María Elena Báez Flores por estar siempre pendiente de la realización de este proyecto y de la culminación de mis estudios de maestría a pesar de la distancia. La expresión SETIS de DNAMET se analizó mediante la reacción en cadena de la polimerasa con transcriptasa inversa cuantitativa (qTR-PCR).
Introducción
Su utilidad ha sido demostrada en la criopreservación de especies en peligro de extinción (Sen-Rong y Ming-Hua, 2009) y especies de importancia agrícola (Panis et al., 2005). La metilación de citosina es una de las modificaciones epigenéticas en plantas más estudiadas (Saze et al., 2012).
Antecedentes
El Cultivo de Vid en Sonora
Conservación de Germoplasma
- Conservación in situ
- Conservación ex situ
- Bancos de semillas
- Cultivo in vitro
- Crioconservación
10 El deterioro fisiológico es otro problema de la conservación de germoplasma en los bancos de semillas. Debido a los problemas antes mencionados, una de las alternativas a los bancos de semillas para la conservación de especies es el cultivo in vitro.
Crioconservación por Vitrificación
12 Primero se forman las sales extracelulares, lo que aumenta la concentración de sales extracelulares. En el Centro Internacional de la Papa (CIP) en Lima, Perú, se conservan 345 accesiones mediante el protocolo de vitrificación.
Conservación de Germoplasma de Vid
- Bancos de Germoplasma de Vid a Nivel Mundial y Nacional
- Crioconservación por Vitrificación en Vitis vinifera
Hace más de dos décadas, la Unión Internacional para la Conservación de la Naturaleza (UICN) la registró en la lista europea de plantas en peligro de extinción (Imazio et al., 2007). 16 El Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP) cuenta con un banco nacional de germoplasma de vid, ubicado en la región de La Laguna, Coahuila.
Modificaciones Moleculares Producidas por la Crioconservación y la
- Modificaciones Genéticas
- Alteraciones Epigenéticas
Esto puede provocar un cambio en la programación normal de la información genética y, a su vez, provocar cambios fenotípicos en los organismos recién formados (Phillips et al., 1994). Existen estudios que han demostrado la presencia de estos cambios por el efecto de criopreservación de la vitrificación en las plantas. En consecuencia, no se observaron bandas polimórficas debido a la criopreservación y los niveles de ploidía se mantuvieron en ambos genotipos analizados a pesar de las diferencias en la capacidad de regeneración.
La remodelación de la cromatina implica la sustitución de histonas en el nucleosoma por otras histonas con propiedades diferentes (Iwatani et al., 2006) o su modificación enzimática. Algunos estudios han demostrado un vínculo entre la modificación de histonas y la metilación del ADN (Saze et al., 2012); experimentos. Los estudios centrados en la evaluación de la estabilidad epigenética en plantas cultivadas in vitro muestran una alta frecuencia de variación en la metilación del ADN.
Aunque existen muchos estudios sobre modificaciones en la metilación del ADN en respuesta al cultivo in vitro, en el caso de la criopreservación son escasos.
Cambios en la Metilación de ADN de Plantas Debidos a la
Lo anterior sugiere que el estrés provoca cambios en la expresión de estos genes, lo que probablemente provoque cambios en el patrón de metilación. Asimismo, se evaluó el efecto de la aplicación de las técnicas antes mencionadas sobre la viabilidad y capacidad de desarrollo de los embriones. La criopreservación mediante vitrificación induce cambios en la expresión de genes que codifican las ADN metiltransferasas (ADNMET) en embriones cigóticos de uva (Vitis vinifera).
4.- Analizar los cambios en la expresión de genes aislados que codifican las enzimas de ADNMET en embriones cigóticos de uva criopreservados por vitrificación. 58 respecto a los criopreservados durante media hora (Mh-V7) y dos horas (2H-V7); Es decir, no parece haber ningún efecto del tiempo de exposición a NL de media hora (Mh-V7) a dos horas (2H-V7) sobre la expresión de MET1-2. La disminución de la expresión de MET1-2 observada en embriones cigóticos puede deberse al estrés provocado por los tratamientos de vitrificación y criopreservación aplicados.
Los resultados de este estudio muestran que disminuyó la expresión de MET1-2 en embriones de V. En este estudio no se observaron cambios en la metilación del promotor para ninguno de los genes evaluados; ni en la expresión de los genes Hat1 y Hdac1 por efecto de la vitrificación. Actualmente, los informes de la literatura indican que la expresión genética y la metilación del ADN cambian dinámicamente en respuesta a factores ambientales.
Los autores sugieren que los genes sobreexpresados pueden estar relacionados con una viabilidad reducida del embrión debido a la vitrificación. Lo anterior indica que la disminución de la expresión de MET1-2 y DRM1 observada en el presente estudio puede deberse a una disminución en la división y diferenciación celular en respuesta a la lesión por criopreservación inducida por la vitrificación.

V Participación de las ADN Metiltransferasas en las Modificaciones
Hipótesis
Objetivos
Objetivo General
Objetivos Específicos
Materiales y Métodos
- Establecimiento del Protocolo de Crioconservación por Vitrificación
- Material Vegetal
- Vitrificación y Crioconservación
- Análisis de Viabilidad
- Regeneración de Plántulas
- Aislamiento de Fragmentos de Genes Codificando Enzimas ADNMET
- Diseño de Oligonucleótidos
- Extracción de ARN Total y Síntesis de ADNc
- Amplificación, Secuenciación y Análisis a Nivel de Aminoácidos
- Análisis de Expresión de Genes Codificando Para Enzimas ADNMET
- Extracción de ARN Total y Diseño de Oligonucleótidos…
- Cuantificación de Expresión de Transcritos Codificando ADNMET
- Análisis Estadístico
DRM1F y R: Oligonucleótidos para amplificar el gen de los dominios modificados de 5-citosina ADN metiltransferasa tipo 1. DRM2F y R: Oligonucleótidos para amplificar el gen de los dominios modificados de 5-citosina ADN metiltransferasa tipo 2. CMT3F y R: Oligonucleótidos para amplificar el gen de 5-citosina ADN metiltransferasa modificadora de la cromatina tipo 3.
Posteriormente, se realizó la estandarización de la retro-transcripción del ARN amplificando el ARNm a y 26 ciclos para identificar el ciclo de amplificación óptimo y evitar la sobreamplificación, tal y como indica el 'SMARTer PCR cDNA SYntesis kit' (Clontech Laboratories USA, CA ). A partir del ADNc y utilizando los oligonucleótidos previamente diseñados, se realizó la estandarización de la amplificación mediante PCR en gradiente de temperatura; Para la cuantificación de la expresión del gen constitutivo se utilizaron los oligonucleótidos VvGAPDH-F y VvGAPDH-R, propuestos por Reid y colaboradores (2006), que se enumeran en la Tabla 3.
Oligonucleótidos diseñados para evaluar la expresión de genes parciales que codifican ADNMET y del gen endógeno utilizado para la normalización.
Resultados y Discusión
Efecto de la Vitrificación y Crioconservación Sobre la Viabilidad
La vitrificación en dos pasos aplicada en el presente estudio (60 min. en 50% PVS2 y 120 min. en 100% PVS) no aumentó la viabilidad de los embriones en comparación con el tratamiento en un solo paso con el mismo tiempo de vitrificación (180 min. .). en PVS2 al 100%). Los resultados sugieren que la disminución en la viabilidad observada en los tratamientos V1 y V2 se debió al tiempo prolongado de exposición a PVS2 y no a la exposición del tejido a la solución (Figura 1). Al comparar la viabilidad de embriones vitrificados (V4) sin criopreservación con tratamientos de criopreservación, se encontró que la inmersión directa en nitrógeno líquido no afecta la viabilidad de los embriones (p>0,05).
Sin embargo, en este trabajo se observó una reducción del 33% en la viabilidad de los embriones V. Sin embargo, al observar los resultados de reducción del TTC en el presente estudio (Figura 2), se observa claramente que el aumento en el tiempo de la criopreservación disminuye la viabilidad de los embriones. 46 tiempos de criopreservación en el presente estudio, pueden deberse a la disminución de los factores mencionados y no a una disminución de la viabilidad per se.
Kartha (1985) sostiene que la evaluación de la viabilidad embrionaria mediante la reducción del TTC por sí sola es insuficiente.

Efecto de la Vitrificación y Crioconservación Sobre el Desarrollo del
Al evaluar el efecto de la vitrificación en el desarrollo embrionario, el tratamiento V7 mostró un mayor porcentaje de recuperación que el V5 (p<0,05). Aunque el V7 reduce en menor medida el poder de regeneración de los embriones cigóticos, V. Kartha (1985) afirma que la vitrificación en sí reduce la viabilidad del material vegetal, pero permite su conservación a temperaturas ultrabajas, lo que permite la regeneración tras la descongelación.
Los porcentajes de regeneración obtenidos en este estudio son similares a los reportados para vitrificación y criopreservación de otros tejidos de la misma especie. No hubo diferencia (p>0,05) en el porcentaje de plántulas recuperadas entre el tratamiento con Mh-V7 y los embriones criopreservados sin vitrificación (T-). Aunque se puede prescindir de la vitrificación para obtener plantas completas tras la criopreservación de tejidos, es necesario aumentar el número de plantas que pueden desarrollarse a partir de embriones.
En relación con esto, Hassan y Haggag (2013) observaron que los meristemas no criopreservados que han sido vitrificados tienen una tasa de regeneración más baja que aquellos que no han sufrido ningún tratamiento, lo que indica un efecto deletéreo de la solución de vitrificación.

Transcritos Codificando ADNMET
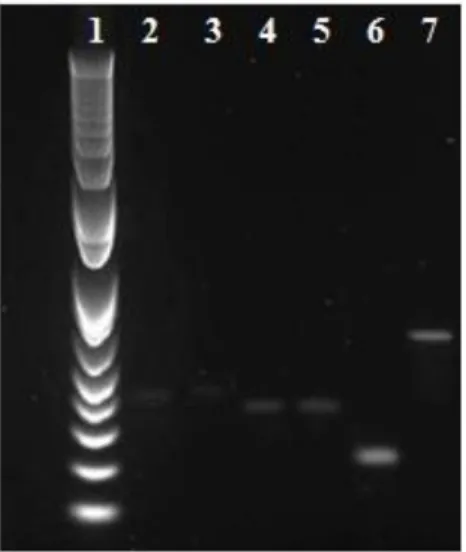
53 Las bandas de ARNr 28S y 18S que se muestran en el gel muestran un ARN libre de proteínas y no degradado (Fleige y Pfaffl, 2006). A partir de 80 ng de ARN total, la estandarización de la síntesis de ADNc se realizó mediante amplificación por PCR junto con transcripción inversa y 26 ciclos. Se obtuvieron amplicones específicos sin la formación de dímeros de oligonucleótidos a partir del ADNc sintetizado y utilizando los oligonucleótidos previamente diseñados.
El tamaño de los amplicones corresponde a los tamaños teóricos esperados; excepto el amplificado por el par de oligonucleótidos CMT3F y R, que aparentemente era de mayor tamaño. Las condiciones óptimas de amplificación establecidas para cada uno de los productos obtenidos se muestran en la Tabla 5. Como resultado de la secuenciación se obtuvieron 5 secuencias de expresión marcadas de genes específicos (SETIS), cuya longitud correspondía al tamaño teórico esperado.
Resultados del análisis de la secuencia de aminoácidos deducida de la secuencia de nucleótidos del SETIS obtenido.

Efecto de la Vitrificación y Crioconservación Sobre la Expresión de
Los resultados del análisis de expresión relativa muestran que el tratamiento de vitrificación V7 provocó una disminución en la expresión (p<0,05) de MET1-2. En el caso del tratamiento V5 se encontró una disminución en la expresión de MET1-2, respecto al control (p<0,05). No se encontró diferencia entre embriones vitrificados con V5 y criopreservados durante dos horas (2H-V5); ambos mostraron una disminución (p<0,05) en la expresión de MET1-2.
Curiosamente, la criopreservación con V5 durante media hora (Mh-V5) no provocó cambios (p>0,05) en la expresión de MET1-2 con respecto al control. Lo anterior sugiere que la disminución en la expresión de MET1-2 observada en el tratamiento con Mh-V7 puede causar una disminución en la metilación del ADN, lo que puede estar relacionado con la sensibilidad de los embriones de uva a la criopreservación. Curiosamente, la vitrificación con V7 disminuyó la expresión de DRM1 (p<0,05), pero no se observó disminución (p>0,05) en la expresión de este gen por efecto de la criopreservación durante media hora (Mh-V7) y dos horas. (2H-V7) (Figura 12).
La disminución en la expresión de DRM1 observada en este estudio puede deberse a una disminución en la diferenciación celular y en los eventos de desarrollo embrionario causados por el efecto nocivo de la vitrificación y la criopreservación.

Conclusiones
Perspectivas
Gene expression profiles and in vitro development after vitrification of pronuclear and 8-cell stage mouse embryos. Molecular stability assessments of trees regenerated from cryopreserved mahogany (Swietenia macrophylla) seed germplasm using non-radioactive techniques to examine chromatin structure and DNA methylation status of ribosomal RNA genes. Intestinal health benefits of water-soluble carbohydrate concentrate of wild grape (Vitis thunbergii) in hamsters.
Changes in lipid peroxidation, total protein and antioxidant enzyme activity in buds of grapevine (Vitis vinifera L.) cv. Cryopreservation of axillary shoot tips of in vitro-grown grapes (Vitis) by a two-step vitrification protocol. Identification and relationships between accessions cultivated in the grapevine (Vitis vinifera L.) Germplasm Bank of Castilla y Léon (Spain) and the cultivars approved in the VQPRD areas of the region by SSR marker analysis.
Highly efficient vitrification protocols for cryopreservation of in vitro grown shoot tips of the rare and endangered plant Emmenopterys henryi Oliv.