Regulación de la síntesis de ácidos grasos en micobacterias
176
0
0
Texto completo
(2) Regulación de la síntesis de ácidos grasos en micobacterias. Sonia Soledad Mondino Licenciada en Biotecnología- Universidad Nacional de Rosario. Esta Tesis es presentada como parte de los requisitos para optar al grado académico de Doctor en Ciencias Biológicas, de la Universidad Nacional de Rosario y no ha sido presentada previamente para la obtención de otro título en esta u otra Universidad. La misma contiene los resultados obtenidos en investigaciones llevadas a cabo en el Laboratorio de Fisiología y Genética de Actinomycetes del Instituto de Biología Molecular y Celular de Rosario (IBR), dependiente de CONICET y de la Facultad de Ciencias Bioquímicas y Farmacéuticas, durante el período comprendido entre el 1 de abril de 2009 y el 3 de febrero de 2014, bajo la dirección de la Dra. Gabriela Gago y la codirección del Dr. Hugo Gramajo.. Directora Dra. Gabriela Gago ................................................ Codirector Dr. Hugo Gramajo ................................................ Dra. Fabiana Bigi ................................................ Dr. Diego José Comerci ................................................ Dr. Diego de Mendoza ................................................ Defendida .................................................
(3) Los resultados obtenidos durante el desarrollo de este trabajo de tesis Doctoral fueron dados a conocer en dos publicaciones internacionales y en distintas presentaciones a congresos científicos, como se detalla a continuación: Publicaciones 1- Mondino, S., Gago, G. and Gramajo, H. (2013) Transcriptional regulation of fatty acid biosynthesis in mycobacteria. Mol Microbiol 89, 372-387. 2- Salzman, V., Mondino, S., Sala, C., Cole, S.T., Gago, G. and Gramajo, H. (2010) Transcriptional regulation of lipid homeostasis in mycobacteria. Mol Microbiol 78, 6477. Presentación a Congresos Internacionales 1- Tuberculosis 2012: Biology, Pathogenesis, Intervention strategies, 2012. Mondino, Sonia; Gramajo, Hugo y Gago, Gabriela. “Transcriptional regulation of lipid biosynthesis in mycobacteria”. 2- Gordon Research Conferences: Tuberculosis Drug Development, 2011. Mondino, Sonia; Salzman, Valentina; Gago, Gabriela y Gramajo, Hugo. “Regulatory network of lipid biosynthesis in mycobacteria: possible new targets for drug discovery”. 3- Mycobacteria: Physiology, Metabolism and Pathogenesis- Back to the Basics, como parte de Keystone Symposia Global Health Series, 2011. Salzman, Valentina; Mondino, Sonia; Gago, Gabriela y Gramajo, Hugo. “Dissecting the regulatory network of lipid homeostasis in mycobacteria”. Presentación a Congresos Nacionales 1- IX Reunión de la Sociedad Argentina de Microbiología General (SAMIGE), 2013. Mondino, Sonia; Gramajo, Hugo y Gago, Gabriela. “Dissecting the transcriptional regulation of fas-acpS operon in mycobacteria”. 2- XLVIII Reunión anual de la Sociedad Argentina de Investigaciones Bioquímicas (SAIB), 2012. Mondino, Sonia; Gramajo, Hugo y Gago, Gabriela. “FasR, a novel positive regulator of the fas gene of mycobacteria”..
(4) 3- XLVII Reunión anual de la Sociedad Argentina de Investigaciones Bioquímicas (SAIB), 2011. Mondino, Sonia; Gramajo, Hugo y Gago, Gabriela. “FasR, a novel transcriptional activator of the fas gene of Mycobacterium tuberculosis”. 4- XLVI Reunión anual de la Sociedad Argentina de Investigaciones Bioquímicas (SAIB), 2010. Mondino, Sonia; Salzman, Valentina; Gramajo, Hugo y Gago, Gabriela. “Regulation of fatty acid biosynthesis in mycobacteria”. 5- IV Reunión de la Sociedad Latinoamericana de Tuberculosis y otras micobacteriosis (SLAMTB), 2009. Salzman, Valentina; Mondino, Sonia; Gago, Gabriela y Gramajo, Hugo. “Characterization of a transcriptional regulator of mycolic acid biosynthesis in Mycobacterium”. 6- XLV Reunión anual de la Sociedad Argentina de Investigaciones Bioquímicas (SAIB), 2009. Salzman, Valentina; Mondino, Sonia; Gago, Gabriela y Gramajo, Hugo. “MabR, a novel transcriptional regulator of mycolic acid biosynthesis in mycobacteria”. 7- XLIV Reunión anual de la Sociedad Argentina de Investigaciones Bioquímicas (SAIB), 2008. Salzman, Valentina; Mondino, Sonia; Gago, Gabriela y Gramajo, Hugo. “Characterization of a transcriptional regulator of mycolic acid biosynthesis in Mycobacterium”. 8- XLIII Reunión anual de la Sociedad Argentina de Investigaciones Bioquímicas (SAIB), 2007. Salzman, Valentina; Mondino, Sonia; Gago, Gabriela y Gramajo, Hugo. “Regulation of mycolic acid biosynthesis in mycobacteria”..
(5) Agradecimientos Regulación de la síntesis de ácidos grasos en micobacterias- Mondino, S. Agradecimientos Y aquí está, el momento en donde los abrazo fuerte a ustedes, y al universo: Gracias a la Facultad de Ciencias Bioquímicas y Farmacéuticas, al IBR y a CONICET, por la oportunidad de formarme profesionalmente y de trabajar en sus establecimientos. Gracias al NIH, a la Agencia Nacional de Promoción Científica y Tecnológica, y a la Fundación Bunge y Born por el financiamiento del presente trabajo de tesis doctoral. Gracias a la Fundación Josefina Prats por el reconocimiento otorgado, siendo estudiante de grado y de posgrado. Gracias a EMBO por la beca otorgada (EMBO-STF) para visitar el laboratorio del Dr. Stewart Cole en Lausanne (Suiza) durante tres meses. Gracias al Dr. Stewart Cole y a la Dra. Claudia Sala por su colaboración, y por la oportunidad de aprender y trabajar en su laboratorio durante tres meses. Gracias a la Dra. Gabriela Gago; directora, maestra y guía. Porque desde el primer día estuvo a mi lado, marcando a paso firme el camino a seguir, y poniendo un gran voto de confianza que valoro desde el corazón. Gracias al Dr. Hugo Gramajo; director y tutor. Por el apoyo, el impulso, los espacios de debate y la enseñanza. Gracias a ambos, por los brindis que acompañaron éxitos, y los empujes que remontaron tropiezos. Gracias del corazón. Gracias a mis viejos; pilares fundamentales, referentes y maestros, hoy y siempre. Por todo; la fuerza, el empuje, el amor, el ejemplo, las palabras, los abrazos, la compañía, la enseñanza y la vida. Los amo desde el alma. Gracias a ustedes, amigos y confidentes entre experimentos; dadores de abrazos y cebadores expertos de mates con debate; a aquellos que se fueron del laboratorio y los que vinieron, a los que descubren y los que enseñan. Gracias a todos ustedes, locos lindos que bailan conmigo cada día; su compañía es un regalo del cielo. Gracias a ellas, viajeras del mundo, maestras de la vida, confidentes del espíritu, sabias almas que el universo me regala cada día desde aquellos primeros días.
(6) Agradecimientos Regulación de la síntesis de ácidos grasos en micobacterias- Mondino, S. de facultad. Gracias mujeres, porque ustedes son uno de los mejores regalos que la vida me ha podido dar. Gracias infinitas, y más allá. Gracias a mi familia; cada uno de ustedes es ejemplo de vida. Gracias por ser, estar, pertenecer y latir conmigo. Me siento orgullosa de ser parte de ésta, nuestra historia. Y por último, gracias a todos aquellos amigos/confidentes/conocidos. A aquellos que cultivo desde los primeros años de secundaria, a los que se han perdido, a aquellos que van y vienen, a aquellos que perduran, a aquellos que están sin estar. Todos ustedes han dejado conmigo momentos que el tiempo no puede borrar; y que seguramente me robarán una sonrisa, algún día, a mitad de camino, mientras observe con la mirada perdida algo, evocándolos sin buscar. Gracias..
(7) Índice Regulación de la síntesis de ácidos grasos en micobacterias- Mondino, S. ÍNDICE ABREVIATURAS. VI. RESUMEN. 1. 1. INTRODUCCIÓN. 4. 1.1 TUBERCULOSIS. 4. 1.2 EL GÉNERO MYCOBACTERIUM. 8. 1.2.1 M. SMEGMATIS COMO MODELO DE ESTUDIO. 9. 1.3 ENVOLTURA CELULAR DE LAS MICOBACTERIAS. 10. 1.3.1 MEMBRANA PLASMÁTICA. 12. 1.3.2 PARED CELULAR. 12. 1.3.3 LÍPIDOS LIBRES. 13. 1.4 BIOSÍNTESIS DE ÁCIDOS GRASOS EN BACTERIAS. 14. 1.5 ASPECTOS. GENERALES DE LA BIOSÍNTESIS DE ÁCIDOS GRASOS Y ÁCIDOS MICÓLICOS EN LAS. MICOBACTERIAS. 17. 1.5.1 CARACTERÍSTICAS DE LOS SISTEMAS FAS-I Y FAS-II MICOBACTERIANOS. 17. 1.5.2 BIOSÍNTESIS DE ÁCIDOS GRASOS Y ÁCIDOS MICÓLICOS EN MICOBACTERIAS. 18. 1.6 ANTECEDENTES SOBRE LA REGULACIÓN DE LA BIOSÍNTESIS DE ÁCIDOS GRASOS EN BACTERIAS. 25. 1.6.1 ANTECEDENTES. SOBRE LA REGULACIÓN DE LA SÍNTESIS DE ÁCIDOS GRASOS Y MICÓLICOS EN LAS. MICOBACTERIAS. 32. 1.6.1.1 Antecedentes sobre el control bioquímico de la síntesis de lípidos en micobacterias. 32. 1.6.1.2 Regulación transcripcional de los genes de la biosíntesis de ácidos micólicos 34 2. MATERIALES Y MÉTODOS. 39. 2.1 CEPAS BACTERIANAS Y PLÁSMIDOS. 39. 2.2 MEDIOS DE CULTIVO Y CONDICIONES DE CRECIMIENTO BACTERIANO. 43. 2.3 TRANSFORMACIÓN DE LAS CEPAS BACTERIANAS. 44. 2.3.1 TRANSFORMACIÓN DE E. COLI. 44. 2.3.2 TRANSFORMACIÓN DE M. SMEGMATIS. 45 I.
(8) Índice Regulación de la síntesis de ácidos grasos en micobacterias- Mondino, S. 2.3.3 TRANSFORMACIÓN DE M. TUBERCULOSIS. 45. 2.4 MANIPULACIÓN DE ADN Y CONSTRUCCIÓN DE PLÁSMIDOS. 46. 2.4.1 TRATAMIENTO CON ENZIMAS DE RESTRICCIÓN Y LIGACIÓN DE FRAGMENTOS DE ADN. 46. 2.4.2 ELECTROFORESIS DE ADN EN GELES DE AGAROSA. 46. 2.4.3 PURIFICACIÓN DE FRAGMENTOS DE ADN A PARTIR DE GELES DE AGAROSA. 46. 2.4.4 SECUENCIACIÓN DE NUCLEÓTIDOS Y ANÁLISIS DE SECUENCIAS. 47. 2.4.5 EXTRACCIÓN DE ADN. 47. 2.4.5.1 Extracción de ADN genómico de M. smegmatis. 47. 2.4.5.2 Extracción de ADN genómico de M. tuberculosis. 47. 2.4.5.3 Extracción de ADN plasmídico de E. coli. 48. 2.4.6 CONSTRUCCIÓN DE PLÁSMIDOS. 48. 2.4.7 REACCIÓN EN CADENA DE LA POLIMERASA (PCR). 53. 2.4.8 ENSAYOS DE RETARDO DE LA MOVILIDAD ELECTROFORÉTICA EN GEL (EMSA). 53. 2.4.8.1 EMSA con extractos proteicos libres de células. 53. 2.4.8.2 EMSA con la proteína purificada His6-FasRMT. 54. 2.4.9 ENSAYO DE PROTECCIÓN A LA DNASA I. 55. 2.4.10 TRANSFERENCIA DE ADN A MEMBRANAS DE NYLON. E HIBRIDIZACIÓN CON SONDAS MARCADAS. (SOUTHERN BLOT). 56. 2.4.10.1 Southern blot de la cepa mutante condicional MSfasR pFR5. 56. 2.4.10.2 Southern blot de la cepa mutante condicional MtbfasR pFR33. 57. 2.5 ANÁLISIS DE PROTEÍNAS. 57. 2.5.1 AISLAMIENTO E IDENTIFICACIÓN DE FASR. 57. 2.5.2 EXPRESIÓN Y PURIFICACIÓN DE FASRMT. 58. 2.5.3 DETERMINACIÓN DEL PESO MOLECULAR DE FASRMT. 59. 2.5.4 DOSAJE DE PROTEÍNAS Y SDS-PAGE. 59. 2.5.4.1. Ánalisis en M. smegmatis. 59. 2.5.4.2. Ánalisis en M. tuberculosis. 60. 2.5.5 TRANSFERENCIA E INMUNODETECCIÓN DE PROTEÍNAS (WESTERN BLOT). 61. 2.5.5.1 Inmunización con FasR y obtención de suero. 61. 2.5.5.2 Western blot con extractos proteicos de M. smegmatis. 61. 2.5.5.3 Western blot con extractos proteicos de M. tuberculosis. 62. 2.5.6 ENSAYOS DE -GALACTOSIDASA. 63 II.
(9) Índice Regulación de la síntesis de ácidos grasos en micobacterias- Mondino, S. 2.6 MANIPULACIÓN DE ARN. 65. 2.6.1 EXTRACCIÓN DE ARN DE MYCOBACTERIUM. 65. 2.6.2 RETROTRANSCRIPCIÓN Y AMPLIFICACIÓN POR PCR (RT-PCR). 65. 2.6.3 DETERMINACIÓN DEL SITIO DE INICIO DE LA TRANSCRIPCIÓN. 65. 2.6.4 PCR EN TIEMPO REAL (QRT-PCR). 66. 2.7 ANÁLISIS DE LÍPIDOS. 67. 2.7.1 ANÁLISIS DE LÍPIDOS POR CROMATOGRAFÍA EN CAPA DELGADA (TLC). 67. 2.8 ANÁLISIS DE LA BIOSÍNTESIS DE MACROMOLÉCULAS. 68. 2.9 ANÁLISIS ESTADÍSTICO. 68. 3. RESULTADOS Y DISCUSIÓN. 69. 3.1 CAPÍTULO 1 IDENTIFICACIÓN DE FASR, UNA PROTEÍNA REGULADORA DEL OPERÓN FAS-ACPS. 69. 3.1.1 OBJETIVOS. 70. 3.1.1.1 Objetivo general. 70. 3.1.1.2 Objetivos particulares. 70. 3.1.2 RESULTADOS. 71. 3.1.2.1 Caracterización de la región promotora del gen fas en M. tuberculosis y M. smegmatis. 71. 3.1.2.2 Identificación de un factor de transcripción de la familia TetR que se une a la región promotora del operón fas-acpS en Mycobacterium. 72. 3.1.2.3 Expresión y purificación de FasR, un regulador transcripcional de la familia TetR que se une específicamente a la región promotora del operón fas-acpS. 75. 3.1.2.4 Determinación del estado oligomérico de FasR en solución. 78. 3.1.2.5 Identificación de los sitios de unión de FasRMT en la región promotora del operón fas-acpS de M. tuberculosis. 78. 3.1.2.6 FasR reconoce y se une a una secuencia palindrómica conservada en micobacterias. 79. 3.1.2.7 FasR es un regulador altamente conservado en actinomicetales productores de ácidos micólicos. 83. 3.1.3 DISCUSIÓN. 85. 3.2 CAPÍTULO 2 III.
(10) Índice Regulación de la síntesis de ácidos grasos en micobacterias- Mondino, S. FASR. ES UN ACTIVADOR TRANSCRIPCIONAL DEL OPERÓN FAS-ACPS MODULADO POR ACIL-COAS DE. CADENA LARGA. 88. 3.2.1 OBJETIVOS. 89. 3.2.1.1 Objetivo general. 89. 3.2.1.2 Objetivos particulares. 89. 3.2.2 RESULTADOS. 90. 3.2.2.1 Construcción de cepas en M. smegmatis para el análisis de fusiones transcripcionales de PfasMT al gen de la -galactosidasa (lacZ) de E. coli. 90. 3.2.2.2 FasRMT es un activador del operón fas-acpS. 91. 3.2.2.3 Las cajas de reconocimiento de FasRMT son esenciales para la función del regulador. 92. 3.2.2.4 Búsqueda de la molécula efectora de FasRMT. 93. 3.2.2.5 Los acil-CoAs de cadena larga modulan la unión de FasRMT a la región promotora del operón fas-acpS. 96. 3.2.3 DISCUSIÓN. 99. 3.3 CAPÍTULO 3 ESTUDIO DEL ROL FISIOLÓGICO DE FASR EN M. SMEGMATIS. 102. 3.3.1 OBJETIVOS. 103. 3.3.1.1 Objetivo general. 103. 3.3.1.2 Objetivos particulares. 103. 3.3.2 RESULTADOS. 104. 3.3.2.1 Construcción de una cepa de M. smegmatis mutante en fasR, utilizando el plásmido termosensible pPR27. 104. 3.3.2.2 Construcción de una cepa de M. smegmatis mutante condicional en fasR, utilizando el sistema TetR/Pip OFF. 107. 3.3.2.3 Caracterización de la cepa MSPptr:fasRMS. 111. 3.3.2.4 FasR es un activador del operón fas-acpS in vivo. 113. 3.3.2.5 Influencia de los niveles de expresión de fasR sobre la expresión del operón fasII in vivo. 116. 3.3.2.6 Construcción de una cepa de M. smegmatis mutante condicional en el operón fas-acpS, utilizando el sistema TetR/Pip OFF. 118. 3.3.2.7 Caracterización de la cepa MSPptr:fasMS. 118 IV.
(11) Índice Regulación de la síntesis de ácidos grasos en micobacterias- Mondino, S. 3.3.3 DISCUSIÓN. 124. 3.4 CAPÍTULO 4 DETERMINACIÓN DE LA ESENCIALIDAD DE FASR EN M. TUBERCULOSIS. 128. 3.4.1 OBJETIVOS. 129. 3.4.1.1 Objetivo general. 129. 3.4.1.2 Objetivos particulares. 129. 3.4.2 RESULTADOS. 131. 3.4.2.1 Expresión de FasR en M. tuberculosis en cultivos in vitro. 131. 3.4.2.2 Construcción de una cepa de M. tuberculosis mutante condicional en fasR utilizando el sistema TetR/Pip OFF. 131. 3.4.2.3 Caracterización de la cepa MtbfasR pFR33. 135. 3.4.3 DISCUSIÓN. 137. 4. CONCLUSIONES. 140. 5. REFERENCIAS BIBLIOGRÁFICAS. 142. V.
(12) Abreviaturas Regulación de la síntesis de ácidos grasos en micobacterias- Mondino, S. Abreviaturas ACC. Acetil-CoA carboxilasa. ACP. Proteína transportadora de acilos. ADN. Ácido desoxirribonucleico. ADNc. ADN complementario. AG. Arabinogalactano. AGI. Ácidos grasos insaturados. AGS. Ácidos grasos saturados. Am. Apramicina. ARN. Ácido ribonucleico. ARNm. ARN mensajero. ARNt. ARN de transferencia. ATc. Anhidrotetraciclina. ATP. trifosfato de adenosina. BCIP. sal de toluidina 5-bromo-4-cloro-3'-indol fosfato. BSA. Albúmina sérica bovina. ChIP-seq. Inmunoprecipitación de la cromatina seguida de secuenciación. CoA. Coenzima A. DAT. Diacil-trehalosas. DNasaI. Desoxirribonucleasa I. dNTPs. Desoxirribonucleótidos. DO. Densidad óptica. DTT. Ditiotreitol. EDTA. Ácido etilendiaminotetraacético. EMB. Etambutol. ETH. Etionamida. FAMEs. Metilésteres de ácidos grasos. FAS. Ácido graso sintasa. G+C. Guanina y Citosina VI.
(13) Abreviaturas Regulación de la síntesis de ácidos grasos en micobacterias- Mondino, S. Gm. Gentamicina. GPL. Glicopeptidolípidos. Hyg. Higromicina. INH. Isoniacida. IPTG. Isopropil--D-tiogalactopiranósido. Km. Kanamicina. LAM. Lipoarabinomanano. LM. Lipomanano. LOS. Lipooligosacáridos. mAGP. Complejo micolil-arabinogalactano-peptidoglicano. MAMEs. Metilésteres de ácidos micólicos. MDR-TB. Tuberculosis multiresistente. NBT. Cloruro de nitroblue tetrazolium. ONPG. orto-nitrofenil-β-D-galactopiranósido. PAT. Pentacil-trehalosas. PCR. Reacción en cadena de la polimerasa. PDIM. Tiocerol dimicocerosato. PG. Peptidoglicano. PGL. Glicolípidos fenólicos. PIMs. Fosfatidilinositolmanósidos. Pip. Pristinamicina. PMSF. Fluoruro de fenilmetilsulfonilo. PPD. Derivado de proteína purificada. PZN. Pirazinamida. qRT-PCR. PCR en tiempo real. RACE. amplificación rápida de los extremos de ADNc. RIF. Rifampicina. RT. Reacción de retrotranscripción. SDS. Dodecil sulfato de sodio VII.
(14) Abreviaturas Regulación de la síntesis de ácidos grasos en micobacterias- Mondino, S. SDS-PAGE. Electroforesis en geles de poliacrilamida en presencia de SDS. SIDA. Síndrome de inmunodeficiencia adquirida. SL. Sulfolípidos. Sm. Estreptomicina. Sp. Espectinomicina. STPK. Serina/treonina proteína quinasa. TAGs. Triacilglicéridos. TB. Tuberculosis. TDM. Trehalosa dimicolato. TLC. Cromatografía en capa delgada. TLM. Tiolactomicina. TMM. Trehalosa monomicolato. TRC. Triclosan. Tris. N‐tris‐(hidroximetil) aminometano. TSS. Sitio de inicio de la transcripción. TST. Prueba cutánea de la tuberculina. VIH. Virus de la Inmunodeficiencia Humana. XDR-TB. Tuberculosis extremadamente resistente. VIII.
(15) Resumen Regulación de la síntesis de ácidos grasos en micobacterias- Mondino, S. Resumen Entre las enfermedades infecciosas, la tuberculosis (TB) continúa siendo una de las principales causas de muerte entre los adultos. Algunos años atrás se pensó que esta enfermedad estaba controlada y que sería erradicada a mediano plazo. Sin embargo, hoy en día la TB está restablecida debido a diversos factores, entre ellas la aparición del SIDA. Mycobacterium tuberculosis, el agente etiológico de la tuberculosis, presenta una pared celular inusual, característica de todas las micobacterias. Esta envoltura celular resulta esencial para la viabilidad y supervivencia de las mismas en ambientes hostiles y consiste de una capa altamente impermeable de ácidos micólicos de 70-90 átomos de carbono unidos covalentemente al peptidoglicano (PG) a través de un polisacárido conector, el arabinogalactano (AG). Los ácidos micólicos son los componentes mayoritarios de la envoltura celular de las micobacterias y juegan un rol crucial en su compleja arquitectura y en su impermeabilidad. La biosíntesis de los ácidos micólicos requiere de dos tipos de sintasas de ácidos grasos (FAS): la enzima multifuncional FAS-I, similar a la presente en eucariotas, y el sistema dependiente de la proteína transportadora de acilos (ACP), FAS-II, el cual consiste de una serie de enzimas donde cada una cataliza un paso en la vía de elongación de acil-CoAs de cadena mediana C12-C16, previamente sintetizados por FAS-I. Los componentes genéticos del sistema FAS-II han sido identificados en M. tuberculosis y se encuentran agrupados en tres unidades transcripcionales principales: fabD-acpM-kasA-kasB-accD6 (operón fasII), mabA-inhA y hadA-hadB-hadC. Análisis de microarreglos demostraron que el tratamiento de M. tuberculosis con diversos antibióticos que afectan la síntesis de ácidos micólicos, como isoniacida (INH), etionamida (ETH) o tiolactomicina (TLM), inducen la transcripción de los genes de operón fasII. A su vez fas, el gen que codifica para la enzima multifuncional FAS-I, también se induce tras el tratamiento de M. tuberculosis con INH, sugiriendo la existencia de señales regulatorias comunes a los dos sistemas FAS. Esta información junto al concepto general sobre la existencia de sistemas reguladores que controlan la homeostasis lipídica en la mayoría de los organismos, llevó a que nuestro grupo de investigación se propusiera estudiar quién y cómo se regulan a nivel transcripcional los sistemas de síntesis de ácidos grasos en. 1.
(16) Resumen Regulación de la síntesis de ácidos grasos en micobacterias- Mondino, S. micobacterias. Es así que fuimos capaces de identificar y caracterizar una proteína reguladora del operón fasII, a quien llamamos MabR (por sus siglas en inglés, Mycolic acid biosynthesis Regulator). Los resultados de esta investigación representaron la primera caracterización de un regulador clave para el metabolismo de ácidos grasos en M. tuberculosis y sentaron las bases para el desarrollo del trabajo de tesis aquí presentado. Los estudios de microarreglos y proteómica antes detallados, junto con la evidencia de que la transcripción del gen fas se veía afectada cuando alterábamos los niveles fisiológicos de MabR, sugirieron la existencia de un mecanismo de regulación coordinado entre los dos sistemas FAS mediado por MabR. La identificación de una repetición invertida en la secuencia promotora del gen fas, similar a la reconocida por MabR en la región promotora del operón fasII, nos llevó a pensar que MabR podría estar regulando de manera directa la transcripción del mismo. Sin embargo, la incapacidad de evidenciar esta interacción in vitro nos condujo a la búsqueda de un nuevo regulador transcripcional del sistema FAS-I. Para alcanzar los objetivos propuestos, en el presente trabajo de tesis se caracterizó la región promotora del gen fas de M. tuberculosis y Mycobacterium smegmatis, comprobando que este gen forma parte de un operón al que denominamos operón fas-acpS. Se identificó y purificó una proteína reguladora de dicho operón, denominada FasR (por sus siglas en inglés, Fatty acid synthase Regulator), la cual fue caracterizada mediante diversos análisis bioquímicos y genéticos. Pudimos determinar que FasR se une a tres repeticiones de una secuencia operadora conservada, en la región promotora del operón fas-acpS. Estudios in vitro e in vivo demostraron que FasR es un activador transcripcional esencial en M. smegmatis, cuya afinidad por la región promotora del operón fas-acpS es modulada por acil-CoAs de cadena larga, productos del sistema FAS-I. La mayoría de los experimentos realizados en este trabajo de tesis utilizaron a M. smegmatis como sistema modelo, elección que responde a razones prácticas. En conclusión, los resultados obtenidos en el presente trabajo de tesis, junto con aquellos previamente publicados por nuestro grupo, sugieren que los dos sistemas FAS deben estar estrictamente co-regulados a nivel transcripcional para mantener la homeostasis lipídica en las micobacterias, y que la disrupción o alteración de dicha. 2.
(17) Resumen Regulación de la síntesis de ácidos grasos en micobacterias- Mondino, S. comunicación conduce a un microorganismo altamente comprometido en su viabilidad y/o capacidad infectiva.. 3.
(18) Introducción Regulación de la síntesis de ácidos grasos en micobacterias- Mondino, S. 1. Introducción 1.1 Tuberculosis La tuberculosis (TB) es una enfermedad infecciosa causada por especies del género Mycobacterium, pertenecientes a un grupo genéticamente relacionado denominado complejo Mycobacterium tuberculosis. El agente etiológico más representativo de esta enfermedad es M.tuberculosis, identificada por primera vez como una especie patógena por Robert Koch en 1882, de allí su denominación como bacilo de Koch [1]. La TB es predominantemente una enfermedad pulmonar (tuberculosis pulmonar) representando el 70% de los casos; sin embargo M. tuberculosis puede diseminarse a otros órganos, incluyendo los nódulos linfáticos, huesos y meninges, provocando una enfermedad extrapulmonar (tuberculosis miliar o extrapulmonar) en 10-42% de los pacientes dependiendo de la edad, estado inmune y grupo étnico [2, 3]. El contagio ocurre por vía aérea cuando un individuo enfermo de tuberculosis pulmonar expele bacterias a través de la tos, el estornudo o la salivación. Si bien basta con que una persona inhale unos pocos bacilos para quedar infectada, en general, una baja proporción de los individuos infectados desarrollan la forma activa de la enfermedad. La mayoría de los individuos son asintomáticos y se piensa que mantienen la infección en una forma latente. Los informes estadísticos estiman que entre el 5-10% de éstos desarrollará la forma activa de la TB durante el transcurso de su vida, fenómeno denominado reactivación o tuberculosis post-primaria (Fig. 1). Este riesgo de reactivación es mucho mayor para las personas cuyo sistema inmunitario está deprimido, como ocurre en casos de personas infectadas con el virus de la Inmunodeficiencia Humana (VIH), desnutrición, diabetes mellitus o abuso de drogas, alcohol o tabaco. El impacto de la TB a nivel mundial sigue siendo extremadamente elevado. En 2012, 8.6 millones de personas enfermaron de TB y 1.4 millones murieron por esta enfermedad [4]. Se estima que una tercera parte de la población mundial está infectada con M. tuberculosis en forma latente [5]. La infección o exposición a esta bacteria puede demostrarse a través de la reactividad del huésped a antígenos del patógeno, mediante la prueba cutánea de la tuberculina (TST). En esta, el paciente es enfrentado intradérmicamente a un extracto de M. tuberculosis, actualmente 4.
(19) Introducción Regulación de la síntesis de ácidos grasos en micobacterias- Mondino, S. compuesto por un cóctel complejo de antígenos conocido como derivado de proteína purificada (PPD) [6]. Sin embargo, los antígenos presentes en la PPD también están en la vacuna BCG, obtenida a partir de cepas de Mycobacterium bovis atenuadas, y en otras micobacterias ambientales no tuberculosas, haciendo que la detección de las personas infectadas con M. tuberculosis esté sobre-estimada y no sea tan específica como se desea.. Figura 1. Epidemiología de la infección por M. tuberculosis- TB activa y latente. La TB resulta de la infección con el patógeno M. tuberculosis, el cual se contagia por transmisión aérea. La mayoría de los individuos infectados no desarrolla la forma activa de la enfermedad. Se estima que un tercio de la población mundial está infectada con M. tuberculosis, sin embargo se mantienen asintomáticos, presentando una forma latente de TB. Estudios epidemiológicos estiman que sólo el 5-10% de estos individuos desarrollarán la forma activa de la enfermedad a lo largo de su vida. Esquema obtenido de O´Garra y colaboradores [3].. El tratamiento de la TB activa involucra un régimen con cuatro drogas de primera línea: isoniazida (INH), rifampicina (RIF), pirazinamida (PZN) y etambutol (EMB), alcanzándose un 90% de recuperación en los pacientes bajo tratamiento. El proceso es prolongado, abarca un mínimo de seis meses dividido en dos fases: una 5.
(20) Introducción Regulación de la síntesis de ácidos grasos en micobacterias- Mondino, S. fase intensiva inicial de dos meses con las cuatro drogas, necesaria para matar los bacilos en división activa, seguida por una fase de continuación de cuatro meses con INH y RIF para asegurar que los bacilos persistentes también sean eliminados. Durante la segunda mitad del siglo XX se pensó que la TB estaba controlada. Sin embargo, la aparición en 1981 del síndrome de inmunodeficiencia adquirida (SIDA) causado por el VIH, la intensificación de las migraciones masivas y las condiciones de pobreza en muchos países subdesarrollados (principalmente en Asia y África) fueron generando un terreno propicio para el resurgimiento de enfermedades que se creían en retroceso como la TB. Como consecuencia del incremento en el número de casos de personas infectadas, en 1993 la Organización Mundial de la Salud declara a esta enfermedad como una emergencia de la salud pública a nivel mundial. El control de la epidemia de TB se vió dificultada por la falta de una vacuna efectiva, de diagnósticos rápidos y eficaces y fundamentalmente por la emergencia de cepas de M. tuberculosis resistentes a múltiples drogas. La TB fármaco-resistente aparece principalmente como consecuencia del incumplimiento efectivo del tratamiento. La TB multiresistente (MDR-TB) es una forma de TB resistente a por lo menos dos de las drogas más importantes utilizadas para el tratamiento de esta enfermedad, INH y RIF [7]. En el año 2006 se han reportado casos de pacientes infectados con cepas de TB extremadamente resistente (XDR-TB), que presentaban resistencias adicionales a dos de las drogas de segunda línea más efectivas: las fluoroquinolonas y al menos una de las tres drogas inyectables de segunda línea, amikacina, kanamicina o capreomicina. En años más recientes se han detectado en distintos países, casos de TB con patrones de resistencia aún más amplios [8-10]. La aparición de estas formas resistentes de TB representa una de las amenazas más importantes para el control de esta enfermedad a nivel mundial. En la mayoría de los casos, se recomienda un tratamiento de veinte meses y requiere de la administración diaria de drogas más costosas, más tóxicas y menos efectivas que aquellas usadas para tratar las formas susceptibles de TB. A su vez, los pacientes infectados que no cumplen adecuadamente con el tratamiento, se vuelven portadores crónicos y diseminan cepas resistentes en su familia y comunidad. En Argentina, la TB no está erradicada y continúa siendo un problema de salud pública. En 2012 se han registrado cerca de 15 000 casos de TB, de los cuales 10 000 han sido nuevos [4]. Existe aún una brecha considerable entre las diferentes 6.
(21) Introducción Regulación de la síntesis de ácidos grasos en micobacterias- Mondino, S. jurisdicciones del país, con elevada concentración de casos en provincias como Salta, Jujuy y Formosa. A su vez, la provincia de Buenos Aires y la Ciudad Autónoma de Buenos Aires notifican la mitad de los casos de todo el país. La pobreza, la marginalidad y el hacinamiento que experimenta gran parte de la población constituyen un obstáculo en la lucha contra esta enfermedad a nivel nacional. La habilidad de M. tuberculosis de persistir bajo la forma de una infección asintomática por largos períodos de tiempo es un tema central en la biología de la enfermedad. Como consecuencia de ello, aún curando clínicamente al paciente, no se logra erradicar completamente al bacilo. En este estado, la mayoría de las bacterias están en un estado de dormancia, no replicativo y con baja actividad metabólica, y sólo una pequeña población se encuentra en división activa [11]. Entre las causas de su persistencia se encuentra la propia naturaleza del proceso infectivo, que genera la ocupación de numerosas cavidades en el huésped con un gran número de bacilos; la resistencia fenotípica del estado dormante que representa uno de los mayores problemas para las quimioterapias y la capacidad limitada del sistema inmune de eliminar los bacilos presentes en las lesiones luego del tratamiento. Se considera que en este estado latente el uso de INH, que actúa sobre las bacterias en división activa, reduce el riesgo de reactivación mientras que la RIF colabora en la eliminación de la subpoblación en estado dormante. Esta resistencia intrínseca de M. tuberculosis está relacionada con la barrera de impermeabilidad que le proporciona su compleja pared celular [12]. En la actualidad se están realizando numerosos esfuerzos para desarrollar drogas que bloqueen la transición entre los distintos estados activos y dormantes, o que sean particularmente efectivos en la eliminación de esta población de bacilos de manera tal de que sirvan como antibióticos o adyuvantes que aumenten la eficacia del tratamiento. Recientemente la Organización Mundial de la Salud publicó la guía para el uso de Bedaquiline, la primera droga con un nuevo mecanismo de acción aprobada luego de 40 años y la primera en ser utilizada específicamente para el tratamiento de la MDR-TB en combinación con otras drogas [13]. La TB podría ser controlada mediante una mejor prevención de la infección, la interrupción de su progreso a enfermedad activa y la detección y aplicación del tratamiento en forma más rápida. Para acelerar el proceso se requiere de una mayor. 7.
(22) Introducción Regulación de la síntesis de ácidos grasos en micobacterias- Mondino, S. intervención junto al desarrollo y la implementación de nuevas drogas, vacunas y herramientas de diagnótico.. 1.2 El género Mycobacterium El género Mycobacterium contiene más de 120 especies reconocidas hasta el momento, muchas de ellas descriptas en los últimos años, incluyendo principalmente organismos patógenos de humanos y animales [14]. Dentro del taxón Actinomycetes, la familia Mycobacteriaceae forma un grupo monofilético con las familias Corynebacteriaceae y Nocardiaceae, el cual contiene a todos los organismos productores de ácidos micólicos, siendo el género Mycobacterium el más extensamente estudiado por su implicancia médica [15]. Filogenéticamente, los miembros de este grupo pertenecen al dominio Bacteria, con alto contenido en G+C en su ADN, desde 53% para algunas especies de Corynebacterium hasta 70% para especies de Nocardia. Las micobacterias presentan un contenido de G+C cercano al 65%. Las micobacterias son actinomicetes aerobios, ácido-alcohol resistente como consecuencia de su compleja pared celular rica en lípidos, que forman bacilos rectos o ligeramente curvos. La separación del género Mycobacterium en dos grupos principales en función de la velocidad de crecimiento de sus especies, constituye la base de la taxonomía micobacteriana. A pesar de no respetar exactamente esta división, la mayoría de las especies de crecimiento lento (tiempo de duplicación de ∼24 h) están asociadas o son las causantes de enfermedades en humanos o animales. En consecuencia, la mayoría de las especies de crecimiento rápido (tiempo de duplicación de ∼3 h), las cuales son comúnmente saprófitas de vida libre, son consideradas no patógenas. Sin embargo, este escenario no debería conducir a una subestimación de la patogenicidad de estas micobacterias, especialmente en huéspedes inmunocomprometidos como aquellos infectados con el VIH [16]. M. tuberculosis forma un complejo junto con otras especies, caracterizado por la alta conservación de sus genomas [17]. El complejo M. tuberculosis comprende varias especies: M. tuberculosis, el agente causal de la mayor parte de los casos de TB 8.
(23) Introducción Regulación de la síntesis de ácidos grasos en micobacterias- Mondino, S. humana; M. africanum, responsable de la TB humana en el África sub-Sahariana; M. microti, el agente causal de la TB en el ratón de campo; M. bovis, capaz de infectar una amplia variedad de especies de mamíferos incluyendo humanos; y M. canettii, una variante raramente encontrada pero capaz de infectar humanos [18]. Otras especies patógenas son M. leprae, agente causal de lepra, una enfermedad granulomatosa crónica de la piel y nervios periféricos [19] y M. ulcerans, que provoca la aparición de úlceras en la piel conocidas como úlceras de Buruli [20].. 1.2.1 M. smegmatis como modelo de estudio El estudio genético de M. tuberculosis se vió retrasado durante muchos años debido a ciertas características que dificultaban el trabajo con esta bacteria, entre ellas su crecimiento lento, el requerimiento de altos niveles de bioseguridad para su manipulación, su tendencia a formar grumos durante el crecimiento en medio líquido y la falta de sistemas eficientes para la transferencia de genes. En un principio, las funciones de determinados genes de M. tuberculosis se estudiaron utilizando Escherichia coli [21-23] y Streptomyces lividans [24] porque existían sistemas de clonado de ADN recombinante eficientes para estos microorganismos. Si bien estos estudios sirvieron para definir la función de algunas proteínas, el uso de hospedadores homólogos como modelo de estudio llevaría a un entendimiento más acertado de la biología de este patógeno [25]. A diferencia de M. tuberculosis, M. smegmatis es una especie de crecimiento rápido, fácilmente cultivable in vitro, donde forma colonias visibles en un período de tres a cinco días. A pesar de haber sido inicialmente aislada de exudados de chancros luéticos en 1884, y de secreciones genitales en 1885, posteriormente no ha podido ser recuperada a partir de estas mismas muestras. Se la considera una micobacteria no patógena y dado que se ha encontrado en suelo y agua se lo considera un microorganismo ambiental. La cepa salvaje presenta una baja eficiencia de transformación con plásmidos, dificultad que fue superada con el aislamiento de una cepa mutante con alta eficiencia de transformación, denominada mc2155 [26], la cual presenta diferencias en la composición de su pared [27]. Por las características anteriormente detalladas, y la disponibilidad de su secuencia genómica, esta cepa de 9.
(24) Introducción Regulación de la síntesis de ácidos grasos en micobacterias- Mondino, S. M. smegmatis se ha convertido en un microorganismo modelo para el estudio de la función de los genes micobacterianos. Por otro lado, M. marinum es un patógeno de hospedadores poiquilotermos como sapos y peces, y está muy relacionado genéticamente a M. tuberculosis. Es una micobacteria de crecimiento rápido, fácil de manipular en el laboratorio, ya que puede transformarse con ADN plasmídico y representa un mínimo riesgo para el laboratorista. La enfermedad que causa M. marinum en sus hospedadores tiene características similares a la TB, incluyendo la etapa de latencia y la formación de granulomas, lo que lo convirtió en un organismo modelo para el estudio del proceso de infección, siendo el pez cebra el hospedador más ampliamente utilizado [28, 29].. 1.3 Envoltura celular de las micobacterias Las micobacterias son relativamente resistentes al desecamiento y al tratamiento con álcali y numerosos desinfectantes químicos, dificultando en consecuencia la prevención de la transmisión de M. tuberculosis en espacios comúnes. Esta resistencia intrínseca general, y en particular a la mayoría de los antibióticos y agentes quimioterapéuticos, se debe a la presencia de una inusual envoltura celular, rica en polisacáridos y lípidos complejos [12]. La elevada impermeabilidad resulta, por un lado debido a la ineficiencia de las porinas micobacterianas de permitir la permeación de solutos hidrofílicos y su existencia en baja concentración [30] y por otro lado, al anormal grosor de la envoltura celular y su baja fluidez, lo cual repercute directamente en la permeación de compuestos hidrofóbicos. La contribución de la envoltura celular en el proceso infectivo y la persistencia de M. tuberculosis se refleja en la capacidad de algunos complejos lipídicos asociados a la pared de actuar en la modulación de la respuesta inmune del huésped y presentar propiedades antigénicas [31-33]. A su vez, numerosas drogas utilizadas actualmente en el tratamiento contra la TB están dirigidas a componentes esenciales de la pared celular de las micobacterias, como INH, etionamida (ETH), tiolactomicina (TLM), triclosán (TRC) y EMB, los cuales afectan la biosíntesis de ácidos micólicos y de otros componentes de la pared celular. 10.
(25) Introducción Regulación de la síntesis de ácidos grasos en micobacterias- Mondino, S. [34-39], demostrando la importancia de esta estructura particular en la patogénesis y persistencia de M. tuberculosis. El contenido lipídico de la envoltura celular de las micobacterias representa hasta un 40% de su peso seco [40], siendo responsable de su crecimiento agrumado en medio líquido y de sus propiedades de tinción ácido-alcohol resistente. En la figura 2 se observa un diagrama de la envoltura celular micobacteriana, compuesta por una membrana plasmática, similar a la presente en otras bacterias, rodeada por una compleja pared rica en carbohidratos y lípidos, la cual a su vez se encuentra rodeada por una estructura denominada cápsula, rica en polisacáridos y proteínas [40].. Figura 2. Representación esquemática de la envoltura celular de M. tuberculosis. Se muestran los ácidos micólicos unidos covalentemente a la capa de arabinogalactano, a su vez unida al peptidoglicano por una unión fosfodiéster. MP: membrana plasmática; PG: peptidoglicano; AG: arabinogalactano; PIMs: fosfatidilinositolmanósidos y LAM: lipoarabinomanano, anclados en la membrana plasmática. Otros lípidos complejos libres presentes en la pared celular son acilglicolípidos, trehalosa monomicolato (TMM) y dimicolato (TDM). 11.
(26) Introducción Regulación de la síntesis de ácidos grasos en micobacterias- Mondino, S. 1.3.1 Membrana plasmática Los fosfolípidos de la membrana plasmática son derivados del ácido fosfatídico, como por ejemplo fosfatidilglicerol, difosfatidilglicerol, fosfatidiletanolamina y fosfatidilinositol y sus manósidos (PIMs). Los PIMs son exclusivos de actinomicetes, siendo componentes principales de la membrana plasmática de las micobacterias y la base lípidica del lipoarabinomanano (LAM) y lipomanano (LM), extensiones multiglicosiladas de los PIMs. Como consecuencia de la presencia de estos carbohidratos, la bicapa lipídica presenta una apariencia asimétrica por microscopía electrónica, sugiriendo que los PIMs se ubican preferentemente en la cara externa de la misma [12].. 1.3.2 Pared celular El principal componente estructural de la pared celular de las micobacterias es el complejo micolil-arabinogalactano-peptidoglicano (mAGP), una macromolécula compuesta por tres elementos unidos covalentemente: peptidoglicano (PG), arabinogalactano (AG) y ácidos micólicos, y que define la forma de las micobacterias [40]. El PG micobacteriano es similar al encontrado en la mayoría de las bacterias, presentando dos diferencias: algunos o todos los residuos de ácido murámico están Nglicosilados y los entrecruzamientos en las cadenas laterales peptídicas ocurren entre dos residuos de ácido diaminopimélico, así como también entre un residuo de éste y D-alanina [41]. Unido al PG, mediante una unión diglicosil-fosfato a la posición C-6 de algunos de los residuos de ácido murámico, se encuentra el AG. Éste es un heteropolisacárido complejo que consta de una cadena lineal formada por residuos de D-galactofuranosa, al cual se unen a su vez dos o tres cadenas laterales de arabinano, estas últimas compuestas por residuos de D-arabinofuranosil. En las dos terceras partes de los extremos no reductores disponibles de estas unidades de arabinano, se ubican los ácidos micólicos en grupos de cuatro [42]. Éstos son ácidos grasos -alquil-hidroxilados muy largos, formados por una cadena carbonada principal, conocida como ácido meromicólico, y una cadena lateral . Las micobacterias presentan los 12.
(27) Introducción Regulación de la síntesis de ácidos grasos en micobacterias- Mondino, S. ácidos micólicos (C70-C90) y las cadenas laterales más largas (C22-C26) que se conocen en la naturaleza. Cada especie tiene una composición característica y esta distinción se ha empleado para la identificación y clasificación de las distintas especies de Mycobacterium [43]. En la figura 3 se muestra un esquema de los ácidos micólicos encontrados en M. tuberculosis y M. smegmatis. Los ácidos micólicos son responsables de algunas de las características distintivas de las micobacterias, como la resistencia a daños químicos, resistencia a deshidratratación, baja permeabilidad a antibióticos hidrofóbicos, virulencia [44-47], tinción ácido-alcohol resistente [48] y la habilidad de persistir en el hospedador [31, 47, 49]. Minnikin fue el primero en proponer un modelo para la organización del complejo mAGP y lípidos asociados a la pared celular. En éste, las cadenas de ácido micólico se disponen de forma paralela entre sí y perpendicularmente al plano de la superficie celular, dando lugar a la cara interna de una bicapa lipídica asimétrica [50]. Esta cara interna, a su vez está cubierta por una cara externa compuesta por lípidos complejos libres.. 1.3.3 Lípidos libres Existe una gran variedad de lípidos de estructuras inusuales en forma de lípidos libres; esto es, extraíbles con solventes orgánicos, que no forman parte del complejo mAGP pero que están asociados a éste. Las principales clases incluyen lipooligosacáridos (LOS), glicopeptidolípidos (GPL), glicolípidos fenólicos (PGL), ceras como el tiocerol dimicocerosato (PDIM), trehalosas aciladas como diacil- y pentaciltrehalosas (DAT y PAT) y la trehalosa dimicolato (TDM), y sulfolípidos (SL) (Fig. 4). Cada especie micobacteriana presenta clases y variantes características [12]. En M. tuberculosis los lípidos mejor caracterizados incluyen los PDIM, SL y trehalosas aciladas, que cumplen funciones en eventos de señalización, en la patogénesis y en la modulación de la respuesta inmune [51]. La TDM representa el lípido extraíble más abundante, granulogénico y significativamente tóxico de la envoltura celular de M. tuberculosis [52]. 13.
(28) Introducción Regulación de la síntesis de ácidos grasos en micobacterias- Mondino, S. Figura 3. Estructura de los principales ácidos micólicos identificados en M. tuberculosis y M. smegmatis. Los ácidos micólicos son ácidos grasos α–alquil β–hidroxilados formados por una cadena larga carbonada (ácido meromicólico) y una cadena lateral más corta. Adaptado de Brown y colaboradores [53].. 1.4 Biosíntesis de ácidos grasos en bacterias La biosíntesis de ácidos grasos es el primer paso en la formación de lípidos de membrana, y representa un aspecto esencial del metabolismo en todos los organismos. A pesar de existir una variación considerable en las estructuras moleculares de las distintas sintasas de ácidos grasos (FAS), el mecanismo de reacción de la síntesis de novo de ácidos grasos es esencialmente el mismo en todos los sistemas biológicos; y en bacterias, las enzimas involucradas y los pasos metabólicos han sido caracterizados utilizando E. coli como modelo de estudio.. 14.
(29) Introducción Regulación de la síntesis de ácidos grasos en micobacterias- Mondino, S. Figura 4. Lípidos complejos extraíbles de Mycobacterium. A. Estructuras genéricas de las tres clases principales de glicolípidos extraíbles de la pared celular micobacteriana. R, cadena larga de acil-ácido graso. B. Ejemplos de algunos lípidos extraíbles de M. tuberculosis.. El primer paso de esta ruta biosintética consiste en la carboxilación de acetilCoA mediada por una acetil-CoA carboxilasa (ACC), para dar malonil-CoA. El residuo malonato luego es transferido a la proteína transportadora de acilos (ACP) por la malonil-CoA:ACP transacilasa (FabD) para dar lugar a malonil-ACP. El primer paso del ciclo de elongación consiste en la condensación de malonil-ACP con acetil-CoA, 15.
(30) Introducción Regulación de la síntesis de ácidos grasos en micobacterias- Mondino, S. catalizada por la β-cetoacil-ACP sintasa III (FabH). Los pasos posteriores de elongación, que determinan la adición sucesiva de dos unidades de carbono hasta sintetizar un ácido graso C16-18, son catalizados por una β-cetoacil-ACP sintasa (FabB-FabF). El βcetoéster resultante, es reducido a un β-hidroxiacil-ACP por una β-cetoacil-ACP reductasa (FabG), seguida de la eliminación de una molécula de agua mediada por una β-hidroxiacil-ACP deshidratasa (FabA-FabZ). La reducción final del intermediario trans2-enoil-ACP es catalizada por una enoil reductasa (FabI-FabK-FabL), dando como producto acil-ACPs saturados, que serán los sustratos de una nueva ronda de elongación [54, 55] (Fig. 5).. Figura 5. Biosíntesis de ácidos grasos en bacterias. ACC: acetil-CoA carboxilasa; FabD: acil-CoA:ACP transacilasa; FabH: β-cetoacil-ACP sintasa III; FabG: β-cetoacil-ACP reductasa; FabA/FabZ: β-hidroxiacilACP deshidratasa/isomerasa y β-hidroxiacil-ACP deshidratasa; FabI, FabK, FabL: enoil-ACP reductasas; FabB, FabF: β-cetoacil-ACP sintasas. Adaptado de Marrakchi y colaboradores [56].. Las FAS son clasificadas en base a la estructura proteica como de tipo I o tipo II. Las FAS de tipo I consisten en una proteína multifuncional, que contiene en una única cadena polipeptídica las distintas actividades catalíticas de la secuencia de reacción, como dominios funcionales discretos. Estas enzimas se encuentran típicamente en el citoplasma de organismos eucariotas y excepcionalmente en el subgrupo productor de 16.
(31) Introducción Regulación de la síntesis de ácidos grasos en micobacterias- Mondino, S. ácidos micólicos de los actinomicetes [57]. Las FAS de tipo II constituyen un sistema disociado formado por proteínas independientes altamente conservadas, codificadas por una serie de genes individuales. Este sistema se encuentra presente en bacterias así como también en organelas de ascendencia procariota (cloroplastos y mitocondrias), donde catalizan la síntesis de ácidos grasos de novo [58]. Los únicos organismos procariotas que utilizan el sistema FAS de tipo I y no de tipo II para la síntesis de ácidos grasos de novo son aquellos productores de ácidos micólicos de la rama de los Actinomycetales, dentro de los cuales se encuentran las micobacterias [5961].. 1.5 Aspectos generales de la biosíntesis de ácidos grasos y ácidos micólicos en las micobacterias 1.5.1 Características de los sistemas FAS-I y FAS-II micobacterianos Las micobacterias son procariotas inusuales dado que poseen los dos tipos de sistemas FAS: FAS-I y FAS-II, siendo ambos necesarios para la biosíntesis de ácidos micólicos [62]. La multienzima bacteriana FAS-I es un hexámero de subunidades idénticas, que cataliza la síntesis de novo de acil-CoAs de cadena larga, los cuales son utilizados para la síntesis de ácidos micólicos, fosfolípidos de membrana y en micobacterias patógenas, son sustratos para la síntesis de ácidos grasos multimetil ramificados (PDIM, TDM entre otros) [57]. Si bien la organización estructural de esta proteína micobacteriana en principio no presenta diferencias con la observada para otros sistemas FAS-I, ciertas propiedades parecerían ser únicas. Entre las más importantes se incluyen la síntesis de producto siguiendo un patrón bimodal, la estimulación debido a ciertos polisacáridos micobacterianos y su capacidad de funcionar como elongasa, catalizando la elongación de palmitoil-CoA [63, 64]. Por otro lado, a diferencia de lo que ocurre en levaduras, la actividad 4’-fosfopanteteinil transferasa encargada de activar el dominio ACP del sistema FAS-I se encuentra codificada por un gen separado (acpS) [65]. A su vez, la multienzima bacteriana se diferencia de otras conocidas en utilizar diferentes reductores, NADPH y NADH, para sus pasos de cetoacil- y enoil-reducción, respectivamente [63].. 17.
(32) Introducción Regulación de la síntesis de ácidos grasos en micobacterias- Mondino, S. Experimentos bioquímicos han demostrado que la síntesis mediada por FAS-I tiene un comportamiento bimodal in vitro, produciendo dos clases de acil-CoAs de cadena larga: C16-18-CoAs y C24-26-CoAs [59, 60, 63]. En presencia de - acilpolisacáridos micobacterianos estas enzimas aumentarían la producción de C16-18-CoA, y la velocidad de síntesis de ácidos grasos en general. Esto se debería a la capacidad de los azúcares de formar un complejo selectivo con estos ácidos grasos, resultando en una regulación por remoción del producto final. Por otro lado, en ausencia de estos polisacáridos se produciría como forma predominante C26-CoA en M. tuberculosis y C24-CoA en M. smegmatis, liberándose el producto final de la enzima por formación de autoagregados [63, 66]. Esta bimodalidad de FAS-I no sería solamente una propiedad intrínseca de la proteína, sino que estaría determinada también por factores externos que influirían en el proceso de elongación. Esto fue demostrado in vivo cuando, al intercambiar la enzima FAS-I de M. smegmatis por la de M. tuberculosis, se encontró que ésta produce principalmente C24-S-enzima y no C26-S enzima, como se esperaba. Esto sugeriría que existen complejas interacciones entre FAS-I y FAS-II, y posiblemente otros elementos aún no dilucidados [67]. Por otro lado, si bien se ha sugerido que el sistema FAS-I in vitro funcionaría como de novo sintasa y elongasa; en las micobacterias patógenas, las cuales posiblemente deriven la mayoría de sus ácidos grasos del huésped, el mismo actuaría principalmente como elongasa convirtiendo los ácidos grasos del huésped en productos C24-26 [64]. Las micobacterias contienen también un sistema disociado FAS-II, dependiente de ACP [68]. Éste se diferencia únicamente de las sintasas de tipo II encontradas en otras bacterias, en su incapacidad de realizar la síntesis de novo de ácidos grasos. En cambio, elonga acil-CoAs de cadena media (C12-16) hasta obtener cadenas aciladas muy largas (C50-60) que dan lugar a la cadena meromicolato del ácido micólico.. 1.5.2 Biosíntesis de ácidos grasos y ácidos micólicos en micobacterias La biosíntesis de ácidos grasos/micólicos ha sido ampliamente estudiada en los últimos años, dada la relevancia de los lípidos en la patogénesis de M. tuberculosis (Fig. 6). FAS-I cataliza la síntesis de novo de acil-CoAs de cadena larga a partir de acetil-CoA. 18.
(33) Introducción Regulación de la síntesis de ácidos grasos en micobacterias- Mondino, S. y malonil-CoA para dar butiril-S-enzima. Las elongaciones subsiguientes mediadas por FAS-I producen C16- y C18-S-enzima, los cuales son convertidos a sus derivados acil-CoA y utilizados principalmente para la síntesis de fosfolípidos de la membrana plasmática. Por otro lado, FAS-I es capaz de continuar con la elongación del palmitoil-CoA para dar lugar en M. tuberculosis, por ejemplo, a C26-S-enzima que sería liberado como C26-CoA, y posteriormente activado mediante una carboxilasa específica para su condensación con el ácido meromicólico, mediada por la policétido sintasa Pks13, convirtiéndose así en la cadena α-alquilo del ácido micólico [62, 69]. Los sistemas FAS del tipo I y II están conectados por una enzima condensante clave, la β-cetoacil-ACP sintasa III o mtFabH. Esta enzima cataliza la condensación decarboxilativa de malonil-ACP con los acil-CoAs producidos por FAS-I [70]. El producto de esta condensación, un β-cetoacil-ACP de cadena mediana, es el sustrato inicial del sistema FAS-II. Estudios bioquímicos llevados a cabo en presencia de malonil-ACP muestran que mtFabH puede utilizar un rango de C 12-20-CoAs como sustrato, lo cual resulta consistente con los estudios estructurales realizados por Scarsdale y colaboradores [71], los cuales diferencian a esta enzima de las FabH de otros sistemas FAS-II, que típicamente usan como sustrato C2-6-CoAs [71, 72]. El mecanismo general de la síntesis de ácidos grasos es común a todas las bacterias, y los genes que codifican para las enzimas involucradas en cada uno de los pasos están generalmente conservados (Tabla 1). En las micobacterias, los genes que codifican para las enzimas del sistema FAS-II se ubican en diferentes regiones del cromosoma, organizados en tres unidades independientes. La primera está formada por los genes fabD-acpM-kasA-kasB-accD6; la segunda tiene dos genes, mabA-inhA; y la tercera cuenta con tres genes, hadA, hadB, hadC [65, 73] (Tabla 2). Pawelczyk y colaboradores [74] demostraron recientemente que los genes fabD, acpM, kasA, kasB y accD6 forman un operón, al que denominamos operón fasII. Su experimento consistió en la amplificación por PCR de las regiones intergénicas ubicadas entre cada uno de estos genes, utilizando como molde ADNc de M. tuberculosis y M. smegmatis. Mediante la obtención de los productos de PCR esperados confirmaron que efectivamente se trataba de una única unidad transcripcional [74].. 19.
(34) Introducción Regulación de la síntesis de ácidos grasos en micobacterias- Mondino, S. Figura 6. Representación esquemática de la vía de biosíntesis de los ácidos micólicos. La biosíntesis de los precursores de los ácidos micólicos requiere de la actividad coordinada de dos ácido graso sintasas: FAS-I y FAS-II. Los pasos de la síntesis de ácidos micólicos se encuentran detallados en el texto. Los componentes del sistema FAS-II están codificados en tres operones. Se resaltan en negrita las proteínas codificadas por los genes del operón fasII (FabD: malonil-ACP transacilasa, AcpM: proteína transportadora de acilos, KasA y KasB: -cetoacil-ACP sintasas y ACCasa: acil-CoA carboxilasa).. El primer gen del operón fasII codifica para la malonil-CoA:ACP transacilasa FabD [75] y el segundo codifica para la ACP micobacteriana (AcpM), involucrada en el transporte de la cadena acilo creciente de un sitio activo a otro dentro del sistema FASII durante la síntesis del ácido graso. Esta proteína es convertida a su forma activa holo-AcpM por la holo-ACP sintasa (AcpS), la cual cataliza la transferencia de un grupo 4-fosfopanteteína de la coenzima A (CoA) a la apo-ACP [76]. AcpM es una ACP inusual 20.
(35) Introducción Regulación de la síntesis de ácidos grasos en micobacterias- Mondino, S. en lo que respecta a su tamaño en comparación con la proteína ACP de E. coli; consiste en un polipéptido de mayor longitud el cual se diferencia de las ACPs bacterianas por presentar una extensión en su extremo carboxilo terminal. Mediante el secuestro de la cadena acilada hidrofóbica del solvente, esta característica adicional podría tener un rol importante en la transferencia del ácido meromicólico en crecimiento, a los diferentes centros del sistema enzimático FAS-II [75, 77].. Tabla 1. Enzimas del sistema FAS-II en diferentes géneros bacterianos Grupo de enzimas. Organismo E. coli. Enzimas condensantes. Enoil-ACP reductasas. M. tuberculosis E. coli S. pneumoniae Bacillus subtilis M. tuberculosis E. coli. Deshidratasas. ACPs β-cetoacilACP reductasas Transacilasas. M. tuberculosis E. coli M. tuberculosis E. coli M. tuberculosis E. coli M. tuberculosis. Proteína. Actividad. FabB FabF FabH mtFabH KasA KasB FabI. β-cetoacil-ACP sintasa I β-cetoacil-ACP sintasa II β-cetoacil-ACP sintasa III β-cetoacil-ACP sintasa III β-cetoacil-ACP sintasa β-cetoacil-ACP sintasa Enoil-ACP reductasa I. FabK. Enoil-ACP reductasa II. FabL. Enoil-ACP reductasa III. InhA. Enoil-ACP reductasa. FabA. β–hidroxiacil-ACP deshidratasa/isomerasa β–hidroxiacil-ACP deshidratasa. FabZ HadAB HadBC AcpP. Proteína transportadora de acilos. AcpM. Proteína transportadora de acilos. FabG. β–cetoacil-ACP reductasa. MabA. β–cetoacil-ACP reductasa. FabD. Malonil-CoA:ACP transacilasa. FabD. Malonil-CoA:ACP transacilasa. β–hidroxiacil-ACP deshidratasas. 21.
(36) Introducción Regulación de la síntesis de ácidos grasos en micobacterias- Mondino, S. Tabla 2. Genes y enzimas del sistema FAS-II en micobacterias Operón putativo. fabD-acpM-kasA-kasB-accD6. Gen. Actividad enzimática. fabD acpM kasA kasB. Malonil-CoA:ACP transacilasa Proteína transportadora de acilos β-cetoacil-ACP sintasa β-cetoacil-ACP sintasa Subunidad carboxil transferasa de la ACCasa 6 β-hidroxiacil-ACP reductasa Enoil-ACP reductasa Hidroxil-acil-deshidratasa Hidroxil-acil-deshidratasa. accD6 mabA-inhA hadA-hadB-hadC. mabA inhA hadA, hadB hadB, hadC. Los pasos de elongación del sistema FAS-II que siguen a continuación de la reacción de condensación dependiente de FabH son llevados a cabo por las β-cetoacilACP sintasas codificadas por los genes kasA y kasB, del operón fasII. Las proteínas KasA y KasB de M. tuberculosis tienen un 67% de identidad y ambas enzimas catalizan la condensación de acil-AcpM con malonil-AcpM, elongando la cadena del meromicolato en dos atomos de carbono [78]. Las dos enzimas tienen distinta especificidad de sustrato, esto sugiere que cumplen roles diferentes en el proceso de elongación. En base a estudios in vitro se propuso que KasA estaría implicada en la elongación inicial de los micolatos, que serían luego extendidos por KasB hasta su longitud final [79, 80]. Las estructuras cristalográficas de estas enzimas son consistentes con la especificidad de sustrato propuesta [78, 81]. Esta hipótesis a su vez fue confirmada in vivo analizando cepas mutantes en el gen kasB de M. marinum y M. tuberculosis. En estas, los ácidos micólicos presentan pequeños cambios en su longitud y composición, lo que determina no sólo una marcada alteración en la permeabilidad de la envoltura celular de estas micobacterias sino también la pérdida de su característica de tinción ácidoalcohol resistente. Por otro lado, la cepa de M. tuberculosis mutante en kasB es incapaz de provocar una enfermedad activa en ratones, pudiendo persistir en estos durante 600 días. Cabe destacar que ninguna de estas anormalidades pudo ser complementada por la sobre-expresión de kasA; demostrando que a pesar de ser altamente homólogas, ambas enzimas no presentan funciones redundantes [48, 82]. Por otro lado, a diferencia de KasB, KasA es un componente esencial del sistema FAS-II 22.
(37) Introducción Regulación de la síntesis de ácidos grasos en micobacterias- Mondino, S. cuya ausencia lleva a la lisis celular. Para caracterizar esta proteína, se construyó una cepa de M. smegmatis mutante condicional en kasA la cual presentó una disminución en los niveles de α- y epoxi-micolatos, y una acumulación de α’-micolatos. Nuevamente, múltiples copias de KasB no pudieron complementar la disminución en los niveles de KasA en esta cepa, confirmando el rol diferencial que presentan durante la vía de síntesis de la cadena de meromicolato [83]. El último gen de este operón es accD6, el cual codifica para la subunidad β de la acil-CoA carboxilasa 6 (ACCasa6), responsable de sintetizar malonil-CoA, el sustrato utilizado por ambos sistemas FAS durante los distintos pasos de elongación [84]. Si bien se ha demostrado que este gen es esencial en M. tuberculosis [74], existen controversias respecto de su esencialidad en M. smegmatis [74, 84]. Las reductasas del sistema FAS-II están codificadas por los genes mabA e inhA, ubicados en la misma región cromosómica de M. tuberculosis formando un operón en esta especie [34]. MabA codifica para una β-cetoacil-ACP reductasa que a diferencia de otras cetoacil reductasas, presenta preferencia por sustratos de cadena larga consistente con su rol en la biosíntesis de ácidos micólicos [85]. El gen inhA codifica para la proteína que cataliza la trans-2-enoil-ACP reducción, con preferencia por sustratos largos [86] y fue identificado como blanco de acción de INH y ETH en M. tuberculosis [34, 87]. Por otro lado, el paso de deshidratación del intermediario β-hidroxiacil-ACP a trans-2-enoil-ACP está catalizado por las β-hidroxiacil-ACP deshidratasas del operón hadA-hadB-hadC de M. tuberculosis, siendo al menos uno de estos genes esenciales para la viabilidad bacteriana. Las proteínas codificadas por estos genes forman dos heterodímeros, HadAB y HadBC. Al igual de lo que ocurre para KasA, HadAB estaría involucrado en los ciclos iniciales de la vía de elongación, mientras que HabBC, al igual que KasB, estaría implicado en los últimos pasos de la biosíntesis del ácido meromicólico [73]. La última etapa en la vía de síntesis de los ácidos micólicos corresponde a la activación y condensación de las cadenas aciladas provenientes de los sistemas FAS-I y FAS-II. En primer lugar, la cadena del ácido meromicólico es activada a acil-AMP a través de la acción de una acil-AMP ligasa (FadD32) [88, 89]. A su vez, el C24-26-CoA 23.
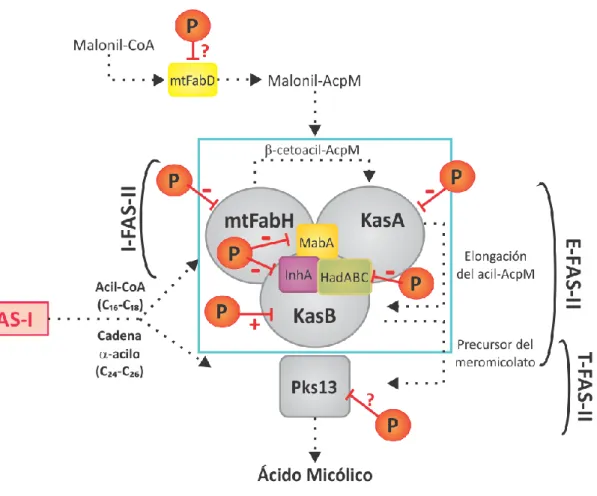
(38) Introducción Regulación de la síntesis de ácidos grasos en micobacterias- Mondino, S. proveniente del sistema FAS-I, es carboxilado por una acil-CoA carboxilasa que involucra a la subunidad carboxil transferasa AccD4 [89, 90]. Finalmente, ambos sustratos activados son condensados por la enzima Pks13 dando lugar al precursor del ácido micólico [91]. Se ha demostrado que los genes que codifican para estas tres proteínas forman el operón fadD32-pks13-accD4 y son esenciales para la viabilidad de las micobacterias [69, 89]. Las. proteínas. del. sistema. FAS-II. de. M.. tuberculosis. interaccionan. específicamente dando lugar a complejos especializados interconectados, sugiriendo que la organización de este sistema se asemejaría a la del sistema FAS-I de mamíferos (Fig. 7). En este modelo, denominado Interactoma de la biosíntesis de ácidos micólicos, un complejo de iniciación (I-FAS-II) formado por las reductasa MabA e InhA, FabD y mtFabH permitiría el pasaje de sustratos desde FAS-I a FAS-II. Luego, el complejo de elongación de tipo I (E1-FAS-II) formado por MabA, InhA, FabD, HadAB y KasA estaría involucrado en los ciclos iniciales de elongación y las cadenas acilo de longitud intermedia sintetizadas servirían entonces como sustratos para el complejo de elongación de tipo II (E2-FAS II), en el cual la deshidratasa sería el complejo HadBC y la enzima condensante sería KasB. Tras los sucesivos pasos de elongación, reducción y deshidratación llevados adelante por este último complejo, los meromicolil-AcpM de longitud completa resultantes son condensados con la cadena acilada -, por el complejo de terminación (T-FAS-II) que involucra a la proteína Pks13, la cual se encontraría interaccionando preferentemente con KasB [92-94]. Las cadenas meromicólicas son modificadas en dos posiciones específicas: distal y proximal (Fig. 3), habiéndose identificado hasta el momento ocho metiltransferasas involucradas en estas modificaciones específicas (MmaA1-MmaA4, CmaA1, CmaA2, PcaA y UmaA). Se ha demostrado que estas proteínas interaccionarían con las enzimas condensantes, siendo específicas de los complejos E-FAS-II, modificando de esta manera la cadena meromicólica durante el proceso de elongación [94]. En consecuencia, la especificidad de cada tipo de complejo de elongación y de las modificaciones realizadas en la cadena del meromicolato, estaría dada por las interacciones específicas ocurridas entre las enzimas condensantes, los heterodímeros de las deshidratasas y las metiltransferasas.. 24.
(39) Introducción Regulación de la síntesis de ácidos grasos en micobacterias- Mondino, S. Figura 7. Esquema de las interacciones proteína-proteína dentro del Interactoma de la biosíntesis de ácidos micólicos. I-FAS-II, complejo de iniciación; E1-FAS-II complejo de elongación I; E2-FAS-II complejo de elongación II; T-FAS II complejo de terminación. Las enzimas condensantes interaccionan entre ellas y con el bloque compuesto por MabA, InhA y FabD. KasA y KasB interaccionan con HadAB y HadBC respectivamente. Las interacciones con las metiltransferasas (indicadas dentro de los recuadros violetas) están representadas con flechas negras. Cada complejo está representado por un recuadro gris. Adaptado de Cantaloube y colaboradores [94].. 1.6 Antecedentes sobre la regulación de la biosíntesis de ácidos grasos en bacterias Los ácidos grasos son los componentes de las membranas biológicas energéticamente más costosos de producir. En consecuencia, las bacterias han desarrollado mecanismos sofisticados que les permiten controlar con precisión la expresión de los genes involucrados tanto en la biosíntesis como en la degradación de los mismos, así como también en la modificación de las cadenas aciladas existentes. De 25.
Figure




+7





Outline
Biosíntesis de ácidos grasos y ácidos micólicos en micobacterias
Construcción de plásmidos
Manipulación de ARN
Identificación de un factor de transcripción de la familia TetR que se une a la región promotora del operón fas-acpS en Mycobacterium
FasR es un regulador altamente conservado en actinomicetales productores de ácidos micólicos
Los acil-CoAs de cadena larga modulan la unión de FasRMT a la región promotora del operón fas-acpS
Construcción de una cepa de M smegmatis mutante en fasR, utilizando el plásmido termosensible pPR
FasR es un activador del operón fas-acpS in vivo
Caracterización de la cepa MSPptr:fasMS
Caracterización de la cepa Mtb fasR pFR
Documento similar
Para recibir todos los números de referencia en un solo correo electrónico, es necesario que las solicitudes estén cumplimentadas y sean todos los datos válidos, incluido el
Abstract: This paper reviews the dialogue and controversies between the paratexts of a corpus of collections of short novels –and romances– publi- shed from 1624 to 1637:
En junio de 1980, el Departamento de Literatura Española de la Universi- dad de Sevilla, tras consultar con diversos estudiosos del poeta, decidió propo- ner al Claustro de la
Sanz (Universidad Carlos III-IUNE): "El papel de las fuentes de datos en los ranking nacionales de universidades".. Reuniones científicas 75 Los días 12 y 13 de noviembre
(Banco de España) Mancebo, Pascual (U. de Alicante) Marco, Mariluz (U. de València) Marhuenda, Francisco (U. de Alicante) Marhuenda, Joaquín (U. de Alicante) Marquerie,
Missing estimates for total domestic participant spend were estimated using a similar approach of that used to calculate missing international estimates, with average shares applied
Habiendo organizado un movimiento revolucionario en Valencia a principios de 1929 y persistido en las reuniones conspirativo-constitucionalistas desde entonces —cierto que a aquellas
The part I assessment is coordinated involving all MSCs and led by the RMS who prepares a draft assessment report, sends the request for information (RFI) with considerations,
