AGRADECIMIENTOS
RESUMEN
ABSTRACT
INTRODUCCIÓN
Las ascidias (Phyllum Chordata, clase Ascidiacea) representan invasores conspicuos en los ecosistemas marinos (Zhan et al. 2015). Las ascidias son organismos exitosos en la competencia por el espacio (Grosberg 1981; Nandakumar et al. 1993, Osman & Whitlach 1995a; Castilla et al. Sus rápidas expansiones poblacionales pueden reducir la abundancia de especies bentónicas previamente establecidas y pueden causar cambios significativos en la estructura de la comunidad bentónica (Lambert 2001; Castilla et al. 2004a, b).
Estos estudios resaltan la importancia de la coexistencia de varios ingenieros ecosistémicos y sus interacciones en el mantenimiento de la biodiversidad (Cerda & Castilla 2001; Caro et al. 2011). En los ambientes marinos costeros, la disponibilidad de sustrato es uno de los factores más importantes que influyen en la distribución y abundancia de organismos macrobentónicos sésiles (Bertness et al. 2014). Varios factores pueden influir en la intensidad de la epibiosis, incluida la estacionalidad, la latitud y la profundidad (Wahl 1989; Barnes 1994; Vasconcelos et al. 2007).
La adición de organismos a menudo crea nuevas estructuras tridimensionales que favorecerían a las comunidades bentónicas (Johnson et al. 2003; Tokeshi & Arakaki 2012). Un estudio realizado en la Bahía San Matías entre 2005 y 2007 registró la presencia de varias ascidias introducidas, entre ellas Diplosoma listerianum, Ciona intestinalis, Lissoclinum frágil, Molgula manhattensis, Cnemidocarpa robinsoni y Corella eumyota (Schwindt et al. 2014). .
MÉTODOS
- Área de estudio
- Muestreo y análisis de datos
- a Abundancia, riqueza, equitatividad y diversidad de la macrofauna asociada a
- b Grupos funcionales de movilidad asociados a Styela clava en Baliza San Matías y
Estos canales forman parte de los niveles más bajos de la zona intermareal donde emergen formaciones rocosas. Fuera de la BSA, hacia el este, se encuentra la Baliza San Matías (BSM). Cada muestra fue fotografiada antes de su recolección (Figura 4), luego se colocó individualmente en una bolsa de plástico con cierre hermético para evitar la pérdida de epibiontes móviles y las bolsas se etiquetaron adecuadamente.
En el laboratorio, el procesamiento de las muestras consistió en la cuidadosa separación de los epibiontes (individuos de macrofauna, es decir, mayores a 500 micras) asociados a cada individuo de S. De cada muestra de macrofauna obtenida se obtiene la abundancia total (número total de individuos) de los taxones registrados. En el caso de los filos, se analizaron las variaciones temporales en las abundancias entre sitios.
Se utilizaron datos de abundancia para todos los taxones por sitio para visualizar la composición de la comunidad en cada sitio. Cuando se detectaron diferencias significativas, se calcularon los porcentajes de contribución de similitud (SIMPER) de cada uno de los taxones para determinar cuál de los taxones considerados contribuyó más a las diferencias observadas.

RESULTADOS
- Abundancia, riqueza, equitatividad y diversidad de la macrofauna asociada aStyela
- Grupos funcionales de movilidad asociados a Styela clava en Baliza San Matías
Especies asociadas a Styela clava en Baliza San Matías (BSM) y Punta Verde (PV) para todos los meses de estudio. Los nematodos fueron agrupados y considerados dentro de la clase Adenophorea, y fueron registrados como una sola especie para ambos sitios para este trabajo. Abundancia, expresada como porcentaje, de la macrofauna asociada a Styela clava, a nivel de Phyla, durante todo el periodo de muestreo en Baliza San Matías (BSM) y Punta Verde (PV).
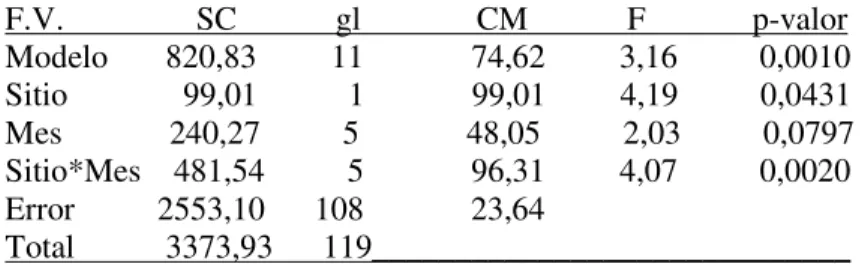
En BSM se observó un patrón inverso al PV, ya que los valores de abundancia más bajos se registraron en los meses de marzo y mayo (464 y 351 individuos, respectivamente) y el pico de abundancia se observó en enero con 1300 individuos (Figura 8). ). Cantidades totales para Baliza San Matías (BSM) y Punta Verde (PV) en todos los meses de muestreo. Respecto a las clases Bivalvia y Hexanauplia, se observaron dos picos de abundancia en los meses de noviembre y enero.
La clase Gastropoda dominó en enero, con valores de abundancia variables en el resto de los meses. Para el sitio PV, como se mencionó, la clase Malacostraca fue la más numerosa, en marzo y mayo se observaron los picos de abundancia, en el resto de los meses mostró valores de abundancia relativamente estables. Se registraron organismos de la clase Insecta con valores de abundancia bajos en enero, marzo y noviembre, lo que coincide con lo observado en BSM.
ANOVA bidireccional: Comparación de abundancia de macrofauna asociada a Styela clava entre Baliza San Matías (BSM) y Punta Verde (PV) y entre meses de estudio. Para los meses restantes, las diferencias entre sitios no fueron significativas (prueba de Tukey, p > 0,05). ANOVA de dos vías: Comparación de la riqueza específica de la macrofauna asociada a Styela clava entre Baliza San Matías (BSM) y Punta Verde (PV) y entre los meses de estudio.
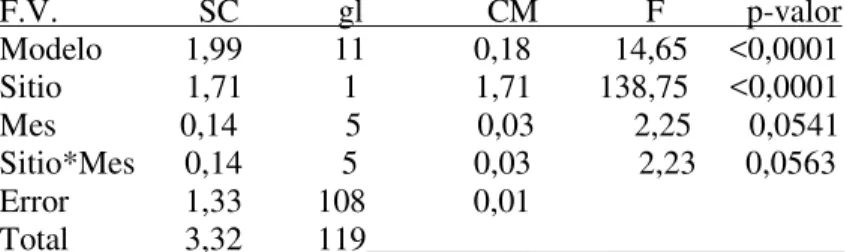
Sin embargo, las diferencias entre las localidades no siempre fueron de la misma magnitud a lo largo del año (interacción débil no significativa entre factores). ANOVA de dos vías: Comparación del índice bursátil Pielou entre Baliza San Matías (BSM) y Punta Verde (PV) y entre los meses estudiados. La interacción significativa indica que estas diferencias no fueron de la misma magnitud en todos los meses examinados (prueba de Tukey, p <.
En BSA, este índice de diversidad se mantuvo relativamente estable en torno al mismo valor medio, mientras que en PV disminuyó hacia los meses cálidos. ANOVA de dos vías: Comparación del índice de diversidad de Shannon-Wiener entre Baliza San Matías (BSM) y Punta Verde (PV) y entre meses estudiados.
DISCUSIÓN…
- Abundancia, riqueza, equitatividad y diversidad de la macrofauna asociada a
- Grupos funcionales de movilidad asociados a Styela clava en Baliza San Matías
49. El interior de la Bahía de San Antonio difiere del exterior; y 2) las diferencias en diversidad y abundancia de organismos asociados con S. Aunque PV mostró una mayor abundancia general que BSM, esta diferencia fue impulsada por la dominancia de la clase Malacostraca. En otro ejemplo, en Chile se registraron entre 72 (Castilla et al. 2004) y 96 (Cerda & Castilla 2001) especies de organismos macrofaunales asociados a la ascidia Pyura praeputialis.
Respecto a estudios con metodología similar a esta tesis, cabe mencionar que por Voltsiadou et al. 2007) en Grecia, quienes registraron 38 especies del superorden Peracarida de la colección de 41 individuos seleccionados al azar de la ascidia Microcosmus sabatieri. 2015), registró 21 especies pertenecientes a 6 filos asociados con 28 individuos de tres especies de ascidias solitarias: Corella antartica, Cnemidocarpa verrucosa y Molgula pedunculata. Entre los diferentes factores que podrían explicar las diferencias en abundancia y en el número y composición taxonómica, se encuentran las diferencias latitudinales (Romero et al. 2014), las condiciones ambientales estresantes (Castilla et al. 2004b), la superficie corporal que ofrece el basibionte ( Rimondino et al. 2015), la composición química de la túnica (Stoecker et al. 1978; McClintock et al. 2004; Nylund & Pavia 2005) o la naturaleza dependiente del contexto de la epibiosis (Berke 2010; en el caso de las ascidias, en PV la disponibilidad de nutrientes también jugará un papel fundamental, ya que el establecimiento de ascidias se ve favorecido en ambientes eutróficos protegidos de la acción de las olas (Lambert et al. 2003; Simkanin et al. 2012).
Además de la influencia directa del aporte de sustrato, el efecto de las ascidias podría ser indirecto al concentrar sedimentos en su superficie. En el PV, la alta abundancia de estos organismos posiblemente se deba a la presencia de sedimentos móviles además de la disponibilidad de nutrientes ya mencionados. Otro grupo que podría beneficiarse de la cantidad de sedimento retenido son los nematodos, aunque esta hipótesis requiere de trabajos experimentales para confirmarla, ya que generalmente son organismos ubicuos (Fauchhald & Jumars 1979).
Por lo tanto, las variables ambientales que no están mediadas por organismos basibones (como la temperatura o el aumento de recursos/alimentos en la columna de agua) pueden influir significativamente en la composición de la comunidad macrofaunal. Este suministro constante de sedimento durante las mareas altas y bajas es forzado por las corrientes de marea de (< 0,5 m s-1) presentes en la BSA (Aliotta et al. 2000; Saad, com. personal). Por lo tanto, en ausencia de sedimentos arenosos o limo-fangosos típicos de ambientes de baja energía (Fucks et al. 2012), no habría suficiente sustrato móvil en BSM para acumularse en la túnica de S.
En estudios realizados con macrofauna asociada a mejillones y ascidias, la retención de sedimentos se correlacionó positivamente con el aumento de la diversidad de organismos (Gibbons 1988; Moreno & Rocha 2006). Esto explicaría por qué los anfípodos, nematodos y, en menor medida, los poliquetos contribuyen a la disparidad entre sitios, al ser taxones con preferencia por sustratos con sedimento móvil (Boeckner et al. 2009). Asimismo, es interesante señalar que otras ascidias solitarias como Pyura praeputialis en Chile (Castilla et al.
Gelós EM, Spagnuolo J, Schillizzi RA (1992) Las unidades morfológicas de la costa occidental del Golfo de San Matías y su evolución. Pereyra PJ, de la Barra P, Gastaldi M, Saad JF, Firstater FN, Narvarte MA (2017) Cuando los pequeños ayudan a los poderosos: facilitación entre dos especies introducidas, una ascidia solitaria y una macroalga en el norte de la Patagonia, Argentina.