123 Figura 29: Composición de la microbiota intestinal a nivel familiar en ratas pertenecientes al grupo de salchichas control (SC1 a SC10) y en ratas pertenecientes al grupo de salchichas funcionales (SF1 a SF10). 132 Figura 39: Composición de la microbiota intestinal a nivel familiar en ratones pertenecientes al grupo de alimentación (Feed 1 a Feed 10), y en ratones pertenecientes al grupo GOS-Lu (GOS-Lu 1 a GOS-Lu 10).
INTRODUCCIÓN
Enfermedades intestinales de origen inflamatorio
- Síndrome de intestino irritable
- Epidemiología
- Etiología y sintomatología
- Tratamiento clínico
- Enfermedad inflamatoria intestinal
- Epidemiología
- Etiología y sintomatología
- Diagnóstico y tratamiento
- Modelos animales
En el último siglo, la incidencia de la EII ha aumentado progresivamente, especialmente en los países desarrollados. La clasificación de Montreal sirve para estandarizar los casos de EII según la intensidad de los brotes: leve (que afecta al 50% de la población afectada), moderada (35%) y grave (15%) (Satsangi et al. 2006).

Carcinoma colorrectal
- Epidemiología
- Desarrollo del cáncer colorrectal
- Cáncer colorrectal hereditario
- Factores que influyen en la aparición de CCR
- Microbiota y CCR
- Cribado del CCR
- Tratamiento del CCR
- Modelos animales de CCR
- Modelos inducidos químicamente
- Modelos animales de CCR genéticos
Todos ellos producen aductos de ADN, estrés oxidativo y compuestos genotóxicos y mutagénicos (Cascella et al. 2018). Estas mutaciones del ADN son las que permitirán el desarrollo del CCR (Tong et al. 2011).

Nutracéuticos y alimentos funcionales
- Prebióticos
- Estructura química de los prebióticos
- Síntesis de SCFAs a partir de prebióticos
- Efectos intestinales de los SCFAs
- Probióticos
- Mecanismo de acción de los probióticos
- Fitoquímicos
- Ácidos grasos insaturados
Además, el mayor tamaño del colon en ratas facilita la detección y seguimiento de tumores (Amos-Landgraf et al. 2007). Esto implica que los FOS pueden derivarse de la inulina mediante hidrólisis debida a endoglicosidasas (van de Wiele et al. 2007). Entre estos filos productores de butirato destacan Actinobacteria, Fusobacteria, Proteobacteria, Spirochaetes y Thermotogae (Vital et al. 2014).
Sin embargo, también pueden metabolizar estos compuestos y evitar que interactúen con las células epiteliales (Wollowski et al. 2001). Aquí, en el grupo tratado con probióticos, se observó una disminución de los ácidos biliares en el intestino y un menor número de adenomas (Urbanska et al. 2009). Además, tienen efectos antiangiogénicos al inhibir los factores de crecimiento endotelial (VEGF) y sus receptores (Li et al. 2008).
En el mismo modelo animal, se añadió a la dieta cianidina-3-glucósido al 0,3%, lo que redujo el número de adenomas en un 45% (Cooke et al. 2006). Sin embargo, los ácidos grasos ω-3 activan la expresión del receptor gamma proliferador de peroxisomas que inhibe el factor NF-ĸB (Scaioli et al. 2017).

OBJETIVOS
Objetivos
Los objetivos generales de la tesis son estudiar y prevenir las enfermedades inflamatorias intestinales, especialmente CU y CCR, con alimentos funcionales con adición de nutracéuticos. Un estudio de alimentos cárnicos funcionales enriquecidos con ácidos grasos monoinsaturados para la prevención de la CU en un modelo animal. Estudio de alimentos cárnicos funcionales con antocianinas para la prevención del CCR en modelo animal.
Estudio de alimentos cárnicos funcionales con prebióticos derivados de lactulosa para la prevención del CCR en un modelo animal. Estudio de alimentos cárnicos funcionales con inulina para la prevención del CCR en un modelo de CCR inducido químicamente y otro de origen genético.
MATERIAL Y MÉTODOS
- Modelos animales
- Modelo animal para CU
- Modelo animal de CCR inducido químicamente
- Modelo animal de CCR genético
- Tipos de alimentación
- Medidas físicas
- Muestras de sangre y de tejidos
- Estudios histológicos
- Ensayo de la actividad de mieloperoxidasa
- Ensayo de permeabilidad de la mucosa
- Capacidad antioxidante en plasma sanguíneo
- Análisis de citoquinas en plasma sanguíneo
- Cuantificación de SCFAs en heces por cromatografía de gases masas
- Cuantificación de ácidos grasos por CG-MS en muestras cárnicas y de
- Extracción de ADN genómico y metagenómica de la subunidad del ARN
- Análisis filogenético de la secuenciación
- Cultivo celular
- Líneas celulares
- Estudios de proliferación celular
- Evaluación de la muerte y ciclo celular
- Análisis estadísticos y representación de datos
Las ratas se colocaron en el animalario (animalario de la Universidad de Oviedo) previa aprobación (n° CE y aprobación de los experimentos por el Comité Ético del Principado de Asturias (código de autorización PROAE 23/2016). Después de una semana con Después de la alimentación, la CCR se indujo en 8 ratas de cada grupo, excepto en las del experimento 3, en el que 7 animales de cada grupo fueron tratados utilizando el carcinógeno AOM (Sigma-Aldrich) disuelto en suero salino (NaCl al 0,9%) a una concentración de 2 mg/ml Las ratas se alojaron en el zoosanitario de la Universidad de Oviedo con permiso (Nº CE y aprobación del Comité Ético del Principado de Asturias (PROAE 15/2017).
A las ratas se les administró GOS-Lu al 10% disuelto en agua potable. Las ratas fueron anestesiadas con isoflurano al 3% (Ecuphar) y la mezcla de probióticos en PBS se administró a través de una cánula orofaríngea unida a una jeringa de 1 ml, lo que permitió administrar la dosis directamente en el estómago. Los ciegos de cada animal fueron pesados e inmediatamente congelados a -20 ºC para su posterior uso de su contenido en análisis de AGCC y estudios metagenómicos.
La identificación de SCFA se basó en el tiempo de retención de los estándares y la asistencia de la biblioteca Wiley 7. La secuenciación se realizó como se describe en la sección anterior con un mínimo de 100.000 lecturas por muestra de cada chip secuenciado.
RESULTADOS
Estudio de alimentos cárnicos funcionales enriquecidos con ácidos grasos
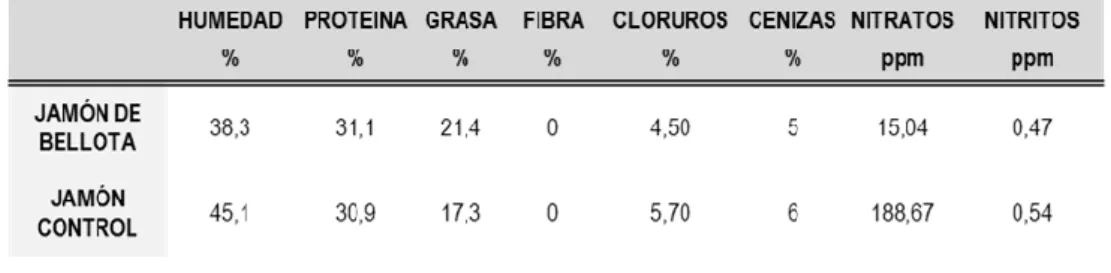
- Comparación nutricional entre el jamón control y el jamón ibérico de
- Efecto de las dietas en los pesos de los animales y en el índice de actividad
- Efecto del jamón de bellota en parámetros histológicos
- Efecto del jamón de bellota sobre los niveles plasmáticos de citoquinas y
- Efecto del jamón de bellota sobre la concentración de SCFAs en heces de
- Efecto del jamón de bellota en la concentración de ácidos grasos en plasma
- Efecto del jamón de bellota sobre la composición de la microbiota
En la cohorte de jamón de control, 3 animales murieron al inicio de la administración de DSS y otros 3 animales murieron al final de este tratamiento [Figura 12B]. Debido a la presencia de cambios reparativos en la mucosa colónica, parámetro que indica la reparación tisular tras la ulceración de la mucosa colónica, en la cohorte de jamón de bellota el valor medio de reepitelización (1,75) fue mayor que en la cohorte de alimentación (1,12). ). . [Figura 14B]. El valor medio de la concentración de ácido propiónico en el ciego de los animales de la cohorte alimentada con jamón de bellota (0,8546 mM) fue mayor que el de la cohorte alimentada (0,7851 mM), pero esto no difirió.
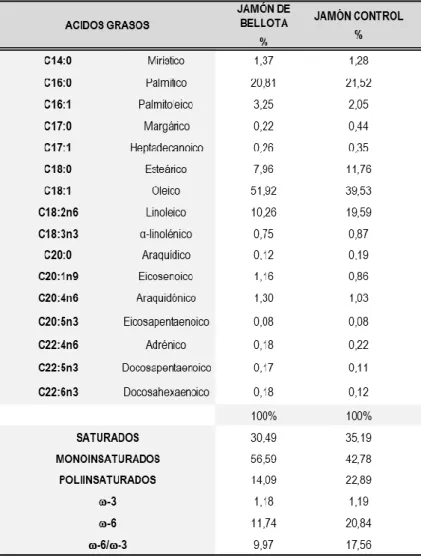
La Tabla 4 muestra los porcentajes de cada ácido graso en el plasma sanguíneo de ratas de la cohorte de jamón de bellota y de ratas de la cohorte de comida. Según el tipo de alimento en cada caso, los animales de la cohorte de jamón alimentado por hectárea presentaron mayores niveles plasmáticos del ácido graso monoinsaturado ácido oleico (31,61%). Este valor fue el doble del contenido de ácido oleico en las ratas de la cohorte de alimento (16,21%), y la diferencia fue estadísticamente significativa [Figura 17A].
La composición de la microbiota a nivel familiar, en general la composición de las familias en ambos tipos de cohortes de jamón fue muy similar y bastante diferente a la de los animales de la cohorte de alimentación [Figura 19]. También es de interés la presencia significativa (13% del total) de la familia Enterococcaceae en los dos animales supervivientes (después del tratamiento con DSS) de la cohorte de jamón de control.

Estudio de alimentos cárnicos funcionales con antocianinas para la
- Efecto de las salchichas funcionales sobre el peso corporal
- Efecto de las salchichas funcionales sobre el número de placas de Peyer
- Efecto de las salchichas funcionales sobre la capacidad antioxidante en
- Efecto de las salchichas funcionales sobre el peso del ciego
- Efecto de las salchichas funcionales sobre el número de tumores y el área
Sin embargo, de la Figura 22A se desprende claramente que durante las dos semanas de tratamiento con DSS (semanas 4 y 15), los animales de la cohorte de alimentación sufrieron una desaceleración en el aumento de peso. En la cohorte que recibió salchichas funcionales, el valor medio de las placas de Peyer fue 0,0018, siendo esta disminución en comparación con la cohorte de salchichas de control estadísticamente significativa (valor p de 0,0018) [Figura 23A]. Sin embargo, no se observó ninguna diferencia estadísticamente significativa cuando se comparó la cohorte de salchichas funcionales con la cohorte de salchichas de control (Figura 26A).
La secuenciación metagenómica de la microbiota intestinal mostró diferencias a nivel de composición del filo al comparar la cohorte de alimentación con las cohortes de salchichas de control y las cohortes de salchichas funcionales [Tabla 5]. La diferencia más significativa entre estas cohortes es la casi ausencia del filo Proteobacteria en la cohorte de alimentación (0,3%), mientras que este filo es el segundo más frecuente en la cohorte de salchichas de control (15,8%) y el tercero más frecuente en la cohorte de salchichas funcionales. cohorte (10,4%) [tabla 5]. En el caso de las ratas pertenecientes a la cohorte de salchichas control, el segundo filo más frecuente es Proteobacteria, pero en el caso de la cohorte de salchichas funcionales, el segundo filo más abundante es Bacteroidetes (12,1%, mientras que en la cohorte de salchichas control es el 12,4%). ) [Figura 27].
Clostridiaceae es una familia más extendida en la cohorte de salchichas funcionales (10,56%) en comparación con la cohorte de salchichas de control (8,28). Desulfovibrionaceae y Enterobacteriaceae están menos presentes en la cohorte de salchichas funcionales (respectivamente 7,96% y 2,38%) con la cohorte de salchichas de control (11,09). % y 4,65%, respectivamente).

Estudio de alimentos cárnicos funcionales con prebióticos derivados de
- Efecto del GOS-Lu sobre el peso corporal
- Efecto del GOS-Lu sobre el peso del ciego intestinal
- Efecto del GOS-Lu sobre el número de pólipos y la extensión colónica
- Efecto del GOS-Lu sobre la concentración de SCFAs en heces del ciego
- Efecto del GOS-Lu sobre la composición de la microbiota intestinal
- Correlaciones entre microbiota intestinal, producción de SCFAs y
- Cultivo celular
- Ensayos de viabilidad celular mediados por SCFAs
- Ensayos de apoptosis celular mediados por SCFAs
- Ensayos de ciclo celular mediados por SCFAs
En la cohorte GOS-Lu también hubo un aumento en la abundancia de Actinobacteria y una disminución en Tenericutes [Tabla 6]. Como descripción general de la composición de la microbiota intestinal, el índice de diversidad de Shannon aumentó en la cohorte GOS-Lu [Figura 37A]. Las familias Bacteroidaceae (4,22% en la cohorte de control, 9,4% en GOS-Lu) y Prevotellaceae (1,59% en la cohorte de alimentación vs. 5,84% en la cohorte de GOS-Lu) fueron las familias representativas del aumento del filo Bacteroidetes en la cohorte GOS -Lu, en comparación con el grupo alimentado [Figuras 38 y 40].
La disminución en el filo Firmicutes que ocurrió en la cohorte CIS-Lu se debió principalmente a una reducción en la familia Lachnospiraceae (33,47% en la cohorte alimentada versus 14,18% en la cohorte CIS-Lu) y, en menor medida, a una reducción en la familia Eubacteriaceae (1,33% en la cohorte de alimentación versus 0,71% en la cohorte GOS-Lu) [Figuras 38 y 40]. En la línea HT29, se lograron reducciones del 80 % en la viabilidad con concentraciones cercanas a 4 mM [Figura 43B]. En la línea HCT116 [Figura 44A] el butirato requirió concentraciones de 0,94 mM para alcanzar la IC50, mientras que en propionato se requirieron concentraciones de 4,06 mM (4,3 veces mayores) [Tabla 8].
Finalmente, en la línea celular T84 [Figura 44C], la concentración requerida de butirato fue de 2,68 mM frente a los 61,64 mM necesarios en el caso del propionato (23 veces mayor) para conseguir el mismo parámetro de inhibición [Tabla 8]. En la línea celular HCT116, tanto el butirato (71% de apoptosis) como el propionato (54% de apoptosis) mostraron diferencias estadísticamente significativas en comparación con el control (línea celular con H2O, 2,62% de apoptosis) [Figura 45A].
Estudio de alimentos cárnicos funcionales con inulina para la prevención de
- Efecto del alimento funcional sobre los niveles plasmáticos de
- Efecto del alimento funcional sobre el peso del ciego
- Efecto del alimento funcional sobre el número placas de Peyer
- Efecto del alimento funcional sobre el número de pólipos y la superficie
- Efecto del alimento funcional sobre la concentración de SCFAs en
Esta disminución fue estadísticamente significativa en comparación con la media del grupo de carne de control (valor de p <0,0001) [Figura 50]. Sin embargo, en el grupo funcional de la carne, las tres familias principales fueron Prevotellaceae (22,8%), Bacteroidaceae (15,8%) y Porphyromonadaceae (12%) [Figura 54]. Así, en el grupo de alimentación se observó que la ganancia de peso fue negativa en el 50% de los animales [Figura 12A].
En la cohorte de jamón alimentado con bellota, la pérdida de microbiota productora de butirato, como las especies de las familias Lachnospiraceae y Ruminococcaceae [Figura 16E], podría haberse compensado con un aumento de microorganismos capaces de producir otros AGCC como isobutirato, isovalerato y valerato. [Figura 16A, B y C]. En este trabajo se han estudiado los animales de la cohorte de jamón de bellota que presentaban altas poblaciones de B. La alta presencia del género Enterococcus en la cohorte de jamón control respecto a las otras dos cohortes (donde representaban menos del 0,05%) podría deberse a la capacidad de estas bacterias de obtener energía a partir de la reducción de los nitritos presentes, generando óxido nítrico a través de la vía de desnitrificación (donde están involucrados los genes nirK y nirS) (Heylen et al. 2006).
Los resultados mostraron un menor número de placas de Peyer hiperplásicas en la cohorte de embutidos funcionales (un 27,1% menos que en los embutidos normales), y este hecho podría explicarse por el efecto antiinflamatorio que ejercen las antocianinas en este tipo de alimentos (un 0,11% menos que en los embutidos normales). del contenido total de esta matriz cárnica). La familia Eubacteriaceae (Firmicutes) también sufrió niveles reducidos en la cohorte GOS-Lu. Sin embargo, en este modelo animal, Bilophila wadsworthia (otro miembro de la familia Desulfovibrionaceae) aumentó en la cohorte GOS-Lu.
La presencia de la familia Ruminococcaceae (Firmicutes) se redujo significativamente en la cohorte funcional en comparación con la cohorte de carne de control.