la relación CO2/O2 disueltos en agua porque la solubilidad del
CO2 decrece más que la del O2. Por tanto, en conjunto, al aumentar la
temperatura, se favorece la fotorespiración. La eficiencia de la fotosíntesis se disminuye con la fotorespiración, pero puede ser un mecanismo de
protección frente a un exceso de luz en condiciones de bajo CO2 (Bieto y
JONATHAN ROMERO CUELLAR Página 21 3.4.2 Ruta Fotosintética C4
A diferencia de la ruta fotosintética C3 las plantas C4 tienen dos células
diferentes para hacer fotosíntesis, células del mesófilo (C3 y C4) y células en
empalizada vascular (solo C4). La ruta C4 posee ventajas frente a la C3. El
paso extra de fijación de CO2 en la ruta C4 hace que el fusfoenolpiruvato
[image:21.612.85.529.204.528.2](PEP) no interactúe con el oxígeno, como lo hace la RuBP esto elimina el dilema de catalizar carboxilaza o oxigenaza (Smith y Smith, 2007) (Figura 4).
Figura 4 Representación de la ruta fotosintética C4 (Smith y Smith, 2007).
Adicionalmente la conversión de los ácidos málicos y ásparticos en CO2
dentro de las células en empalizada hace que se concentre el CO2, esto
incrementa la eficiencia de la reacción entre el CO2 y la RuBP catalizada
por la rubisco. Por lo tanto las plantas tipo C4 tiene una mayor tasa
JONATHAN ROMERO CUELLAR Página 22 3.4.3 Ruta Fotosintética CAM
Es muy similar a la ruta C4 la única diferencia con la ruta CAM
(metabolismo ácido de las crasuláceas) es que esta ruta utiliza las células
del mesófilo para hacer los dos pasos de las C4, pero en momentos
diferentes.
De acuerdo con Smith y Smith (2007) las plantas tipo CAM capturan el CO2
en las noches y lo convierten en ácido málico durante el día, el vegetal
cierra sus estomas y reconvierte el ácido málico en CO2. Esta ruta es lenta
e ineficiente en la fijación de CO2 pero hace que las plantas no pierdan
agua por transpiración.
En este metabolismo el nitrógeno es un macronutriente principal porque participa activamente en la fotosíntesis, debido a que forma parte de la
rubisco (cataliza la transformación de CO2 en monosacáridos) y de la
clorofila (absorbe la energía de la luz). Por consiguiente, la tasa máxima de fotosíntesis (saturación de luz) está relacionada con el contenido de nitrógeno en la hoja (Smith y Smith, 2007).
3.4.4 Fotosíntesis en el contexto del cambio climático
Tanto por condiciones naturales o antrópicas la concentración de CO2 en
la atmósfera crece a razón de 2 ppm por año, junto con otros gases de efecto invernadero. A la larga la acumulación de dichos gases puede generar cambios climáticos regionales o globales reflejados en cambios bruscos de parámetros como: la precipitación, la temperatura, la humedad del suelo, el nivel del mar etc. Adicionalmente de acuerdo con las predicciones del Panel Intergubernamental del Cambio Climático (IPCC) para el año 2100 la temperatura aumentara de 1 a 3.5 °C y el nivel del mar aumentara de 15 a 95 cm. (Martínez y Patiño, 2010).
En la actualidad es un tema de interés para la ciencia los efectos del cambio climático sobre los ecosistemas terrestres y específicamente se requiere conocer cuáles serán las respuestas adaptativas de las plantas
frente a los cambios: del CO2 en la atmósfera, de temperatura y de
régimen hídrico. Básicamente el interés se centra en conocer el papel de la vegetación en la mitigación de los efectos del cambio climático, dada
su gran capacidad de absorción de CO2 por fotosíntesis (Bieto y Talón,
2008).
De hecho en primera instancia un aumento de CO2 produce un
JONATHAN ROMERO CUELLAR Página 23
tipo C3. Por otro lado, en presencia de temperaturas altas (40 – 50 °C) la
fotosíntesis se reduce rápidamente y la fotorespiración aumenta con gran
velocidad, hasta un punto donde todo el CO2 asimilado es liberado de
nuevo. Las plantas tiene la capacidad de aclimatarse de acuerdo con el cambio de régimen térmico esta es la principal característica de adaptación genotípica (Bieto y Talón, 2008).
Igualmente el aumento del CO2 atmosférico puede producir una
aclimatación de la fotosíntesis. Este fenómeno consiste en cambios bioquímicos que disminuyen la capacidad fotosintética, cuando las
plantas crecen continuamente con CO2 elevado y estos cambios pueden
ocurrir en semanas o meses. Se cree que la aclimatación de la fotosíntesis se debe a que la planta no es capaz de utilizar todos los carbohidratos adicionales y en segundo lugar en presencia de grandes cantidades de
Co2 la rubisco disminuye (Bieto & Talón 2008).
3.5 Interacciones de la capa límite en la hoja
La capa límite en la hoja es una capa de fluido (aire) que se mueve más lenta sobre los contornos de los cuerpos, debido a la acción de las fuerzas de fricción sobre una superficie. A menor velocidad del viento mayor es el espesor de la capa límite (Taiz y Zeiger, 2006).
Luego la capa límite se relaciona con el intercambio de calor y de gases entre un cuerpo (hojas) y la atmósfera, dichos cuerpos utilizan la ecofisiología vegetal y más específicamente la conductancia estomática para interactuar con la capa límite de igual forma intervienen la temperatura foliar, la transpiración y la fotosíntesis (Golberg, 2010).
En síntesis el grosor de la capa límite es función de la velocidad del viento y el tamaño de la hoja. Pronto la capa límite afecta el intercambio de calor entre la hoja y la atmósfera, el paso de vapor de agua de la cavidad del
estoma hacia la atmósfera, y la absorción de CO2 desde la atmósfera.
JONATHAN ROMERO CUELLAR Página 24
[image:24.612.81.533.222.499.2]Finalmente para tratar la complejidad de los procesos estudiados se presenta un diagrama causal de factores que interactúan entre sí; se asignó un signo positivo (+) cuando el aumento de la intensidad de un factor le corresponde un incremento correlativo del otro, por ejemplo: un aumento del DPV produce un aumento de la transpiración (Figura 5). El signo negativo (-) ha sido asignado a las interacciones en las cuales el aumento de un factor está asociado con la disminución del otro, por ejemplo, la disminución del contenido de agua en el suelo determina una disminución de la conductancia estomática.
Figura 5 Diagrama causal de las interacciones entre el ambiente y la planta. Adaptado de (Golberg, 2010).
Analizando una de las rutas del diagrama se observa que al aumentar la intensidad del viento se disminuye el espesor de la capa límite y esto genera un aumento en la conductancia de la capa límite que a su vez aumenta el intercambio de calor entre la hoja y el ambiente, ocasionando la disminución de la temperatura de la hoja. A su vez la esta disminución genera una disminución del déficit de presión de vapor en el estoma que ocasiona la disminución de la transpiración.
JONATHAN ROMERO CUELLAR Página 25
temperatura foliar ocasionada por el adelgazamiento de la capa límite produce una disminución de la transpiración; por otra parte, ese mismo adelgazamiento ocasionará un incremento de la transpiración al disminuir la resistencia al flujo de vapor de agua entre la hoja y la atmósfera (Golberg, 2010). Este tipo de fenómenos expresan la complejidad de los sistemas vivos.
Paralelamente la planta presenta un dilema funcional porque la producción de nueva biomasa (fotosíntesis) y el gasto de agua
(transpiración) ocurren en simultánea, la entrada de CO2 y la salida de
agua, utilizan la misma vía (los estomas de la hoja), entre más abierto está
el estoma más fácil entra el CO2 y sale el H2O. Por otro lado, el cierre
estomático en las horas más calurosas (medio día) produce la reducción
de la perdida de agua, entrada de CO2 y la disipación del calor a través
del enfriamiento evaporativo. Por lo tanto, la fotosíntesis disminuye y las temperaturas foliares pueden elevarse (Bieto y Talón, 2008).
Igualmente es importante recordar que el aumento de la temperatura genera un aumento de la presión de vapor y por lo tanto se aumenta el gradiente de humedad y la transpiración. Así mismo la tasa de respiración de una planta aumenta con el aumento de la temperatura (Smith y Smith, 2007).
3.6 Descripción del modelo implementado
El modelo representa proceso ecofisiológicos acoplados a nivel de hoja (fotosíntesis y conductancia estomática) con procesos físico (balance de energía y conductancia de la capa límite), procesos que determinan el
flujo de CO2, el vapor de agua y el intercambio de calor entre las hojas y el
ambiente. La interdependencia natural de los procesos a nivel de hoja y de follaje producen un conjunto acoplado de estrechas relaciones físicas y biológicas que controlan los flujos entre las plantas y la atmósfera (Drewry et al., 2010b).
JONATHAN ROMERO CUELLAR Página 26 3.6.1 Fotosíntesis C3
Luego de introducir los conceptos generales de la fotosíntesis, se presenta un modelo bioquímico de la ruta fotosintética características de las plantas
tipo C3. Los aportes de CO2 a las plantas se producen por difusión de gas a
través de la capa límite en la hoja, el poro estomático y la solución del mesófilo. En general, los estomas se abren o cierran respondiendo a las
variaciones ambientales para mantener la concentración de CO2
intercelular (Ci) a una fracción relativamente constante del CO2
atmosférico (Ca), si no, se presentan otros factores que obligue al cierre
estomático (Nikolov et al., 1995).
A continuación se muestran conceptos y modelos basados en procesos biológicos mecanísticos creados con el ánimo de sintetizar la información fisiológica y bioquímica a nivel de proceso y discutidos por (Farquhar et al., 1980; Collatz et al., 1991; Harley et al., 1992; Baldocchi y Meyers, 1998; Tuzet et al., 2003; Yin and Struik, 2009; Drewry et al., 2010a, 2010b; Gracia, 2011).
Por lo tanto, la tasa de asimilación neta de CO2 (An, μmol CO2 m-2 s-1) en la
hoja es:
Donde A es la tasa bruta de adsorción de CO2 (también llamada
fotosíntesis bruta) y Rd es la respiración oscura es decir producto de
procesos como la fotorespiración. La fotosíntesis bruta es definida por el factor más limitante de tres tasas potenciales:
{ }
Donde es la tasa de asimilación de CO2 restringida solamente por la
activación y las propiedades cinéticas de la enzima rubisco y representa el
mecanismo de carboxilación; es igualmente la tasa de asimilación pero
limitada solamente por la regeneración del sustrato de la enzima es decir la capacidad de regeneración de la ribulosa bifosfato (RuBP) dependiente
del flujo de electrones; y limita la tasa de asimilación por la capacidad
para utilizar los productos de la fotosíntesis, por ejemplo la síntesis desde la triosa fosfato en el ciclo de calvin.
Por consiguiente la dinámica fotosintética de las plantas no está limita
solamente por un factor potencial ( ) en realidad es el conjunto de
los tres y en forma de transición gradual que se presenta la tasa de
JONATHAN ROMERO CUELLAR Página 27
función continua con transiciones suaves por medio de dos ecuación cuadrática anidadas (Collatz et al., 1991):
( )
( )
Aquí es una tasa intermedia de asimilación de CO2 que representa el
mínimo de y , y son coeficientes convexos definidos por las raíces
de la transición. En este modelo y . La tasa de fotosíntesis
limitada por la rubisco (mecanismo de carboxilación):
Donde es la tasa máxima de carboxilación (μmol m-2 s-1), y son la
presión parcial (Pa) de CO2 y O2 en el espacio intercelular de la hoja, es
el punto de compensación (Pa) de CO2 en ausencia de respiración
oscura, y son parámetros cinéticos (Pa) de Michaelis-Menten para
CO2 y O2, respectivamente.
Por otro lado, la regeneración de la RuBP es controlada por la tasa del transporte de electrones. Asumiendo que el transporte del electrón es típicamente involucrado en la producción de adenosin trifosfato, la tasa de asimilación sería:
Donde es la tasa potencial del transporte de electrones (μmol m-2 s-1).
Collatz et al. (1991) define la tasa de asimilación de CO2 limitada por la
triosa fosfato como:
Luego, el punto de compensación de CO2 depende de la presión parcial
de O2 y de las propiedades cinéticas de la rubisco que a su vez dependen
enormemente de la temperatura. Por lo tanto, se utilizó un polinomio de segundo orden propuesto por Brooks y Farquhar (1985) para calcular el
punto de compensación de CO2
JONATHAN ROMERO CUELLAR Página 28
Donde es la temperatura de la hoja en (°C). También la tasa potencial
de transporte de electrones está limitada por la densidad de flujo de
fotones fotosintéticos incidentes (PPFD, por sus siglas en inglés) y la temperatura de la hoja. Una ecuación cuadrática es aproximada para
representar el control de PPFD y la temperatura de la hoja sobre .
√
Donde es un coeficiente de convexidad entre 0.7 y 0.9 para plantas C3,
es la tasa potencial de luz saturada del transporte de electrones, es
la intensidad de PPFD (ambos y están en μmol m-2 s-1), es la
eficiencia de conversión de energía para el transporte de electrones.
Donde (a) es el coeficiente de absorción para PPFD (0.85 para hojas
verdes) y ( ) es un factor de perdida de energía (por ejemplo la fracción
de PPFD absorbida disponible para fotosíntesis). El parámetro ( ) es
específico para cada especie y oscila entre 0.05 a 0.5 (Ziegler-Jöns y Selinger, 1987). Es necesario aclarar que en la ecuación [9] en condiciones de baja irradiancia, la regeneración de la RuPB es insensible a la temperatura.
La tasa potencial máxima de transporte de electrones es controlada
por la temperatura de la hoja (Farquhar et al., 1980)
[( )
]
[ ]
Donde es la tasa de a 25 °C, es la temperatura de la hoja en
°K, R es la constante universal de los gases (8.3143 J mol-1 K-1), E es la
energía de activación de la reacción (81993 J/mol), S y H son parámetros
relacionados con la entropía y la entalpia de la reacción (711.36 J mol-1 K-1
y 219814 J/mol respectivamente) estos parámetros fueron asumidos del
estudio realizo por Harley et al. (1992). Se presenta la dependencia de
con la temperatura:
[
]
JONATHAN ROMERO CUELLAR Página 29
Donde es la tasa de a 25 °C. Los parámetros y
dependen del tipo de especie, la fisiología, la edad de las hojas, las condiciones de crecimiento en términos de temperatura, concentración
atmosférica de CO2, la disponibilidad de luz, agua y nitrógeno mineral
(Seemann et al., 1984; Carter and Smith, 1985; Leuning et al., 1991; Harley et
al., 1992). Los parámetros cinéticos de CO2 y O2 son calculados:
[ ]
[ ]
Donde P es la presión atmosférica (Pa), y son parámetros
correspondientes a 25 °C (mol/mol). Estos parámetros varían con la
especie, pero se presentan unos valores típicos para plantas C3
y
(Woodrow y Berry, 1988). Además, la
respiración oscura (μmol CO2 m-2 s-1) aumenta exponencialmente con la
temperatura y se asume que es proporcional a la tasa de carboxilación a 25 °C,
[ ]
El coeficiente de proporcionalidad varía con la concentración
atmosférica de CO2 y el tipo de especie. En este trabajo de grado se utiliza
un valor sugerido por Collatz et al. (1991), , los mecanismo que
causan la inhibición de ante los efectos de luz son todavía
desconocidos, pero es incrementado linealmente hasta el 67% del valor
calculado por la ecuación [15] cuando PPFD < 50 (μmol m-2 s-1) (Brooks y
Farquhar, 1985).
Finalmente la concentración intercelular de CO2 depende de la tasa
neta de asimilación de CO2, la fisiología de la planta y la resistencia de la
capa límite en la superficie de la hoja. Farquhar et al., (1980) desarrollo una relación entre la fotosíntesis neta y la conductancia estomática:
Donde es la concentración de CO2 en el aire del ambiental (tanto
como tiene unidades de mol/mol), y son las conductancias al
vapor de agua (μmol m-2 s-1) del estoma de la hoja y de la capa límite
JONATHAN ROMERO CUELLAR Página 30
moleculares de vapor de agua y CO2 en condiciones de aire quieto. Esta
relación se reduce a 1.37 en la capa límite en la hoja debido al efecto del
flujo de aire. Para estimar la presión parcial de CO2 intercelular es
necesario multiplicar por la presión total (P) (Nikolov et al., 1995).
3.6.2 Conductancia Estomática
[image:30.612.82.528.288.570.2]El cierre estomático parcial reduce más intensamente la transpiración que la fotosíntesis, lo cual permite a las plantas continuar sus procesos de crecimiento (Figura 6). Las mayores remociones de vapor de agua en la hoja por efectos del viento ocurren a bajas velocidades, puesto que a velocidades altas tiene lugar el cierre estomático, bien por efectos mecánicos o por posible deshidratación (García Torres & Fuentes M 1994).
Figura 6 Esquema de la anatomía de la hoja de una planta C3 indicando las resistencias al flujo de entrada de C02 y de salida de vapor de agua a través del estoma (Gracia, 2011).
Ball-Berry (1982) presentan un modelo de origen empírico (basado en experimentos con intercambio de gases), pero que demuestra robustez al momento de capturar las respuestas de la conductancia estomática al
vapor de agua para diferentes condiciones ambientales y varias
especies C3 y C4 (Ball, 1988; Leuning et al., 1991). El modelo relaciona
con la asimilación de CO2 neta , la humedad relativa en la superficie
JONATHAN ROMERO CUELLAR Página 31
En esta ecuación (m) representa un parámetro adimensional específico de
cada especie que determina la sensibilidad de con . Este modelo
aparentemente simple esconde dependencias implícitas de la hoja como
Q y a través de la relación con . No obstante, tratar de modelar la
relación entre y no es fácil, porque esta relación no está entendida
conceptualmente del todo.
Por lo tanto, la ecuación [17] es una buena aproximación porque maneja elementos interesantes como: el establecimiento de una interrelación clara
entre y ; la ganancia de carbono y perdida de agua puede estar
influenciada por factores que afectan el grosor de la capa límite en la hoja como la velocidad del viento y la geometría de la hoja; esto presupone que la planta no responde directamente a las condiciones ambientales sino a su entorno inmediato es decir, la cavidad subestomática y la capa límite (Nikolov et al., 1995).
3.6.3 Condiciones intercelular y superficial de la hoja
Como la resistencia de la capa límite en la hoja siempre es mayor que cero, es de esperar que la humedad relativa en la superficie de la hoja es
mayor y la concentración de CO2 es menor que en el aire del ambiente.
Por consiguiente se plantea un balance de masas de vapor de agua y
CO2 entre la superficie de la hoja y la parte superior de la capa límite en la
hoja, se puede decir que para una superficie de hoja seca:
y
Donde es la presión de vapor de agua en el espacio intercelular de la
hoja, es la presión de vapor saturado a la temperatura de la hoja,
es la presión de vapor en la atmósfera (todas las presiones de vapor en Pa). En el caso donde la hoja tenga gotas de agua sobre su superficie, se asume que la ecuación [19] sigue siendo válida, suponiendo que las gotas de agua no afectan el intercambio gaseoso; por el lado de la ecuación [19] no hay problema porque simplemente se asume vapor de agua
JONATHAN ROMERO CUELLAR Página 32
En síntesis, en la superficie de la hoja con la capa límite formada por el flujo
de aire la concentración de CO2 es determinada por la ley de Fick’s en
una dimensión (Campbell y Norman, 1997).
3.6.4 Conductancia de la capa límite en la hoja
El flujo de aire sobre el follaje de la planta y las fuerzas de empuje por gradientes de temperatura entre la hoja y el ambiente son factores que afectan la conductancia de la capa límite en la hoja. Esta juega un papel importante en el acople y modulación entre el estoma y el ambiente (Collatz et al., 1991).
La conductancia en la capa límite se relaciona con la velocidad del viento y la dimensión característica de la hoja:
√
Donde (mol/m2*s), es la velocidad del viento en (m/s) y es la
dimensión característica de la hoja (m). La dimensión característica de la hoja se puede aproximar:
Siendo el grosor de la hoja en (m).
3.6.5 Balance de energía en la hoja
El balance de energía en las hojas determina , por lo tanto, la partición
de energía en color latente y sensible proporcionan un control sobre la fotosíntesis de las hojas (Drewry et al., 2010b). De hecho en el caso de estado estable (temperatura de la hoja constante), la energía absorbida por las dos caras de las hojas se disipa a través de los flujo de calor latente y sensible, la radiación de onda corta y el almacenamiento metabólico (Nikolov et al., 1995).
Donde es la energía absorbida desde ambos lados de la hoja (radiación
JONATHAN ROMERO CUELLAR Página 33
J kg-1 K-1), es la constante psicométrica (Pa/K), es la conductancia total
de la hoja al vapor de agua (m/s), es la temperatura ambiente (°C), es
la emisividad térmica de la hoja (alrededor de 0.975), es la constante de
Stefan-Boltzmann (5.67*10-8 W m-2 K-4) y es la energía almacenada por
efectos de reacciones bioquímicas la cual representa una fracción muy
pequeña de (0.506 J/ μmol CO2) y puede ser ignorada (Nikolov et al.,
1995).
Luego la conductancia total al vapor de agua para una superficie de hoja seca y la constante psicométrica son:
Y
Donde 0.622 es la proporción del peso molecular del agua en el aire seco, y L es el calor latente de vaporización (J/kg):
Finalmente, como la presión de vapor de agua saturado incrementa exponencialmente con la temperatura y puede ser calculado con una buena precisión:
[ ]
3.6.6 Acople de procesos en la hoja
Hasta el momento se presentan conceptos y sus equivalentes expresiones matemáticas, pero en este subcapítulo se desarrolla la forma de resolver numéricamente los procesos biofísicos y bioquímicos que ocurren en la hoja. Luego retomando algunos conceptos claves, se conoce que la tasa
neta de fotosíntesis (An) es controlada por la temperatura, PPFD y la presión
parcial de CO2 intercelular (Ci).
Sin embargo, (Ci) depende (An) y de la conductancia estomática ( ).
Igualmente esta última también depende de (An), y regula en conjunto
con la conductancia de la capa límite ( ) y la temperatura de la hoja
( ) al restringir la cantidad de energía absorbida por la superficie de la
JONATHAN ROMERO CUELLAR Página 34
del aire entre la superficie de la hoja y el ambiente, esto implica que la conductancia de la capa límite es sensible a la temperatura de la hoja y por lo tanto a la conductancia estomática (Collatz et al., 1991).
En ese orden de ideas los tres modelos (fotosíntesis, conductancia estomática y balance de energía en la hoja) son interdependientes. Por esta razón se implementó un procedimiento iterativo anidado para resolver el sistema numéricamente.
3.7 Cultivo de Maracuyá
3.7.1 Descripción botánica
Es un bejuco leñoso trepador, perenne del género passiflora de 5 a 8 m de altura, su periodo vegetativo no supera por lo general la década. Su tallo es rígido y leñoso, presenta hojas alternadas, gruesas, de gran tamaño (8 -20 cm de largo y 6 -5 cm de ancho), con márgenes finamente dentados, lisas de color verde oscuro. El sistema radicular es superficial y poco distribuido, sin raíz pivotante, las raíces generalmente se ubican en los primeros 30 cm de profundidad y alrededor del 80% se localizan a una distancia menor de 50 cm desde el tallo (Correa, 2004; Robert J. Knight, 2006).
3.7.2 Fotosíntesis y Transpiración del cultivo Maracuyá
Cuando comienza la actividad fotosintética, fijando el CO2 para ser
reducido a carbohidratos, los cuales sirven de sustrato principal de la respiración y cuya función es liberar la energía obtenida en forma de ATP y producir compuestos, comienza la producción de materia seca en la planta, como resultado neto del balance de estos procesos metabólicos básicos.
En etapa de producción, las hojas cercanas a los frutos transpiran más para traer nutrientes a los frutos. El cultivo de Maracuyá tiene un sistema
fotosintético tipo C3 esta característica se relaciona con una baja tasa de
JONATHAN ROMERO CUELLAR Página 35
De acuerdo con Correa (2004) en estudios realizados en la Universidad de Sao Paulo Brasil, el cultivo de maracuyá tiene una tasa de
evapotranspiración en promedio de 2.68 mm d-1.
3.7.3 Requerimientos climáticos y edáficos
El cultivo de Maracuyá se adapta a rangos de temperaturas desde los 21 hasta los 35 °C, con respecto a la altitud se cultiva desde el nivel del mar hasta los 1000 m y con humedad relativa del 60%. Requiere precipitación
de 800 – 1750 mm año-1 y una mínima mensual de 80 mm, las lluvias
intensas producen enfermedades fungosas y los periodos secos provocan la caída de las hojas y reducción del tamaño de los frutos (García, 2002).
De acuerdo con el CENTA (Centro Nacional de Tecnología Agropecuaria y Forestal) el maracuyá es una planta fotoperiódica que requiere mínimo de 11 horas de luz para poder florecer. Se puede cultivar en suelos desde arenosos hasta arcillosos, sueltos, bien drenados, con profundidad mínima
de 60 cm de profundidad, fertilidad media alta y pH de 5.5 – 7.0 también
es importante mencionar que las raíces del cultivo de maracuyá son muy susceptibles al daño por encharcamiento(García, 2002).
4.
Metodología, diseño experimental y descripción del caso
de estudio
Es necesario mencionar que los datos recolectados a nivel de campo son
producto del proyecto de investigación titulado “Optimización del manejo
del agua en los cultivos de Granadilla (Passiflora Ligularis Jus) y Maracuyá
(Passiflora edullis variedad flavicarpa) para mejorar la competitividad”
desarrollado por el Grupo de Investigación Hidroingeniería y Desarrollo
Agropecuario – GHIDA de la Universidad Surcolombiana en convenio con
JONATHAN ROMERO CUELLAR Página 36
4.1 Ubicación zona experimental
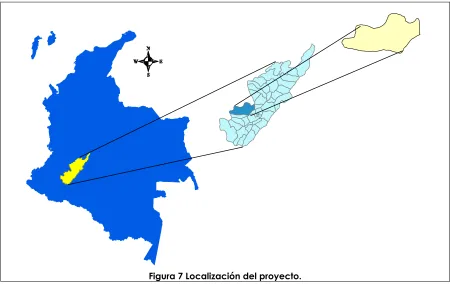
La zona experimental se ubica en la región sur-occidente de Colombia en el departamento del Huila, municipio de la Plata, vereda Fátima a 3 km del perímetro urbano del municipio y tiene un área total de 0.25 ha. El
experimento se encuentra a 2° 23’ 19.9’’ latitud norte y 75° 54’ 54.5’’
longitud oeste. La altitud del experimento es 1134 msnm, la temperatura promedio es 24 °C, humedad relativa del 72%, precipitación de 1600 mm año-1 y evaporación de 1180 mm año-1 y la velocidad del viento varía
entre 0.8 – 1.6 m s-1. El cultivo de maracuyá se dispuso en espaldera con
[image:36.612.79.529.264.548.2]riego por goteo (descarga de gotero 4 lph) y la distancia de siembra es de 2.5 m x 4 m entre calles y plantas respectivamente.
Figura 7 Localización del proyecto.
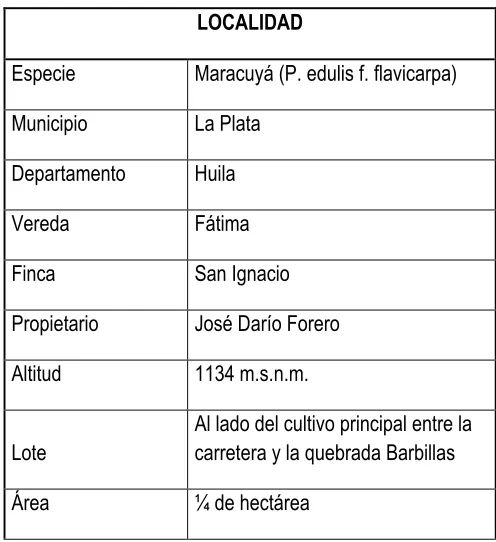
JONATHAN ROMERO CUELLAR Página 37 Tabla 1 Ubicación de la zona experimental.
LOCALIDAD
Especie Maracuyá (P. edulis f. flavicarpa)
Municipio La Plata
Departamento Huila
Vereda Fátima
Finca San Ignacio
Propietario José Darío Forero
Altitud 1134 m.s.n.m.
Lote
Al lado del cultivo principal entre la carretera y la quebrada Barbillas
Área ¼ de hectárea
4.2 Estación meteorológica automatizada
Los datos meteorológicos que soportan la información de entrada del modelo (Temperatura ambiente [°C], velocidad del viento [m s-1], radiación solar [w m-2] y humedad relativa [%]) proviene de una estación automatizada marca Weatherhawk. El periodo de información analizada corresponde del 20 de enero al 29 de diciembre de 2011. El equipo se programó para el registro de lecturas cada minuto y el promedio horario, por lo tanto se cuenta con un total de 8160 datos disponibles.
4.3 Protocolo de modelación implementado
En el contexto del presente trabajo de grado se utilizó la modelación matemática como método cognoscitivo, es decir, aplicación del protocolo de modelación en ingeniería como método para aprender procesos y analizar sistemas.
JONATHAN ROMERO CUELLAR Página 38
matemática (modelo matemático) la cual, implementa un algoritmo
lógico – numérico que permite estudiar las cualidades del proceso original.
Esto implica que el modelo seleccionado debe tener condiciones de analogía y similitud con el objeto de estudio.
De hecho la modelación matemática es muy utilizada en la actualidad para analizar sistemas y por ende resolver problemas. Esta herramienta es muy útil cuando el objeto de estudio es inviable, costoso o riesgoso. Adicionalmente la modelación matemática permite analizar las propiedades del objeto de estudio en situaciones inimaginables (Domínguez, 2010).
En ese orden de ideas es necesario presentar las fases del protocolo de modelación que se utilizaron en este trabajo:
I. Definir el objeto de la modelación, ¿Para qué modelar un
proceso? Esta pregunta nos permite preliminarmente establecer el tipo de modelo a utilizar, estimar la precisión que se requiere y el tiempo del proyecto.
II. Establecer el sistema de datos que luego de su procesamiento se
convierte en información (materia prima), luego esta materia prima es analizada y se convierte en conocimiento y finalmente cuando el conocimiento es utilizado para hacer el bien se convierte en sabiduría.
III. Formulación del modelo conceptual determina la complejidad
de los procesos a tomar en cuenta (suposiciones) y comprende la percepción del usuario sobre el proceso objeto de modelación. Tal vez esta fase sea la más importante del protocolo pues en esta se estudian las: teorías, mecanismos, escalas (temporal y espacial) propias del fenómeno estudiado.
IV. Selección del modelo matemático debido a que existen varios
operadores matemáticos disponibles el modelador debe escoger de acuerdo a su conveniencia, la complejidad del problema y la disponibilidad de información entre: ecuaciones diferenciales (determinísticas o estocásticas), sistemas lineales, dinámica no lineal, ecuaciones empíricas, estadística etc.
V. Selección del aplicativo computacional para lo cual se presentan
JONATHAN ROMERO CUELLAR Página 39
VI. Verificación de los modelos, generalmente son experimentos
controlados o llamados ejemplos de libros para confirmar que los modelos están funcionando bien.
VII. Calibración y validación, se utiliza un 70% de la información
disponible para identificar los parámetros del modelo y el 30% restante para validar la variable de salida del modelo por medio de alguna métrica de desempeño con el fin de probar el rango de bondad de ajuste (ej: Coeficiente de determinación de Nash
(R2)), una herramienta de utilidad para esta fase es el MCAT
(Monte-Carlo Analysis Tool).
VIII. Análisis de sensibilidad luego de la identificación de los
parámetros puede ser interesante conocer que tanta influencia tiene un determinado parámetro sobre la variable de salida, esto se realiza con el propósito de hacer futuras mejoras al modelo o de planificar salidas a campo para una parametrización adecuadamente del modelo.
IX. Crear escenarios en esta fase se formula la pregunta ¿Qué
pasaría si? Generalmente se hacen reuniones con los actores del problema y se plantea una prospección.
X. Simulación es donde el modelo es explotado matemáticamente y
luego se analizan los resultados, es decir, la información que brindan las simulaciones de utilidad para tomar decisiones.
4.4 Caso de estudio
4.4.1 Información meteorológica
De acuerdo con las variables climáticas que activan el modelo biofísico y bioquímico a nivel de hoja: Temperatura Ambiente [°C], Velocidad del
Viento [m/s], Humedad Relativa [%] y Radiación Neta en la Hoja [w/m2]. Se
presenta su información gráfica a nivel horario. Los datos disponibles son registrados cada hora desde el 21 de enero de 2011 hasta el 28 de diciembre de 2011 para un total de 8160 datos.
4.4.1.1 Temperatura ambiente
JONATHAN ROMERO CUELLAR Página 40
[image:40.612.82.535.128.430.2]porque determina la temperatura de las plantas junto con otros factores como: los flujos de calor radiante, latente, sensible y el almacenamiento de calor.
Figura 8 Representación de la serie de Temperatura ambiente registrada por la estación automatizada de la zona experimental registros horarios a lo largo del día.
La figura 8 es de utilidad para analizar la tendencia y la estructura de la señal, igualmente válida para poder afirmar que la temperatura ambiental de la zona experimental oscila a lo largo del día entre los 12 y los 32 °C.
Por otro lado, analizando la Figura 8 y 11 se observa que los máximos de temperatura se presentan un tiempo después de la incidencia de máxima energía radiante. Este tipo de retraso es común en sistemas con almacenamiento y resistencia (Campbell y Norman, 1997).
4.4.1.2 Humedad ambiental
JONATHAN ROMERO CUELLAR Página 41
[image:41.612.83.534.158.458.2](Allen et al., 1998). La humedad relativa es la segunda variable de entrada que direcciona el modelo, es importante en el transporte de energía, principalmente cuando hay cambios de fase asociados con el transporte de agua, grandes cantidades de energía pueden ser transferidas (Campbell y Norman, 1997).
Figura 9 Representación de la serie de humedad relativa registrada por la estación automatizada de la zona experimental registros horarios a lo largo del día.
En efecto la figura 9 representa un ambiente húmedo especialmente en horas de la mañana y la noche, donde se observan condiciones de saturación de vapor de agua, esta condición ambiental puede propiciar el crecimiento de hongos y plagas dañinos para el cultivo de maracuyá.
De igual forma, las plantas están muy pocas veces en equilibrio hídrico con su medio ambiente. Por lo tanto es importante mantener un adecuado balance interno de agua para el buen funcionamiento del cultivo (Monteith, 1965).
JONATHAN ROMERO CUELLAR Página 42
4.4.1.3 Velocidad del viento
[image:42.612.80.534.174.474.2]Es la tercera variable que direcciona el modelo presentado con anterioridad. Esta es altamente variable en el espacio y en el tiempo y participa en el transporte de calor y en la mezcla efectiva de la capa límite de la atmósfera de la tierra (Figura 10).
Figura 10 Representación de la serie de velocidad del viento registrada por la estación automatizada de la zona experimental registros horarios a lo largo del día.
En general el comportamiento del viento en la zona de estudio es errático pero se puede decir que conserva una banda media entre los 0 y 5 (m/s). El comportamiento del viento, es gobernado por las tasas de transferencia turbulentas en la capa límite de la superficie (Campbell y Norman, 1997).
4.4.1.4 Radiación neta absorbida por la hoja
JONATHAN ROMERO CUELLAR Página 43
[image:43.612.81.534.127.430.2]corta es la radiación (directa, reflejada o dispersa) originada en el sol; y la radiación de onda larga es la radiación originada en fuentes terrestres (Brutsaert, 1982).
Figura 11 Representación de la serie de radiación neta registrada por la estación automatizada de la zona experimental registros horarios a lo largo del día.
La figura 11 presenta un pico extremo al medio para todos los días estudiados.
4.4.1.5 Textura de las señales
JONATHAN ROMERO CUELLAR Página 44 Figura 12 Apariencia de las señales registradas por la estación automatizada de la zona
experimental cada hora.
La figura anterior es de mucha utilidad al momento de analizar tendencias, por ejemplo para el mes de marzo se aumenta exageradamente la velocidad del viento casi el triple de su valor normal, cuando este comportamiento es de esperarse para el mes de agosto.
5.
Herramienta computacional
Luego de estudiar la biofísica, bioquímica y la matemática del modelo a implementar, se ingresa a la fase V del protocolo de modelación (Ver sección 4.3). No obstante, debido a que estos modelos no son todavía de uso comercial es muy difícil encontrar software para utilizar en la modelación. Por esta razón y teniendo en cuenta que es más conveniente para el modelador estar en control de todos los procesos se decidió desarrollar una herramienta computacional propia.
0 1000 2000 3000 4000 5000 6000 7000 8000 9000 12 14 16 18 20 22 24 26 28 30 32
Serie de Temperatura Ambiente La Plata - Huila
Tiempo [h] T e m p e ra tu ra A m b ie n te [ °C ]
0 1000 2000 3000 4000 5000 6000 7000 8000 9000 10 20 30 40 50 60 70 80 90 100
Serie de la Humedad Relativa La Plata - Huila
Tiempo [h] H u m e d a d R e la tiv a [ % ]
0 1000 2000 3000 4000 5000 6000 7000 8000 9000 0
5 10 15
Serie de la Velocidad del viento La Plata - Huila
Tiempo [h] V e lo c id a d d e l v ie n to [ m /s ]
0 1000 2000 3000 4000 5000 6000 7000 8000 9000 0 100 200 300 400 500 600 700 800
Serie de la Radiación Neta absorbida por la hoja La Plata - Huila