UNIVERSIDAD NACIONAL DEL CENTRO DEL PERÚ
FACULTAD DE INGENIERÍA QUÍMICA
TESIS
PRESENTADA POR LOS BACHILLERES:
IZARRA PORRAS CESAR MIGUEL PEÑA PAUCAR YENNY BEATRIZ
Para optar el Título Profesional de Ingeniero Químico
HUANCAYO-PERÚ 2022
EFECTO DE LA TEMPERATURA, pH Y CELULASA EN LA CONCENTRACION DE GLUCOSA PRODUCIDA POR HIDRÓLISIS CON
RESIDUOS DE MAIZ MORADO
_____________________________________________________
ASESOR:
Dr. WILDER EFRAÍN EUFRACIO ARIAS
_____________________________________________________
DEDICATORIA Primero que todo a Dios por llenarme de
Bendiciones
A mis padres Hermogenes e Hilda, una pareja exitosa en la profesión más difícil, ser padres
A mis maestros, que consideraron en mi la profesión y la persona
A mis hermanos Walter, Fredy, Luis, José y Saúl y familiares, que son solo el hecho de existir en mi vida y sin saberlo, me acompañaron al logro de esta meta.
Finalmente, dedicado a todo aquel que creyó en mí, Teófila Porras, Macedoño porras.
Izarra Porras Cesar Miguel
A mi amado Dios, por el milagro de la vida y el regalo de la capacidad de aprender.
Con amor para mis padres Emiliana Paucar Ayuque y Donato Peña Torres, guerreros incansables que nunca dejaron de estar a mi lado.
A mis hermanos Viviana Ramirez, Erika Peña, Fidel Peña y Jhonatan Peña, mis soportes, mis amigos y mis cómplices en los momentos buenos y malos, que me han permitido ver lo bello que es la vida.
A mis familiares, que sin saberlo con el solo hecho de pensar en mí, han aligerado este camino.
Finalmente, para todos los que confiaron en mí.
Yenny Beatriz Peña Paucar
AGRADECIMIENTO
La realización de esta tesis no habría sido posible sin la participación de muchas voluntades. Desde mi formación profesional hasta a lo largo de todo el proceso he contraído deudas de gratitud con muchas personas a las que me gustaría expresar, desde estas páginas, mi más sincero agradecimiento.
Gracias le damos a Dios por iluminarnos y bendecirnos durante toda nuestra carrera y por este gran objetivo alcanzado.
Eterna gratitud a nuestros padres por su inmenso amor, que se plasma en nuestro triunfo que no es más que el reflejo del trabajo y la formación que nos han dado a lo largo de una vida.
A nuestros hermanos, por su ejemplo, confianza, amor y la fortaleza que nos han brindado en los momentos de debilidad.
A nuestro asesor Dr. Wilder Efrain Eufracio Arias, cuya visión profesional y humana ha ido más allá de la simple labor de un profesor asesor. Ha sido una satisfacción trabajar con él.
A la Universidad y a su cuerpo docente por la formación que nos brindaron, su interés constante por facilitarnos sus conocimientos y su expertis.
Finalmente, sin el ánimo de olvidarnos de quienes son importantes en nuestras vidas
¡¡¡¡Gracias a todos¡¡¡¡
RESUMEN
En el pretratamiento se altera la estructura de la celulosa y la producción de azucares es baja, por lo que es necesario la hidrólisis para incrementar la conversión de la celulosa y la hemicelulosa en azúcares simples, en este proceso influyen las características de la biomasa y enzimas. Por lo cual en la tesis se determinó el efecto de la temperatura, pH y dosis de celulasa en la concentración de glucosa producida por hidrólisis de residuos de maíz morado.
En los experimentos se utilizó 25 g de residuos de maíz morado y se realizó el pretratamiento con 250 mL de solución de H2SO4 al 0.1% y 65 °C durante 2 h. Luego se realizó la hidrólisis enzimática y se evaluó la temperatura (55, 60 y 65 °C), pH (3.5, 4.5 y 5.5) y dosis de celulasa (0.25, 0.50 y 0.75 mL/L). Se usó el diseño experimental de Box-Behnken y para la optimización el método de superficie de respuesta.
La temperatura, pH y dosis de celulasa óptima de la hidrólisis en la producción de glucosa con residuos de maíz morado es 55 °C, 5.5 y 0.25 mL/L, respectivamente.
La dosis de celulasa, [temperatura]2, [pH]2 y [dosis de celulasa]2 influyen en la concentración de glucosa en la hidrólisis enzimática de los residuos de maíz morado pretratados. La mayor concentración de glucosa obtenido es 14.50 g/L y el óptimo estimado por el método de superficie de respuesta es 16.92±1.00 g/L.
Palabras claves: hidrólisis enzimático, azucares reductores, glucosa, residuos de maíz morado.
INTRODUCCIÓN
La producción de bioetanol de segunda generación involucra el pretratamiento, hidrólisis y fermentación, en la cual cada una tiene sus propias variables que influyen en el rendimiento de cada proceso y en forma global en la cantidad de etanol producido.
En el pretratamiento se incrementa la superficie, rompe estructuras complejas y disuelve la hemicelulosa y/o lignina de la biomasa para facilitar la accesibilidad de las enzimas en la hidrólisis y convertir los materiales celulósicos en azúcares simples. En la fermentación se produce la conversión de los azucares en etanol con la ayuda de una levadura. Los tres procesos tienen sus propias variables que tienen que ser determinados experimentalmente influyen en su rendimiento y en forma global en la cantidad de etanol producido.
La tendencia creciente en el mundo de la generación de residuos se refleja en una contaminación ambiental cada vez más crítica, otro aspecto que agrava este problema en la mayoría de lugares es la limitada implementación de un sistema de gestión de residuos sólidos. Al respecto (Li & Yang, 2016, p. 619), indica que los alimentos se desperdician a lo largo de la cadena de suministro de alimentos, desde la producción agrícola inicial hasta el consumo doméstico final y que los productos vegetales suman alrededor del 85% de la FW total y el 15% restante proviene de productos de origen animal.
Esta es la razón para investigar alternativas de solución y aprovechar los residuos de maíz morado que contienen gran cantidad de carbohidratos para la obtención de glucosa u otros azúcares y luego convertirlos en etanol, con ello se aporta en la mitigación del impacto ambiental negativo debido a este residuo.
Las condiciones de la hidrólisis enzimática tienen que ser determinadas experimentalmente por las características particulares de la biomasa o sustrato (materia prima) y las enzimas que se utilizan, por ello en la tesis se determinó el efecto de la temperatura, pH y celulasa en la concentración de glucosa producida por hidrólisis de residuos de maíz morado. Los granos del maíz que quedan como residuo después de la cocción para la elaboración de la chicha morada se utilizaron en la investigación.
El cuerpo del informe de la tesis contiene tres capítulos, en el primer capítulo se presenta la revisión bibliográfica (antecedentes, bases teóricas y marco conceptual), en el que se detalla el sustento teórico de nuestro estudio. El segundo capítulo se presenta la parte experimental en la que se describe los materiales, equipos e instrumentos que se utilizó, y el procedimiento seguido en los experimentos. Finalmente, en el último capítulo se desarrolla el tratamiento de datos y discusión de resultados. El informe termina con las conclusiones, recomendaciones y anexos.
OBJETIVOS Objetivo General
• Determinar el efecto de la temperatura, pH y dosis de celulasa en la concentración de glucosa producida por hidrólisis de residuos de maíz morado.
Objetivos Específicos
• Determinar la temperatura de la hidrólisis en la producción de glucosa con residuos de maíz morado.
• Determinar el pH de la hidrólisis en la producción de glucosa con residuos de maíz morado.
• Determinar la dosis de celulasa para la hidrólisis en la producción de glucosa con residuos de maíz morado.
• Determinar la concentración de glucosa en la hidrólisis de residuos de maíz morado.
GLOSARIO DE TERMINOS ABE = Acetona, butanol y etanol
BC = Celulosa bacteriana BG o BGL = β-glucosidasas
CBD = Dominio de unión a celulosa CBH = Celobiohidrolasas
CBM = Módulo de unión a carbohidratos CBP = Bioprocesamiento consolidado CCP = Polvo de mazorca de maíz CD = Dominio catalítico
CS = Rastrojo de maíz seco CS-DW = CS con agua destilada
CS-DW-GNa = CS con agua destilada, glicerol y NaHCO3
CSP = Polvo de rastrojo de maíz
EC = Enzyme Commission o Comisión de Enzimas EG = Endoglucanasas o endo-β-1,4-glucanasa FSC = Cadena de suministro de alimentos FW = Desechos o desperdicios de cocina
GA = Glucoamilasas
GAU = Unidad de glucoamilasa GEI = Gases de efecto invernadero GH = Glucósido hidrolasas
GP = Grado de polimerización
IL = Líquidos iónicos
IU = Unidad internacional LCB = Biomasa lignocelulósica
LPMO = Monooxigenasas de polisacáridos líticos
MS = Materia seca
RSU = Desechos sólidos municipales SHF = Hidrólisis y fermentación separadas
SSCF = Sacarificación y cofermentación simultáneas SSF = Sacarificación y fermentación simultáneas TAG = Triacilglicéridos
TCA = Ácido tricarboxílico
WB = Salvado de trigo
SIMBOLOGIA
Símbolo Descripción Unidades
pH Potencial de hidrógeno Adimensional
T Temperatura OC
ÍNDICE
ASESOR ii
DEDICATORIA iii
AGRADECIMIENTO iv
RESUMEN v
INTRODUCCIÓN vi
OBJETIVOS vii
GLOSARIO DE TERMINOS viii
SIMBOLOGIA ix
ÍNDICE x
Índice de Tablas xii
Índice de Figuras xiii
CAPITULO I
REVISIÓN BIBLIOGRÁFICA
1.1 Antecedentes 1
1.2 Bases teóricas 5
1.2.1 Maíz morado 5
a) Taxonomía. 8
b) Composición química. 8
c) Usos. 9
1.2.2 Hidrólisis 9
a) Tipos de hidrolisis. 10
b) Factores críticos de la hidrolisis. 11
1.2.3 Hidrólisis enzimática 11
a) Enzimas. 13
b) Clasificación de las enzimas. 13
c) Estructura y mecanismos de las enzimas. 14
d) Cofactores y coenzimas. 15
e) Función biológica de las enzimas. 16
f) Factores que influyen en la hidrolisis enzimática. 16
1.2.4 Alfa-amilasa, glucoamilasa y celulasa 23
a) Alfa amilasa. 23
b) Glucoamilasa. 24
c) Celulasa. 25
1.2.5 Energía de biomasa 28
1.2.6 Biocombustibles 30
1.2.7 Producción de bioetanol de segunda generación 33
1.2.8 Biomasa lignocelulósica 35
Análisis proximal de residuos agrícolas 37
a) Componentes de la biomasa. 38
1.2.9 Desechos o desperdicios de cocina 44
1.2.10 Almidón 46
1.3 Marco conceptual 50
CAPITULO II PARTE EXPERIMENTAL
2.1. Materia prima 52
2.2. Materiales 52
2.3. Reactivos 53
2.4. Enzimas 53
2.5. Equipos e instrumentos 53
2.6. Procedimiento 54
2.6.1 Determinación de la concentración de azucares reductores por el método
de DNS 54
a) Preparación del reactivo DNS. 54
b) Preparación de la curva estándar. 54
2.6.2 Pretratamiento ácido de los residuos de maíz morado 55
2.6.3 Hidrólisis enzimática 55
2.7. Diseño de investigación 56
CAPITULO III
TRATAMIENTO DE DATOS Y DISCUSIÓN DE RESULTADOS
3.1 Presentación y análisis de los datos 57
3.1.1 Aspectos descriptivos de las variables 57
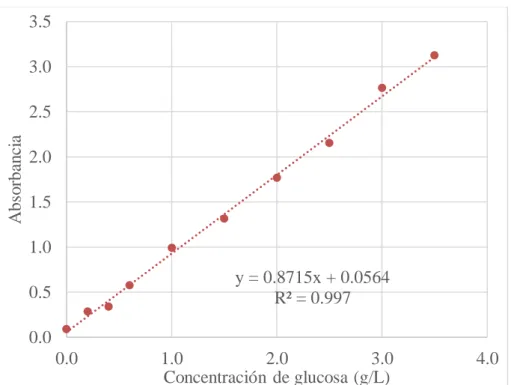
3.1.2 Curva de estándar 57
3.1.3 Determinación de la temperatura, pH y dosis de celulasa de la hidrólisis en la producción de glucosa con residuos de maíz morado. 58 3.1.4 Optimización de la concentración de azucares reductores en la hidrólisis enzimática de los residuos de maíz morado pretratados 60
3.2 Proceso de la prueba de hipótesis 63
3.3 Discusión de resultados 64
CONCLUSIONES 67
RECOMENDACIONES 68
BIBLIOGRAFÍA 69
ANEXOS 72
Anexo A: Fotografías de los experimentos 73
Índice de Tablas
Tabla 1. Composición química del maíz morado INIA-615 Negro Canaán 8 Tabla 2. Disponibilidad global de biomasa apta para la conversión de energía según
Woolf et al. (2010) 30
Tabla 3. Materia prima más utilizada para producir bioetanol 34 Tabla 4. Composición lignocelulósica de diferentes residuos agrícolas 37
Tabla 5. Análisis proximal de residuos agrícolas 37
Tabla 6. Producción agrícola total versus volúmenes de desperdicio de alimentos a
nivel mundial 46
Tabla 7. Contenido de amilosa y amilopectina (% de almidón total) en diferentes
fuentes de almidón 47
Tabla 8. Reactivos utilizados en la investigación 53
Tabla 9. Enzimas y especificaciones técnicas utilizadas en la investigación 53
Tabla 10. Niveles de las variables 56
Tabla 11. Matriz de diseño 56
Tabla 12. Estadísticos descriptivos de la concentración de azucares reductores en función a la temperatura de la hidrólisis enzimática 57 Tabla 13. Estadísticos descriptivos de la concentración de azucares reductores en
función al pH de la hidrólisis enzimática 57
Tabla 14. Estadísticos descriptivos de la concentración de azucares reductores en función a la dosis de celulasa de la hidrólisis enzimática 57 Tabla 15. Absorbancia en función a la temperatura, pH y dosis de celulasa de la hidrólisis enzimática de los residuos de maíz morado pretratados 59 Tabla 16. Concentración de azucares reductores en función a la temperatura, pH y dosis de celulasa de la hidrólisis enzimática de los residuos de maíz morado
pretratados 59
Tabla 17. Análisis de varianza
64
Índice de Figuras
Figura 1. Estructura de un grano de maíz 7
Figura 2. Características composicionales de tres variedades 9 Figura 3. Diagrama esquemático del proceso de hidrólisis enzimática de la biomasa lignocelulósica: 1) difusión enzimática, 2) adsorción y 3) hidrólisis 12
Figura 4. Papel de la α-amilasa 24
Figura 5. Mecanismo de acción de la celulasa 26
Figura 6. Mecanismo de acción de la celulasa 28
Figura 7. Producción de biocombustibles a partir de biomasa (izquierda) y principales rutas biosintéticas utilizadas para la producción de biocombustibles (derecha) 32 Figura 8. Evolución de la investigación en biocombustibles 33 Figura 9. Procesos para la conversión de biomasa lignocelulósica en bioetanol 35 Figura 10. Composición y estructura de la pared celular de la biomasa lignocelulósica
36 Figura 11. Fraccionamiento de biomasa lignocelulósica 37
Figura 12. Diferentes constituyentes de la biomasa 38
Figura 13. Estructura de la celulosa 40
Figura 14. Componentes básicos de la lignina 43
Figura 15. Desperdicio mundial relativo de alimentos por fase de la cadena de
suministro de alimentos 45
Figura 16. Estructura de amilosa que muestra una cadena de unidades de D-
glucopiranosa unidas a α-1,4 que dan como resultado una hélice 48 Figura 17. Estructura de la amilopectina que muestra una cadena de unidades de d-
glucopiranosa unidas a α-1,4 con ramificación α-1,6 49
Figura 18. Representación en forma de árbol de la amilopectina que muestra los
diferentes tipos de cadenas poliméricas 50
Figura 19. Curva estándar 58
Figura 20. Gráfica de contorno de la concentración de azucares reductores en función al pH y temperatura de la hidrólisis enzimática de los residuos de maíz
morado 60
Figura 21. Gráfica de contorno de la concentración de azucares reductores en función a la dosis de celulasa y temperatura de la hidrólisis enzimática de los
residuos de maíz morado 61
Figura 22. Gráfica de contorno de la concentración de azucares reductores en función a la dosis de celulasa y pH de la hidrólisis enzimática de los residuos de
maíz morado 61
Figura 23. Gráfica de superficie de la concentración de azucares reductores en función al pH y temperatura de la hidrólisis enzimática de los residuos de maíz
morado 62
Figura 24. Gráfica de superficie de la concentración de azucares reductores en función a la dosis de celulasa y temperatura de la hidrólisis enzimática de los
residuos de maíz morado 62
Figura 25. Gráfica de superficie de la concentración de azucares reductores en función a la dosis de celulasa y pH de la hidrólisis enzimática de los residuos de
maíz morado 63
Figura 26. Valores óptimos de la temperatura, pH y dosis de celulasa de la hidrólisis
enzimática de los residuos de maíz morado 63
Figura 27. Muestras de los residuos de maíz morado 72
Figura 28. Cubetas con contenido de las muestras para la curva estándar 72 Figura 29. Solución de H2SO4 al 0.1% v/v para el pretratamiento ácido de los residuos
de maíz morado 72
Figura 30. Hidrólisis enzimática de los residuos de maíz morado 73 Figura 31. Toma de 1 mL del líquido de la hidrólisis para la determinación de los
azucares reductores por el método DNS 73
Figura 32. Lectura de la absorbancia de la muestra en el espectrofotómetro de UV 74
CAPITULO I
REVISIÓN BIBLIOGRÁFICA 1.1 Antecedentes
Hui Zhang & Wu (2021), en su publicación “Statistical optimization of aqueous ammonia pretreatment and enzymatic hydrolysis of corn cob powder for enhancing sugars production”, aplicaron la metodología de superficie de respuesta (RSM) para la optimización sistemática del pretratamiento con amoníaco acuoso y la hidrólisis enzimática del polvo de mazorca de maíz (CCP) utilizando celulasas de producción propia de Aspergillus niger HQ-1. Utilizaron polvo de mazorca de maíz (CCP) y polvo de rastrojo de maíz (CSP) con diferentes tamaños de partículas de CCP, incluidos 0.3 mm y 0.5 mm, para optimizar el pretratamiento con amoníaco acuoso. Prepararon un medio con pH inicial de 4.7 mezclando 10.0 g de sustratos sólidos que contenían 5.0 g de CSP y 5.0 g de WB con 23.67 mL de solución mineral que contenía 1.267% (p/v) de (NH4)2SO4, 0.253% (p/v) de KH2PO4 y 0.127% (p/v) de MgSO4⋅7H2O. Prepararon el inóculo de semillas con una solución de esporas y un medio precultivado de glucosa con un pH inicial de 7.0 que contenía 1.5% (p/v) de glucosa, 0.4% (p/v) de extracto de levadura, 0.2% (p/v) de K2HPO4 y 0.1% de MgSO4⋅7H2O. Las variables significativas que estudiaron fue la concentración de amoníaco acuoso, temperatura y tiempo durante el pretratamiento. Los resultados mostraron que el CCP con un tamaño de partícula de 0.3 mm debe pretratarse con una solución acuosa de amoníaco al 11.6% en peso con una carga de sólidos al 20.0% (p/v) a 71.3 °C durante 8.8 h. A las condiciones optimizadas de pretratamiento lograron un 97.1% de recuperación de celulosa, 66.6%
de recuperación de hemicelulosa y 75.7% de eliminación de lignina. En la hidrólisis enzimática optimizaron la carga de enzimas, tiempo de reacción y carga de biomasa, y obtuvieron el azúcar reductor máximo (822.31 mg/g de sustrato seco, mg/gds) y xilosa (316.72 mg/gds) mientras que el CCP pretratado se hidrolizó a 50.0 °C, pH 4.8 durante 36.6 h con 10.9 FPU/gds (FPU/g sustrato seco) de carga de enzima, 6.95% (p/v) de carga de biomasa y 0.5% (p/v) de Tween-80. Las condiciones optimizadas de hidrólisis enzimática podrían resultar en un 92.1% de conversión de holocelulosa y un 79.3% de conversión de hemicelulosa. Sin embargo, se requiere más investigación para la optimización de la fermentación para la producción de etanol por cepas de levadura utilizando el hidrolizado del CCP pretratado.
He et al. (2020), en el artículo “Dynamics of fermentation quality, physiochemical property and enzymatic hydrolysis of high-moisture corn stover ensiled with sulfuric acid or sodium hydroxide”, investigaron la dinámica de la calidad de la fermentación, propiedades fisicoquímicas e hidrólisis enzimática del rastrojo de maíz con alto
contenido de humedad ensilado con ácido sulfúrico o hidróxido de sodio. En la hidrólisis enzimática utilizaron la mezcla de enzimas celulolíticas SAE0020 (Sigma) y una preparación comercial de β-glucosidasa (Macklin). El fluido de incubación se preparó utilizando tampón de citrato de sodio 50 mM (pH 4.8), trabajaron con una carga de sólidos del 5% (p/v) y cargas de enzimas de 20 FPU/g y 15 CBU/g de MS en matraces Erlenmeyer de 125 mL en un agitador de baño de aire termostatizado a 50 °C y 180 rpm durante 72 h. El rastrojo de maíz con alto contenido de humedad se ensiló con ácido sulfúrico (0.3% y 0.6%) o hidróxido de sodio (0.5% y 1.0%) y se investigaron las características del ensilaje, perfil lignocelulósico y sacarificación enzimática los días 3, 7, 15 y 30 y 60 de ensilado. Los resultados mostraron que el 0.6% de H2SO4 redujo la pérdida de materia seca (9.81% a 6.34%) y contenido de nitrógeno amoniacal (3.89 g/kg a 1.04 g/kg MS) durante el ensilado, mientras que fue el caso contrario para el tratamiento con NaOH (19.89%, 5.74 g/kg MS). La hemicelulosa se redujo (27.98% a 22.61%, 16.81% MS) con 0,6% de H2SO4 o 1.0% de NaOH. El rendimiento de sacarificación disminuyó (de 306 mg/g a 229 mg/g MS) durante el ensilaje, el cual mejoró (de 229 mg/g a 356, 277 mg/g MS) con los tratamientos con H2SO4 y NaOH. Este estudio sugiere que el ensilaje con la adición de 0.6% de H2SO4 podría mejorar la conservación de nutrientes y el rendimiento de sacarificación del rastrojo de maíz con alto contenido de humedad.
Wang et al. (2020), en la investigación “Enhancing enzymatic hydrolysis of corn stover by twin-screw extrusion pretreatment”, mejoraron la hidrólisis enzimática del rastrojo de maíz mediante pretratamiento de extrusión de doble tornillo. Los tallos fueron cortados en pedazos pequeños y secado en un horno a 80 °C, y el rastrojo de maíz seco (CS) se trituró y tamizó. La hidrólisis enzimática de muestras de rastrojo de maíz por hemicelulasa y celulasa lo realizaron en un matraz Erlenmeyer de 250 mL que contenía 50 mL de tampón acetato de sodio (pH = 4.8) y 0.02% (p/v) de ázida de sodio (NaN₃) para inhibir el crecimiento microbiano. Después colocaron 1 g de muestra de rastrojo de maíz en el matraz y la carga de enzima fue de 15 FPU/g de materia seca. Luego, los matraces fueron incubados a 50 °C y 150 rpm durante 72 h y en un intervalo de tiempo de 12 h, extrajeron 5 mL de hidrolizado para determinar el rendimiento de glucosa o xilosa. En este estudio, el pretratamiento de extrusión de doble tornillo cambió la distribución del tamaño de las partículas y la estructura espacial de las muestras de rastrojo de maíz, pero básicamente no tuvo efectos en su estructura química. El rendimiento de glucosa de CS con agua destilada aumentó de 25 g/L de CS con 48 h de tiempo de hidrólisis a 34 g/L de CS con 36 h de tiempo de hidrólisis, y el rendimiento de glucosa de CS con agua destilada (CS-DW) y de CS con agua destilada, glicerol y NaHCO3 (CS-DW-GNa) aumentó a 45 g/L con 24 h de hidrólisis. Mientras tanto, el
rendimiento de xilosa de CS-DW y CS-DW-GNa aumentó de 19 g/L de CS (48 h) a 27 g/L de CS (36 h) y 40 g/L de CS (24 h), respectivamente. Estos resultados brindan una buena orientación para mejorar la conversión de lignocelulosas en monosacáridos para su posterior utilización.
Santamaria Vilca (2019) publicó el “Efecto de la concentración de ácido fosfórico y tiempo de hidrólisis en la obtención de azúcares reductores de grano de maíz (Zea mays) morado cocido”, donde su objetivo fue evaluar el efecto de la concentración de ácido fosfórico y tiempo de hidrólisis a 121 °C en la obtención de azúcares reductores (expresado como glucosa equivalente) en grano de maíz (Zea mays) morado cocido.
Las muestras de maíz morado cocido residual fueron trituradas en una licuadora con presencia de agua destilada en porción 1:1 p/p para generar una pasta. La pasta generada fue secada en una estufa a 40 °C. Luego se transformó en harina con un tamaño de partícula inferior a 1.0 mm (malla 16 en escala Tyler que corresponde a una abertura de 0.991 mm), en un mortero para posteriormente ser hidrolizado con ácido fosfórico en una relación harina/solución de 1/10. El diseño experimental consistió en un diseño compuesto central rotable, siendo la concentración de ácido fosfórico y tiempo de hidrolisis las variables independientes en los niveles de 1-5% p/p y 15-90 min respectivamente. Los resultados demuestran una mayor obtención de azúcares reductores cuando la concentración de ácido fosfórico es de 4.4% p/p con tiempo de 26 min (a 121 °C) donde se obtuvo 19.43 g azúcares reductores/100 g solución. Mientras que la menor obtención fue de 8.59 g azúcares reductores/100 g solución, que corresponde a una concentración de ácido de 1% y tiempo de hidrólisis de 52.5 min. El análisis demuestra un efecto estadísticamente significativo por parte de las variables independientes en la obtención de azúcares reductores. El modelo matemático obtenido al ser validado muestra errores inferiores al 10% indicando un buen ajuste. Los valores óptimos para la máxima obtención de azúcares reductores se encuentran entre 3.5- 4.5% p/p de ácido fosfórico y de 33-52 minutos de hidrólisis a 121 °C.
Sewsynker-Sukai & Gueguim Kana (2018), en el artículo “Simultaneous saccharification and bioethanol production from corn cobs: Process optimization and kinetic studies”, investigaron el proceso simultáneo de sacarificación y fermentación (SSF) para la producción de bioetanol a partir de mazorcas de maíz con prehidrólisis (PSSF) y sin prehidrólisis (OSSF). Desarrollaron dos modelos de superficie de respuesta con altos coeficientes de determinación (> 0.90). La optimización del proceso proporcionó altas concentraciones de bioetanol y conversiones de bioetanol para los modelos PSSF (36.92 ± 1.34 g/L y 62.36 ± 2.27%) y OSSF (35.04 ± 0.170 g/L y 58.13 ± 0.283%) respectivamente. Además, usaron los modelos logísticos y modificados de Gompertz para estudiar la cinética del crecimiento de células microbianas y la formación
de etanol en condiciones microaerófilas y anaeróbicas. El crecimiento celular en el proceso microaerofílico OSSF proporcionó la velocidad de crecimiento específico máxima (µmáx) de 0.274 h−1. El bioproceso microaerofílico PSSF proporcionó la concentración máxima de bioetanol potencial más alto (42.24 g/L). Este estudio demostró que las condiciones de cultivo microaerófilo en lugar de anaeróbico mejoraron el crecimiento celular y la producción de bioetanol, y que los pasos adicionales de prehidrólisis no tienen un impacto significativo en la concentración y conversión de bioetanol en el proceso de SSF.
Lancho Ruiz (2015) investigó sobre la “Obtención de jarabe a partir del almidón del maíz morado Zea maydis”, fue realizado en los laboratorios de la Facultad de Ingeniería pesquera y de Alimentos de la Universidad Nacional del Callao y en la Facultad de Ciencias Biológicas de la Universidad Nacional Mayor de San Marcos, donde extrajo el almidón para la evaluación de la hidrólisis enzimática, utilizando ácido clorhídrico al 1% y alfa amilasa al 1%, y a temperaturas de 65 y 90 °C, en suspensiones de almidón del maíz de muestra al 30%. El mejor resultado fue en la hidrólisis enzimática con una pregelificación del almidón antes de añadir la enzima alfa amilasa, logrando obtener una hidrólisis del almidón nativo hasta 24% en azucares solubles y formar un jarabe a partir del almidón del maíz morado Zea maydis L con una calidad superior.
Vavouraki et al. (2014), en el artículo “Optimization of thermo-chemical pretreatment and enzymatic hydrolysis of kitchen wastes”, manifiestan que optimizaron el efecto combinado de pre tratamiento termoquímico e hidrólisis enzimática de los desechos de cocina para maximizar la producción de azúcares solubles fermentables.
Evaluaron la concentración de HCl (0-3%), temperatura de tratamiento (30-100 °C) y tiempo de residencia (0-120 min) a volumen constante de 2.2 mL. Determinaron la influencia de la carga de sólidos en los rendimientos de azúcar después de la hidrólisis ácido-enzimático (2% de HCl, 85 °C, 80 min, 0.1% amilasa y 0.1% amiloglucosidasa) y alcalino-enzimática (1% KOH, 0 °C, 20 min, 1.1% amilasa, 0.4% amiloglucosidasa).
Concluyeron que el pre tratamiento termoquímico solo puede dar lugar a un mayor rendimiento azúcares solubles totales, 121% frente al 75.6% que se obtuvo en el óptimo combinado termo-químico a condiciones enzimáticas de este estudio.
Yan et al. (2011), en el artículo “Enzymatical hydrolysis of food waste and ethanol production from the hydrolysate”, evaluaron la factibilidad del uso de residuos de alimentos y residuos municipales principales como un sustrato potencial para generar hidrolizados para la producción de etanol. Según los resultados del análisis de superficie de respuesta, las condiciones óptimas para la producción de azúcar fueron: dosis de glucoamilasa de 142.2 u/g, pH de 4.82; temperatura de 55 ºC y tiempo de 2.48 h.
Concluyeron que la producción de azúcar (164.8 g/L) a las condiciones óptimas son
próximas al valor estimado por el modelo cuadrático (164.3 g/L), demostrando su validez. Por otra parte, evaluaron el efecto de la fermentación no esterilizada y esterilizada del hidrolizado (líquido) de los residuos de alimentos con la levadura Saccharomyces cerevisiae H058 en la producción de etanol, donde demostraron que a condiciones sin esterilizar es mejor por los nutrientes vírgenes en su interior.
1.2 Bases teóricas 1.2.1 Maíz morado
El maíz morado “es un fruto utilizado como alimento en el Perú prehispánico. Z.
mays habita en las vertientes occidentales del Perú alrededor de 8°00’28,8” S, 78°41’18,5” W y 1390 de elevación” (Leiva González et al., 2016, p. 296).
Respecto al cultivo en el Perú, aproximadamente 5000 ha corresponde al maíz morado, siendo las zonas agroecológicas de mayor área de cultivo Arequipa, Ica, Lima, Huánuco, Cajamarca y Ayacucho (MINAGRI, 2012).
En el Perú existen muchas variedades de maíz morado como: Morado Canteño, Morado Mejorado, Morado Caráz, Arequipeño, Cuzco Morado, Negro Junín y Negro Canaán.
Sin embargo, la variedad más comercial es el maíz morado Canteño porque se desarrolla bien entre los 1800 a 2500 msnm. En general la floración ocurre entre los 110-125 días, es tolerante a plagas y se adapta por ser nativa a las diferentes zonas.
Asimismo, altitudes entre 1000 y 2900 msnm contribuyen a una óptima producción de maíz morado; sin embargo, se puede producir también a 3000 msnm (INIA, 2007; Risco, 2007). (Pinedo Taco, 2015, p. 15)
El maíz morado se cultiva desde hace siglos en la Región Andina. En Perú, la gente consume una bebida típica hecha de maíz morado llamada “Chicha Morada”, que se cree que el folclore usa para mejorar la salud (Brack-Egg, 1999). El maíz morado es una importante fuente de antocianinas, pigmentos naturales ampliamente distribuidos en el reino vegetal con características peculiares en cuanto a la coloración de los alimentos.
Las antocianinas ya caracterizadas en mazorcas y semillas de maíz morado incluyen cianidina-3-glucósido, pelargonidina-3-glucósido, peonidina-3-glucósido y sus respectivas contrapartes malonadas (Aoki, Kuze, & Kato, 2001; Pascual-Teresa, Santos-Buelga, & Rivas-Gonzalo, 2002).
A las antocianinas se les atribuyen muchas actividades biológicas. Las actividades antioxidantes, antimutagénicas y anticancerígenas de las antocianinas han sido ampliamente reportadas (Bomser, Madhavi, Singletary, & Smith, 1996; Kähkönen &
Heinonen, 2003; Kamei et al., 1995; Katsube, Iwashita, Tsushida, Yamaki, & Kobori, 2003; Remeuf, Dorange y Dupuy, 1983; Yoshimoto, Okuno, Kumagai, Yoshinaga y Yamakawa, 1999; Yoshimoto, Okuno, Yamaguchi y Yamakawa, 2001). Además, las antocianinas se han utilizado en el tratamiento de diversos trastornos de la circulación
sanguínea resultantes de la fragilidad capilar (Wang, Cao y Prior, 1997), en estudios vasoprotectores y antiinflamatorios (Lietti, Cristoni y Picci, 1976), en la inhibición de la agregación plaquetaria (Morazzoni & Magistretti, 1986), y en el mantenimiento de la permeabilidad vascular normal (Wang et al., 1997).
También se han informado ampliamente los efectos beneficiosos para la salud de otros compuestos fenólicos no antocianínicos, incluidos los antioxidantes (Adom y Liu, 2002;
Cakir et al., 2003; Dewanto, Wu y Liu, 2002; Friedman, 1997; Ghiselli, Nardini, Baldi y Scaccini, 1998; Lien, Ren, Bui y Wang, 1999; Rice-Evans, Miller y Paganga, 1996;
Stintzing, Stintzing, Carle, Frei y Wrolstad, 2002; Wang y Lin, 2000); antimutagénico (Cardador-Martínez, CastañoTostado, & Loarca-Pin˜a, 2002; Edenharper, Keller, Platt,
& Unger, 2001; Edenharper, von Petersdoff, & Rauscher, 1993; Miyazawa & Hisama, 2003; Wang & Lee, 1996), actividad anticancerígena (Ferguson, 2001; Hertog, Van Poppel y Verhoeven, 1997; Iwashita, Kobori, Yamaki y Tsushida, 2000; Katsube et al., 2003; Loo, 2003; Surh, 1999; Yang, Landau, Huang, & Newmark, 2001) y otras propiedades biológicas. [...] la caracterización de los compuestos fenólicos no antocianínicos presentes en el maíz morado no ha sido reportada hasta donde sabemos.
La mayoría de las propiedades atribuidas a los extractos de maíz morado, incluyendo atributos colorantes (Duhard, Garner, & Megard, 1997; Cevallos-Casals & Cisneros- Zevallos, 2004), antioxidante (Cevallos-Casals & Cisneros-Zevallos, 2003a), antimicrobiana (Cevallos-Casals & Cisneros-Zevallos, 2003b), la actividad contra la obesidad y la mejora de la hiperglucemia (Tsuda, Horio, Uchida, Aoki, & Osawa, 2003) y las propiedades anticancerígenas (Hagiwara et al., 2001) se relacionaron con las antocianinas. (Pedreschi & Cisneros-Zevallos, 2007, pp. 956–957)
El maíz morado (Zea mays L.), originario de la región andina, América Latina, es una rica fuente de antocianinas. [...] el contenido de antocianinas en el maíz morado es mucho mayor, incluso hasta 82.3 mg/g materia seca (Cevallos-Casals y Cisneros- Zevallos, 2003; Wu et al., 2006). (Lao & Giusti, 2018, p. 87)
La mayoría de las variedades comerciales de maíz dulce (Zea mays L. var. rugosa) son de color amarillo o blanco, con un pericarpio incoloro y una capa de aleurona incolora (Lindstrom, 1923). El maíz no dulce pigmentado contiene antocianina generalmente en uno de estos tejidos, lo que les da a los granos un color púrpura, azul o púrpura rojizo (Salinas Moreno, Sánchez, Hernández y Lobato, 2005). Históricamente, el pericarpio pigmentado tiende a asociarse con algunas variedades de maíz peruano, mientras que la pigmentación con aleurona a menudo se asocia con variedades mexicanas de “maíz indio” (Grobman, Salhuana y Sevilla, 1961; Prasanna, 2012). En general, el maíz pigmentado con pericarpio tiende a tener una concentración de antocianina significativamente más alta que el maíz pigmentado con aleurona, debido a una mayor
cantidad de capas de células pigmentadas en el pericarpio, en comparación con una aleurona de una sola capa (Lao & Giusti, 2016; Li et al., 2019; Nankar et al., 2016) [...].
El maíz pigmentado es una fuente rica en antocianinas, con una concentración total de antocianinas que varía de 21 mg/100 g a 618 mg/100 g materia seca (Chatham, West, Berhow, Vermillion y Juvik, 2018; Nankar et al., 2016; Zhao et al. al., 2008). Las principales antocianinas en el maíz pigmentado son los glucósidos a base de cianidina, peonidina y pelargonidina (Chatham et al., 2018; Lao & Giusti, 2016; Nankar et al., 2016). Se ha demostrado que las antocianinas poseen varios beneficios potenciales para la salud, como antiinflamatorios, antioxidantes, antidiabéticos (Liu, Li, Zhang, Sun y Xia, 2014), anticancerígenos (Afrin et al., 2016; Hakimuddin, Paliyath y Meckling, 2004). ; Joshi et al., 2017), protección cardiovascular (Cassidy, 2018; Cassidy et al., 2016) y actividades antihipertensivas (Liu et al., 2014), así como actividad preventiva en el deterioro cognitivo relacionado con la edad y pérdida de memoria (Khoo, Azlan, Tang y Lim, 2017; Yousuf, Gul, Wani y Singh, 2016), lo que hace que el maíz dulce pigmentado no solo sea un producto visualmente diferente al maíz dulce amarillo o blanco, sino que potencialmente sea un producto con diferentes beneficios para la salud del maíz dulce amarillo. (Hong et al., 2020, p. 1)
En la Figura 1 se presenta la estructura del grano de maíz en estado maduro, el cual está formado por cuatro estructuras fundamentales o básicas: embrión (parte que de la estructura del maíz que da origen a una nueva planta), testa (parte superior del grano), pericarpio (cubierta) y endosperma (parte importante del grano en la que se encuentra el almidón y proteínas).
Figura 1
Estructura de un grano de maíz
Adaptado de (Frame et al., 2006, p. 75)
a) Taxonomía.
Takhtajan (1980) reportó la taxonomía del maíz morado la cual es:
Reino: Plantae
División: Magnoliophyta Clase: Liliopsida
Orden: Poales Familia: Poaceae
Tribu: Andropogoneae Género: Zea
Especie: Zea mays L.
Nombre común: Maíz morado b) Composición química.
En la Tabla 1 se presenta la composición química del maíz morado INIA-615 Negro Canaán diferenciando el grano y coronta.
Tabla 1
Composición química del maíz morado INIA-615 Negro Canaán
Componente Maíz grano Coronta
Humedad 1.67 2.33
Proteína 9.26 4.38
Extracto etéreo (%) 22.18 30.55
Energía (kcal/100 g) 496.70 694.53
Cenizas totales (%) 1.89 0.35
Carbohidratos 65.01 62.40
Contenido en 100 g
Laboratorio y nutrición de la UNSCH citado por Fernández (2009). Adaptado de (Pinedo Taco, 2015, p. 34)
En la Figura 2 se presenta la composición de tres variedades de maíz, en la que se puede observar que el contenido de almidón del maíz morado es menor al maíz amarillo y azul; pero respecto a proteína, fibra y aceite la variación es mínima entre estas variedades.
Figura 2
Características composicionales de tres variedades
Adaptado de (Somavat et al., 2018, p. 79) c) Usos.
Pinedo Taco (2015) destacó los usos del maíz morado, los cuales son:
• Colorante natural para la chicha y mazamorra morada a nivel casero.
• Materia prima para obtener colorante a nivel industrial, específicamente de la coronta, por su contenido de antocianinas. Este colorante se utiliza para la coloración de bebidas, productos lácteos, productos de panadería, productos vegetales, conservas de pescado, grasas, aceites, mermeladas, jaleas, frutas confitadas, frutas en almíbar, jarabes de frutas y otros; también se usa para el teñido y en la industria de cosméticos.
• Con el grano se puede extraer almidones y/o derivados o en la elaboración de alimentos balanceados para animales.
• Por sus propiedades anticarcinogénicas y antioxidantes puede ser utilizado en alimentos funcionales y nutracéuticos.
1.2.2 Hidrólisis
Es el proceso de “descomponer el complejo de celulosa y lignina-hemicelulosas- celulosa para hacerlo más susceptible a la hemicelulosa en azúcares monoméricos”
(Krishnan et al., 2020, p. 127) o donde los “carbohidratos poliméricos se convierten en monómeros de azúcar” (B. Sharma et al., 2020, p. 4).
“La hidrólisis es el paso crucial donde la celulosa se convierte en azúcares. Se dispone de diferentes métodos para la generación de azúcares a partir de biomasa lignocelulósica, de los cuales los métodos químicos y enzimáticos han demostrado ser más exitosos” (Binod et al., 2019, p. 447).
a) Tipos de hidrolisis.
Los tipos de hidrólisis donde se realiza la sacarificación son: química y enzimática.
Hidrólisis química.
En la hidrólisis química, los ácidos se utilizan para la hidrólisis de la celulosa para la generación de azúcares. La hidrólisis química generalmente se realiza mediante el uso de ácidos. Los ácidos minerales concentrados como H2SO4 y HCl se usan comúnmente. Los principales inconvenientes de este proceso son la corrosión del reactor, separación del producto, escasa reciclabilidad del catalizador y necesidad de un tratamiento adecuado de los efluentes generados durante el proceso (Huang & Fu, 2013). El uso de líquidos iónicos (IL) también se está volviendo popular hoy en día donde el proceso es menos peligroso.
Recientemente, varios estudios revelaron que los ácidos de base carbónica sulfonados, los ácidos sólidos magnéticos y los ácidos de base polimérica sirven como un buen agente para la hidrólisis de la celulosa (Huang & Fu, 2013).
Aunque la velocidad de hidrólisis ácida es mucho más rápida que la hidrólisis enzimática, el principal inconveniente es que la glucosa también se degrada rápidamente en condiciones ácidas (Cheung & Anderson, 1996). (Binod et al., 2019, p. 448)
“La hidrólisis catalizada por ácido ocurre principalmente a alta temperatura (100- 240 °C) y una alta probabilidad de formación de inhibidores debido a la alta temperatura y la condición ácida. De hecho, es una combinación de procesos de pretratamiento e hidrólisis” (B. Sharma et al., 2020, p. 4).
“Durante la hidrólisis ácida, los ácidos penetran en la biomasa y descomponen la lignina y, en última instancia, descomponen los polímeros de celulosa y hemicelulosa en moléculas de azúcar individuales. En este proceso se utilizan diferentes ácidos como H2SO4, HCl, HNO3, etc” (Binod et al., 2019, p. 448).
Hidrólisis enzimática.
El principal propósito comercial de la hidrólisis enzimática de los celulósicos es destruir la celulosa y otros polímeros de carbohidratos en azúcares fermentables, incluida la glucosa y/o los oligómeros que pueden convertirse en productos valiosos a través de enfoques biológicos o químicos. Aunque la hidrólisis enzimática de la celulosa se complica por la existencia de otros componentes (p. ej., hemicelulosa y lignina) y sus derivados después del pretratamiento, es esencial comprender los efectos de las características clave de la propia celulosa sobre la velocidad y la eficacia de la hidrólisis enzimática.
(Yang et al., 2011, p. 423)
La hidrólisis enzimática se lleva a cabo en condiciones suaves a pH 4.8 y una temperatura de 45 a 50 °C, que es óptima para la enzima celulasa. La principal ventaja de la hidrólisis enzimática sobre la hidrólisis química es que no crea un problema de corrosión (Duff, W.D. Murra, 1996). El tiempo necesario para la hidrólisis enzimática es mayor que el de la hidrólisis química. Además, el producto final de la hidrólisis enzimática inhibe la enzima y, en última instancia, afecta el proceso, a menos que se eliminen inmediatamente después de su formación. Aparte de esto, un cuello de botella importante en la producción de etanol lignocelulósico, en la actualidad, es el costo de las enzimas. (Binod et al., 2019, p. 449). Otra ventaja de la hidrólisis enzimática es la alta recuperación de azúcar sin formación de inhibidores.
b) Factores críticos de la hidrolisis.
En función al tipo de hidrólisis, química o enzimática, utilizado en el proceso de producción de azúcares monoméricos, los factores son diferentes, por ejemplo, los factores que influyen en la hidrólisis enzimática son “la temperatura, pH, carga de enzimas y tiempo que afecta la productividad de la sacarificación enzimática” (B.
Sharma et al., 2020, p. 4). Los tres primeros factores están relacionadas a las condiciones de operación de la enzima que se utiliza en la hidrólisis.
En la hidrólisis química, los factores críticos son la concentración del ácido, ácido que se utiliza, temperatura y pH. Cualquiera sea el tipo de hidrólisis, los valores de sus factores o variables varían en función a las características de la materia prima o biomasa.
1.2.3 Hidrólisis enzimática
Actualmente, la conversión de materiales celulósicos en azúcares simples mediante hidrólisis enzimática es una de las tecnologías más viables hacia el concepto de biorrefinería. La hidrólisis enzimática de la celulosa es una reacción heterogénea que normalmente comienza con la rápida adsorción de celulasas en regiones fácilmente accesibles de la superficie del sustrato, lo que caracteriza una velocidad de hidrólisis inicial relativamente rápida. Sin embargo, se logra una fase intermedia con una velocidad de reacción moderada cuando se ha hidrolizado alrededor del 50-70% del sustrato original. A partir de entonces, se observa una desaceleración dramática de la velocidad cuando ya se han alcanzado los grados más altos de conversión. Estas tres fases de hidrólisis dependen de la composición del material lignocelulósico y del método de pretratamiento empleado (Mooney et al., 1998; Saddler y Arantes, 2010). En algunos casos, se requieren tiempos de hidrólisis prolongados y/o altas cargas de enzimas para lograr una conversión casi completa de la celulosa, aunque es importante señalar que la carga de enzimas y el rendimiento de la hidrólisis no muestran una relación lineal
(Ramos et al., 1992). En general, las altas cargas de enzimas dan como resultado altos rendimientos de glucosa, pero existe un límite porque las celulasas compiten por los sitios de sustrato más accesibles, y parte se pierde debido a su unión no específica y/o improductiva a la lignina y a los complejos de lignina-carbohidrato (Xiao et al., 2004).
(Ramos et al., 2020, pp. 77–78)
En la Figura 3 se muestra esquemáticamente el proceso de la hidrólisis enzimática, en la etapa inicial el sustrato y enzima no están en contacto en la mezcla para luego mediante la difusión, las moléculas de las enzimas se aproximan a la biomasa y por adsorción se unen a la celulosa en zonas accesibles, en la cual se produce la hidrólisis ennzimática y se da la formación de azucares.
Figura 3
Diagrama esquemático del proceso de hidrólisis enzimática de la biomasa lignocelulósica: 1) difusión enzimática, 2) adsorción y 3) hidrólisis
Adaptado de (H. Zhang et al., 2021, p. 4)
A diferencia de la hidrólisis química, la hidrólisis enzimática se lleva a cabo en condiciones suaves a un pH de 4.8 y una temperatura de 45 a 50 °C, que es óptima para la enzima celulasa. La principal ventaja de la hidrólisis enzimática sobre la hidrólisis química es que no crea un problema de corrosión (Duff & Murray, 1996). El tiempo necesario para la hidrólisis enzimática es mayor que el de la hidrólisis química. Además, el producto final de la hidrólisis enzimática inhibe la enzima y, en última instancia, afecta el proceso, a menos que se eliminen inmediatamente después de su formación. Aparte de esto, un cuello de botella importante en la producción de etanol lignocelulósico, en la actualidad, es el costo de las enzimas.
Teniendo en cuenta la complejidad y heterogeneidad de la biomasa lignocelulósica, una sola enzima no hidrolizará toda la biomasa y se requiere el desarrollo de un cóctel de enzimas como celulasas, hemicelulasas y otras enzimas accesorias para la hidrólisis completa. (Binod et al., 2019, p. 449)
a) Enzimas.
Las enzimas son sustancias que catalizan las reacciones bioquímicas en las células de todos los organismos. Estas reacciones constituyen la base química de la vida. La mayoría de las enzimas son proteínas (unas pocas son ácidos ribonucleicos o ribonucleoproteínas) y la maquinaria catalítica está ubicada en un sitio activo relativamente pequeño, donde los sustratos se unen y se procesan químicamente en productos. (Frey & Hegeman, 2007, p. 1)
El costo de la enzima es el mayor obstáculo en las refinerías de bioetanol debido a su enorme consumo en el mercado. sacarificación (Wang, 2013). Hoy en día, las enzimas microbianas se utilizan ampliamente en diversas industrias para hacer que el proceso sea económicamente viable. Hay una serie de microorganismos que secretan naturalmente enzimas celulolíticas. Clostridium, Thermomonospora, Trichoderma, Aspergillus son las cepas bacterianas y fúngicas comunes para la producción de celulasas (Kuhad et al., 2011). Entre todas, las especies de Trichoderma son las candidatas más deseables y potenciales para la producción de celulasa (Imran et al., 2016). (B. Sharma et al., 2020, p. 4)
Las enzimas son macromoléculas de proteínas grandes que realizan tareas catalíticas en biología (Kluger, 1998; Leadlay, 1978; Lipscomb, 1981; Yonaha &
Soda, 1994; Moss, 1968). El sitio activo de una enzima típica contiene aminoácidos, conocidos como grupos catalíticos, que participan simultáneamente en la localización, unión y catalización de transformaciones de moléculas de reactivo (sustrato) (Fersht, 1984; Palmer, 1995).
Las enzimas tienen un modo de acción molecular, aunque son más bien moléculas grandes. Operan en solución abierta. Sin embargo, sus capacidades catalíticas generalmente superan con creces las de los sistemas comparables fabricados por el hombre. De hecho, las enzimas como clase se consideran el tipo de catalizador más eficaz, por un margen considerable. (Swiegers, p. 41) b) Clasificación de las enzimas.
Las enzimas se clasifican numéricamente y a cada enzima luego de las letras
“EC” (Enzyme Commission numbers, en español Números de la Comisión de Enzimas) se le designa cuatro números separados por puntos; primer, segundo, tercer y cuarto número indican la clase, subclase, subsubclase y orden en la lista, respectivamente.
Ejemplo de esta clasificación tenemos a la pululanasa y la isoamilasa cuyos códigos son EC 3.2.1.41 y EC 3.2.1.68, respectivamente.
Cardellá Rosales et al. (2007) indican que las enzimas presentan dos propiedades fundamentales: eficiencia catalítica y especificidad de acción. La última sirve como criterio para la clasificación y nomenclatura de las enzimas, así se establecen seis grupos fundamentales o clases principales teniendo en cuenta la reacción global, que a su vez se dividen en subclases y subsubclases considerando otras características del tipo de reacción como son los grupos involucrados, cofactores y otros. Los grupos principales son:
1. Oxidoreductasas: Catalizan reacciones de oxidorreducción, es decir, la transferencia de electrones o sus equivalentes entre un donante y un aceptor.
2. Transferasas: Catalizan la transferencia de un grupo químico entre un donante y un aceptor. Se excluyen aquellas que transfieren electrones o sus equivalentes pues pertenecen al primer grupo, y aquellas en que el aceptor del grupo es el agua pues son parte del siguiente grupo.
3. Hidrolasas: Catalizan la ruptura de enlaces químicos con la participación de las moléculas del agua.
4. Liasas: Catalizan reacciones en las cuales se produce la adición o sustracción de grupos químicos a dobles enlaces.
5. Isomerasas: Catalizan la interconversión de dos isómeros.
6. Ligasas: Catalizan la unión covalente de dos sustratos utilizando para ello la energía de hidrólisis de nucleósidos trifosfatados, generalmente el ATP.
c) Estructura y mecanismos de las enzimas.
Muy pocos mecanismos de reacción enzimática involucran un solo evento químico y un solo estado de transición. La mayoría procede en múltiples pasos que involucran la formación transitoria de intermediarios reactivos. En una reacción de varios pasos, una enzima debe catalizar más de un proceso químico.
Incluso una reacción tan simple como la hidrólisis de péptidos por quimotripsina procede en dos pasos, acilación y desacilación, y los estados de transición de los dos procesos no pueden ser idénticos, aunque pueden ser muy similares.
Dos reacciones completamente diferentes son catalizadas en el mismo sitio […].
Los estados de transición deben ser diferentes, y la enzima seguramente sufre cambios estructurales en su adaptación a la catálisis de las dos reacciones. Los dos estados de transición en una reacción que involucra transferencia de acilo en ambos pasos a menudo son similares, pero por muy relacionados que estén los pasos, siempre hay diferencias. El reconocimiento de estas diferencias por
parte del sitio activo enzimático debe implicar al menos cambios menores en su estructura. (Frey & Hegeman, 2007, p. 6)
d) Cofactores y coenzimas.
La mayoría de las reacciones enzimáticas proceden con cambios químicos que no pueden provocar las cadenas laterales de los residuos de aminoácidos. Estas enzimas funcionan en cooperación con coenzimas y cofactores, que les otorgan potencialidades fisicoquímicas que no se encuentran en los aminoácidos.
Muchas coenzimas son moléculas orgánicas que incorporan grupos funcionales con propiedades químicas que les permiten facilitar reacciones de ciertos tipos.
Estas moléculas se unen a sitios activos diseñados para ellas a través de la evolución y equipados para ayudar en sus funciones coenzimáticas. Muchas de estas coenzimas se derivaron de las vitaminas y, en las primeras investigaciones bioquímicas, las vitaminas y coenzimas a menudo se consideraban estrechamente vinculadas e incluso sinónimas. Sin embargo, las vitaminas como la vitamina D son más parecidas a las hormonas que a las coenzimas y, en la bioquímica moderna, las coenzimas recién descubiertas no están relacionadas con las vitaminas y tienen identidades independientes de cualquier origen nutricional […].
Cada coenzima y enzima afín forman una unión que les permite actuar como una sola entidad catalítica que funciona en concierto para provocar una transformación química difícil. Cada coenzima proporciona la química requerida para una clase de procesos enzimáticos, y los mecanismos de catálisis enzimática a menudo se revelan a través de las acciones de las coenzimas participantes. (Frey & Hegeman, 2007, p. 129)
Las enzimas, como otras proteínas, tienen pesos moleculares que oscilan entre 12000 y más de un millón. Algunas enzimas no necesitan grupos químicos distintos de sus propios residuos de aminoácidos. Otros requieren un componente químico adicional llamado cofactor, que puede ser uno o más iones inorgánicos como Fe2+, Mg2+, Mn2+ o Zn2+ o una molécula orgánica compleja o metalorgánica llamada coenzima. Las coenzimas actúan como transportadores transitorios de grupos funcionales específicos. La mayoría de ellos se derivan de vitaminas, nutrientes orgánicos cuya presencia en la dieta es necesaria en pequeñas cantidades […]. Algunas enzimas requieren tanto una coenzima como uno o más iones metálicos para estar activas. Una coenzima o un ion metálico que se une muy fuerte, o incluso covalentemente, a una enzima se denomina grupo prostético. Una enzima completa, catalíticamente activa junto con su coenzima y/o iones metálicos, se denomina holoenzima. La parte proteica de
una de estas enzimas se denomina apoenzima o apoproteína. Finalmente, algunas enzimas se modifican covalentemente por fosforilación, glicosilación y otros procesos. Muchas de estas modificaciones están implicadas en la regulación de la actividad enzimática. (Nelson & Cox, 2014, p. 190)
e) Función biológica de las enzimas.
La catálisis enzimática de las reacciones es esencial para los sistemas vivos.
Bajo las condiciones biológicas relevantes, las reacciones no catalizadas tienden a ser lentas; la mayoría de las moléculas biológicas son muy estables en las condiciones internas de las células con pH neutro, temperaturas suaves y un ambiente acuoso. Además, muchos procesos químicos comunes, como la formación transitoria de intermediarios cargados inestables o la colisión de dos o más moléculas exactamente en la orientación exacta necesaria para que ocurran las reacciones, son desfavorables o improbables en el entorno celular.
Las reacciones necesarias para digerir los alimentos, enviar señales nerviosas o contraer los músculos simplemente no ocurren a un ritmo adecuado sin catálisis. Las enzimas eluden estos problemas al proporcionar un entorno específico adecuado para que una reacción dada ocurra más rápidamente. La propiedad característica de las reacciones catalizadas por enzimas es que la reacción ocurre confinada a un bolsillo de la enzima llamado sitio activo. La molécula que se une al sitio activo y sobre la que actúa la enzima se denomina sustrato. El contorno de la superficie del sitio activo está delimitado por residuos de aminoácidos con grupos en las cadenas laterales que se unen al sustrato y catalizan su transformación química. A menudo, el sitio activo envuelve el sustrato, separándolo completamente de la solución. El complejo enzima- sustrato, cuya existencia fue propuesta por primera vez por Charles-Adolphe Wurtz en 1880, es fundamental para la acción enzimática. (Nelson & Cox, 2014, p. 192)
f) Factores que influyen en la hidrolisis enzimática.
Para que el proceso sea económicamente viable, se han estudiado y optimizado tentativamente varios factores relacionados con el sustrato, la enzima y el reactor que limitan la eficiencia de la hidrólisis de la celulosa (Gatt et al., 2018; Yang et al., 2011a; Saddler y Arantes, 2010; Chandra et al., 2007; Mansfield et al., 1999).
Estos factores pueden explicar la desaceleración de la velocidad de reacción y la hidrólisis incompleta de los materiales lignocelulósicos pretratados, particularmente en sólidos totales de sustrato altos. (Ramos et al., 2020, pp. 77–
78)
La hidrólisis enzimática está influenciada tanto por las características estructurales de la celulosa como por el modo de acción de la enzima. Debido a la complejidad del sustrato de celulosa y el sistema de celulasa, el mecanismo de hidrólisis del sustrato de celulosa aún no se comprende completamente, aunque el conocimiento detallado de algunos aspectos de la estructura de la enzima, las propiedades moleculares de la enzima y la ultraestructura de la celulosa se han obtenido a través de extensos estudios durante las últimas décadas. (Yang et al., 2011, p. 422)
A continuación, se presenta los factores que influyen en la hidrólisis enzimática:
Factores relacionados con el sustrato.
La velocidad de hidrólisis enzimática de la lignocelulosa se ve profundamente afectada por las características estructurales de la celulosa (Penner & Liaw, 1994), que incluyen la cristalinidad de la celulosa, área de superficie disponible/accesible, organización estructural, es decir, macroestructura (fibra) y microestructura (elementos-microfibrilla), tamaño de partícula y presencia de materiales asociados como hemicelulosa y lignina. Se ha demostrado que el efecto de la cristalinidad del sustrato juega un papel importante en la limitación de la hidrólisis en algunos estudios.
La concentración de sustrato es uno de los principales factores que afectan el rendimiento y la velocidad inicial de hidrólisis enzimática de la celulosa. Mantener altas concentraciones de sólidos durante todo el proceso de conversión de biomasa a etanol es importante desde el punto de vista de la viabilidad energética y económica. La hidrólisis enzimática con alto contenido de sólidos tiene lugar a niveles de sólidos donde inicialmente no hay presente una cantidad significativa de agua libre. Esto permite una mayor capacidad del sistema, menor demanda de energía para calentar y enfriar la pulpa y también menos descarga de efluentes. En cuanto a la viabilidad económica general de la producción de etanol lignocelulósico, una alta concentración de sustrato permite la producción de una solución concentrada de azúcar, que a su vez es beneficiosa para la fermentación posterior. (Binod et al., 2019, p. 455)
Las características del sustrato tienen un papel clave en el control de las velocidades y los rendimientos de la hidrólisis de la celulosa, y estos incluyen el área superficial, el tamaño de las partículas, el volumen de los poros, la cristalinidad, el grado de polimerización y el contenido y la composición de la lignina, así como la presencia de compuestos fenólicos y hemicelulosa.
productos derivados como xilosa y xilo-oligómeros. (Kim et al., 2011; Ximenes et
al., 2010; Yun et al., 2002; Zhang y Lynd, 2004). El área superficial de celulosa accesible a las enzimas celulasas es uno de los factores más importantes para la hidrólisis eficiente de los polisacáridos vegetales [...]. Independientemente de las diferencias sustanciales en la fuente, la estructura y la composición química de las materias primas, y las diferencias en la tecnología de pretratamiento aplicada, la carga de proteínas requerida para lograr una hidrólisis eficiente dependía en gran medida de la accesibilidad de la celulosa en cada uno de los materiales pretratados. Los sustratos ricos en celulosa con celulosa altamente accesible requerían una carga de proteína más baja por gramo de glucano para lograr una hidrólisis eficiente en comparación con los sustratos que contenían celulosa menos accesible. Por lo tanto, los resultados sugirieron que el paso limitante de la velocidad durante la hidrólisis no es la escisión catalítica de las cadenas de celulosa, sino la accesibilidad limitada de las enzimas a las cadenas de celulosa debido a la estructura física del sustrato celulósico.
El grado de cristalinidad de la celulosa también juega un papel importante en la susceptibilidad de los materiales lignocelulósicos a la hidrólisis enzimática. Una disminución en la cristalinidad de la celulosa influye en la velocidad inicial de hidrólisis (Laureano-Perez et al., 2005) [...].
Altos grados de deslignificación no garantizan la obtención de rendimientos teóricos de glucosa por hidrólisis enzimática [...]. Por el contrario, la presencia de lignina residual crea una estructura de pared celular interna porosa que expone la superficie de las microfibrillas y facilita el acceso a las enzimas.
La lignina y sus derivados producidos por el pretratamiento, dependiendo de factores como la concentración, distribución y composición, pueden interferir con el acceso de las celulasas al componente celulósico (Mooney et al., 1998; Sun y Cheng, 2002). Además, algunas de las enzimas responsables de la hidrólisis de la celulosa pueden inactivarse por adsorción improductiva en la superficie de los fragmentos de lignina. La fracción de celulasa que se vuelve improductiva debido a este tipo de adsorción puede alcanzar hasta el 70% del total de enzimas añadidas al medio de reacción (Wang et al., 2003). Además, los fragmentos de lignina modificados químicamente que se generan durante el pretratamiento pueden actuar como inhibidores no competitivos de celulasas y hemicelulasas.
Por otro lado, las β-glucosidasas están fuertemente influenciadas por la presencia de pequeñas cantidades de compuestos fenólicos. La β-glucosidasa de T. reesei es cuatro veces más sensible a los derivados fenólicos que la β- glucosidasa de A. niger. Con esto, la suplementación de celulasas y
hemicelulasas comerciales con β-glucosidasa de fuentes como A. niger es una práctica común en la literatura (Dos Santos et al., 2019).
Dependiendo de las condiciones del pretratamiento, particularmente cuando se lleva a cabo en condiciones ácidas, los fragmentos de lignina solubilizados en el medio de reacción tienden a reaccionar entre sí y con derivados de carbohidratos como los compuestos de furano para producir materiales de alta masa molecular que también son muy perjudiciales para la hidrólisis enzimática (Balat, 2011).
Estos compuestos se denominan pseudolignina porque su estructura no se parece a la de la lignina nativa (Li et al., 2007) [...].
Los materiales lignocelulósicos con bajo contenido de lignina generalmente exhiben una alta accesibilidad a la celulosa, lo que requiere cargas de enzimas más bajas para lograr altos rendimientos de glucosa. Por lo tanto, las tecnologías de pretratamiento que son efectivas en la deslignificación de biomasa eliminan las barreras hidrofóbicas a la hidrólisis enzimática [...].
Un enfoque prometedor para aumentar la hidrólisis enzimática de la celulosa es la suplementación con surfactantes (Tween, Triton), polímeros (polietilenglicol) o proteínas no catalíticas (albúmina de suero bovino) (Börjesson et al., 2007;
Eriksson et al., 2002; Kristensen et al., 2007). El principal mecanismo de estos aditivos se debe a la prevención de la unión inespecífica de las enzimas a la lignina. Se cree que los tensioactivos y los polímeros forman una capa hidratada en la superficie de la lignina, formando un obstáculo estérico para la unión improductiva de la celulasa (Eriksson et al., 2002). (Ramos et al., 2020, pp. 77–
80)
Factores relacionados con enzimas.
Se ha sugerido que varios factores asociados con la naturaleza del sistema enzimático de celulasa influyen durante el proceso de hidrólisis. Estos incluyen la concentración de enzimas, la adsorción de enzimas, el sinergismo, la inhibición del producto final, la desactivación mecánica (tensión de cizallamiento de fluidos o interfaz gas-líquido), la inactivación térmica y la unión irreversible (no productiva) a la lignina. En el proceso de hidrólisis enzimática, la naturaleza del sistema enzimático utilizado, el modo de acción (endoenzimas frente a exoenzimas) y su mecanismo estereoquímico de hidrólisis (inversión frente a retención) están interrelacionados. Además, la sinergia entre las enzimas puede resultar significativamente beneficiosa para aumentar las velocidades de hidrólisis del sustrato complejo. El sinergismo también depende del sustrato, y algunas mezclas muestran una acción cooperativa sobre sustratos amorfos, pero no sobre celulosa microcristalina. Todos estos factores pueden influir